Long Noncoding RNA E2F4as Promotes Progression and Predicts Patient Prognosis in Human Ovarian Cancer


Abstract
Simple Summary
Abstract
1. Introduction
2. Results
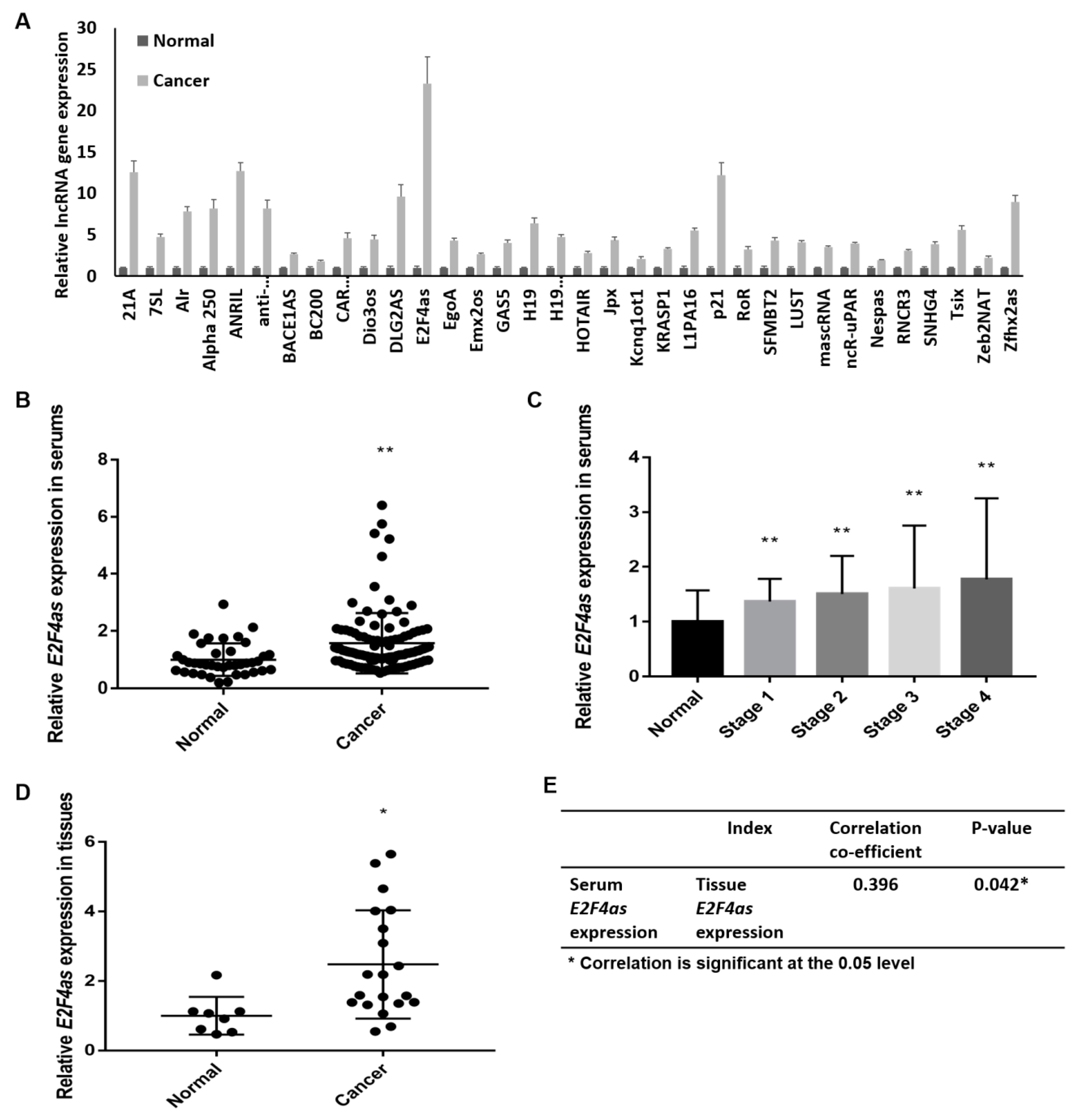
2.1. E2F4as Overexpressed in Ovarian Cancer Patient Serum
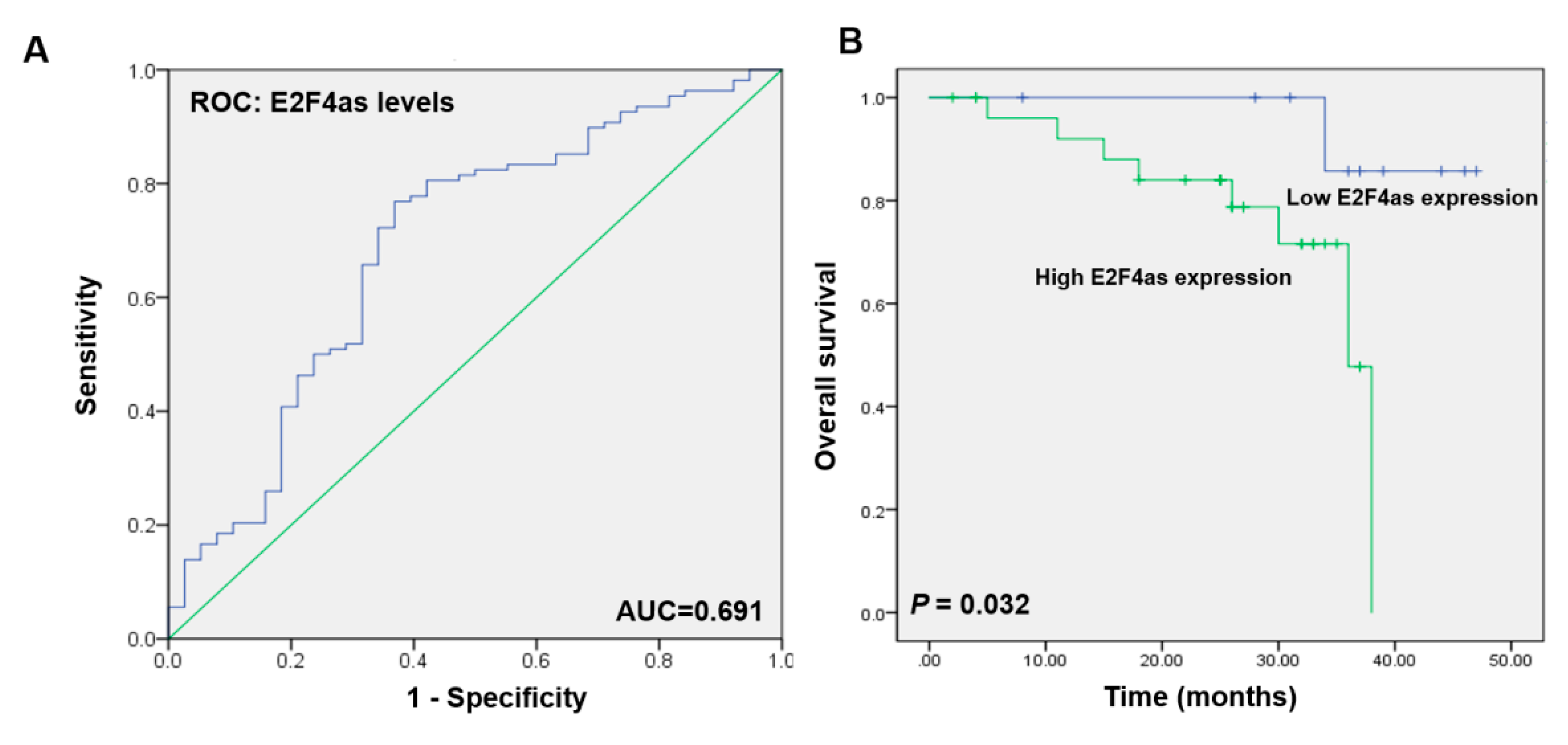
2.2. E2F4as Levels Elevated in Patient Serum with Ovarian Cancer Having Poor Prognoses
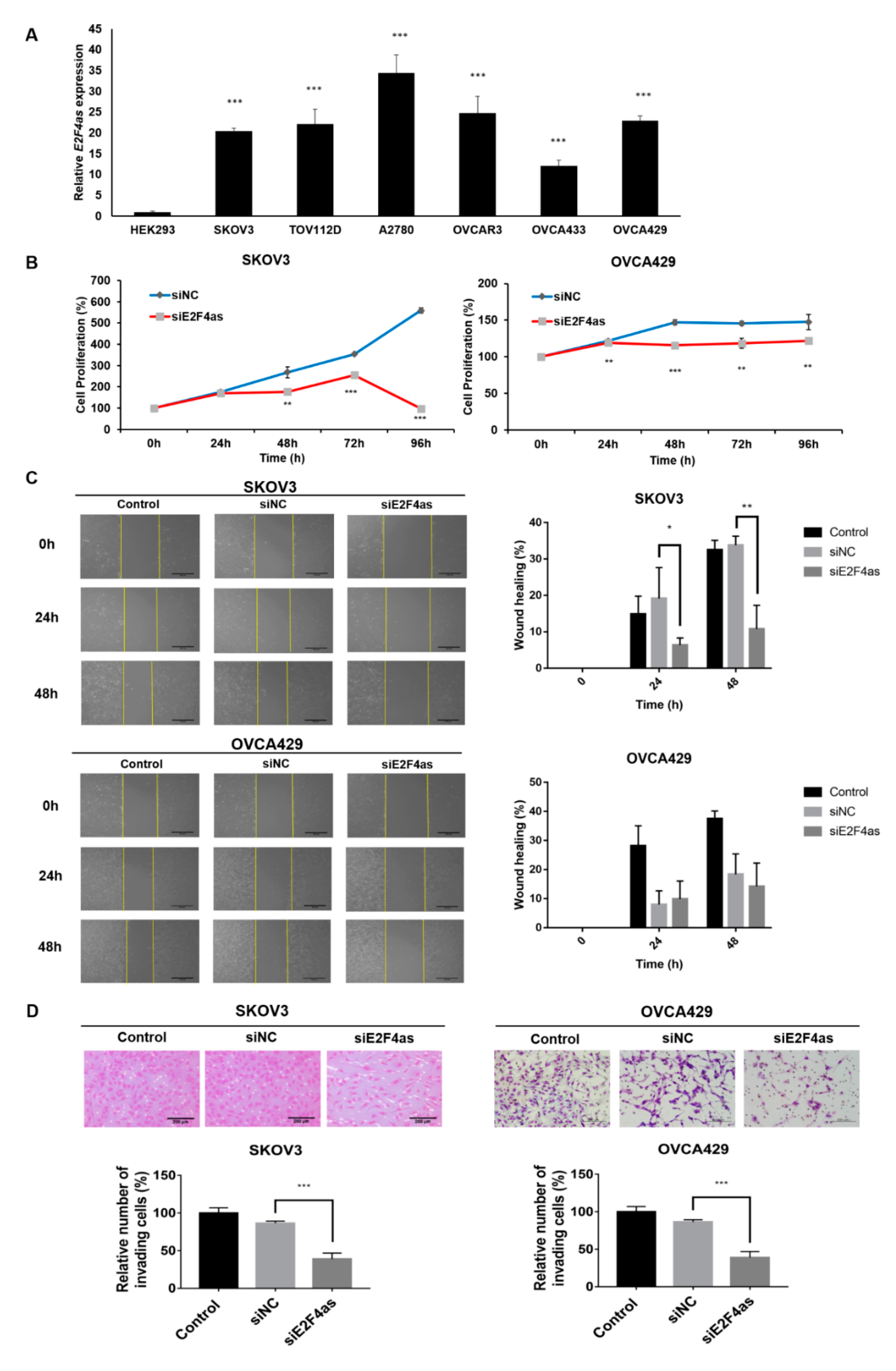
2.3. Inhibition of E2F4as Could Suppress Proliferation and Metastasis of Ovarian Cancer Cells
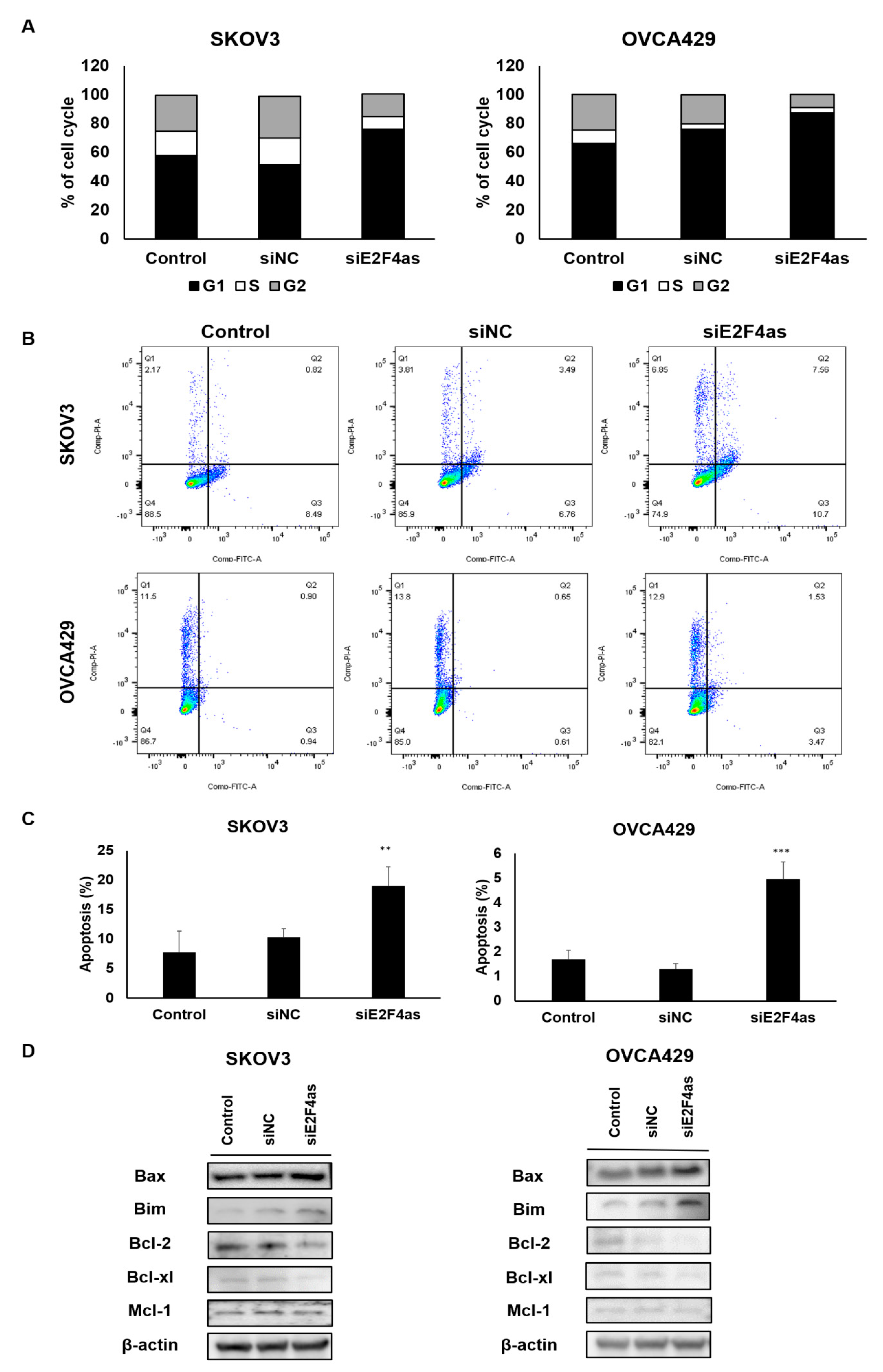
2.4. Downregulated E2F4as Increases Cell Cycle Arrest and Apoptosis in Ovarian Cancer Cells
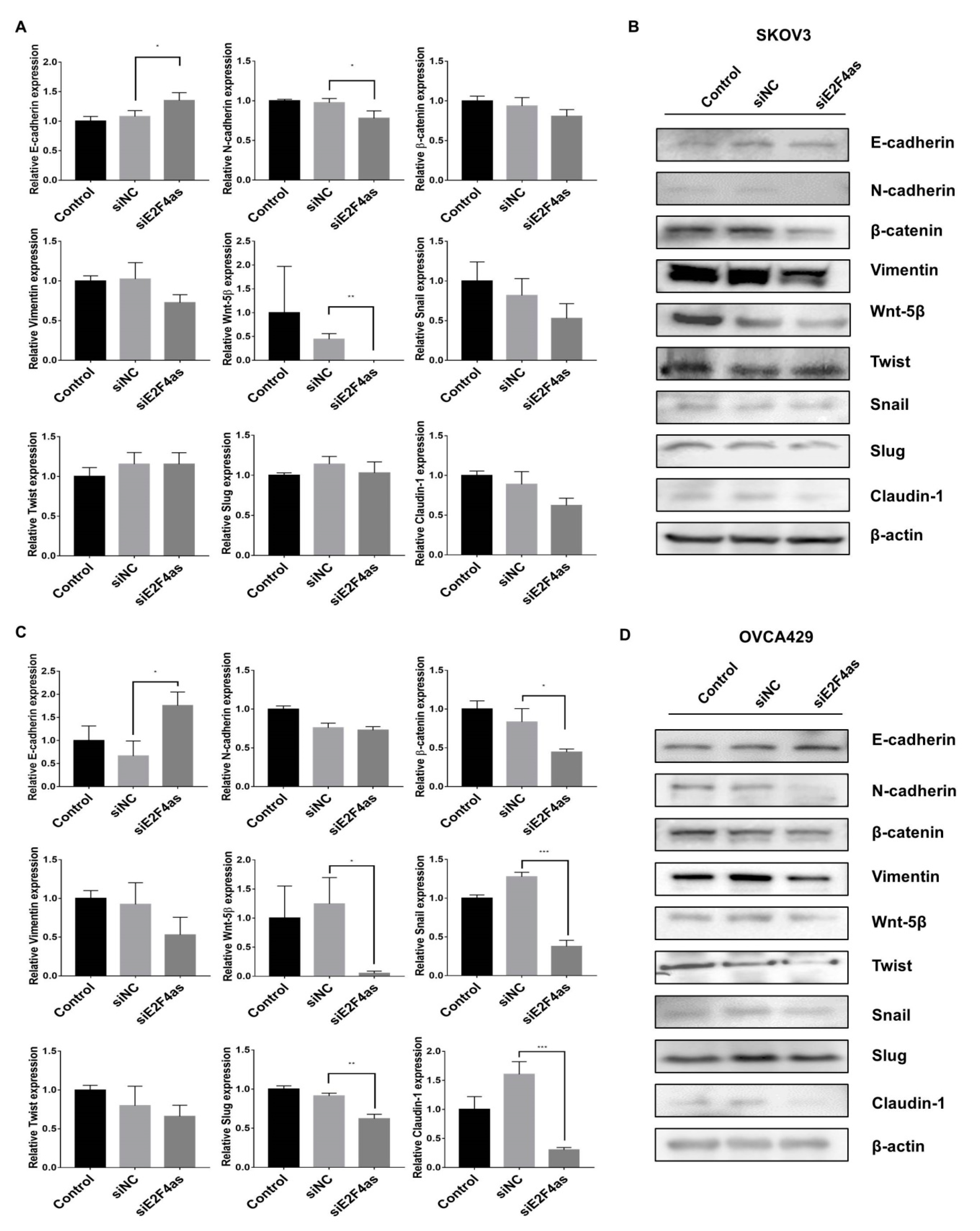
2.5. Effects of Knockdown E2F4as on EMT-Associated Genes in Ovarian Cancer Cells
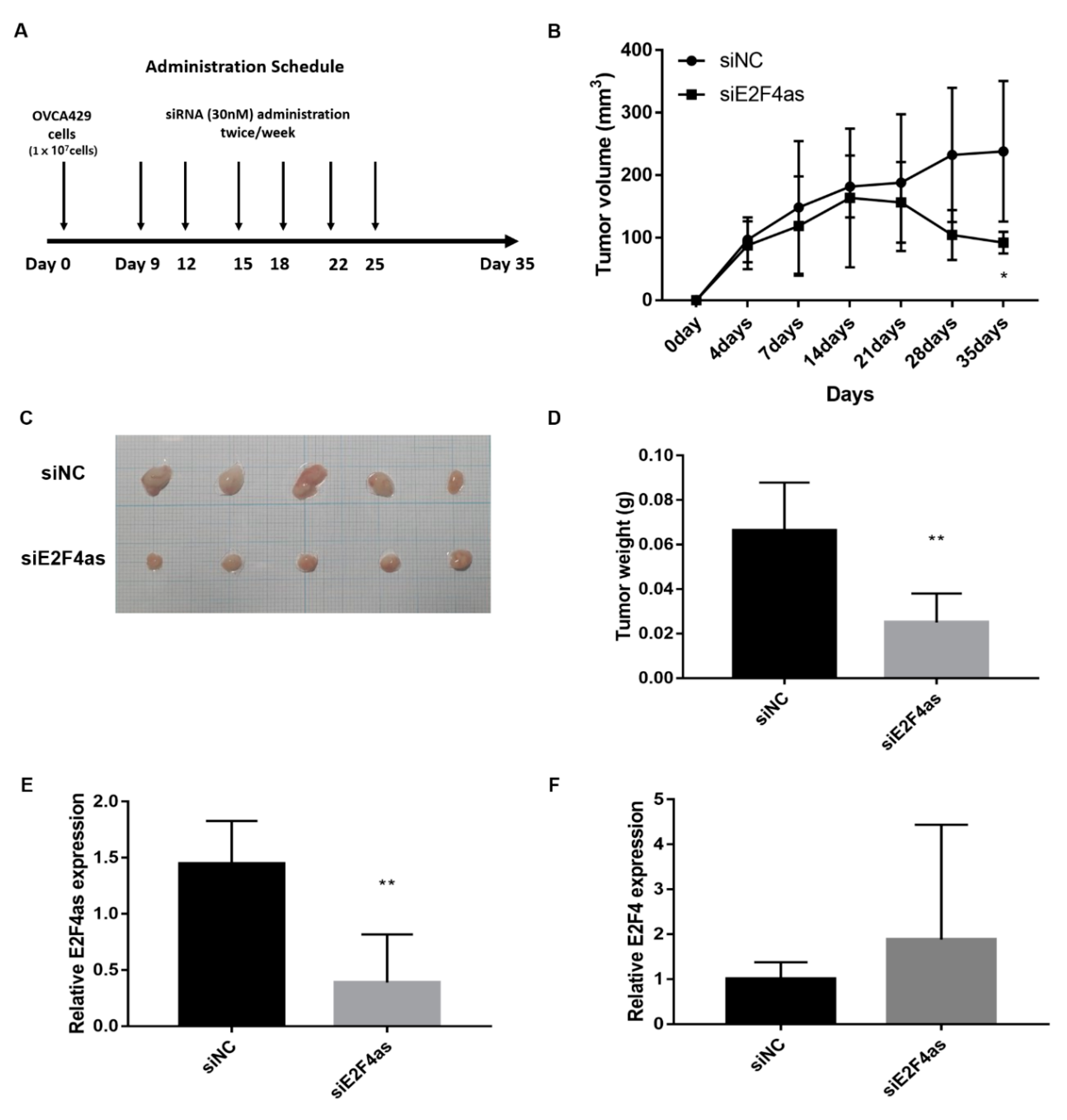
2.6. Silencing of lncE2F4as Inhibits OVCA429 Tumour Growth In Vivo
3. Discussion
4. Materials and Methods
4.1. Patient Tissue and Serum Samples
4.2. Ethics Approval and Consent to Participate
4.3. Serum RNA Extraction and LncRNA Expression Profiling
4.4. Quantitative Real-Time PCR Analysis (qRT-PCR)
4.5. Cell Lines
4.6. Small Interfering RNA (siRNA) Transfection
4.7. Cell Proliferation Assay
4.8. Wound Healing Assay and Matrigel Invasion Assay
4.9. Western Blot Analysis
4.10. Flow Cytometry Apoptosis Analysis
4.11. Flow Cytometry Cell Cycle Analysis
4.12. Xenograft in Mice
4.13. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
| E2F4as | E2F4 antisense |
| EMT | epithelial-mesenchymal transition |
| FIGO | The International Federation of Gynecology and Obstetrics |
| KCLB | Korean Cell Line Bank |
| siRNA | small interfering RNA |
| siE2F4as | E2F4as-targeting siRNA |
| siNC | negative control siRNA |
| qRT-PCR | quantitative real-time PCR |
| CCK-8 | Cell Counting Kit-8 |
| RIPA | radioimmunoprecipitation assay |
| HR | hazard ratio |
| NRF | National Research Foundation of Korea |
References
- Zhan, L.; Li, J.; Wei, B. Long noncoding RNAs in ovarian cancer. J. Exp. Clin. Cancer Res. 2018, 37, 120. [Google Scholar] [CrossRef] [PubMed]
- Agarwal, R.; Kaye, S.B. Ovarian cancer: Strategies for overcoming resistance to chemotherapy. Nat. Rev. Cancer 2003, 3, 502–516. [Google Scholar] [CrossRef] [PubMed]
- Siegel, R.L.; Miller, K.D.; Jemal, A. Cancer statistics, 2018. CA Cancer J. Clin. 2018, 68, 7–30. [Google Scholar] [CrossRef] [PubMed]
- Chen, T.; Xie, W.; Xie, L.; Sun, Y.; Zhang, Y.; Shen, Z.; Sha, N.; Xu, H.; Wu, Z.; Hailong, H.; et al. Expression of long noncoding RNA lncRNA-n336928 is correlated with tumor stage and grade and overall survival in bladder cancer. Biochem. Biophys. Res. Commun. 2015, 468, 666–670. [Google Scholar] [CrossRef]
- Yang, G.; Lu, X.; Yuan, L. LncRNA: A link between RNA and cancer. Biochim. Biophys. Acta 2014, 1839, 1097–1109. [Google Scholar] [CrossRef]
- Wilusz, J.E.; Sunwoo, H.; Spector, D.L. Long noncoding RNAs: Functional surprises from the RNA world. Genes Dev. 2009, 23, 1494–1504. [Google Scholar] [CrossRef]
- Yim, G.W.; Kim, H.J.; Kim, L.K.; Kim, S.W.; Kim, S.; Nam, E.J.; Kim, Y.T. Long non-coding RNA HOXA11 antisense promotes cell proliferation and invasion and predicts patient prognosis in serous ovarian cancer. Cancer Res. Treat. 2017, 49, 656–668. [Google Scholar] [CrossRef]
- Zielke, N.; Kim, K.J.; Tran, V.; Shibutani, S.T.; Bravo, M.J.; Nagarajan, S.; van Straaten, M.; Woods, B.; von Dassow, G.; Rottig, C.; et al. Control of Drosophila endocycles by E2F and CRL4 CDT2. Nature 2011, 480, 123–127. [Google Scholar] [CrossRef]
- Lammens, T.; Li, J.; Leone, G.; de Veylder, L. Atypical E2Fs: New players in the E2F transcription factor family. Trends Cell Biol. 2009, 19, 111–118. [Google Scholar] [CrossRef]
- Dyson, N. The regulation of E2F by pRB-family proteins. Genes Dev. 1998, 12, 2245–2262. [Google Scholar] [CrossRef]
- Tsantoulis, P.K.; Gorgoulis, V.G. Involvement of E2F transcription factor family in cancer. Eur. J. Cancer 2005, 41, 2403–2414. [Google Scholar] [CrossRef] [PubMed]
- DeGregori, J.; Johnson, D.G. Distinct and overlapping roles for E2F family members in transcription, proliferation and apoptosis. Curr. Mol. Med. 2006, 6, 739–748. [Google Scholar] [PubMed]
- Dimova, D.K.; Dyson, N.J. The E2F transcriptional network: Old acquaintances with new faces. Oncogene 2005, 24, 2810–2826. [Google Scholar] [CrossRef]
- Gaubatz, S.; Lindeman, G.J.; Ishida, S.; Jakoi, L.; Nevins, J.R.; Livingstone, D.M.; Rempel, R.E. E2F4 and E2F5 play an essential role in pocket protein-mediated G1 control. Mol. Cell. 2000, 6, 729–735. [Google Scholar] [CrossRef]
- Yochum, G.S.; Cleland, R.; McWeeney, S.; Goodman, R.H. An antisense transcript induced by Wnt/β-catenin signaling decreases E2F4. J. Biol. Chem. 2007, 282, 871–878. [Google Scholar] [CrossRef] [PubMed]
- Yang, A.D.; Fan, F.; Camp, E.R.; van Buren, G.; Liu, W.; Somcio, R.; Gray, M.J.; Cheng, H.; Hoff, P.M.; Ellis, L.M. Chronic oxaliplatin resistance induces epithelial-to-mesenchymal transition in colorectal cancer cell lines. Clin. Cancer Res. 2006, 12, 4147–4153. [Google Scholar] [CrossRef] [PubMed]
- Lee, M.; Kim, H.J.; Kim, S.W.; Park, S.-A.; Chun, K.-H.; Cho, N.H.; Song, Y.S.; Kim, Y.T. The long non-coding RNA HOTAIR increases tumour growth and invasion in cervical cancer by targeting the Notch pathway. Oncotarget 2016, 7, 44558–44571. [Google Scholar] [CrossRef]
- Kim, H.J.; Lee, D.W.; Yim, G.W.; Nam, E.J.; Kim, S.; Kim, S.W.; Kim, Y.T. Long non-coding RNA HOTAIR is associated with human cervical cancer progression. Int. J. Oncol 2015, 46, 521–530. [Google Scholar] [CrossRef]
- Zhao, M.; Qiu, Y.; Yang, B.; Sun, L.; Hei, K.; Du, X.; Li, Y. Long non-coding RNAs involved in gynecological cancer. Int. J. Gynecol. Cancer 2014, 24, 1140–1145. [Google Scholar] [CrossRef]
- Fan, C.; Wang, J.; Tang, Y.; Zhang, S.; Xiong, F.; Guo, C.; Zhou, Y.; Li, Z.; Li, X.; Li, Y.; et al. Upregulation of long non-coding RNA LOC284454 may serve as a new serum biomarker for head and neck cancers. Oncol. Cancer Biol. 2020, 20, 1–9. [Google Scholar] [CrossRef]
- Freedman, J.A.; Chang, J.T.; Jakoi, L.; Nevins, J.R. A combinatorial mechanism for determining the specificity of E2F activation and repression. Oncogene 2009, 28, 2873–2881. [Google Scholar] [CrossRef] [PubMed]
- Baum, B.; Settleman, J.; Quinlan, M.P. Transitions between epithelial and mesenchymal states in development and disease. Seminars in cell and developmental biology. Semin. Cell Dev. Biol. 2008, 19, 294–308. [Google Scholar] [CrossRef] [PubMed]
- Thiery, J.P.; Sleeman, J.P. Complex networks orchestrate epithelial–mesenchymal transitions. Nat. Rev. Mol. Cell Biol. 2006, 7, 131. [Google Scholar] [CrossRef] [PubMed]
- Wang, Q.; López-Ozuna, V.M.; Baloch, T.; Bithras, J.; Amin, O.; Kessous, R.; Kogan, L.; Laskov, I.; Yasmeen, A. Biguanides in combination with olaparib limits tumorigenesis of drug-resistant ovarian cancer cells through inhibition of snail. Cancer Med. 2020, 9, 1307–1320. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Z.; Zhou, W.; Han, Y.; Peng, F.; Wang, R.; Yu, R.; Wang, C.; Liang, H.; Guo, Z.; Gu, Y. EMT-regulome: A database for EMT-related regulatory interactions, motifs and network. Cell Death Dis. 2017, 8, e2872. [Google Scholar] [CrossRef] [PubMed]
- Li, C.; Wan, L.; Liu, Z.; Xu, G.; Wang, S.; Su, Z.; Zhang, Y.; Zhang, Z.; Liu, X.; Lei, Z.; et al. Long non-coding RNA XIST promotes TGF-β-induced epithelial-mesenchymal transition by regulating miR-367/141-ZEB2 axis in non-small-cell lung cancer. Cancer Lett. 2018, 418, 185–195. [Google Scholar] [CrossRef]
- Coticchia, C.M.; Yang, J.; Moses, M.A. Ovarian cancer biomarkers: Current options and future promise. J. Natl. Compr. Cancer Netw. 2008, 6, 795–802. [Google Scholar] [CrossRef]
- Nowak, M.; Janas, Ł.; Stachowiak, G.; Stetkiewicz, T.; Wilczyński, J.R. Current clinical application of serum biomarkers to detect ovarian cancer. Prz. Menopauzalny Menopause Rev. 2015, 14, 254. [Google Scholar] [CrossRef]
- Beaufort, C.M.; Helmijr, J.C.; Piskorz, A.M.; Hoogstraat, M.; Ruigrok-Ritstier, K.; Besselink, N.; Murtaza, M.; van, I.W.F.; Heine, A.A.; Smid, M.; et al. Ovarian cancer cell line panel (OCCP): Clinical importance of in vitro morphological subtypes. PLoS ONE 2014, 9, e103988. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| E2F4as Expression | ||||
|---|---|---|---|---|
| Variables | n | Low | High | p-Value a |
| Age (years, mean ± SD) | 108 | 51.06 ± 9.5 | 53.75 ± 12.8 | 0.084 |
| Grade | 0.235 | |||
| 1 | 10 | 5 | 5 | |
| 2 | 28 | 8 | 20 | |
| 3 | 70 | 17 | 53 | |
| Stage | 0.006 * | |||
| I | 26 | 14 | 12 | |
| II | 21 | 5 | 16 | |
| III | 31 | 7 | 24 | |
| IV | 30 | 4 | 26 | |
| Lymph-node metastasis | 0.004 * | |||
| Yes | 33 | 3 | 30 | |
| No | 75 | 27 | 48 | |
| BMI (Body mass index) | 0.393 | |||
| 25> | 75 | 19 | 56 | |
| 25< | 33 | 11 | 22 | |
| Tumour size | 0.201 | |||
| 2_5.9 | 30 | 11 | 19 | |
| ≥6 | 78 | 19 | 59 | |
| Recurrence | 0.676 | |||
| Yes | 32 | 8 | 24 | |
| No | 76 | 22 | 54 | |
| Menopause | 0.401 | |||
| Yes | 68 | 17 | 51 | |
| No | 40 | 13 | 27 | |
| Cell type | 0.168 | |||
| Serous Carcinoma | 66 | 14 | 52 | |
| Clear cell carcinoma | 7 | 3 | 4 | |
| Endometrioid | 14 | 6 | 8 | |
| Mucinous | 16 | 4 | 12 | |
| Other | 5 | 2 | 3 | |
| Univariate Analysis | Multivariate Analysis | |||
|---|---|---|---|---|
| Factor | Hazard Ratio (95% CI) | p-Value | Hazard Ratio (95% CI) | p-Value |
| E2F4as expression | 0.575 (0.187–1.766) | 0.327 | 0.218 (0.048–0.996) | 0.049 * |
| Age (years) | 1.044 (0.994–1.095) | 0.084 | 0.996 (0.925–10.73) | 0.924 |
| Grade | 1.961 (0.653–5.886) | 0.216 | 1.310 (0.343–5.002) | 0.693 |
| Stage | 1.676 (0.969–2.899) | 0.055 | 2.130 (1.005–4.513) | 0.048 * |
| Lymph-node metastasis | 4.527 (1.458–14.056) | 0.004 ** | 4.324 (1.392–13.425) | 0.011 * |
| BMI | 2.025 (0.675–6.081) | 0.199 | 1.484 (0.444–4.964) | 0.521 |
| Tumour size | 0.801 (0.246–2.612) | 0.712 | 2.025 (0.471–8.705) | 0.343 |
| Recurrence | 2.379 (0.798–7.092) | 0.109 | 0.863 (0.232–3.213) | 0.826 |
| Menopause | 7.368 (0.957–56.723) | 0.024 * | 6.985 (0.907–53.809) | 0.062 |
| Cell type | 0.848 (0.531–1.355) | 0.487 | 1.490 (0.444–4.964) | 0.146 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Park, S.-A.; Kim, L.K.; Kim, Y.T.; Heo, T.-H.; Kim, H.J. Long Noncoding RNA E2F4as Promotes Progression and Predicts Patient Prognosis in Human Ovarian Cancer. Cancers 2020, 12, 3626. https://doi.org/10.3390/cancers12123626
Park S-A, Kim LK, Kim YT, Heo T-H, Kim HJ. Long Noncoding RNA E2F4as Promotes Progression and Predicts Patient Prognosis in Human Ovarian Cancer. Cancers. 2020; 12(12):3626. https://doi.org/10.3390/cancers12123626
Chicago/Turabian StylePark, Sun-Ae, Lee Kyung Kim, Young Tae Kim, Tae-Hwe Heo, and Hee Jung Kim. 2020. "Long Noncoding RNA E2F4as Promotes Progression and Predicts Patient Prognosis in Human Ovarian Cancer" Cancers 12, no. 12: 3626. https://doi.org/10.3390/cancers12123626
APA StylePark, S.-A., Kim, L. K., Kim, Y. T., Heo, T.-H., & Kim, H. J. (2020). Long Noncoding RNA E2F4as Promotes Progression and Predicts Patient Prognosis in Human Ovarian Cancer. Cancers, 12(12), 3626. https://doi.org/10.3390/cancers12123626