Non-Surgical Cancer Risk Reduction in BRCA1 Mutation Carriers: Disabling the Remote Control



{kind=link}
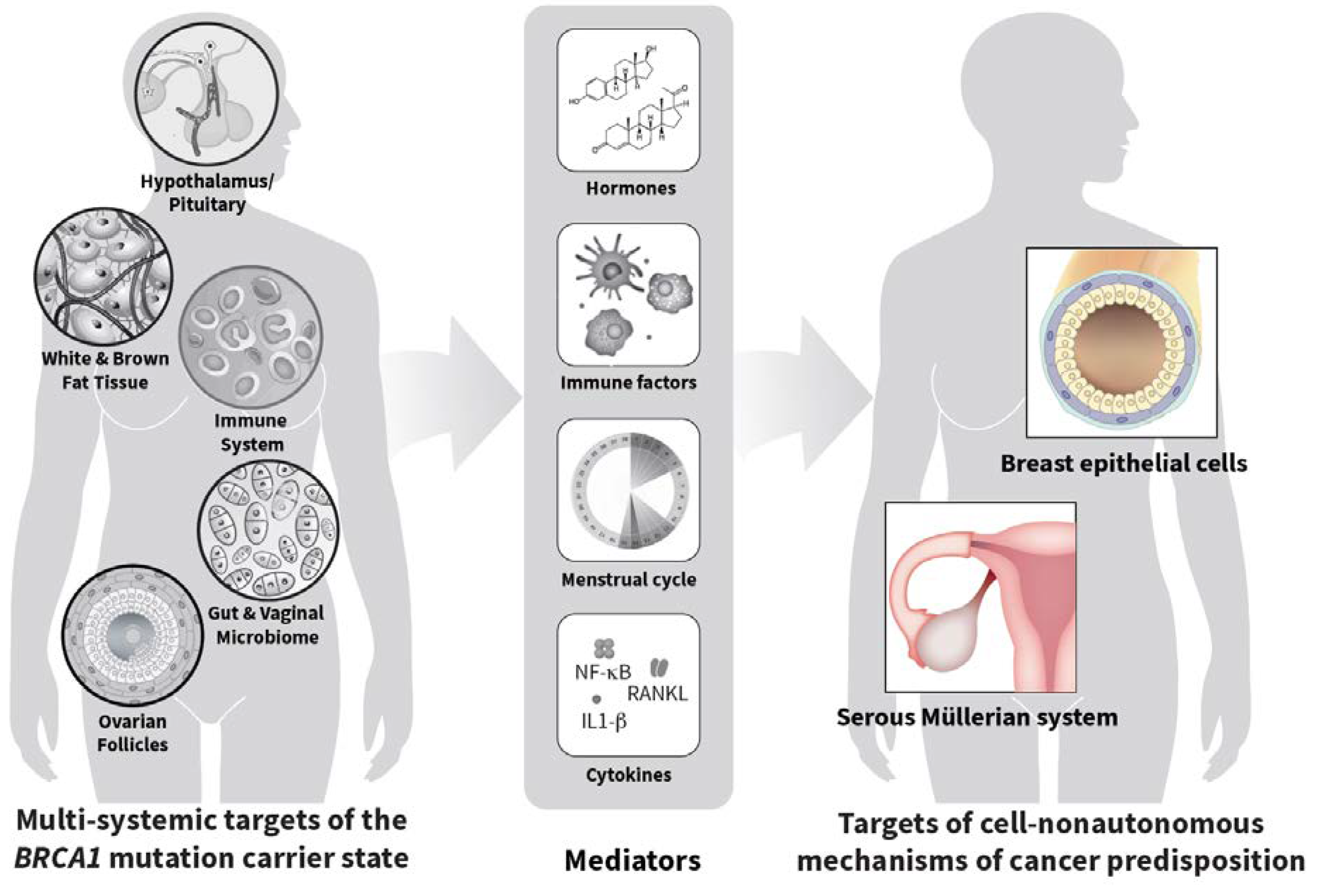
Abstract
1. Introduction
2. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Cuzick, J.; Sestak, I.; Cawthorn, S.; Hamed, H.; Holli, K.; Howell, A.; Forbes, J.F. Tamoxifen for prevention of breast cancer: Extended long-term follow-up of the IBIS-I breast cancer prevention trial. Lancet Oncol. 2015, 16, 67–75. [Google Scholar] [CrossRef]
- Cuzick, J.; Sestak, I.; Forbes, J.F.; Dowsett, M.; Knox, J.; Cawthorn, S.; Saunders, C.; Roche, N.; Mansel, R.E.; Von, M.G.; et al. Anastrozole for prevention of breast cancer in high-risk postmenopausal women (IBIS-II): An international, double-blind, randomised placebo-controlled trial. Lancet 2014, 383, 1041–1048. [Google Scholar] [CrossRef]
- Scott, C.L.; Swisher, E.M.; Kaufmann, S.H. Poly(ADP-ribose) polymerase inhibitors: Recent advances and future development. J. Clin. Oncol. 2015, 33, 1397–1406. [Google Scholar] [CrossRef]
- Austria, T.; Marion, C.; Yu, V.; Widschwendter, M.; Hinton, D.R.; Dubeau, L. Mechanism of cytokinesis failure in ovarian cystadenomas with defective BRCA1 and P53 pathways. Int. J. Cancer 2018, 143, 2932–2942. [Google Scholar] [CrossRef] [PubMed]
- Narod, S.A.; Risch, H.; Moslehi, R.; Dorum, A.; Neuhausen, S.; Olsson, H.; Provencher, D.; Radice, P.; Evans, G.; Bishop, S.; et al. Oral contraceptives and the risk of hereditary ovarian cancer. Hereditary Ovarian Cancer Clinical Study Group. N. Engl. J. Med. 1998, 339, 424–428. [Google Scholar] [CrossRef]
- Antoniou, A.C.; Rookus, M.; Andrieu, N.; Brohet, R.; Chang-Claude, J.; Peock, S.; Cook, M.; Evans, D.G.; Eeles Embrace, R.; Nogues, C.; et al. Reproductive and hormonal factors, and ovarian cancer risk for BRCA1 and BRCA2 mutation carriers: Results from the International BRCA1/2 Carrier Cohort Study. Cancer Epidemiol. Biomark. Prev. 2009, 18, 601–610. [Google Scholar] [CrossRef]
- Pike, M.C.; Henderson, B.E.; Casagrande, J.T.; Rosario, I.; Gray, G.E. Oral contraceptive use and early abortion as risk factors for breast cancer in young women. Br. J. Cancer 1981, 43, 72–76. [Google Scholar] [CrossRef]
- Beaber, E.F.; Malone, K.E.; Tang, M.T.; Barlow, W.E.; Porter, P.L.; Daling, J.R.; Li, C.I. Oral contraceptives and breast cancer risk overall and by molecular subtype among young women. Cancer Epidemiol. Biomark. Prev. 2014, 23, 755–764. [Google Scholar] [CrossRef]
- Chodankar, R.; Kwang, S.; Sangiorgi, F.; Hong, H.; Yen, H.Y.; Deng, C.; Pike, M.C.; Shuler, C.F.; Maxson, R.; Dubeau, L. Cell-nonautonomous induction of ovarian and uterine serous cystadenomas in mice lacking a functional Brca1 in ovarian granulosa cells. Curr. Biol. 2005, 15, 561–565. [Google Scholar] [CrossRef]
- Hong, H.; Yen, H.Y.; Brockmeyer, A.; Liu, Y.; Chodankar, R.; Pike, M.C.; Stanczyk, F.Z.; Maxson, R.; Dubeau, L. Changes in the mouse estrus cycle in response to BRCA1 inactivation suggest a potential link between risk factors for familial and sporadic ovarian cancer. Cancer Res. 2010, 70, 221–228. [Google Scholar] [CrossRef]
- Yen, H.Y.; Gabet, Y.; Liu, Y.; Martin, A.; Wu, N.L.; Pike, M.C.; Frenkel, B.; Maxson, R.; Dubeau, L. Alterations in Brca1 expression in mouse ovarian granulosa cells have short-term and long-term consequences on estrogen-responsive organs. Lab. Investig. 2012, 92, 802–811. [Google Scholar] [CrossRef] [PubMed]
- Widschwendter, M.; Rosenthal, A.N.; Philpott, S.; Rizzuto, I.; Fraser, L.; Hayward, J.; Intermaggio, M.P.; Edlund, C.K.; Ramus, S.J.; Gayther, S.A.; et al. The sex hormone system in carriers of BRCA1/2 mutations: A case-control study. Lancet Oncol. 2013, 14, 1226–1232. [Google Scholar] [CrossRef]
- Widschwendter, M.; Burnell, M.; Fraser, L.; Rosenthal, A.N.; Philpott, S.; Reisel, D.; Dubeau, L.; Cline, M.; Pan, Y.; Yi, P.C.; et al. Osteoprotegerin (OPG), The Endogenous Inhibitor of Receptor Activator of NF-kappaB Ligand (RANKL), is Dysregulated in BRCA Mutation Carriers. eBioMedicine 2015, 2, 1331–1339. [Google Scholar] [CrossRef] [PubMed]
- Schramek, D.; Leibbrandt, A.; Sigl, V.; Kenner, L.; Pospisilik, J.A.; Lee, H.J.; Hanada, R.; Joshi, P.A.; Aliprantis, A.; Glimcher, L.; et al. Osteoclast differentiation factor RANKL controls development of progestin-driven mammary cancer. Nature 2010, 468, 98–102. [Google Scholar] [CrossRef]
- Nolan, E.; Vaillant, F.; Branstetter, D.; Pal, B.; Giner, G.; Whitehead, L.; Lok, S.W.; Mann, G.B.; Kathleen, C.; Soriano, R.; et al. RANK ligand as a potential target for breast cancer prevention in BRCA1-mutation carriers. Nat. Med. 2016, 22, 933–939. [Google Scholar] [CrossRef]
- Phelan, C.M.; Kuchenbaecker, K.B.; Tyrer, J.P.; Kar, S.P.; Lawrenson, K.; Winham, S.J.; Dennis, J.; Pirie, A.; Riggan, M.J.; Chornokur, G.; et al. Pharoah, Identification of 12 new susceptibility loci for different histotypes of epithelial ovarian cancer. Nat. Genet. 2017, 49, 680–691. [Google Scholar] [CrossRef]
- Pike, M.C. Age-related factors in cancers of the breast, ovary, and endometrium. J. Chronic Dis. 1987, 40 (Suppl. 2), 59S–69S. [Google Scholar] [CrossRef]
- Pike, M.C.; Pearce, C.L.; Peters, R.; Cozen, W.; Wan, P.; Wu, A.H. Hormonal factors and the risk of invasive ovarian cancer: A population-based case-control study. Fertil. Steril. 2004, 82, 186–195. [Google Scholar] [CrossRef]
- Whittemore, A.S.; Harris, R.; Itnyre, J. Characteristics relating to ovarian cancer risk: Collaborative analysis of 12 US case-control studies. II. Invasive epithelial ovarian cancers in white women. Collaborative Ovarian Cancer Group. Am. J. Epidemiol. 1992, 136, 1184–1203. [Google Scholar] [CrossRef]
- Nur, U.; El Reda, D.; Hashim, D.; Weiderpass, E. A prospective investigation of oral contraceptive use and breast cancer mortality: Findings from the Swedish women’s lifestyle and health cohort. BMC Cancer 2019, 19, 807. [Google Scholar] [CrossRef]
- Schrijver, L.H.; Olsson, H.; Phillips, K.A.; Terry, M.B.; Goldgar, D.E.; Kast, K.; Engel, C.; Mooij, T.M.; Adlard, J.; Barrowdale, D.; et al. Ibccs, Oral Contraceptive Use and Breast Cancer Risk: Retrospective and Prospective Analyses From a BRCA1 and BRCA2 Mutation Carrier Cohort Study. JNCI Cancer Spectr. 2018, 2, pky023. [Google Scholar] [CrossRef] [PubMed]
- Dubeau, L. The cell of origin of ovarian epithelial tumours. Lancet Oncol. 2008, 9, 1191–1197. [Google Scholar] [CrossRef]
- Lee, Y.; Miron, A.; Drapkin, R.; Nucci, M.R.; Medeiros, F.; Saleemuddin, A.; Garber, J.; Birch, C.; Mou, H.; Gordon, R.W.; et al. A candidate precursor to serous carcinoma that originates in the distal fallopian tube. J. Pathol. 2007, 211, 26–35. [Google Scholar] [CrossRef] [PubMed]
- Yu, V.M.; Marion, C.M.; Austria, T.M.; Yeh, J.; Schonthal, A.H.; Dubeau, L. Role of BRCA1 in controlling mitotic arrest in ovarian cystadenoma cells. Int. J. Cancer 2012, 130, 2495–2504. [Google Scholar] [CrossRef] [PubMed]
- Anjum, S.; Fourkala, E.O.; Zikan, M.; Wong, A.; Gentry-Maharaj, A.; Jones, A.; Hardy, R.; Cibula, D.; Kuh, D.; Jacobs, I.J.; et al. A BRCA1-mutation associated DNA methylation signature in blood cells predicts sporadic breast cancer incidence and survival. Genome Med. 2014, 6, 47. [Google Scholar] [CrossRef]
- Ramadan, S.; Arm, J.; Silcock, J.; Santamaria, G.; Buck, J.; Roy, M.; Leong, K.M.; Lau, P.; Clark, D.; Malycha, P.; et al. Lipid and Metabolite Deregulation in the Breast Tissue of Women Carrying BRCA1 and BRCA2 Genetic Mutations. Radiology 2015, 275, 675–682. [Google Scholar] [CrossRef]
- Smirnova, T.G.; Savochkina Dolgushin, A.Y., II; Nikushkina, K.V.; Samuseva, I.V. Changes in Functional Activity of Neutrophils and Monocytes Isolated from the Peripheral Blood of Women at Different Phases of the Menstrual Cycle. Bull. Exp. Biol. Med. 2018, 166, 222–224. [Google Scholar] [CrossRef]
- Makinde, J.; Jones, C.; Bartolf, A.; Sibeko, S.; Baden, S.; Cosgrove, C.; Shattock, R.J. Localized cyclical variations in immunoproteins in the female genital tract and the implications on the design and assessment of mucosal infection and therapies. Am. J. Reprod. Immunol. 2018, 79, e12801. [Google Scholar] [CrossRef]
- Lorenz, T.K.; Heiman, J.R.; Demas, G.E. Interactions Among Sexual Activity, Menstrual Cycle Phase, and Immune Function in Healthy Women. J. Sex Res. 2018, 55, 1087–1095. [Google Scholar] [CrossRef]
- Nene, N.R.; Reisel, D.; Leimbach, A.; Franchi, D.; Jones, A.; Evans, I.; Knapp, S.; Ryan, A.; Ghazali, S.; Timms, J.F.; et al. Association between the cervicovaginal microbiome, BRCA1 mutation status, and risk of ovarian cancer: A case-control study. Lancet Oncol. 2019, 20, 1171–1182. [Google Scholar] [CrossRef]
- Liu, Y.; Pike, M.C.; Wu, N.; Lin, Y.G.; Mucowski, S.; Punj, V.; Tang, Y.; Yen, H.Y.; Stanczyk, F.Z.; Enbom, E.; et al. Brca1 Mutations Enhance Mouse Reproductive Functions by Increasing Responsiveness to Male-Derived Scent. PLoS ONE 2015, 10, e0139013. [Google Scholar] [CrossRef]
- Widschwendter, M.; Jones, A.; Evans, I.; Reisel, D.; Dillner, J.; Sundstrom, K.; Steyerberg, E.W.; Vergouwe, Y.; Wegwarth, O.; Rebitschek, F.G.; et al. Epigenome-based cancer risk prediction: Rationale, opportunities and challenges. Nat. Rev. Clin. Oncol. 2018, 15, 292–309. [Google Scholar] [CrossRef] [PubMed]
- Bartlett, T.E.; Chindera, K.; McDermott, J.; Breeze, C.E.; Cooke, W.R.; Jones, A.; Reisel, D.; Karegodar, S.T.; Arora, R.; Beck, S.; et al. Epigenetic reprogramming of fallopian tube fimbriae in BRCA mutation carriers defines early ovarian cancer evolution. Nat. Commun. 2016, 7, 11620. [Google Scholar] [CrossRef] [PubMed]
- Trabert, B.; Ness, R.B.; Lo-Ciganic, W.H.; Murphy, M.A.; Goode, E.L.; Poole, E.M.; Brinton, L.A.; Webb, P.M.; Nagle, C.M.; Jordan, S.J.; et al. Aspirin, nonaspirin nonsteroidal anti-inflammatory drug, and acetaminophen use and risk of invasive epithelial ovarian cancer: A pooled analysis in the Ovarian Cancer Association Consortium. J. Natl. Cancer Inst. 2014, 106, djt431. [Google Scholar] [CrossRef] [PubMed]
- Widschwendter, M.; Fiegl, H.; Egle, D.; Mueller-Holzner, E.; Spizzo, G.; Marth, C.; Weisenberger, D.J.; Campan, M.; Young, J.; Jacobs, I.; et al. Epigenetic stem cell signature in cancer. Nat. Genet. 2007, 39, 157–158. [Google Scholar] [CrossRef]
- Noreen, F.; Roosli, M.; Gaj, P.; Pietrzak, J.; Weis, S.; Urfer, P.; Regula, J.; Schar, P.; Truninger, K. Modulation of age- and cancer-associated DNA methylation change in the healthy colon by aspirin and lifestyle. J. Natl. Cancer Inst. 2014, 106. [Google Scholar] [CrossRef]
- Yang, Z.; Wong, A.; Kuh, D.; Paul, D.S.; Rakyan, V.K.; Leslie, R.D.; Zheng, S.C.; Widschwendter, M.; Beck, S.; Teschendorff, A.E. Correlation of an epigenetic mitotic clock with cancer risk. Genome Biol. 2016, 17, 205. [Google Scholar] [CrossRef]

© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Widschwendter, M.; Dubeau, L. Non-Surgical Cancer Risk Reduction in BRCA1 Mutation Carriers: Disabling the Remote Control. Cancers 2020, 12, 547. https://doi.org/10.3390/cancers12030547
Widschwendter M, Dubeau L. Non-Surgical Cancer Risk Reduction in BRCA1 Mutation Carriers: Disabling the Remote Control. Cancers. 2020; 12(3):547. https://doi.org/10.3390/cancers12030547
Chicago/Turabian StyleWidschwendter, Martin, and Louis Dubeau. 2020. "Non-Surgical Cancer Risk Reduction in BRCA1 Mutation Carriers: Disabling the Remote Control" Cancers 12, no. 3: 547. https://doi.org/10.3390/cancers12030547
APA StyleWidschwendter, M., & Dubeau, L. (2020). Non-Surgical Cancer Risk Reduction in BRCA1 Mutation Carriers: Disabling the Remote Control. Cancers, 12(3), 547. https://doi.org/10.3390/cancers12030547