Suicide Gene Therapy Mediated with Exosomes Produced by Mesenchymal Stem/Stromal Cells Stably Transduced with HSV Thymidine Kinase


{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Results
2.1. A Schematic Workflow of All Steps Performed in This Study
2.2. MSCs Obtained from Various Human Tissues Can Be Stably Transduced with the HSVTK Suicide Gene
2.3. The Homogenous Population of Mesenchymal Stem Cells (MSCs) Transduced with Herpes Simplex Virus Thymidine Kinase (HSVTK) Gene Release Exosomes
2.4. HSVTK-MSCs and Corresponding Conditional Medium (CM) Induce Tumor Cell Death in the Prodrug Ganciclovir Presence in a Dose-Dependent Manner
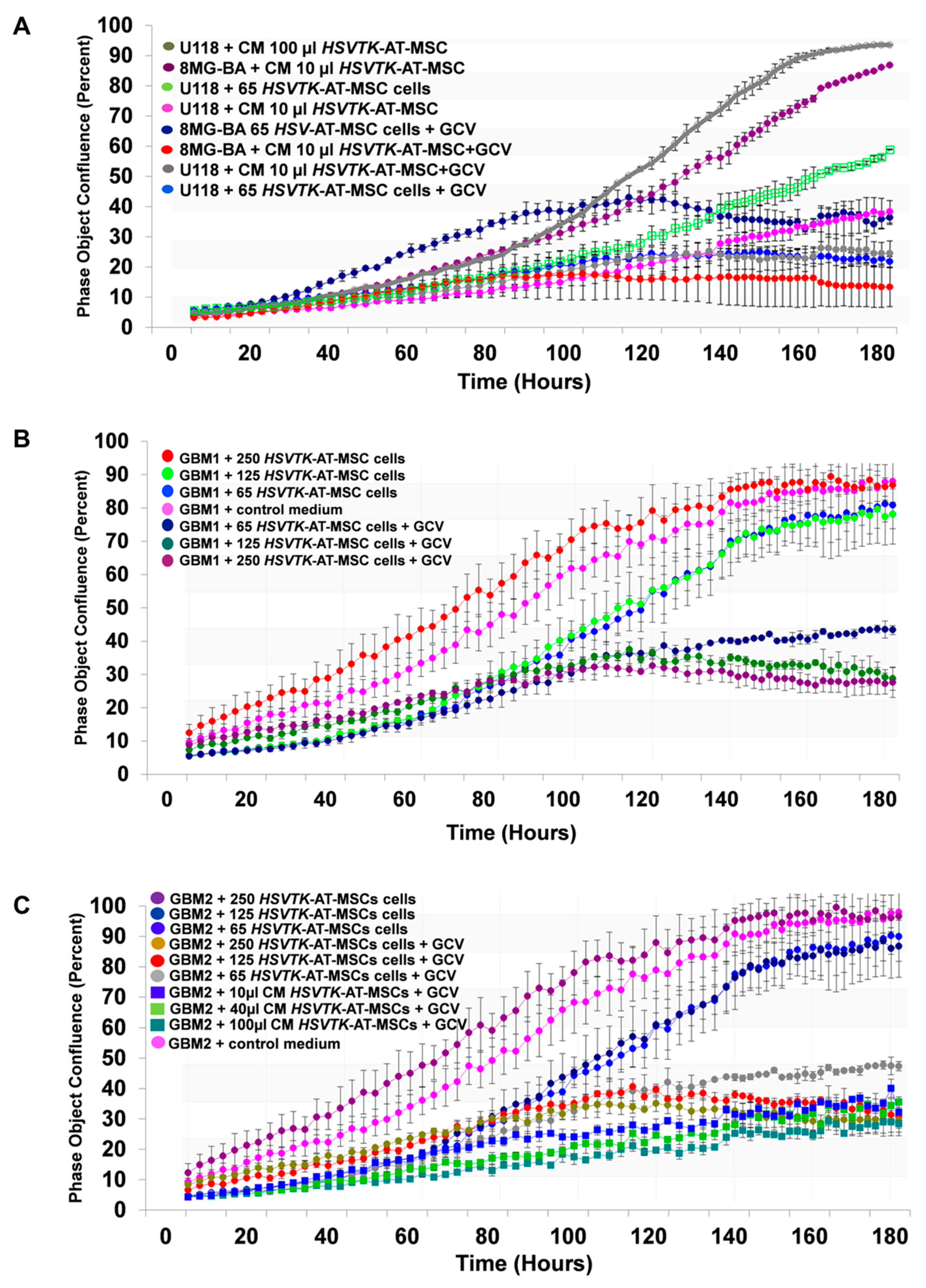
2.5. The Efficiency of HSVTK-AT-MSCs Compared to Matched CM in the Growth Inhibiting Activity of Glioma Cell Lines and Primary Glioblastoma Cells
2.6. Exosomes Are Responsible for the Tumor Cell Killing Activity in HSVTK-MSC CM
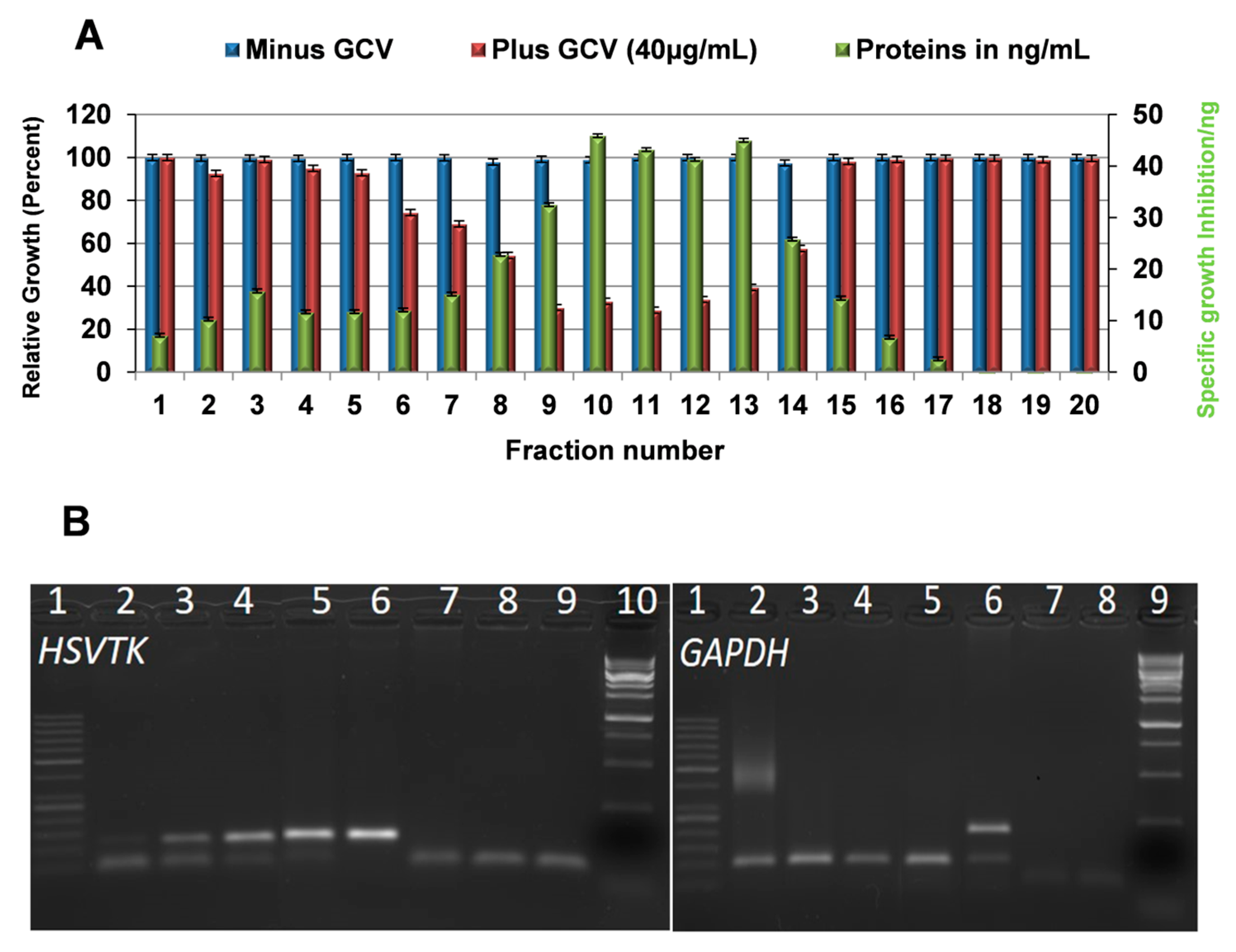
2.7. Messenger Ribonucleic Acid (mRNA) of the Prodrug Converting Gene Is Present in the Cargo of Exosomes Released from HSVTK-MSCs
2.8. Tumor Cell Growth Inhibition Efficacy of Exosomes from HSVTK Gene Transduced MSCs of Various Tissues Origin
3. Discussion
4. Materials and Methods
4.1. Culture Isolation and Maintenance of MSCs
4.2. Preparation of HSVTK-Retrovirus-Producing Cells
4.3. Cell Transduction with Retrovirus
4.4. Cell Growth Assessment Using IncuCyte Live Cell Monitoring System
4.5. Exosomes Isolation
4.6. Polymerase Chain Reaction (PCR), RT-PCR, and qRT-PCR Analysis
- 5′ GGAGGACAACACATCGACCG 3′ (forward) and
- 5′ GCAGATACCGCACCGTATTGGC 3′ (reverse) for HSVTK (122 bp),
- 5′ CCACTCCTCCACCTTTGAC 3′ (forward) and
- 5′ ACCCTGTTGCTGTAGCCA 3′ (reverse) for GAPDH DNA (206 bp), cDNA (102 bp),
- 5′ TAGTCAAACCGGCACGATGTC 3′ (forward) and
- 5′ ATCAAGCAGTCGGAGCAGTTC 3′ (reverse) for DSP (80 bp).
4.7. Isolation and Detection of mRNA from HSVTK-MSC Exosomes
4.8. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Studeny, M.; Marini, F.C.; Champlin, R.E.; Zompetta, C.; Fidler, I.J.; Andreeff, M. Bone marrow derived mesenchymal stem cells as vehicles for interferon-beta delivery into tumors. Cancer Res. 2002, 62, 3603–3608. [Google Scholar] [PubMed]
- Altaner, C. Prodrug Cancer Gene Therapy, Review. Cancer Lett. 2008, 270, 191–201. [Google Scholar] [CrossRef] [PubMed]
- Kucerova, L.; Altanerova, V.; Matuskova, M.; Tyciakova, S.; Altaner, C. Adipose tissue-derived human mesenchymal stem cells mediated prodrug cancer gene therapy. Cancer Res. 2007, 67, 6304–6313. [Google Scholar] [CrossRef]
- Kucerova, L.; Matuskova, M.; Pastorakova, A.; Tyciakova, S.; Jakubikova, J.; Bohovic, R.; Altanerova, V.; Altaner, C. Cytosine deaminase expressing human mesenchymal stem cells mediated tumour regression in melanoma bearing mice. J. Gene Med. 2008, 10, 1071–1082. [Google Scholar] [CrossRef]
- Cavarretta, I.T.; Altanerova, V.; Matuskova, M.; Kucerova, L.; Culig, Z.; Altaner, C. Adipose tissue-derived mesenchymal stem cells expressing prodrug-converting enzyme inhibit human prostate tumor growth. Mol. Ther. 2010, 18, 223–231. [Google Scholar] [CrossRef]
- Abrate, A.; Buono, R.; Canu, T.; Esposito, A.; Del Maschio, A.; Lucianò, R.; Bettiga, A.; Colciago, G.; Guazzoni, G.; Benigni, F.; et al. Mesenchymal stem cells expressing therapeutic genes induce autochthonous prostate tumour regression. Eur. J. Cancer 2014, 50, 2478–2488. [Google Scholar] [CrossRef]
- Altanerova, V.; Cihova, M.; Babic, M.; Rychly, B.; Ondicova, K.; Mravec, B.; Altaner, C. Human adipose tissue-derived mesenchymal stem cells expressing yeast cytosinedeaminase::uracil phosphoribosyltransferase inhibit intracerebral rat glioblastoma. Int. J. Cancer 2012, 130, 2455–2463. [Google Scholar] [CrossRef]
- Altaner, C.; Altanerova, V.; Cihova, M.; Ondicova, K.; Rychly, B.; Baciak, L.; Mravec, B. Complete regression of glioblastoma by mesenchymal stem cells mediated prodrug gene therapy simulating clinical therapeutic scenario. Int. J. Cancer 2014, 134, 1458–1465. [Google Scholar] [CrossRef]
- Altaner, C. Stem Cell-Mediated Prodrug Gene Therapy of High-Grade Brain Tumors, In Stem Cell Therapeutics for Cancer; Shah, K., Ed.; Wiley-Blackwell: Hoboken, NJ, USA, 2013; pp. 57–72. [Google Scholar]
- Moolten, F.L. Tumor chemosensitivity conferred by inserted herpes thymidine kinase genes: Paradigm for a prospective cancer control strategy. Cancer Res. 1986, 46, 5276–5281. [Google Scholar]
- Matuskova, M.; Hlubinova, K.; Pastorakova, A.; Hunakova, L.; Altanerova, V.; Altaner, C.; Kucerova, L. HSV-tk expressing mesenchymal stem cells exert bystander effect on human glioblastoma cells. Cancer Lett. 2010, 290, 58–67. [Google Scholar] [CrossRef]
- Altaner, C.; Altanerova, U. Mesenchymal Stem Cell Exosome-Mediated Prodrug Gene Therapy for Cancer. Suicide Gene Therapy: Methods and Protocols, Methods in Molecular Biology; Duzgunes, N., Ed.; Springer Science+Business Media, LLC: Berlin/Heidelberg, Germany, 2019; pp. 75–85. [Google Scholar]
- Kanada, M.; Kim, B.D.; Hardy, J.W.; Ronald, J.A.; Bachmann, M.H.; Bernard, M.P.; Perez, G.I.; Zarea, A.A.; Ge, T.J.; Withrow, A.; et al. Microvesicle-mediated delivery of minicircle DNA results in effective gene-directed enzyme prodrug cancer therapy. Mol. Cancer Ther. 2019, 18, 2331–2342. [Google Scholar] [CrossRef] [PubMed]
- Altanerova, U.; Jakubechova, J.; Benejova, K.; Priscakova, P.; Pesta, M.; Pitule, P.; Topolcan, O.; Kausitz, J.; Zduriencikova, M.; Repiska, V.; et al. Prodrug suicide gene therapy for cancer targeted intracellular by mesenchymal stem cell exosomes. Int. J. Cancer 2019, 144, 897–908. [Google Scholar] [CrossRef] [PubMed]
- Zhang, H.; Freitas, D.; Kim, H.S.; Fabijanic, K.; Li, Z.; Chen, H.; Mark, M.T.; Molina, H.; Martin, A.B.; Bojmar, L.; et al. Identification of distinct nanoparticles and subsets of extracellular vesicles by asymmetric flow field-flow fractionation. Nat. Cell Biol. 2018, 20, 332–343. [Google Scholar] [CrossRef]
- Freeman, S.M.; Abboud, C.N.; Whartenby, K.A.; Packman, C.H.; Koeplin, D.S.; Moolten, F.L. Abraham, G.N. The “bystander effect”: Tumor regression when a fraction of the tumor mass is genetically modified. Cancer Res. 1993, 53, 5274–5283. [Google Scholar] [PubMed]
- Kruse, C.A.; Roper, M.D.; Kleinschmidt-DeMasters, B.K.; Banuelos, S.J.; Smiley, W.R.; Robbins, J.M.; Burrows, F.J. Purified herpes simplex thymidine kinase retrovector particles. I. In vitro characterization, in situ transduction efficiency, and histopathological analyses of gene therapy-treated brain tumors. Cancer Gene Ther. 1997, 4, 18–28. [Google Scholar]
- Rainov, N.G. A phase III clinical evaluation of herpes simplex virus type 1 thymidine kinase and ganciclovir gene therapy as an adjuvant to surgical resection and radiation in adults with previously untreated glioblastoma multiforme. Hum. Gene Ther. 2000, 11, 2389–2401. [Google Scholar] [CrossRef] [PubMed]
- Elshami, A.A.; Saavedra, A.; Zhang, H.; Kucharczuk, J.C.; Spray, D.C.; Fishman, G.I.; Amin, K.M.; Kaiser, L.R.; Albelda, S.M. Gap junctions play a role in the ’bystander effect’ of the herpes simplex virus thymidine kinase/ganciclovir system in vitro. Gene Ther. 1996, 3, 85–92. [Google Scholar]
- Miletic, H.; Fischer, Y.; Litwak, S.; Giroglou, T.; Waerzeggers, Y.; Winkeler, A.; Li, H.; Himmelreich, U.; Lange, C.; Stenzel, W.; et al. Bystander killing of malignant glioma by bone marrow-derived tumor-infiltrating progenitor cells expressing a suicide gene. Mol. Ther. 2007, 15, 1373–1381. [Google Scholar] [CrossRef]
- Cottin, S.; Gould, P.V.; Cantin, L.; Caruso, M. Gap junctions in human glioblastomas: Implications for suicide gene therapy. Cancer Gene Ther. 2011, 18, 674–681. [Google Scholar] [CrossRef]
- Singh, S.K.; Hawkins, C.; Clarke, I.D.; Squire, J.; Bayani, J.; Hide, T.; Henkelman, R.M.; Cusimano, M.D.; Dirks, P.B. Identification of human brain tumour initiating cells. Nature 2004, 432, 396–401. [Google Scholar] [CrossRef]
- Hu, W.; Liu, W. Side populations of glioblastoma cells are less sensitive to HSV-TK/GCV suicide gene therapy system than the non-side population. Vitro Cell Dev. Biol. Anim. 2010, 46, 497–501. [Google Scholar] [CrossRef] [PubMed]
- Huszthy, P.C.; Giroglou, T.; Tsinkalovsky, O.; Euskirchen, P.; Skaftnesmo, K.O.; Bjerkvig, R.; von Laer, D.; Miletic, H. Remission of invasive, cancer stem-like glioblastoma xenografts using lentiviral vector-mediated suicide gene therapy. PLoS ONE 2009, 4, e6314. [Google Scholar] [CrossRef] [PubMed]
- Kim, S.S.; Harford, J.B.; Moghe, M.; Slaughter, T.; Doherty, C.; Chang, E.H. A tumor-targeting nanomedicine carrying the p53 gene crosses the blood-brain barrier and enhances anti-PD-1 immunotherapy in mouse models of glioblastoma. Int. J. Cancer 2019, 145, 2535–2546. [Google Scholar] [CrossRef] [PubMed]
- Collet, G.; Grillon, C.; Nadim, M.; Kieda, C. Trojan horse at cellular level for tumor gene therapies. Gene 2013, 525, 208–216. [Google Scholar] [CrossRef]
- Buddhadev Layek, B.; Sadhukha, T.; Panyam, J.; Prabh, S. Nano-engineered mesenchymal stem cells increase therapeutic efficacy of anticancer drug through true active tumor targeting. Mol. Cancer Ther. 2018, 17, 1196–1206. [Google Scholar] [CrossRef]
- Tamura, R.; Miyoshi, H.; Yoshida, K.; Okano, H.; Toda, M. Recent progress in the research of suicide gene therapy for malignant glioma. Neurosurg. Rev. 2019, 1–21. [Google Scholar] [CrossRef]
- Altanerova, U.; Benejova, K.; Altanerova, V.; Tyciakova, S.; Rychly, B.; Szomolanyi, P.; Ciampor, F.; Cihova, M.; Repiska, V.; Ondicova, K.; et al. Dental pulp mesenchymal stem/stromal cells labeled with iron sucrose release exosomes and cells applied intra-nasally migrate to intracerebral glioblastoma. Neoplasma 2016, 63, 925–933. [Google Scholar] [CrossRef]
- Mueller, S.; Jain, P.; Liang, W.S.; Kilburn, C.; Kline, L.; Gupta, N.; Panditharatna, E.; Magge, S.N.; Zhang, B.; Zhu, Y.; et al. A pilot precision medicine trial for children with diffuse intrinsic pontine glioma-PNOC003: A report from the Pacific Pediatric Neuro-Oncology Consortium. Int. J. Cancer 2019, 145, 1889–1901. [Google Scholar] [CrossRef]
- Altaner, C.; Altanerova, V.; Cihova, M.; Hunakova, L.; Kaiserova, K.; Klepanec, A.; Vulev, I.; Madaric, J. Characterization of mesenchymal stem cells of “no-options” patients with critical limb ischemia treated by autologous bone marrow mononuclear cells. PLoS ONE 2013, 8, e73722. [Google Scholar] [CrossRef]
- Stanko, P.; Kaiserova, K.; Altanerova, V.; Altaner, C. Comparison of human mesenchymal stem cells derived from dental pulp, bone marrow, adipose tissue, and umbilical cord tissue by gene expression. Biomed. Pap. Med. Fac. Univ. Palacky Olomouc. Czech Repub. 2014, 158, 373–377. [Google Scholar] [CrossRef]
- Hlavaty, J.; Hlubinova, K.; Altaner, C. Construction and testing of gene therapy retroviral vector expressing bacterial cytosine deaminase gene. Neoplasma 1999, 6, 267–276. [Google Scholar]




© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Pastorakova, A.; Jakubechova, J.; Altanerova, U.; Altaner, C. Suicide Gene Therapy Mediated with Exosomes Produced by Mesenchymal Stem/Stromal Cells Stably Transduced with HSV Thymidine Kinase. Cancers 2020, 12, 1096. https://doi.org/10.3390/cancers12051096
Pastorakova A, Jakubechova J, Altanerova U, Altaner C. Suicide Gene Therapy Mediated with Exosomes Produced by Mesenchymal Stem/Stromal Cells Stably Transduced with HSV Thymidine Kinase. Cancers. 2020; 12(5):1096. https://doi.org/10.3390/cancers12051096
Chicago/Turabian StylePastorakova, Andrea, Jana Jakubechova, Ursula Altanerova, and Cestmir Altaner. 2020. "Suicide Gene Therapy Mediated with Exosomes Produced by Mesenchymal Stem/Stromal Cells Stably Transduced with HSV Thymidine Kinase" Cancers 12, no. 5: 1096. https://doi.org/10.3390/cancers12051096
APA StylePastorakova, A., Jakubechova, J., Altanerova, U., & Altaner, C. (2020). Suicide Gene Therapy Mediated with Exosomes Produced by Mesenchymal Stem/Stromal Cells Stably Transduced with HSV Thymidine Kinase. Cancers, 12(5), 1096. https://doi.org/10.3390/cancers12051096