Pharmacological Modulation of SAMHD1 Activity by CDK4/6 Inhibitors Improves Anticancer Therapy

 ,
,  , , , ,
, , , , 
Abstract
1. Introduction
2. Results
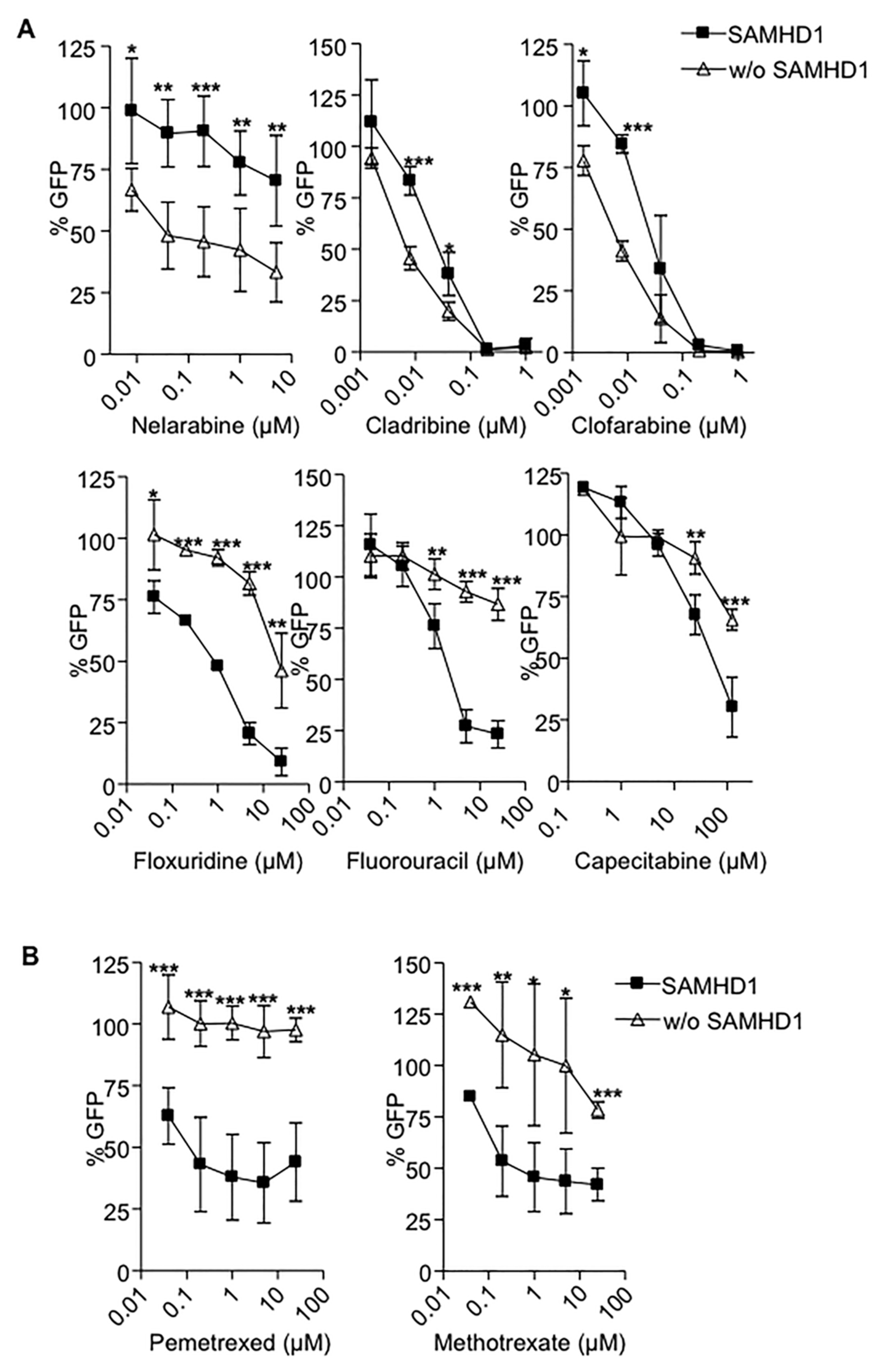
2.1. SAMHD1 Regulates Antiviral Efficacy of Antimetabolites in Primary Cells
2.2. SAMHD1 Is Required for Antiviral Activity of CDK4/6 Inhibitors
2.3. Pharmacological Inhibition of CDK4/6 Enhances Antiviral Activity of Antimetabolites
2.4. Cytotoxic Efficacy of Antimetabolites Is Enhanced by CDK4/6 Inhibitors
2.5. Alternative Pathways of dNTP Metabolism Control Are Responsible for Drug Synergy
2.6. SAMHD1 Is Expressed in Different Tumor Tissues
3. Discussion
4. Materials and Methods
4.1. Cells
4.2. Drugs
4.3. Virus
4.4. Virus Infections
4.5. Western Blot
4.6. Evaluation of Cytotoxicity
4.7. Evaluation of Drug Combination
4.8. Immunohistochemistry
4.9. Statistical Methods
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Goldstone, D.C.; Ennis-Adeniran, V.; Hedden, J.J.; Groom, H.C.; Rice, G.I.; Christodoulou, E.; Walker, P.A.; Kelly, G.; Haire, L.F.; Yap, M.W.; et al. HIV-1 restriction factor SAMHD1 is a deoxynucleoside triphosphate triphosphohydrolase. Nature 2011, 480, 379–382. [Google Scholar] [CrossRef]
- Franzolin, E.; Pontarin, G.; Rampazzo, C.; Miazzi, C.; Ferraro, P.; Palumbo, E.; Reichard, P.; Bianchi, V. The deoxynucleotide triphosphohydrolase SAMHD1 is a major regulator of DNA precursor pools in mammalian cells. Proc. Natl. Acad. Sci. USA 2013, 110, 14272–14277. [Google Scholar] [CrossRef]
- Rice, G.I.; Bond, J.; Asipu, A.; Brunette, R.L.; Manfield, I.W.; Carr, I.M.; Fuller, J.C.; Jackson, R.M.; Lamb, T.; Briggs, T.A.; et al. Mutations involved in Aicardi-Goutières syndrome implicate SAMHD1 as regulator of the innate immune response. Nat. Genet. 2009, 41, 829–832. [Google Scholar] [CrossRef]
- Clifford, R.; Louis, T.; Robbe, P.; Ackroyd, S.; Burns, A.; Timbs, A.T.; Wright Colopy, G.; Dreau, H.; Sigaux, F.; Judde, J.G.; et al. SAMHD1 is mutated recurrently in chronic lymphocytic leukemia and is involved in response to DNA damage. Blood 2014, 123, 1021–1031. [Google Scholar] [CrossRef]
- Rentoft, M.; Lindell, K.; Tran, P.; Chabes, A.L.; Buckland, R.J.; Watt, D.L.; Marjavaara, L.; Nilsson, A.K.; Melin, B.; Trygg, J.; et al. Heterozygous colon cancer-associated mutations of SAMHD1 have functional significance. Proc. Natl. Acad. Sci. USA 2016, 113, 4723–4728. [Google Scholar] [CrossRef]
- de Silva, S.; Wang, F.; Hake, T.S.; Porcu, P.; Wong, H.K.; Wu, L. Downregulation of SAMHD1 expression correlates with promoter DNA methylation in Sézary syndrome patients. J. Investig. Dermatol. 2014, 134, 562–565. [Google Scholar] [CrossRef]
- Wang, J.; Lu, F.; Shen, X.-Y.; Wu, Y.; Zhao, L. SAMHD1 is down regulated in lung cancer by methylation and inhibits tumor cell proliferation. Biochem. Biophys. Res. Commun. 2014, 455, 229–233. [Google Scholar] [CrossRef]
- Coquel, F.; Silva, M.-J.; Técher, H.; Zadorozhny, K.; Sharma, S.; Nieminuszczy, J.; Mettling, C.; Dardillac, E.; Barthe, A.; Schmitz, A.-L.; et al. SAMHD1 acts at stalled replication forks to prevent interferon induction. Nature 2018, 557, 57–61. [Google Scholar] [CrossRef]
- Ballana, E.; Esté, J.A. SAMHD1: At the crossroads of cell proliferation, immune responses, and virus restriction. Trends Microbiol. 2015, 23, 680–692. [Google Scholar] [CrossRef]
- Laguette, N.; Sobhian, B.; Casartelli, N.; Ringeard, M.; Chable-Bessia, C.; Ségéral, E.; Yatim, A.; Emiliani, S.; Schwartz, O.; Benkirane, M. SAMHD1 is the dendritic- and myeloid-cell-specific HIV-1 restriction factor counteracted by Vpx. Nature 2011, 474, 654–657. [Google Scholar] [CrossRef]
- Lahouassa, H.; Daddacha, W.; Hofmann, H.; Ayinde, D.; Logue, E.C.; Dragin, L.; Bloch, N.; Maudet, C.; Bertrand, M.; Gramberg, T.; et al. SAMHD1 restricts the replication of human immunodeficiency virus type 1 by depleting the intracellular pool of deoxynucleoside triphosphates. Nat. Immunol. 2012, 13, 223–228. [Google Scholar] [CrossRef]
- Hrecka, K.; Hao, C.; Gierszewska, M.; Swanson, S.K.; Kesik-Brodacka, M.; Srivastava, S.; Florens, L.; Washburn, M.P.; Skowronski, J. Vpx relieves inhibition of HIV-1 infection of macrophages mediated by the SAMHD1 protein. Nature 2011, 474, 658–661. [Google Scholar] [CrossRef]
- Cribier, A.; Descours, B.; Valadão, A.L.C.; Laguette, N.; Benkirane, M. Phosphorylation of SAMHD1 by cyclin A2/CDK1 regulates its restriction activity toward HIV-1. Cell Rep. 2013, 3, 1036–1043. [Google Scholar] [CrossRef]
- Pauls, E.; Ruiz, A.; Badia, R.; Permanyer, M.; Gubern, A.; Riveira-Muñoz, E.; Torres-Torronteras, J.; Álvarez, M.; Mothe, B.; Brander, C.; et al. Cell Cycle Control and HIV-1 Susceptibility Are Linked by CDK6-Dependent CDK2 Phosphorylation of SAMHD1 in Myeloid and Lymphoid Cells. J. Immunol. 2014, 193, 1988–1997. [Google Scholar] [CrossRef]
- White, T.E.; Brandariz-Nuñez, A.; Valle-Casuso, J.C.; Amie, S.; Nguyen, L.A.; Kim, B.; Tuzova, M.; Diaz-Griffero, F. The retroviral restriction ability of SAMHD1, but not its deoxynucleotide triphosphohydrolase activity, is regulated by phosphorylation. Cell Host Microbe 2013, 13, 441–451. [Google Scholar] [CrossRef]
- Bermejo, M.; López-Huertas, M.R.; García-Pérez, J.; Climent, N.; Descours, B.; Ambrosioni, J.; Mateos, E.; Rodríguez-Mora, S.; Rus-Bercial, L.; Benkirane, M.; et al. Dasatinib inhibits HIV-1 replication through the interference of SAMHD1 phosphorylation in CD4+ T cells. Biochem. Pharmacol. 2016, 106, 30–45. [Google Scholar] [CrossRef]
- Mlcochova, P.; Caswell, S.J.; Taylor, I.A.; Towers, G.J.; Gupta, R.K. DNA damage induced by topoisomerase inhibitors activates SAMHD1 and blocks HIV-1 infection of macrophages. EMBO J. 2018, 37, 50–62. [Google Scholar] [CrossRef]
- Mlcochova, P.; Sutherland, K.A.; Watters, S.A.; Bertoli, C.; de Bruin, R.A.; Rehwinkel, J.; Neil, S.J.; Lenzi, G.M.; Kim, B.; Khwaja, A.; et al. A G1-like state allows HIV-1 to bypass SAMHD1 restriction in macrophages. EMBO J. 2017, 36, 604–616. [Google Scholar] [CrossRef]
- Pauls, E.; Badia, R.; Torres-Torronteras, J.; Ruiz, A.; Permanyer, M.; Riveira-Muñoz, E.; Clotet, B.; Marti, R.; Ballana, E.; Esté, J.A. Palbociclib, a selective inhibitor of cyclin-dependent kinase4/6, blocks HIV-1 reverse transcription through the control of sterile α motif and HD domain-containing protein-1 (SAMHD1) activity. AIDS 2014, 28, 2213–2222. [Google Scholar] [CrossRef]
- Szaniawski, M.A.; Spivak, A.M.; Cox, J.E.; Catrow, J.L.; Hanley, T.; Williams, E.S.C.P.; Tremblay, M.J.; Bosque, A.; Planelles, V. SAMHD1 Phosphorylation Coordinates the Anti-HIV-1 Response by Diverse Interferons and Tyrosine Kinase Inhibition. MBio 2018, 9, e00819-18. [Google Scholar] [CrossRef]
- Jordheim, L.P.; Durantel, D.; Zoulim, F.; Dumontet, C. Advances in the development of nucleoside and nucleotide analogues for cancer and viral diseases. Nat. Rev. Drug Discov. 2013, 12, 447–464. [Google Scholar] [CrossRef] [PubMed]
- Badia, R.; Pujantell, M.; Torres-Torronteras, J.; Menéndez-Arias, L.; Martí, R.; Ruzo, A.; Pauls, E.; Clotet, B.; Ballana, E.; Esté, J.A.; et al. SAMHD1 is active in cycling cells permissive to HIV-1 infection. Antivir. Res. 2017, 142, 123–135. [Google Scholar] [CrossRef]
- Ballana, E.; Badia, R.; Terradas, G.; Torres-Torronteras, J.; Ruiz, A.; Pauls, E.; Riveira-Muñoz, E.; Clotet, B.; Martí, R.; Esté, J.A. SAMHD1 Specifically Affects the Antiviral Potency of Thymidine Analog HIV Reverse Transcriptase Inhibitors. Antimicrob. Agents Chemother. 2014, 58, 4804–4813. [Google Scholar] [CrossRef][Green Version]
- Amie, S.M.; Daly, M.B.; Noble, E.; Schinazi, R.F.; Bambara, R.A.; Kim, B. Anti-HIV host factor SAMHD1 regulates viral sensitivity to nucleoside reverse transcriptase inhibitors via modulation of cellular deoxyribonucleoside triphosphate (dNTP) levels. J. Biol. Chem. 2013, 288, 20683–20691. [Google Scholar] [CrossRef]
- Huber, A.D.; Michailidis, E.; Schultz, M.L.; Ong, Y.T.; Bloch, N.; Puray-Chavez, M.N.; Leslie, M.D.; Ji, J.; Lucas, A.D.; Kirby, K.A.; et al. SAMHD1 has differential impact on the efficacies of HIV nucleoside reverse transcriptase inhibitors. Antimicrob. Agents Chemother. 2014, 58, 4915–4919. [Google Scholar] [CrossRef]
- Ordonez, P.; Kunzelmann, S.; Groom, H.C.T.; Yap, M.W.; Weising, S.; Meier, C.; Bishop, K.N.; Taylor, I.A.; Stoye, J.P. SAMHD1 enhances nucleoside-analogue efficacy against HIV-1 in myeloid cells. Sci. Rep. 2017, 7, 42824. [Google Scholar] [CrossRef]
- Herold, N.; Rudd, S.G.; Sanjiv, K.; Kutzner, J.; Bladh, J.; Paulin, C.B.J.; Helleday, T.; Henter, J.-I.; Schaller, T. SAMHD1 protects cancer cells from various nucleoside-based antimetabolites. Cell Cycle 2017, 16, 1029–1038. [Google Scholar] [CrossRef]
- Herold, N.; Rudd, S.G.; Ljungblad, L.; Sanjiv, K.; Myrberg, I.H.; Paulin, C.B.J.; Heshmati, Y.; Hagenkort, A.; Kutzner, J.; Page, B.D.G.; et al. Targeting SAMHD1 with the Vpx protein to improve cytarabine therapy for hematological malignancies. Nat. Med. 2017, 23, 256–263. [Google Scholar] [CrossRef]
- Schneider, C.; Oellerich, T.; Baldauf, H.-M.; Schwarz, S.-M.; Thomas, D.; Flick, R.; Bohnenberger, H.; Kaderali, L.; Stegmann, L.; Cremer, A.; et al. SAMHD1 is a biomarker for cytarabine response and a therapeutic target in acute myeloid leukemia. Nat. Med. 2017, 23, 250–255. [Google Scholar] [CrossRef]
- Knecht, K.M.; Buzovetsky, O.; Schneider, C.; Thomas, D.; Srikanth, V.; Kaderali, L.; Tofoleanu, F.; Reiss, K.; Ferreirós, N.; Geisslinger, G.; et al. The structural basis for cancer drug interactions with the catalytic and allosteric sites of SAMHD1. Proc. Natl. Acad. Sci. USA 2018, 115, E10022–E10031. [Google Scholar] [CrossRef]
- Hollenbaugh, J.A.; Shelton, J.; Tao, S.; Amiralaei, S.; Liu, P.; Lu, X.; Goetze, R.W.; Zhou, L.; Nettles, J.H.; Schinazi, R.F.; et al. Substrates and Inhibitors of SAMHD1. PLoS ONE 2017, 12, e0169052. [Google Scholar] [CrossRef] [PubMed]
- Rassidakis, G.Z.; Herold, N.; Myrberg, I.H.; Tsesmetzis, N.; Rudd, S.G.; Henter, J.-I.; Schaller, T.; Ng, S.-B.; Chng, W.J.; Yan, B.; et al. Low-level expression of SAMHD1 in acute myeloid leukemia (AML) blasts correlates with improved outcome upon consolidation chemotherapy with high-dose cytarabine-based regimens. Blood Cancer J. 2018, 8, 98. [Google Scholar] [CrossRef] [PubMed]
- Uhlen, M.; Zhang, C.; Lee, S.; Sjöstedt, E.; Fagerberg, L.; Bidkhori, G.; Benfeitas, R.; Arif, M.; Liu, Z.; Edfors, F.; et al. A pathology atlas of the human cancer transcriptome. Science 2017, 357, eaan2507. [Google Scholar] [CrossRef] [PubMed]
- Aird, K.M.; Zhang, R. Nucleotide metabolism, oncogene-induced senescence and cancer. Cancer Lett. 2015, 356, 204–210. [Google Scholar] [CrossRef]
- Herold, N.; Rudd, S.G.; Sanjiv, K.; Kutzner, J.; Myrberg, I.H.; Paulin, C.B.J.; Olsen, T.K.; Helleday, T.; Henter, J.-I.; Schaller, T. With me or against me: Tumor suppressor and drug resistance activities of SAMHD1. Exp. Hematol. 2017, 52, 32–39. [Google Scholar] [CrossRef]
- Joerger, M.; Omlin, A.; Cerny, T.; Früh, M. The role of pemetrexed in advanced non small-cell lung cancer: Special focus on pharmacology and mechanism of action. Curr. Drug Targets 2010, 11, 37–47. [Google Scholar] [CrossRef]
- Tsesmetzis, N.; Paulin, C.B.J.; Rudd, S.G.; Herold, N. Nucleobase and Nucleoside Analogues: Resistance and Re-Sensitisation at the Level of Pharmacokinetics, Pharmacodynamics and Metabolism. Cancers 2018, 10, 240. [Google Scholar] [CrossRef]
- Turner, N.C.; Ro, J.; André, F.; Loi, S.; Verma, S.; Iwata, H.; Harbeck, N.; Loibl, S.; Huang Bartlett, C.; Zhang, K.; et al. Palbociclib in Hormone-Receptor-Positive Advanced Breast Cancer. N. Engl. J. Med. 2015, 373, 209–219. [Google Scholar] [CrossRef]
- Sherr, C.J.; Beach, D.; Shapiro, G.I. Targeting CDK4 and CDK6: From Discovery to Therapy. Cancer Discov. 2016, 6, 353–367. [Google Scholar] [CrossRef]
- O’Leary, B.; Finn, R.S.; Turner, N.C. Treating cancer with selective CDK4/6 inhibitors. Nat. Rev. Clin. Oncol. 2016, 13, 417–430. [Google Scholar] [CrossRef]
- Patnaik, A.; Rosen, L.S.; Tolaney, S.M.; Tolcher, A.W.; Goldman, J.W.; Gandhi, L.; Papadopoulos, K.P.; Beeram, M.; Rasco, D.W.; Hilton, J.F.; et al. Efficacy and Safety of Abemaciclib, an Inhibitor of CDK4 and CDK6, for Patients with Breast Cancer, Non-Small Cell Lung Cancer, and Other Solid Tumors. Cancer Discov. 2016, 6, 740–753. [Google Scholar] [CrossRef] [PubMed]
- Malumbres, M.; Barbacid, M. Cell Cycle, CDKs and Cancer: A Changing Paradigm. Nat. Rev. Cancer 2009, 9, 153–166. [Google Scholar] [CrossRef] [PubMed]
- Pujantell, M.; Franco, S.; Galván-Femenía, I.; Badia, R.; Castellví, M.; Garcia-Vidal, E.; Clotet, B.; de Cid, R.; Tural, C.; Martínez, M.A.; et al. ADAR1 affects HCV infection by modulating innate immune response. Antivir. Res. 2018, 156, 116–127. [Google Scholar] [CrossRef] [PubMed]
- Badia, R.; Ballana, E.; Castellví, M.; García-Vidal, E.; Pujantell, M.; Clotet, B.; Prado, J.G.; Puig, J.; Martínez, M.A.; Riveira-Muñoz, E.; et al. CD32 expression is associated to T-cell activation and is not a marker of the HIV-1 reservoir. Nat. Commun. 2018, 9, 2739. [Google Scholar] [CrossRef]
- Zhang, N.; Fu, J.-N.; Chou, T.-C. Synergistic combination of microtubule targeting anticancer fludelone with cytoprotective panaxytriol derived from panax ginseng against MX-1 cells in vitro: Experimental design and data analysis using the combination index method. Am. J. Cancer Res. 2016, 6, 97–104. [Google Scholar]
- Chou, T.C.; Talalay, P. Quantitative analysis of dose-effect relationships: The combined effects of multiple drugs or enzyme inhibitors. Adv. Enzym. Regul. 1984, 22, 27–55. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Drugs | Drug Type (Base Targeted) | EC50 (µM) | FC | ||
|---|---|---|---|---|---|
| SAMHD1 (+) | SAMHD1 (-) | -/+ | +/- | ||
| AZT | NRTI (dT) | 0.006 | 0.11 | 18 | - |
| NVP | NNRTI (none) | 0.88 | 0.95 | 1 | 1 |
| AraC | Pyrimidine (dC) | 3.24 | 0.11 | - | 30 |
| Nelarabine | Purine (dG) | 13.96 | 1.83 | - | 8 |
| Cladribine | Purine (dA) | 0.029 | 0.007 | - | 4 |
| Clofarabine | Purine (dG) | 0.034 | 0.006 | - | 6 |
| Floxuridine | Pyrimidine (dU) | 0.73 | 20.28 | 28 | - |
| Fluorouracil | Pyrimidine (dU) | 2.40 | >25 | >10 | - |
| Pemetrexed | Anti-folate | 0.25 | >25 | >100 | - |
| Methotrexate | Anti-folate | 0.42 | 79.24 | 190 | - |
| Drug Combination | Combination Index (CI) | Effect | |
|---|---|---|---|
| Pemetrexed | 25 | 0.0049 | Synergy |
| + | 5 | 0.0097 | Synergy |
| Palbociclib 0.04 µM | 1 | 0.0285 | Synergy |
| 0.2 | 0.0415 | Synergy | |
| 0.04 | 0.0673 | Synergy | |
| Pemetrexed | 25 | 0.079 | Synergy |
| + | 5 | 0.069 | Synergy |
| Midostaurin 0.2 µM | 1 | 0.056 | Synergy |
| 0.2 | 0.045 | Synergy | |
| 0.04 | 0.064 | Synergy | |
| Fluorouracil | 5 | 1.871 | Antagonism |
| + | 1 | 0.572 | Synergy |
| Palbociclib 0.04 µM | 0.2 | 0.658 | Synergy |
| 0.04 | 1.818 | Antagonism | |
| 0.008 | 2.967 | Antagonism | |
| Midostaurin | 5 | 2.074 | Antagonism |
| + | 1 | 0.427 | Synergy |
| Palbociclib 0.04 µM | 0.2 | 0.223 | Synergy |
| 0.04 | 0.419 | Synergy | |
| 0.008 | 0.324 | Synergy | |
| Cell Type | Drug Combination | Combination Index (CI) | Effect | |
|---|---|---|---|---|
| TZM-bl | Pemetrexed | 1 | 0.806 | Synergy |
| + | 0.2 | 0.689 | Synergy | |
| Palbociclib 5 µM | 0.04 | 0.71 | Synergy | |
| 0.008 | 0.72 | Synergy | ||
| Fluorouracil | 5 | 0.726 | Synergy | |
| + | 1 | 0.766 | Synergy | |
| Palbociclib 5 µM | 0.2 | 0.764 | Synergy | |
| 0.04 | 0.735 | Synergy | ||
| T47D | Pemetrexed | 5 | 0.783 | Synergy |
| + | 1 | 0.707 | Synergy | |
| Palbociclib 5 µM | 0.2 | 0.779 | Synergy | |
| 0.04 | 0.874 | Synergy | ||
| Fluorouracil | 25 | 0.848 | Synergy | |
| + | 5 | 0.804 | Synergy | |
| Palbociclib 5 µM | 1 | 0.966 | Additive | |
| 0.2 | 1.01 | Additive | ||
| MDA-MB-468 | Pemetrexed | 1 | 0.886 | Synergy |
| + | 0.2 | 0.886 | Synergy | |
| Palbociclib 5 µM | 0.04 | 0.921 | Synergy | |
| 0.008 | 1.01 | Additive | ||
| Fluorouracil | 25 | 2.844 | Antagonism | |
| + | 5 | 1.745 | Antagonism | |
| Palbociclib 5 µM | 1 | 1.356 | Antagonism | |
| 0.2 | 1.125 | Antagonism | ||
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Castellví, M.; Felip, E.; Ezeonwumelu, I.J.; Badia, R.; Garcia-Vidal, E.; Pujantell, M.; Gutiérrez-Chamorro, L.; Teruel, I.; Martínez-Cardús, A.; Clotet, B.; et al. Pharmacological Modulation of SAMHD1 Activity by CDK4/6 Inhibitors Improves Anticancer Therapy. Cancers 2020, 12, 713. https://doi.org/10.3390/cancers12030713
Castellví M, Felip E, Ezeonwumelu IJ, Badia R, Garcia-Vidal E, Pujantell M, Gutiérrez-Chamorro L, Teruel I, Martínez-Cardús A, Clotet B, et al. Pharmacological Modulation of SAMHD1 Activity by CDK4/6 Inhibitors Improves Anticancer Therapy. Cancers. 2020; 12(3):713. https://doi.org/10.3390/cancers12030713
Chicago/Turabian StyleCastellví, Marc, Eudald Felip, Ifeanyi Jude Ezeonwumelu, Roger Badia, Edurne Garcia-Vidal, Maria Pujantell, Lucía Gutiérrez-Chamorro, Iris Teruel, Anna Martínez-Cardús, Bonaventura Clotet, and et al. 2020. "Pharmacological Modulation of SAMHD1 Activity by CDK4/6 Inhibitors Improves Anticancer Therapy" Cancers 12, no. 3: 713. https://doi.org/10.3390/cancers12030713
APA StyleCastellví, M., Felip, E., Ezeonwumelu, I. J., Badia, R., Garcia-Vidal, E., Pujantell, M., Gutiérrez-Chamorro, L., Teruel, I., Martínez-Cardús, A., Clotet, B., Riveira-Muñoz, E., Margelí, M., Esté, J. A., & Ballana, E. (2020). Pharmacological Modulation of SAMHD1 Activity by CDK4/6 Inhibitors Improves Anticancer Therapy. Cancers, 12(3), 713. https://doi.org/10.3390/cancers12030713