PLAG Exerts Anti-Metastatic Effects by Interfering with Neutrophil Elastase/PAR2/EGFR Signaling in A549 Lung Cancer Orthotopic Model


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Results
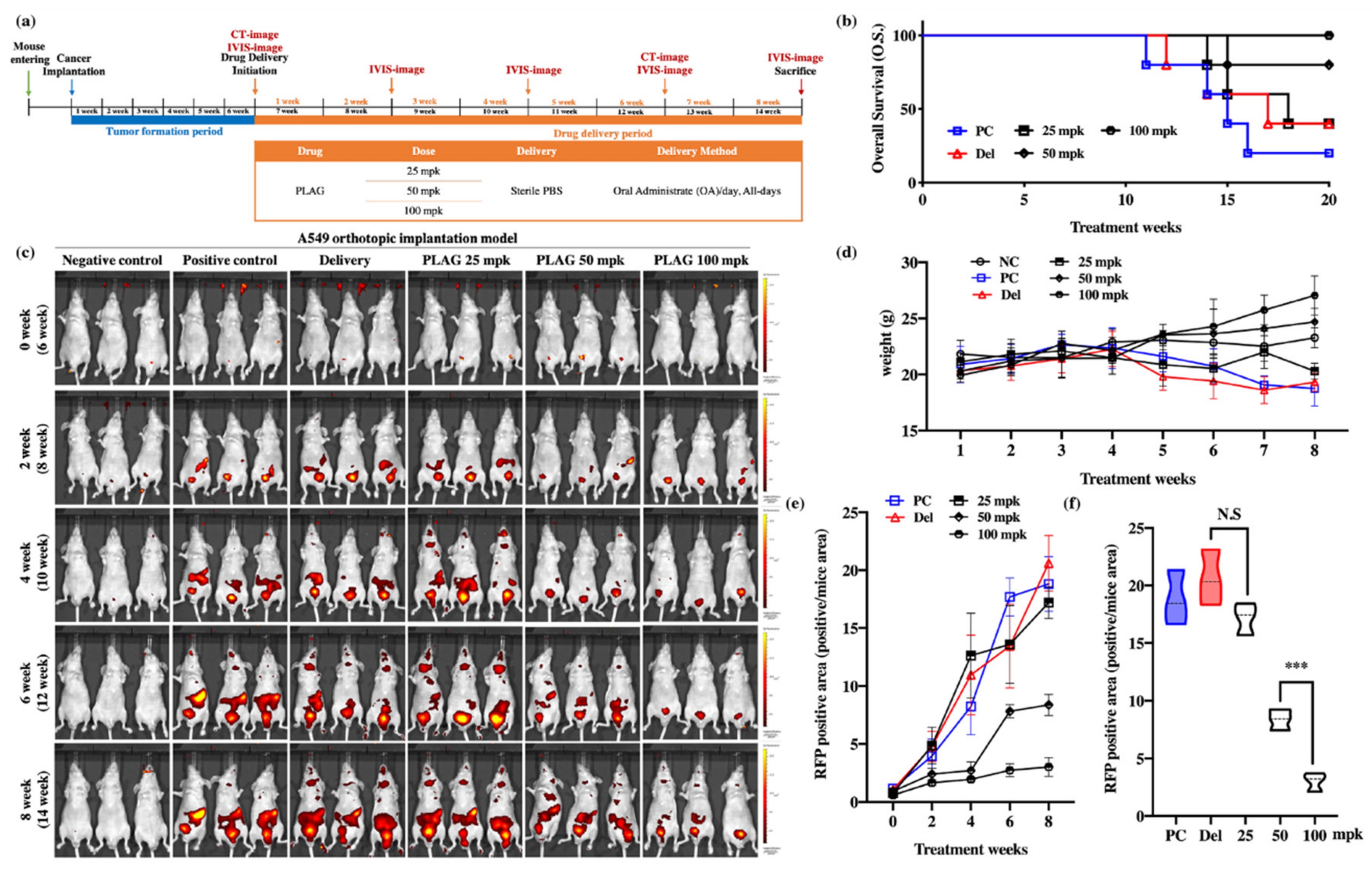
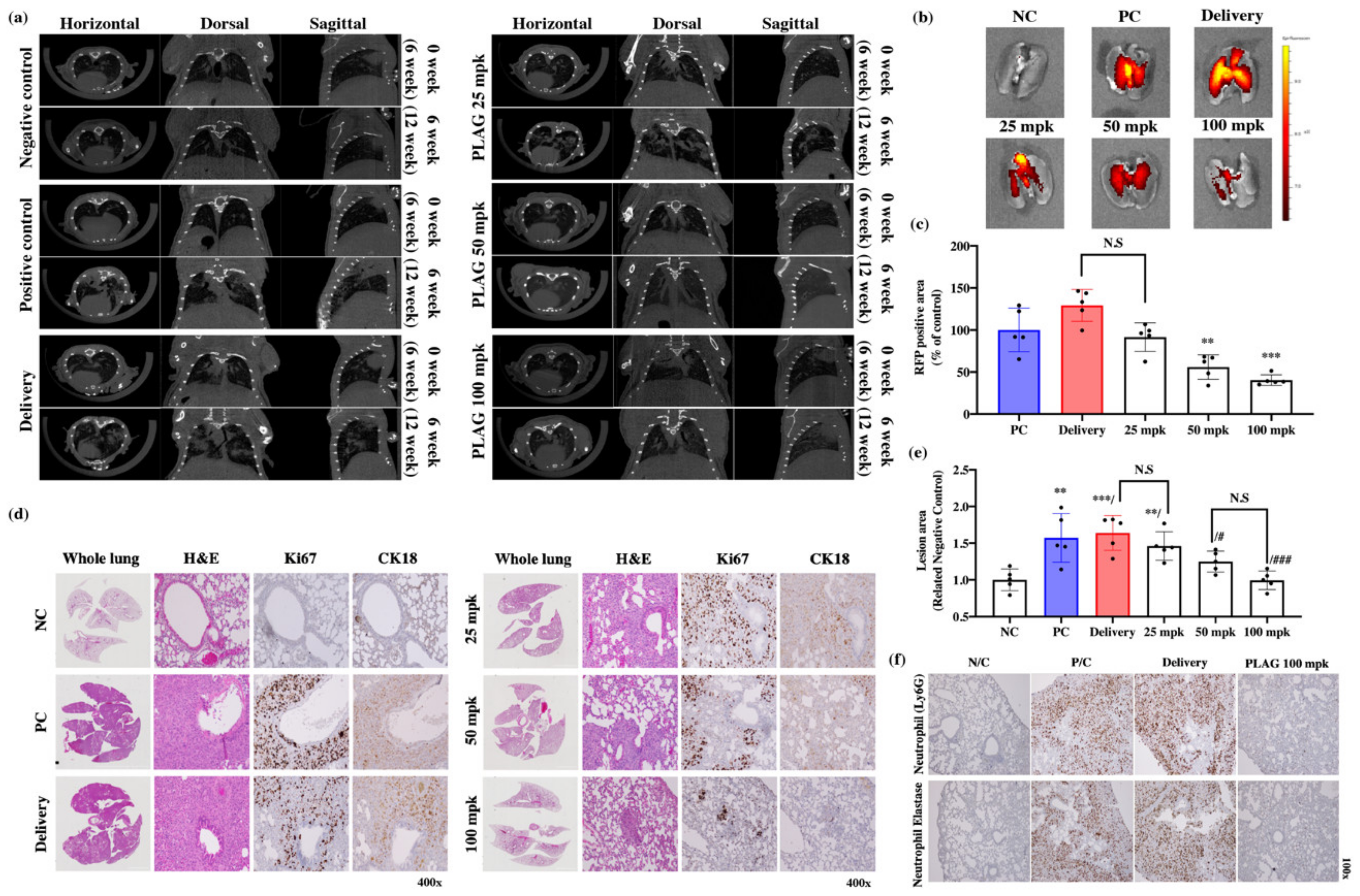
2.1. PLAG Inhibited Metastasis of A549 Human Lung Cancer in an Orthotopic Mouse Model.
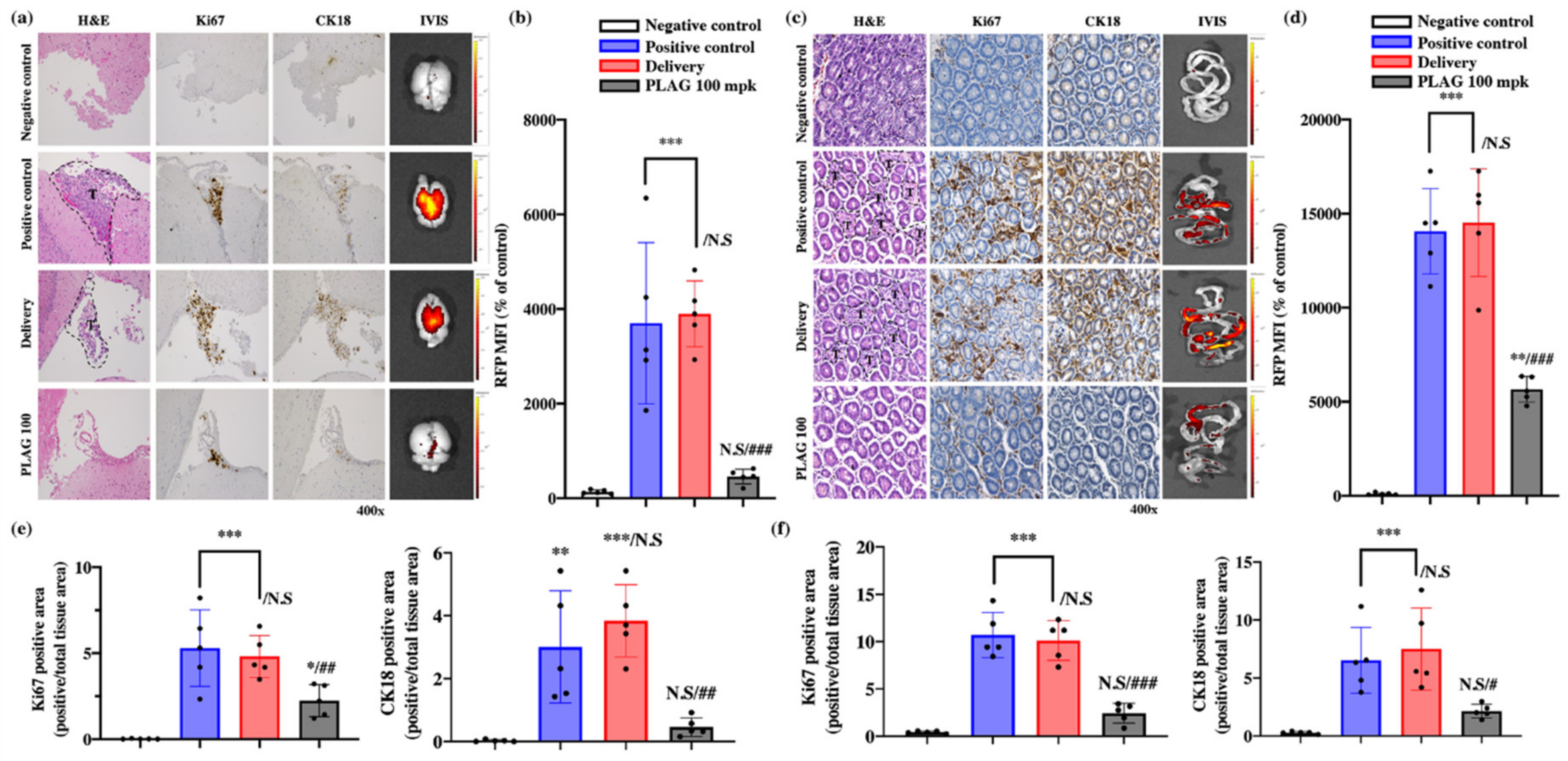
2.2. Treatment with 100 mpk PLAG Reduced Metastasis to the Brain and Intestines.
2.3. PLAG Treatment Inhibited the Growth of A549 Human Lung Cancer in Mice.
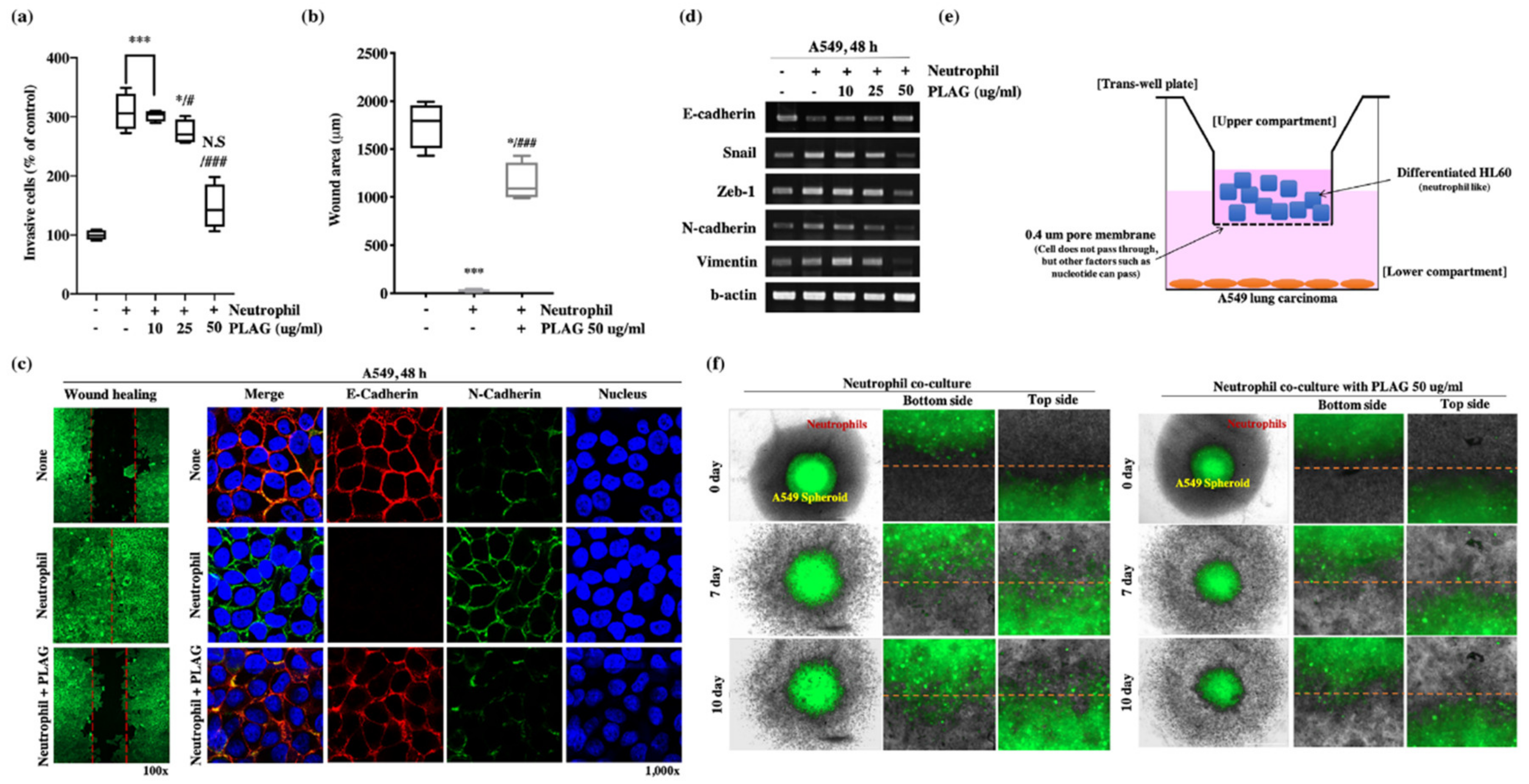
2.4. The Metastatic Properties of A549 Cells were Promoted by the Tumor-Infiltrating Neutrophil Environment and Attenuated by PLAG.
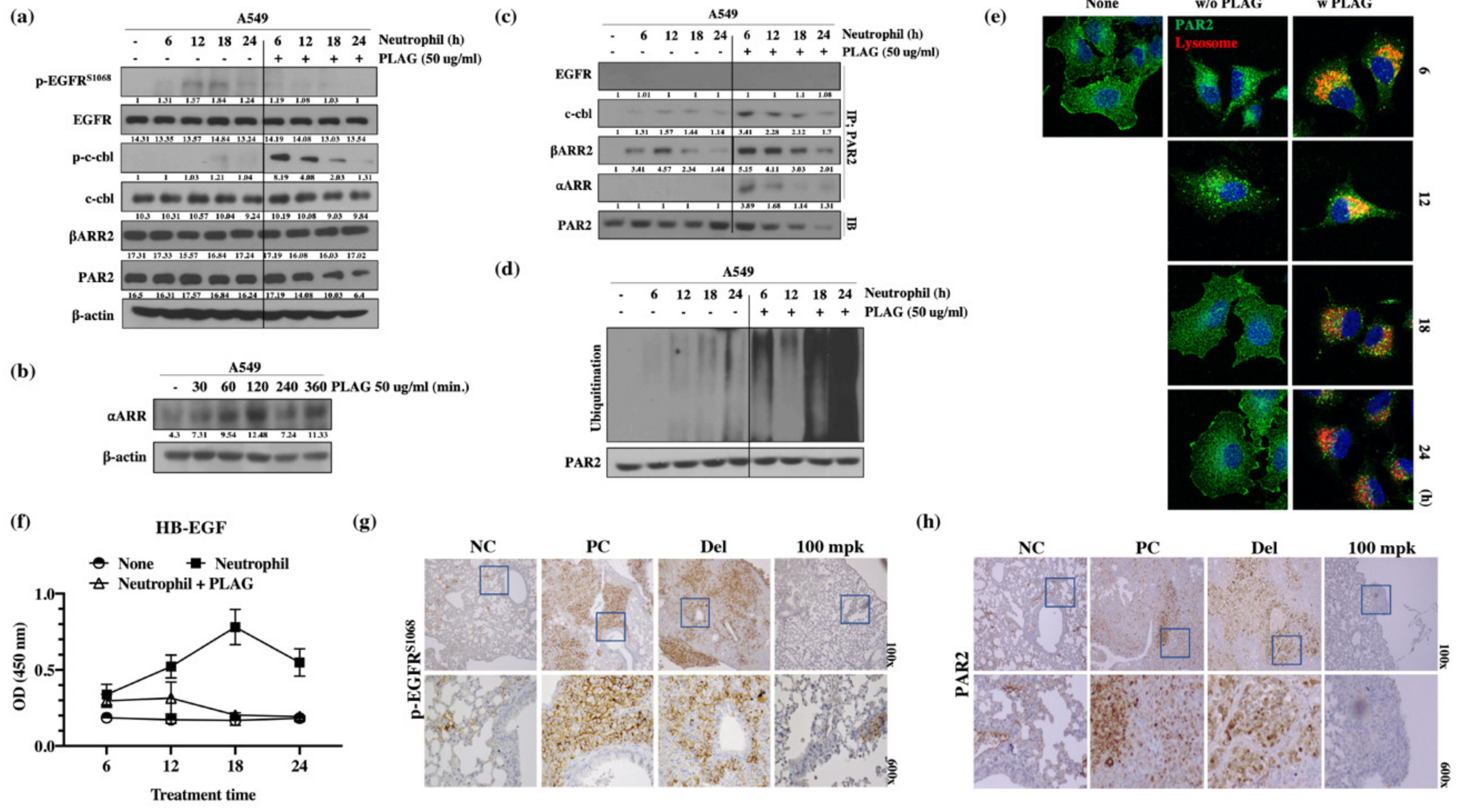
2.5. EGFR Transactivation for Metastatic Activity was Mediated by the PAR2 Receptor in the Presence of Neutrophils.
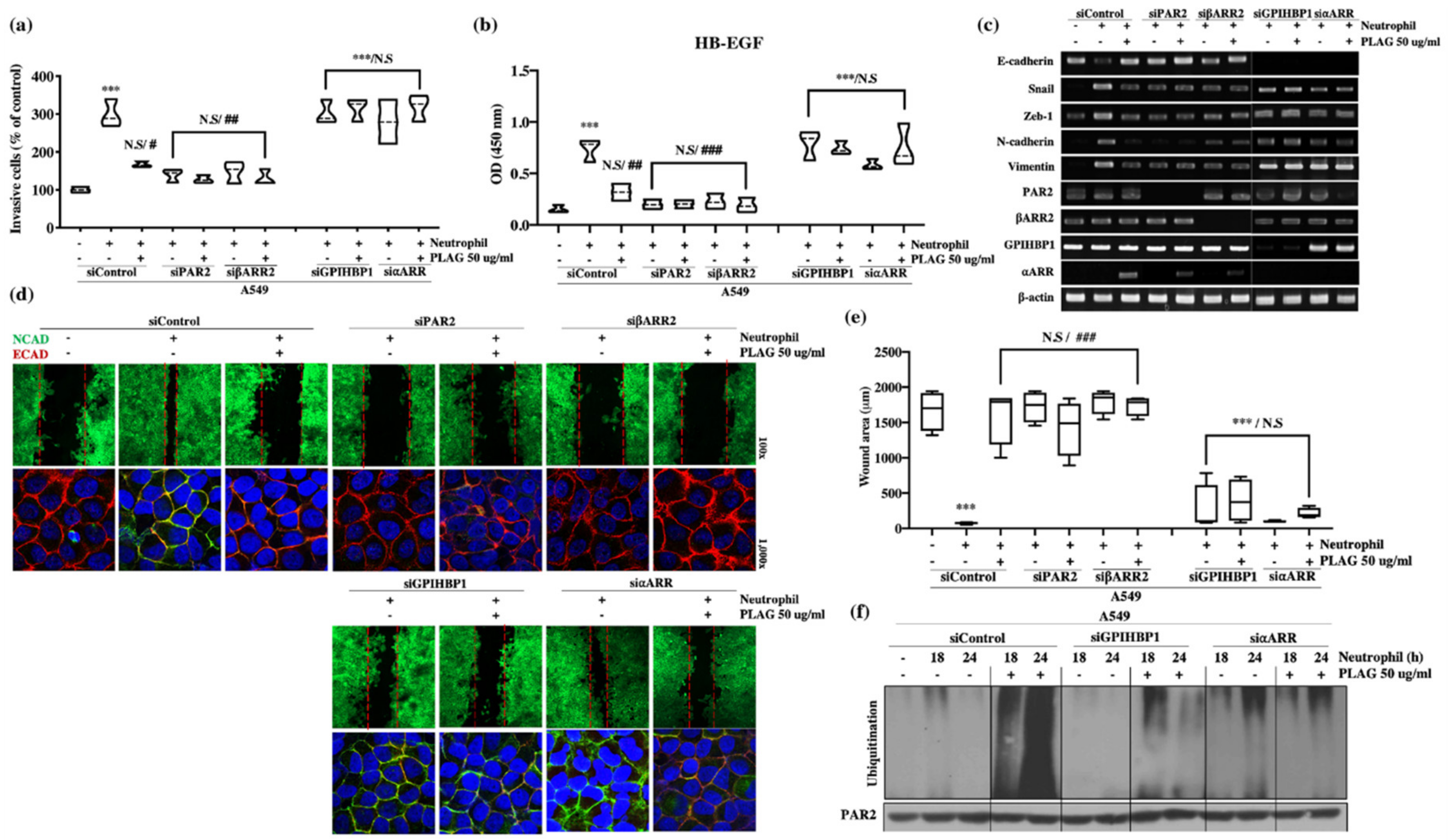
2.6. Neutrophil-Induced Metastatic Activity of A549 Cells was Dependent on PAR2 and β-Arrestin-2.
2.7. The Inhibitory Effect of PLAG on Neutrophil-Induced Metastatic Activity in A549 Cells was Dependent on GPIHBP1 and ⍺-Arrestin.
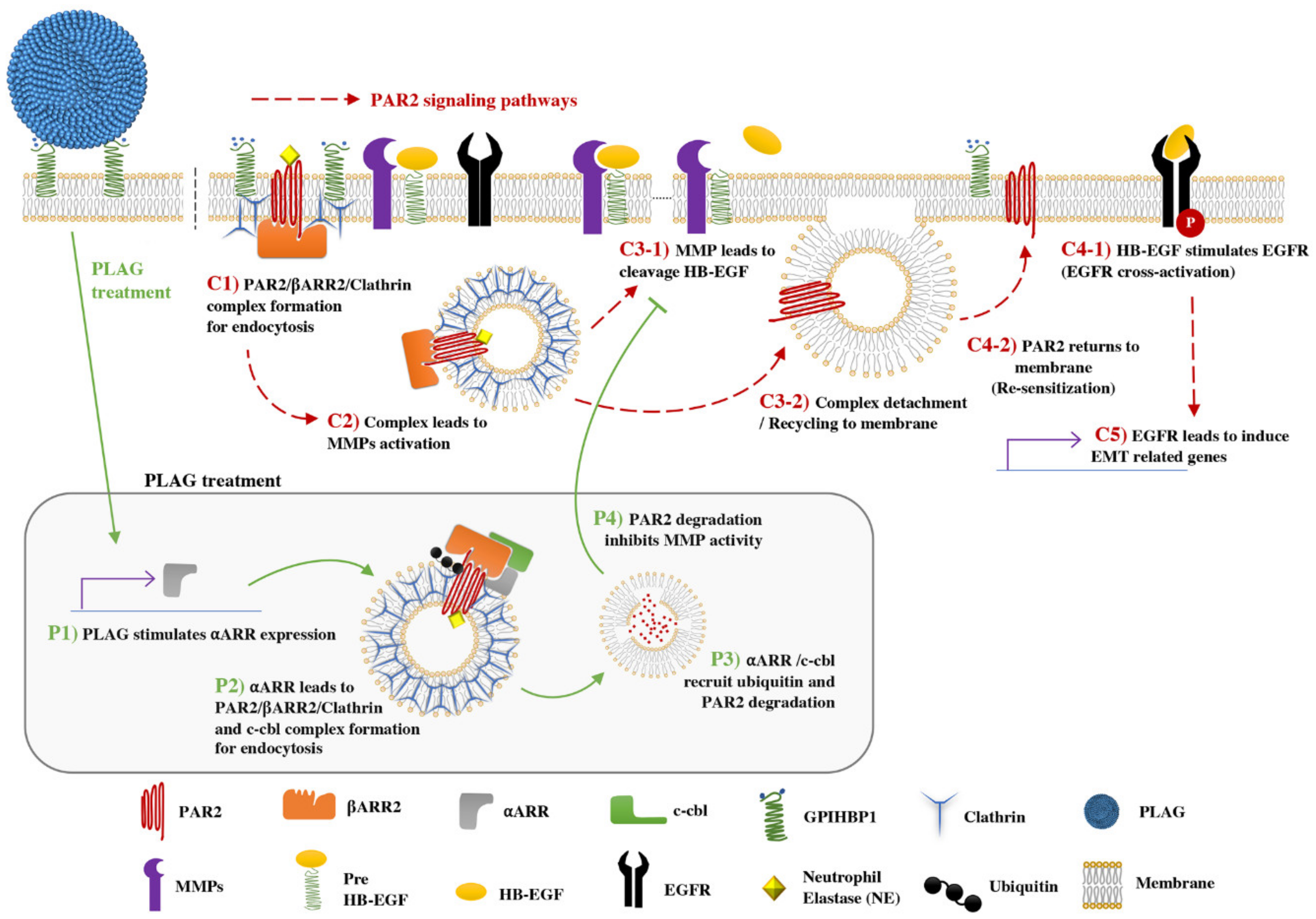
3. Discussion
4. Materials and Methods
4.1. Cell Culture
4.2. Invasion Assay to Measure the Migration Activity of Cancer Cells
4.3. Co-Culture System for Indirect Contact using Separated Chambers
4.4. Immunofluorescence Staining and Wound Healing Assay
4.5. Spheroid-Formation Assay to Examine Cancer Cell Scattering
4.6. Analysis of Protein Degradation by Ubiquitination Assay
4.7. Analysis of Secreted Proteins by ELISA
4.8. Examination by Confocal Microscopy for Lysosome Co-Localization
4.9. Analysis of Protein Assemblies by Immunoprecipitation
4.10. Analysis of Expressed Transcripts using Quantitative PCR
4.11. Gene Silencing by Small Interfering RNA (siRNA)
4.12. Lung Cancer Orthotopic Implantation Model
4.13. Overall Survival Analysis
4.14. Tumor Mass Scan by Computed Tomography
4.15. Immunohistochemistry Staining
4.16. Hematoxylin and Eosin Staining
4.17. Statistics
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Bray, F.; Ferlay, J.; Soerjomataram, I.; Siegel, R.L.; Torre, L.A.; Jemal, A. Global cancer statistics 2018: Globocan estimates of incidence and mortality worldwide for 36 cancers in 185 countries. CA A Cancer J. Clin. 2018, 68, 394–424. [Google Scholar] [CrossRef] [PubMed]
- Miller, K.D.; Siegel, R.L.; Lin, C.; Mariotto, A.B.; Kramer, J.L.; Rowland, J.H.; Stein, K.D.; Alteri, R.; Jemal, A. Cancer treatment and survivorship statistics, 2016. CA A Cancer J. Clin. 2016, 66, 271–289. [Google Scholar] [CrossRef] [PubMed]
- Pérez-Tomás, R. Multidrug resistance: Retrospect and prospects in anti-cancer drug treatment. Curr. Med. Chem. 2006, 13, 1859–1876. [Google Scholar] [CrossRef] [PubMed]
- Molina, J.R.; Yang, P.; Cassivi, S.D.; Schild, S.E.; Adjei, A.A. Non-small cell lung cancer: Epidemiology, risk factors, treatment, and survivorship. Mayo Clin. Proc. 2008, 83, 584–594. [Google Scholar] [CrossRef]
- Poullis, M.; Shackcloth, M.; Page, R.; Asanti-Siaw, J.; Woolley, S.; Mediratta, N. Metastatic index of non-small-cell lung cancer and long-term survival. Asian Cardiovasc. Thorac. Ann. 2015, 23, 185–190. [Google Scholar] [CrossRef] [PubMed]
- Di, J.-Z.; Peng, J.-Y.; Wang, Z.-G. Prevalence, clinicopathological characteristics, treatment, and prognosis of intestinal metastasis of primary lung cancer: A comprehensive review. Surg. Oncol. 2014, 23, 72–80. [Google Scholar] [CrossRef]
- Milovanovic, I.; Stjepanovic, M.; Mitrovic, D. Distribution patterns of the metastases of the lung carcinoma in relation to histological type of the primary tumor: An autopsy study. Ann. Thorac. Med. 2017, 12, 191–198. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.; Zhang, W.; Yuan, X.; Fu, M.; Qian, H.; Xu, W. Neutrophils in cancer development and progression: Roles, mechanisms, and implications (review). Int. J. Oncol. 2016, 49, 857–867. [Google Scholar] [CrossRef]
- Li, S.; Cong, X.; Gao, H.; Lan, X.; Li, Z.; Wang, W.; Song, S.; Wang, Y.; Li, C.; Zhang, H.; et al. Tumor-associated neutrophils induce emt by il-17a to promote migration and invasion in gastric cancer cells. J. Exp. Clin. Cancer Res. 2019, 38, 6. [Google Scholar] [CrossRef]
- Kim, Y.; Lee, D.; Lee, J.; Lee, S.; Lawler, S. Role of tumor-associated neutrophils in regulation of tumor growth in lung cancer development: A mathematical model. PLoS ONE 2019, 14(1), e0211041. [Google Scholar] [CrossRef]
- Lerman, I.; de la Garcia-Hernandez, M.; Rangel-Moreno, J.; Chiriboga, L.; Pan, C.; Nastiuk, K.L.; Krolewski, J.J.; Sen, A.; Hammes, S.R. Infiltrating myeloid cells exert protumorigenic actions via neutrophil elastase. Mol. Cancer Res. 2017, 15, 1138–1152. [Google Scholar] [CrossRef] [PubMed]
- Moroy, G.; Alix, A.J.P.; Sapi, J.; Hornebeck, W.; Bourguet, E. Neutrophil elastase as a target in lung cancer. Anti Cancer Agents Med. Chem. 2012, 12, 565–579. [Google Scholar] [CrossRef] [PubMed]
- Feliciano, P. Cxcl1 and cxcl2 link metastasis and chemoresistance. Nat. Genet. 2012, 44. [Google Scholar] [CrossRef]
- Li, L.; Xu, L.; Yan, J.; Zhen, Z.-J.; Ji, Y.; Liu, C.-Q.; Lau, W.; Zheng, L.; Xu, J. Cxcr2–cxcl1 axis is correlated with neutrophil infiltration and predicts a poor prognosis in hepatocellular carcinoma. J. Exp. Clin. Cancer Res. 2015, 34, 129. [Google Scholar] [CrossRef] [PubMed]
- Shi, H.; Han, X.; Sun, Y.; Shang, C.; Wei, M.; Ba, X.; Zeng, X. Chemokine (c-x-c motif) ligand 1 and cxcl2 produced by tumor promote the generation of monocytic myeloid-derived suppressor cells. Cancer Sci. 2018, 109, 3826–3839. [Google Scholar] [CrossRef] [PubMed]
- Hjortoe, G.M.; Petersen, L.C.; Albrektsen, T.; Sorensen, B.B.; Norby, P.L.; Mandal, S.K.; Pendurthi, U.R.; Rao, V.L. Tissue factor-factor viia–specific up-regulation of il-8 expression in mda-mb-231 cells is mediated by par-2 and results in increased cell migration. Blood 2004, 103, 3029–3037. [Google Scholar] [CrossRef]
- Fan, Z.; Li, Y.; Zhao, Q.; Fan, L.; Tan, B.; Zuo, J.; Hua, K.; Ji, Q. Highly expressed granulocyte colony-stimulating factor (g-csf) and granulocyte colony-stimulating factor receptor (g-csfr) in human gastric cancer leads to poor survival. Med Sci. Monit. Int. Med. J. Exp. Clin. Res. 2018, 24, 1701–1711. [Google Scholar] [CrossRef]
- Kawano, M.; Mabuchi, S.; Matsumoto, Y.; Sasano, T.; Takahashi, R.; Kuroda, H.; Kozasa, K.; Hashimoto, K.; Isobe, A.; Sawada, K.; et al. The significance of g-csf expression and myeloid-derived suppressor cells in the chemoresistance of uterine cervical cancer. Sci. Rep. 2015, 5, 18217. [Google Scholar] [CrossRef]
- Albrengues, J.; Shields, M.A.; Ng, D.; Park, C.; Ambrico, A.; Poindexter, M.E.; Upadhyay, P.; Uyeminami, D.L.; Pommier, A.; Küttner, V.; et al. Neutrophil extracellular traps produced during inflammation awaken dormant cancer cells in mice. Science 2018, 361, eaao4227. [Google Scholar] [CrossRef]
- Wculek, S.K.; Malanchi, I. Neutrophils support lung colonization of metastasis-initiating breast cancer cells. Nature 2015, 528, 413. [Google Scholar] [CrossRef]
- García-Mendoza, M.G.; Inman, D.R.; Ponik, S.M.; Jeffery, J.J.; Sheerar, D.S.; Doorn, R.R.; Keely, P.J. Neutrophils drive accelerated tumor progression in the collagen-dense mammary tumor microenvironment. Breast Cancer Res. 2016, 18, 49. [Google Scholar] [CrossRef] [PubMed]
- Bergin, D.A.; Greene, C.M.; Sterchi, E.E.; Kenna, C.; Geraghty, P.; Belaaouaj, A.; Belaaouaj, A.; Taggart, C.C.; O’Neill, S.J.; McElvaney, N.G. Activation of the epidermal growth factor receptor (egfr) by a novel metalloprotease pathway. J. Biol. Chem. 2008, 283, 31736–31744. [Google Scholar] [CrossRef] [PubMed]
- Darmoul, D.; Gratio, V.; Devaud, H.; Laburthe, M. Protease-activated receptor 2 in colon cancer trypsin-induced mapk phosphorylation and cell proliferation are mediated by epidermal growth factor receptor transactivation. J. Biol. Chem. 2004, 279, 20927–20934. [Google Scholar] [CrossRef] [PubMed]
- De Almeida, V.; dos Guimarães, I.; Almendra, L.R.; Rondon, A.; Tilli, T.M.; de Melo, A.C.; Sternberg, C.; Monteiro, R.Q. Positive crosstalk between egfr and the tf-par2 pathway mediates resistance to cisplatin and poor survival in cervical cancer. Oncotarget 2018, 9, 30594–30609. [Google Scholar] [CrossRef] [PubMed]
- Olayan, A.; Hussaini, H.; Jazieh, A. The roles of epidermal growth factor receptor (egfr) inhibitors in the management of lung cancer. J. Infect. Public Health 2012, 5, 1054–1062. [Google Scholar] [CrossRef] [PubMed]
- Prabhakar, C.N. Epidermal growth factor receptor in non-small cell lung cancer. Transl. Lung Cancer Res. 2015, 4, 110–118. [Google Scholar]
- Sharma, S.V.; Bell, D.W.; Settleman, J.; Haber, D.A. Epidermal growth factor receptor mutations in lung cancer. Nat. Rev. Cancer 2007, 7, 169–181. [Google Scholar] [CrossRef]
- Weng, C.-H.; Chen, L.-Y.; Lin, Y.-C.; Shih, J.-Y.; Lin, Y.-C.; Tseng, R.-Y.; Chiu, A.-C.; Yeh, Y.-H.; Liu, C.; Lin, Y.-T.; et al. Epithelial-mesenchymal transition (emt) beyond egfr mutations per se is a common mechanism for acquired resistance to egfr tki. Oncogene 2019, 38, 455–468. [Google Scholar] [CrossRef]
- Poh, M.-E.; Liam, C.-K.; Rajadurai, P.; Chai, C.-S. Epithelial-to-mesenchymal transition (emt) causing acquired resistance to afatinib in a patient with epidermal growth factor receptor (egfr)-mutant lung adenocarcinoma. J. Thorac. Dis. 2018, 10, E560–E563. [Google Scholar] [CrossRef] [PubMed]
- George, A.J.; Hannan, R.D.; Thomas, W.G. Unravelling the molecular complexity of gpcr-mediated egfr transactivation using functional genomics approaches. FEBS J. 2013, 280, 5258–5268. [Google Scholar] [CrossRef]
- Wang, Z. Transactivation of epidermal growth factor receptor by g protein-coupled receptors: Recent progress, challenges and future research. Int. J. Mol. Sci. 2016, 17, 95. [Google Scholar] [CrossRef] [PubMed]
- Yang, H.; Kim, S.; Cho, S.-H.; Kim, M.-G.; Seo, J.-Y.; Park, J.-S.; Jhon, G.-J.; Han, S.-Y. Purification and structural determination of hematopoietic stem cell-stimulating monoacetyldiglycerides from cervus nippon (deer antler). Chem. Pharm. Bull. 2004, 52, 874–878. [Google Scholar] [CrossRef] [PubMed]
- Jeong, J.; Kim, Y.-J.; Lee, D.; Moon, B.-G.; Sohn, K.-Y.; Yoon, S.; Kim, J. 1-palmitoyl-2-linoleoyl-3-acetyl-rac-glycerol (plag) attenuates gemcitabine-induced neutrophil extravasation. Cell Biosci. 2019, 9, 4. [Google Scholar] [CrossRef] [PubMed]
- Lee, H.-R.; Yoo, N.; Kim, J.; Sohn, K.-Y.; Kim, H.-J.; Kim, M.-H.; Han, M.; Yoon, S.; Kim, J. The therapeutic effect of plag against oral mucositis in hamster and mouse model. Front. Oncol. 2016, 6, 209. [Google Scholar] [CrossRef] [PubMed]
- Kim, Y.-J.; Shin, J.; Shin, S.-H.; Kim, J.; Sohn, K.-Y.; Kim, H.-J.; Kang, J.-K.; Yoon, S.; Kim, J. 1-palmitoyl-2-linoleoyl-3-acetyl-rac-glycerol ameliorates arthritic joints through reducing neutrophil infiltration mediated by il-6/stat3 and mip-2 activation. Oncotarget 2014, 5, 96636–96648. [Google Scholar] [CrossRef] [PubMed]
- Powell, D.R.; Huttenlocher, A. Neutrophils in the tumor microenvironment. Trends Immunol. 2016, 37, 41–52. [Google Scholar] [CrossRef] [PubMed]
- Cools-Lartigue, J.; Spicer, J.; Najmeh, S.; Ferri, L. Neutrophil extracellular traps in cancer progression. Cell. Mol. Life Sci. 2014, 71, 4179–4194. [Google Scholar] [CrossRef]
- Friedl, P.; Wolf, K. Tumour-cell invasion and migration: Diversity and escape mechanisms. Nat. Rev. Cancer 2003, 3, 362–374. [Google Scholar] [CrossRef]
- DeFea, K.A.; Zalevsky, J.; Thoma, M.S.; Déry, O.; Mullins, R.D.; Bunnett, N.W. B-arrestin-dependent endocytosis of proteinase-activated receptor 2 is required for intracellular targeting of activated erk1/2. J. Cell Biol. 2000, 148, 1267–1282. [Google Scholar] [CrossRef]
- Gieseler, F.; Ungefroren, H.; Settmacher, U.; Hollenberg, M.D.; Kaufmann, R. Proteinase-activated receptors (pars)—focus on receptor-receptor-interactions and their physiological and pathophysiological impact. Cell Commun. Signal. 2013, 11, 86. [Google Scholar] [CrossRef]
- Young, S.G.; Davies, B.S.J.; Fong, L.G.; Gin, P.; Weinstein, M.M.; Bensadoun, A.; Beigneux, A.P. Gpihbp1: an endothelial cell molecule important for the lipolytic processing of chylomicrons. Curr. Opin. Lipidol. 2007, 18, 389–396. [Google Scholar] [PubMed]
- Puca, L.; Chastagner, P.; Meas-Yedid, V.; Israël, A.; Brou, C. A-arrestin 1 (arrdc1) and β-arrestins cooperate to mediate notch degradation in mammals. J. Cell Sci. 2013, 126, 4457–4468. [Google Scholar] [CrossRef]
- Hinshaw, D.C.; Shevde, L.A. The tumor microenvironment innately modulates cancer progression. Cancer Res. 2019. [Google Scholar] [CrossRef]
- Yuan, M.; Zhu, H.; Xu, J.; Zheng, Y.; Cao, X.; Liu, Q. Tumor-derived cxcl1 promotes lung cancer growth via recruitment of tumor-associated neutrophils. J. Immunol. Res. 2016, 2016, 6530410. [Google Scholar] [CrossRef] [PubMed]
- Liubomirski, Y.; Lerrer, S.; Meshel, T.; Rubinstein-Achiasaf, L.; Morein, D.; Wiemann, S.; Körner, C.; Ben-Baruch, A. Tumor-stroma-inflammation networks promote pro-metastatic chemokines and aggressiveness characteristics in triple-negative breast cancer. Front. Immunol. 2019, 10, 757. [Google Scholar] [CrossRef] [PubMed]
- Neutrophils help circulating tumor cells metastasize. Cancer Discov. 2019, 9, 458–459.
- Szczerba, B.; Castro-Giner, F.; Vetter, M.; Krol, I.; Gkountela, S.; Landin, J.; Scheidmann, M.C.; Donato, C.; Scherrer, R.; Singer, J.; et al. Neutrophils escort circulating tumour cells to enable cell cycle progression. Nature 2019, 566, 553–557. [Google Scholar] [CrossRef]
- Zhuo, C.; Wu, X.; Li, J.; Hu, D.; Jian, J.; Chen, C.; Zheng, X.; Yang, C. Chemokine (c-x-c motif) ligand 1 (cxcl1) is associated with tumor progression and poor prognosis in patients with colorectal cancer. Biosci. Rep. 2018, 38. [Google Scholar]
- Acharyya, S.; Oskarsson, T.; Vanharanta, S.; Malladi, S.; Kim, J.; Morris, P.G.; Manova-Todorova, K.; Leversha, M.; Hogg, N.; Seshan, V.E.; et al. A cxcl1 paracrine network links cancer chemoresistance and metastasis. Cell 2012, 150, 165–178. [Google Scholar] [CrossRef]
- Pawar, N.R.; Buzza, M.S.; Antalis, T.M. Membrane-anchored serine proteases and protease-activated receptor-2–mediated signaling: Co-conspirators in cancer progression. Cancer Res. 2019, 79, 301–310. [Google Scholar] [CrossRef]
- Lerman, I.; Ma, X.; Seger, C.; Maolake, A.; de la Garcia-Hernandez, M.; Rangel-Moreno, J.; Ackerman, J.; Nastiuk, K.L.; Susiarjo, M.; Hammes, S.R. Epigenetic suppression of serpinb1 promotes inflammation-mediated prostate cancer progression. Mol. Cancer Res. 2019, 17, 845–859. [Google Scholar] [CrossRef] [PubMed]







© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kim, G.T.; Hahn, K.W.; Yoon, S.Y.; Sohn, K.-Y.; Kim, J.W. PLAG Exerts Anti-Metastatic Effects by Interfering with Neutrophil Elastase/PAR2/EGFR Signaling in A549 Lung Cancer Orthotopic Model. Cancers 2020, 12, 560. https://doi.org/10.3390/cancers12030560
Kim GT, Hahn KW, Yoon SY, Sohn K-Y, Kim JW. PLAG Exerts Anti-Metastatic Effects by Interfering with Neutrophil Elastase/PAR2/EGFR Signaling in A549 Lung Cancer Orthotopic Model. Cancers. 2020; 12(3):560. https://doi.org/10.3390/cancers12030560
Chicago/Turabian StyleKim, Guen Tae, Kyu Woong Hahn, Sun Young Yoon, Ki-Young Sohn, and Jae Wha Kim. 2020. "PLAG Exerts Anti-Metastatic Effects by Interfering with Neutrophil Elastase/PAR2/EGFR Signaling in A549 Lung Cancer Orthotopic Model" Cancers 12, no. 3: 560. https://doi.org/10.3390/cancers12030560
APA StyleKim, G. T., Hahn, K. W., Yoon, S. Y., Sohn, K.-Y., & Kim, J. W. (2020). PLAG Exerts Anti-Metastatic Effects by Interfering with Neutrophil Elastase/PAR2/EGFR Signaling in A549 Lung Cancer Orthotopic Model. Cancers, 12(3), 560. https://doi.org/10.3390/cancers12030560