Tumor-Associated Macrophages Induce Migration of Renal Cell Carcinoma Cells via Activation of the CCL20-CCR6 Axis

 , ,
, ,  , , , , , and
, , , , , and
Abstract
1. Introduction
2. Results
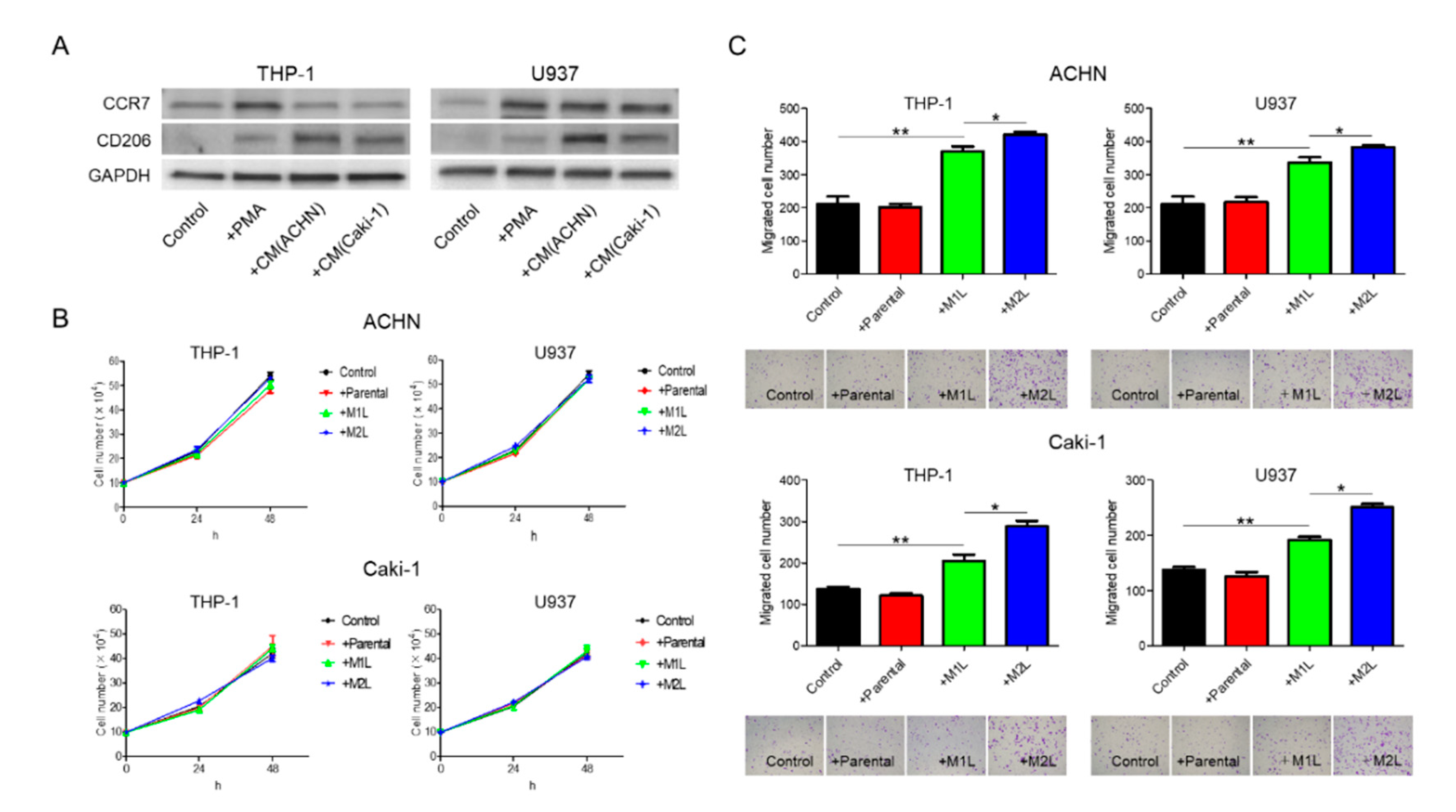
2.1. RCC Cells Could Induce M2-Type-Like (M2L) Macrophages from M1-Type-Like (M1L) Macrophages
2.2. Macrophages Increased RCC Cell Migration
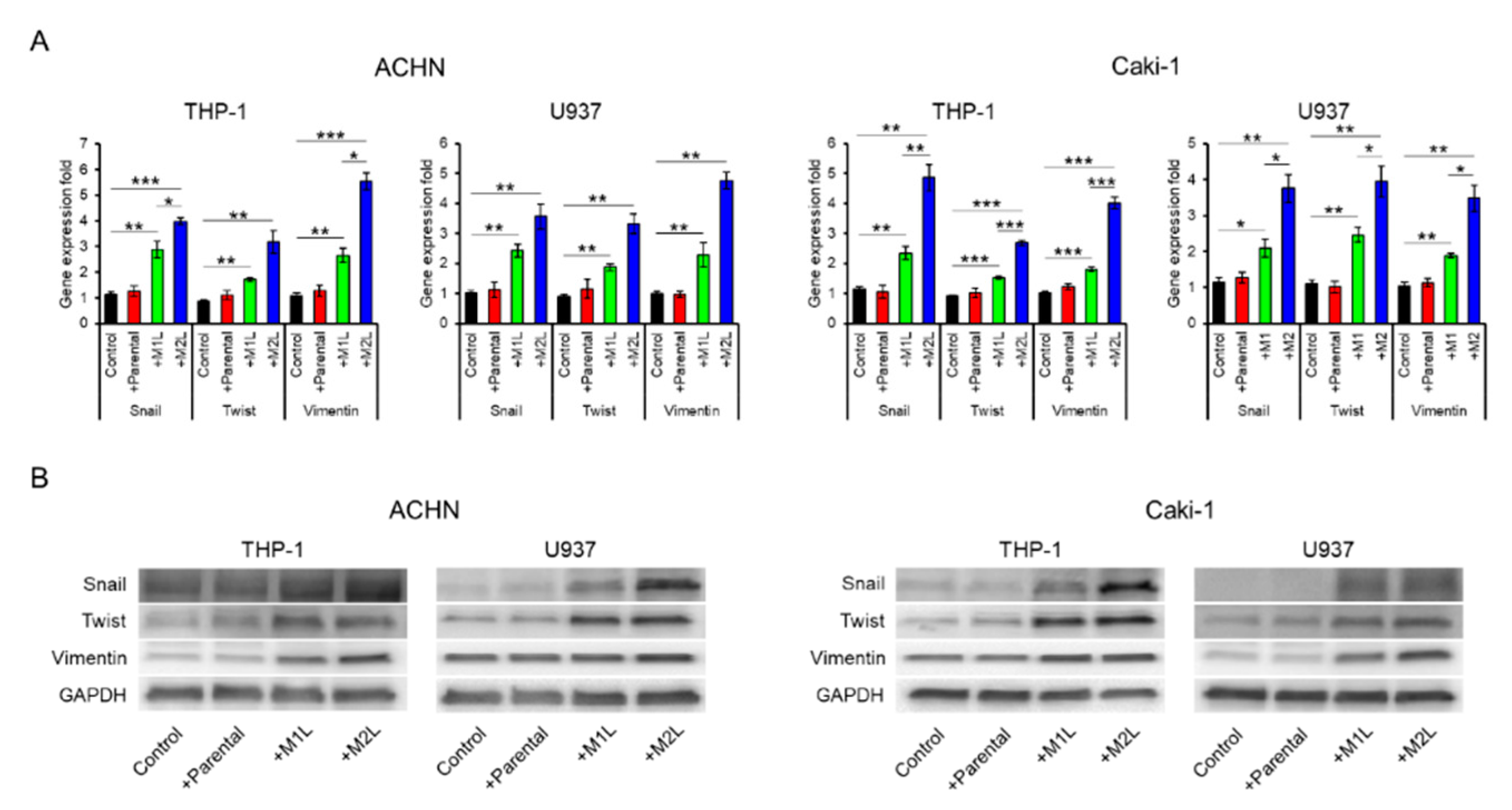
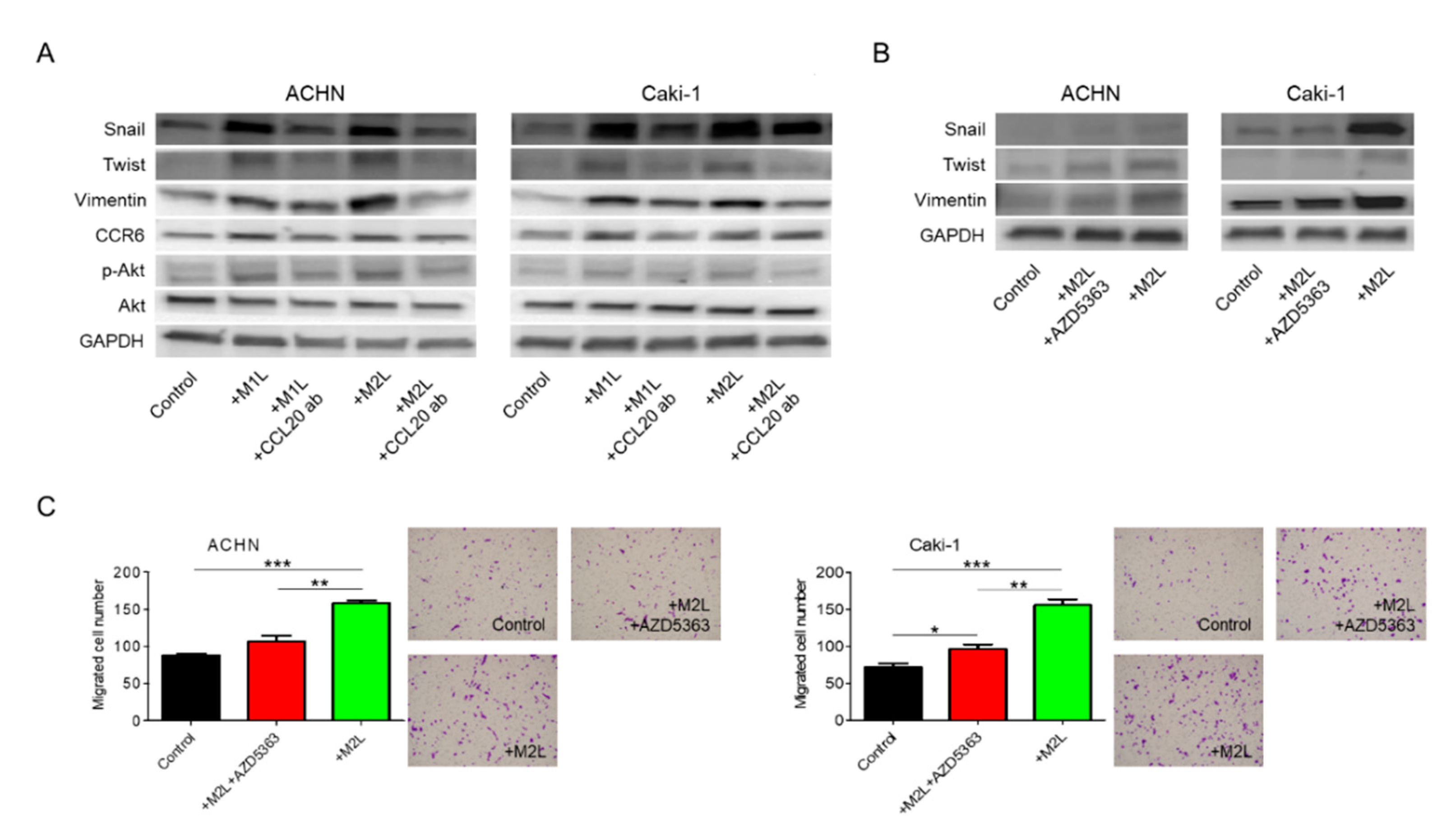
2.3. Macrophages Enhanced the EMT of RCC Cells
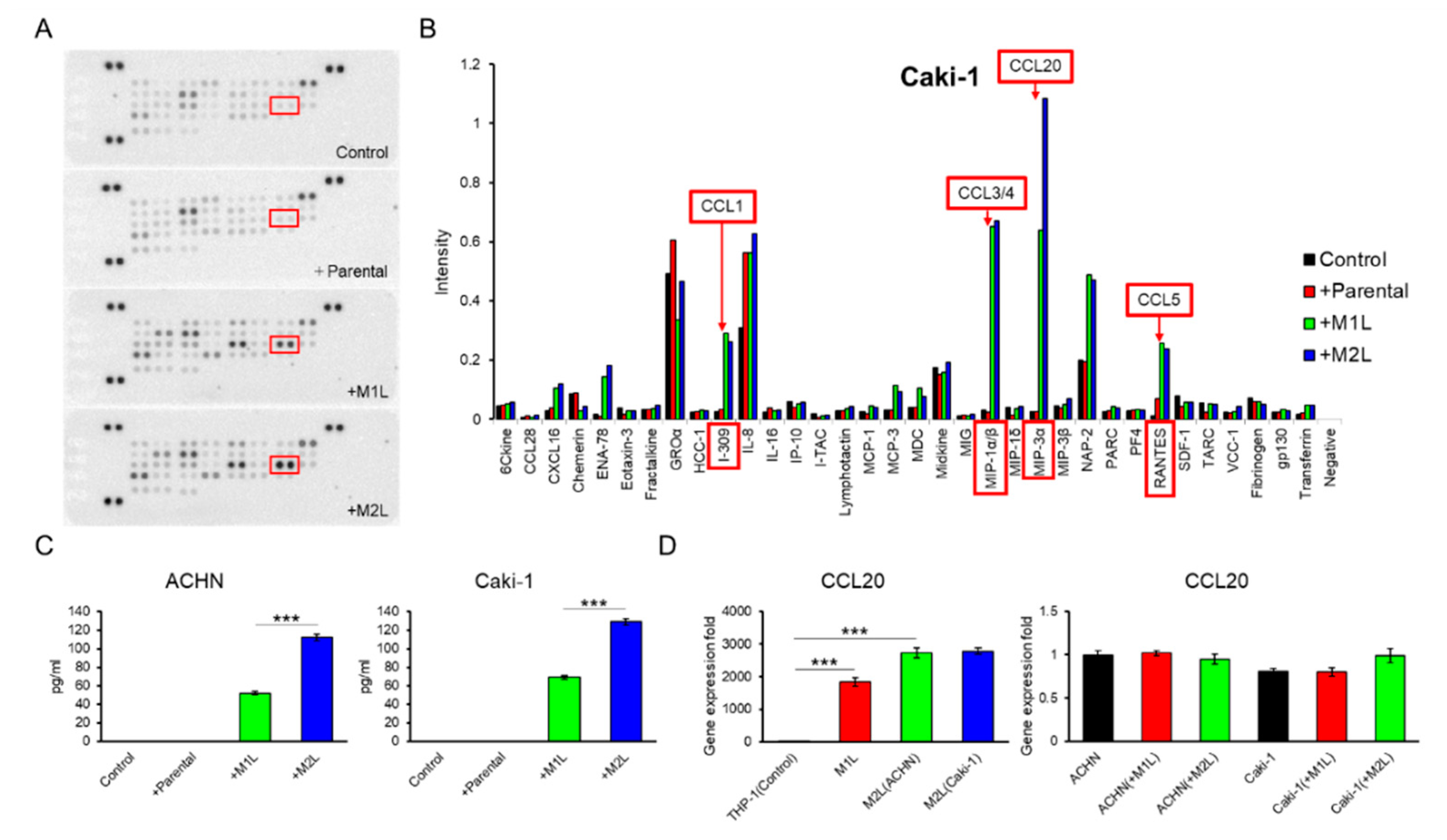
2.4. Macrophage-Like Cells Secreted CCL20
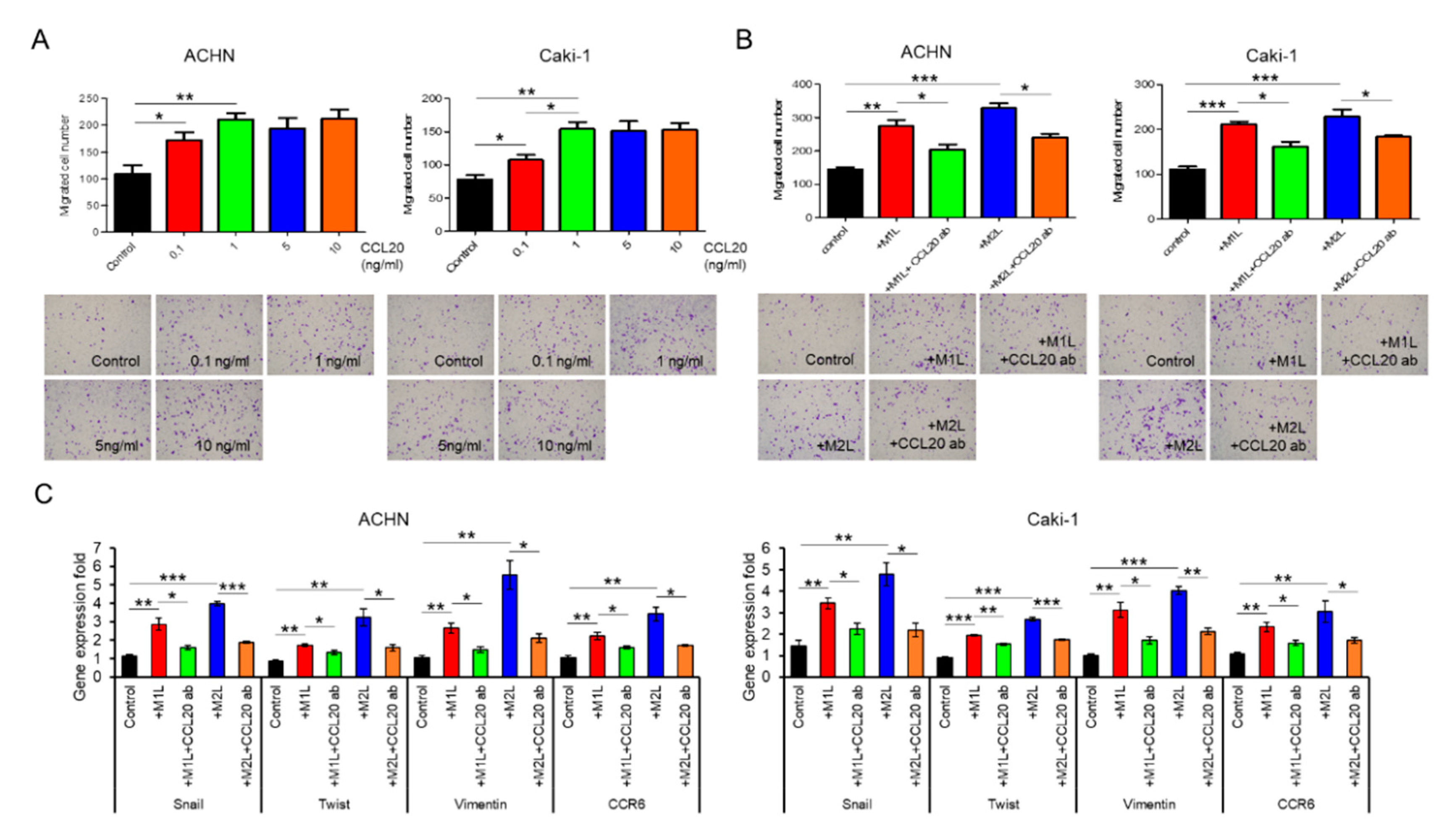
2.5. CCL20 Promoted ACHN and Caki-1 Cell Migration
2.6. CCL20-CCR6 Axis Activated Akt
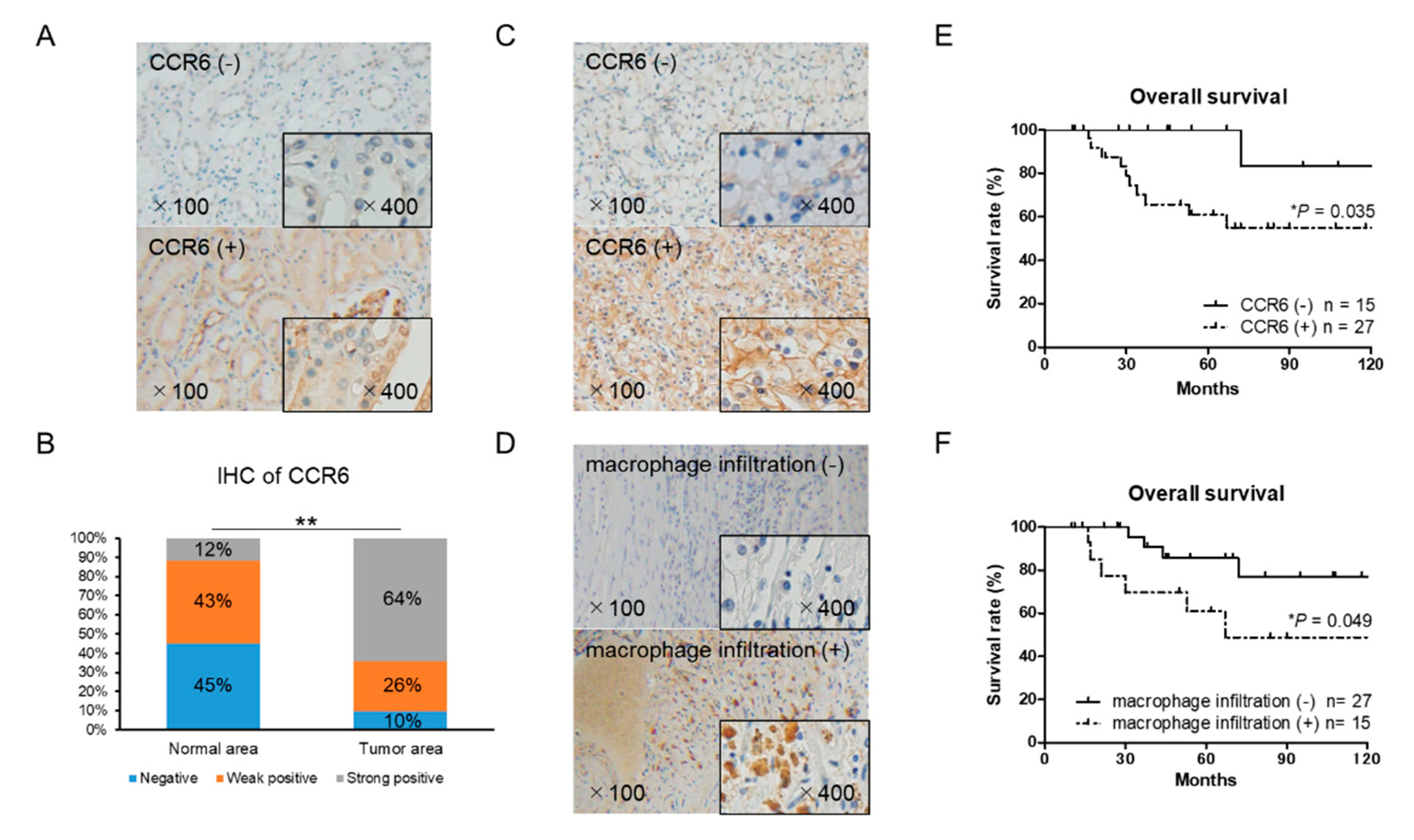
2.7. CCR6 Expression is a Poor Prognostic Factor in RCC
3. Discussion
4. Materials and Methods
4.1. Reagents and Antibodies
4.2. Cell Culture
4.3. Conditioned Medium (CM)
4.4. Preparation of Macrophage-Like Cells
4.5. Co-Culture Assay
4.6. Proliferation Assays
4.7. Cell Migration Assays
4.8. RNA Extraction and RT-qPCR Analysis
4.9. Western Blot Analysis
4.10. Chemokine Array
4.11. Enzyme-Linked Immunosorbent Assay (ELISA)
4.12. Immunohistochemistry
4.13. Statistical Analyses
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Siegel, R.L.; Miller, K.D.; Jemal, A. Cancer statistics, 2019. CA Cancer J. Clin. 2019, 69, 7–34. [Google Scholar] [CrossRef] [PubMed]
- Molina, A.M.; Motzer, R.J. Current algorithms and prognostic factors in the treatment of metastatic renal cell carcinoma. Clin. Genitourin. Cancer 2008, 6 (Suppl. 1), S7–S13. [Google Scholar] [CrossRef] [PubMed]
- Harada, K.; Nozawa, M.; Uemura, M.; Tatsugami, K.; Osawa, T.; Yamana, K.; Kimura, G.; Fujisawa, M.; Nonomura, N.; Eto, M.; et al. Treatment patterns and outcomes in patients with unresectable or metastatic renal cell carcinoma in Japan. Int. J. Urol. 2019, 26, 202–210. [Google Scholar] [CrossRef] [PubMed]
- Prasad, S.R.; Humphrey, P.A.; Catena, J.R.; Narra, V.R.; Srigley, J.R.; Cortez, A.D.; Dalrymple, N.C.; Chintapalli, K.N. Common and uncommon histologic subtypes of renal cell carcinoma: Imaging spectrum with pathologic correlation. Radiographics 2006, 26, 1795–1806, disscussion 1806–1810. [Google Scholar] [CrossRef] [PubMed]
- Takezawa, Y.; Izumi, K.; Shimura, Y.; Aerken, M.; Natsagdorji, A.; Iijima, M.; Shigehara, K.; Nohara, T.; Narimoto, K.; Kadono, Y.; et al. Treatment Outcome of Low-dose Interleukin-2 Therapy in Patients with Metastatic Renal Cell Carcinoma. Anticancer Res. 2016, 36, 4961–4964. [Google Scholar] [CrossRef]
- Kovaleva, O.V.; Samoilova, D.V.; Shitova, M.S.; Gratchev, A. Tumor Associated Macrophages in Kidney Cancer. Anal. Cell. Pathol. 2016, 2016, 9307549. [Google Scholar] [CrossRef]
- Cannarile, M.A.; Ries, C.H.; Hoves, S.; Ruttinger, D. Targeting tumor-associated macrophages in cancer therapy and understanding their complexity. Oncoimmunology 2014, 3, e955356. [Google Scholar] [CrossRef]
- Izumi, K.; Fang, L.Y.; Mizokami, A.; Namiki, M.; Li, L.; Lin, W.J.; Chang, C. Targeting the androgen receptor with siRNA promotes prostate cancer metastasis through enhanced macrophage recruitment via CCL2/CCR2-induced STAT3 activation. EMBO Mol. Med. 2013, 5, 1383–1401. [Google Scholar] [CrossRef]
- Komohara, Y.; Hasita, H.; Ohnishi, K.; Fujiwara, Y.; Suzu, S.; Eto, M.; Takeya, M. Macrophage infiltration and its prognostic relevance in clear cell renal cell carcinoma. Cancer Sci. 2011, 102, 1424–1431. [Google Scholar] [CrossRef]
- Lin, T.H.; Izumi, K.; Lee, S.O.; Lin, W.J.; Yeh, S.; Chang, C. Anti-androgen receptor ASC-J9 versus anti-androgens MDV3100 (Enzalutamide) or Casodex (Bicalutamide) leads to opposite effects on prostate cancer metastasis via differential modulation of macrophage infiltration and STAT3-CCL2 signaling. Cell Death Dis. 2013, 4, e764. [Google Scholar] [CrossRef]
- Maolake, A.; Izumi, K.; Shigehara, K.; Natsagdorj, A.; Iwamoto, H.; Kadomoto, S.; Takezawa, Y.; Machioka, K.; Narimoto, K.; Namiki, M.; et al. Tumor-associated macrophages promote prostate cancer migration through activation of the CCL22-CCR4 axis. Oncotarget 2017, 8, 9739–9751. [Google Scholar] [CrossRef] [PubMed]
- Maolake, A.; Izumi, K.; Natsagdorj, A.; Iwamoto, H.; Kadomoto, S.; Makino, T.; Naito, R.; Shigehara, K.; Kadono, Y.; Hiratsuka, K.; et al. Tumor necrosis factor-alpha induces prostate cancer cell migration in lymphatic metastasis through CCR7 upregulation. Cancer Sci. 2018, 109, 1524–1531. [Google Scholar] [CrossRef] [PubMed]
- Bingle, L.; Brown, N.J.; Lewis, C.E. The role of tumour-associated macrophages in tumour progression: Implications for new anticancer therapies. J. Pathol. 2002, 196, 254–265. [Google Scholar] [CrossRef] [PubMed]
- Sica, A.; Schioppa, T.; Mantovani, A.; Allavena, P. Tumour-associated macrophages are a distinct M2 polarised population promoting tumour progression: Potential targets of anti-cancer therapy. Eur. J. Cancer 2006, 42, 717–727. [Google Scholar] [CrossRef] [PubMed]
- Qian, B.Z.; Pollard, J.W. Macrophage diversity enhances tumor progression and metastasis. Cell 2010, 141, 39–51. [Google Scholar] [CrossRef] [PubMed]
- Condeelis, J.; Pollard, J.W. Macrophages: Obligate partners for tumor cell migration, invasion, and metastasis. Cell 2006, 124, 263–266. [Google Scholar] [CrossRef] [PubMed]
- Liu, B.; Jia, Y.; Ma, J.; Wu, S.; Jiang, H.; Cao, Y.; Sun, X.; Yin, X.; Yan, S.; Shang, M.; et al. Tumor-associated macrophage-derived CCL20 enhances the growth and metastasis of pancreatic cancer. Acta Biochim. Biophys. Sin. 2016, 48, 1067–1074. [Google Scholar] [CrossRef]
- Zohny, S.F.; Fayed, S.T. Clinical utility of circulating matrix metalloproteinase-7 (MMP-7), CC chemokine ligand 18 (CCL18) and CC chemokine ligand 11 (CCL11) as markers for diagnosis of epithelial ovarian cancer. Med. Oncol. 2010, 27, 1246–1253. [Google Scholar] [CrossRef]
- Zeng, W.; Chang, H.; Ma, M.; Li, Y. CCL20/CCR6 promotes the invasion and migration of thyroid cancer cells via NF-kappa B signaling-induced MMP-3 production. Exp. Mol. Pathol. 2014, 97, 184–190. [Google Scholar] [CrossRef]
- Zhang, X.P.; Hu, Z.J.; Meng, A.H.; Duan, G.C.; Zhao, Q.T.; Yang, J. Role of CCL20/CCR6 and the ERK signaling pathway in lung adenocarcinoma. Oncol. Lett. 2017, 14, 8183–8189. [Google Scholar] [CrossRef]
- Oldham, K.A.; Parsonage, G.; Bhatt, R.I.; Wallace, D.M.; Deshmukh, N.; Chaudhri, S.; Adams, D.H.; Lee, S.P. T lymphocyte recruitment into renal cell carcinoma tissue: A role for chemokine receptors CXCR3, CXCR6, CCR5, and CCR6. Eur. Urol. 2012, 61, 385–394. [Google Scholar] [CrossRef] [PubMed]
- Zhu, C.C.; Chen, C.; Xu, Z.Q.; Zhao, J.K.; Ou, B.C.; Sun, J.; Zheng, M.H.; Zong, Y.P.; Lu, A.G. CCR6 promotes tumor angiogenesis via the AKT/NF-kappaB/VEGF pathway in colorectal cancer. Biochim. Biophys. Acta Mol. Basis Dis. 2018, 1864, 387–397. [Google Scholar] [CrossRef] [PubMed]
- Han, G.; Wu, D.; Yang, Y.; Li, Z.; Zhang, J.; Li, C. CrkL meditates CCL20/CCR6-induced EMT in gastric cancer. Cytokine 2015, 76, 163–169. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Zhu, D.; Lv, Q.; Yi, Y.; Li, F.; Zhang, W. The key role of astrocyte elevated gene-1 in CCR6-induced EMT in cervical cancer. Tumor. Biol. 2015, 36, 9763–9767. [Google Scholar] [CrossRef] [PubMed]
- Izumi, K.; Mizokami, A.; Lin, H.P.; Ho, H.M.; Iwamoto, H.; Maolake, A.; Natsagdorj, A.; Kitagawa, Y.; Kadono, Y.; Miyamoto, H.; et al. Serum chemokine (CC motif) ligand 2 level as a diagnostic, predictive, and prognostic biomarker for prostate cancer. Oncotarget 2016, 7, 8389–8398. [Google Scholar] [CrossRef]
- Urata, S.; Izumi, K.; Hiratsuka, K.; Maolake, A.; Natsagdorj, A.; Shigehara, K.; Iwamoto, H.; Kadomoto, S.; Makino, T.; Naito, R.; et al. C-C motif ligand 5 promotes migration of prostate cancer cells in the prostate cancer bone metastasis microenvironment. Cancer Sci. 2018, 109, 724–731. [Google Scholar] [CrossRef]
- Genin, M.; Clement, F.; Fattaccioli, A.; Raes, M.; Michiels, C. M1 and M2 macrophages derived from THP-1 cells differentially modulate the response of cancer cells to etoposide. BMC Cancer 2015, 15, 577. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| CCR6 | CD68 | |||||||
|---|---|---|---|---|---|---|---|---|
| Parameters | n | Pos | Neg * | p | Pos | Neg | p | |
| Gender | Male | 31 | 20 | 11 | n.s. | 12 | 19 | n.s. |
| Female | 11 | 7 | 4 | 3 | 8 | |||
| Age | ≤60 | 16 | 9 | 7 | n.s. | 6 | 10 | n.s. |
| >60 | 26 | 18 | 8 | 9 | 17 | |||
| Histology | Pure CC | 37 | 24 | 13 | n.s. | 13 | 24 | n.s. |
| CC with SC | 5 | 3 | 2 | 2 | 3 | |||
| TNM | T1N0M0 | 16 | 6 | 10 | 0.008 | 7 | 9 | n.s. |
| Other | 26 | 21 | 5 | 8 | 18 | |||
| GAPDH | forward | 5′-TCT CTG CTC CTC CTG TTC GA-3′ |
| reverse | 5′-GCG CCC AAT ACG ACC AAA TC-3′ | |
| Snail | forward | 5′-ATG GCC ATT TCT GTG GAG GG-3′ |
| reverse | 5′-CAA AAA CCC ACG CAG ACA GG-3′ | |
| Twist | forward | 5′-ATT CAG ACC CTC AAG CTG GC-3′ |
| reverse | 5′-AGC TTG CCA TCT TGG AGT CC-3′ | |
| Vimentin | forward | 5′-AAC TTA GGG GCG CTC TTG TC-3′ |
| reverse | 5′-ATT CAA GTC TCA GCG GGC TC-3′ | |
| CCR6 | forward | 5′-AAT CGC TTG AAC CCA GAA GG-3′ |
| reverse | 5′-GAG TCT CGC TTT GTC ACC CA-3′ | |
| CCL20 | forward | 5′-GGA ATG GAA TTG GAC ATA GCC-3′ |
| reverse | 5′-CCT CCA TGA TGT GCA AGT GA-3′ |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kadomoto, S.; Izumi, K.; Hiratsuka, K.; Nakano, T.; Naito, R.; Makino, T.; Iwamoto, H.; Yaegashi, H.; Shigehara, K.; Kadono, Y.; et al. Tumor-Associated Macrophages Induce Migration of Renal Cell Carcinoma Cells via Activation of the CCL20-CCR6 Axis. Cancers 2020, 12, 89. https://doi.org/10.3390/cancers12010089
Kadomoto S, Izumi K, Hiratsuka K, Nakano T, Naito R, Makino T, Iwamoto H, Yaegashi H, Shigehara K, Kadono Y, et al. Tumor-Associated Macrophages Induce Migration of Renal Cell Carcinoma Cells via Activation of the CCL20-CCR6 Axis. Cancers. 2020; 12(1):89. https://doi.org/10.3390/cancers12010089
Chicago/Turabian StyleKadomoto, Suguru, Kouji Izumi, Kaoru Hiratsuka, Taito Nakano, Renato Naito, Tomoyuki Makino, Hiroaki Iwamoto, Hiroshi Yaegashi, Kazuyoshi Shigehara, Yoshifumi Kadono, and et al. 2020. "Tumor-Associated Macrophages Induce Migration of Renal Cell Carcinoma Cells via Activation of the CCL20-CCR6 Axis" Cancers 12, no. 1: 89. https://doi.org/10.3390/cancers12010089
APA StyleKadomoto, S., Izumi, K., Hiratsuka, K., Nakano, T., Naito, R., Makino, T., Iwamoto, H., Yaegashi, H., Shigehara, K., Kadono, Y., Nakata, H., Saito, Y., Nakagawa-Goto, K., & Mizokami, A. (2020). Tumor-Associated Macrophages Induce Migration of Renal Cell Carcinoma Cells via Activation of the CCL20-CCR6 Axis. Cancers, 12(1), 89. https://doi.org/10.3390/cancers12010089