Molecular Mode of Action of TRAIL Receptor Agonists—Common Principles and Their Translational Exploitation


Abstract
1. Introduction
2. Death Signaling by Tumor Necrosis Factor-Related Apoptosis-Inducing Ligand Receptors 1 and 2
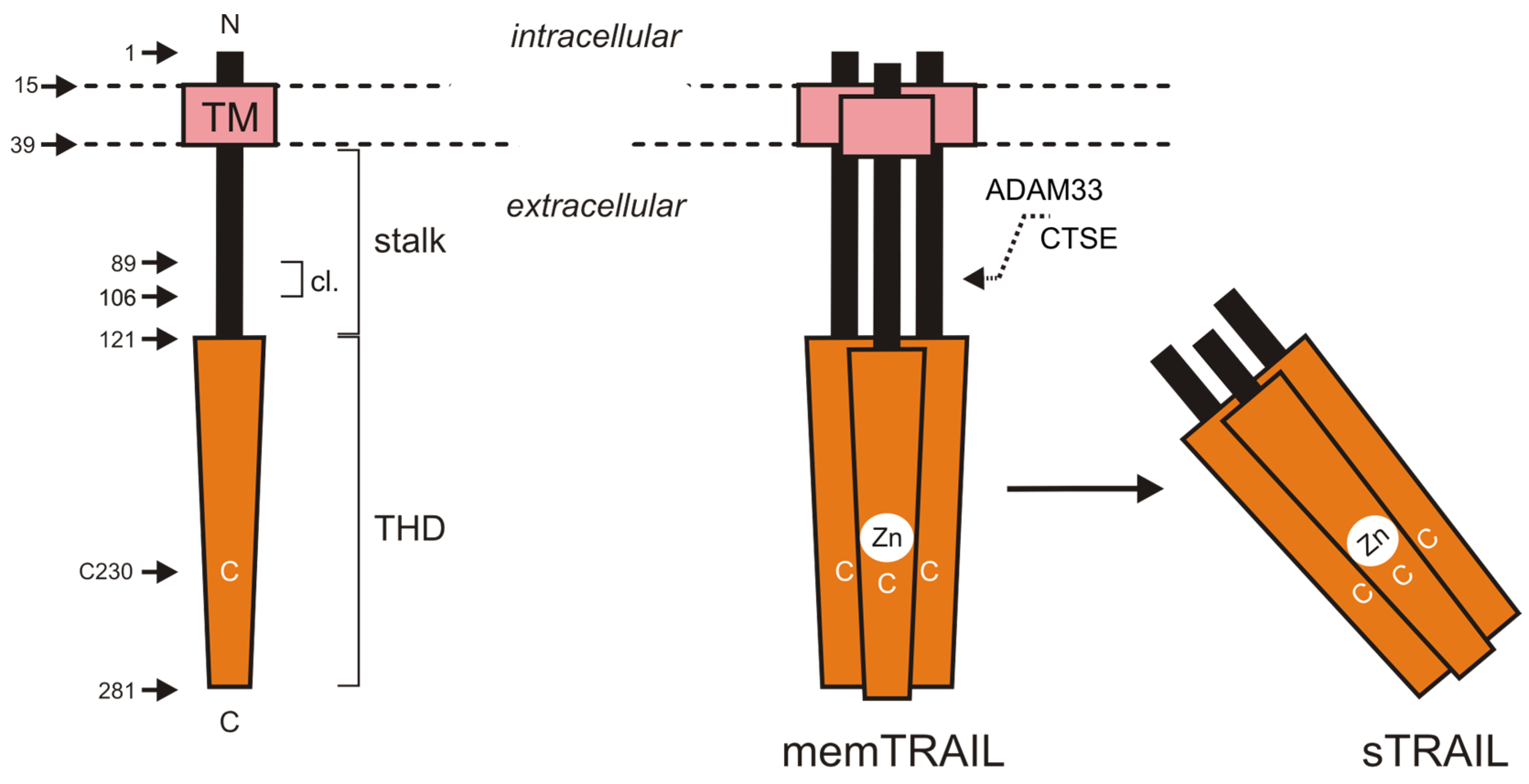
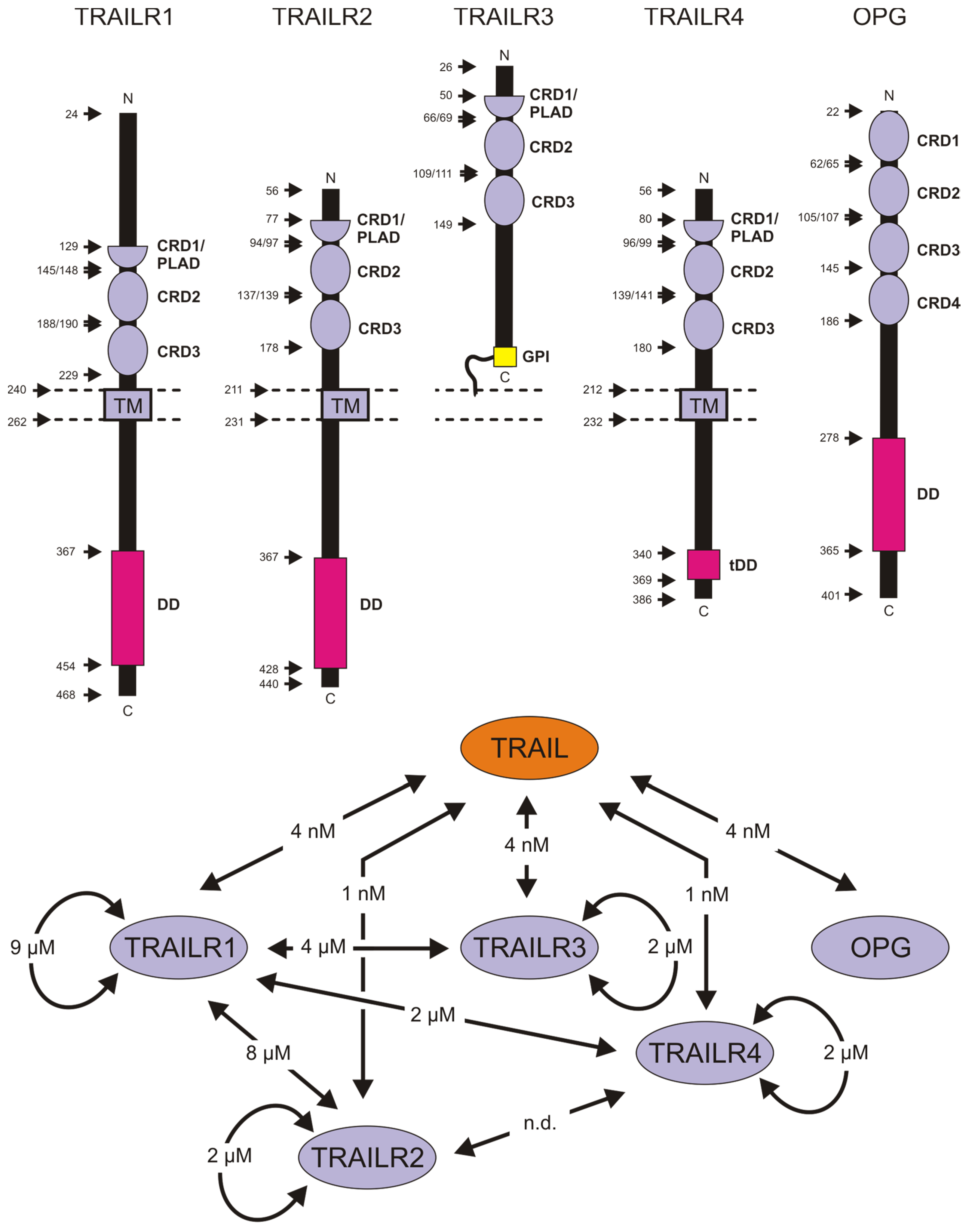
2.1. Tumor Necrosis Factor-Related Apoptosis-Inducing Ligand and Its Receptors
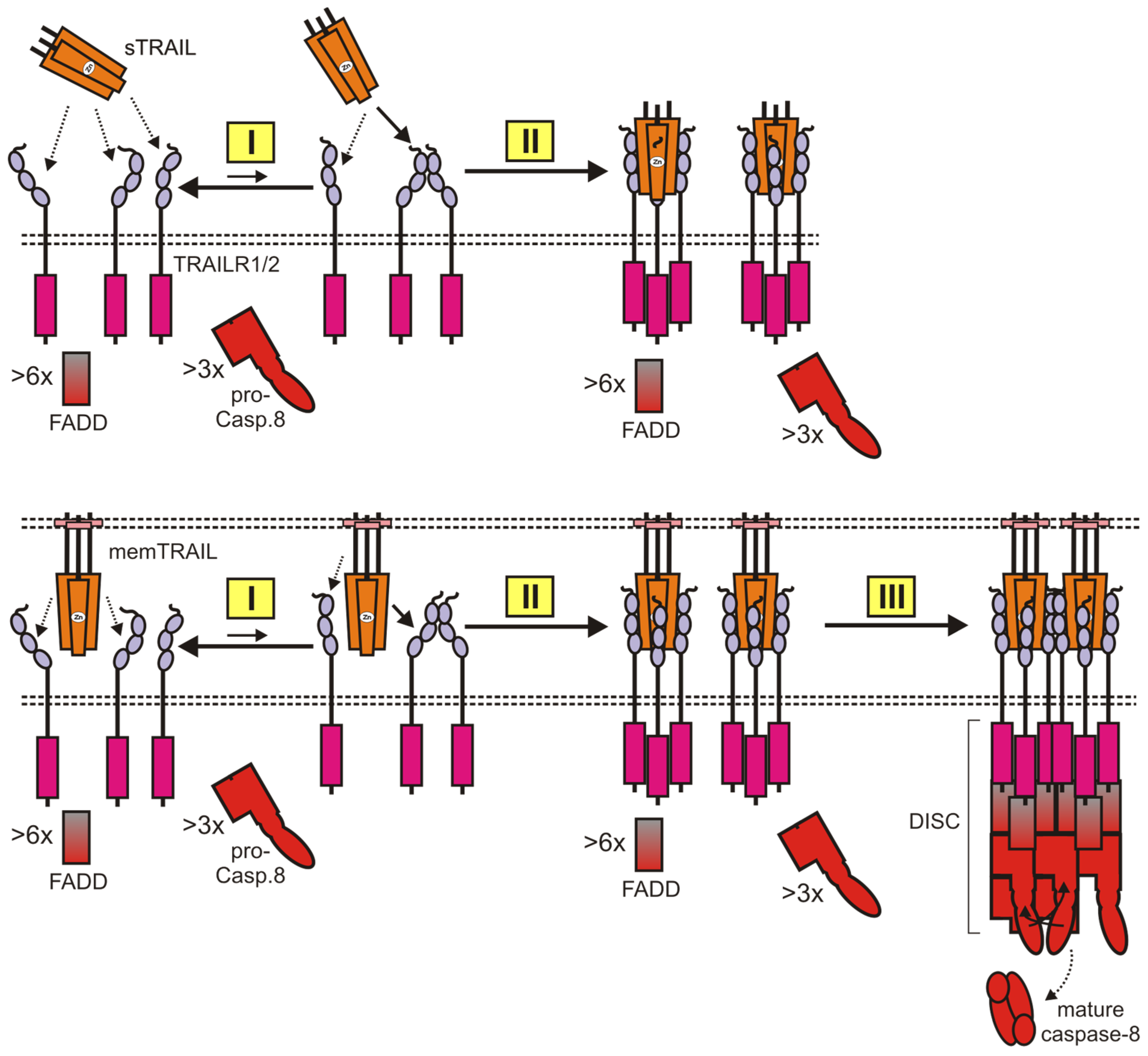
2.2. TRAIL Death Receptor-Induced Engagement of Cytotoxic Signaling Pathways
2.3. TRAIL-Stimulated Non-Cell Death Signaling
3. Conventional TRAIL Death Receptor Agonists and Their Limitations: The Lessons from Preclinical Studies and Clinical Trials
4. Next Generation Ligand-Based TRAIL Death Receptor Agonists
4.1. Stabilized TRAIL Trimers and TRAILR1- and TRAILR2-Specific TRAIL Mutants
4.2. TRAIL Variants with Superior Specific Activity
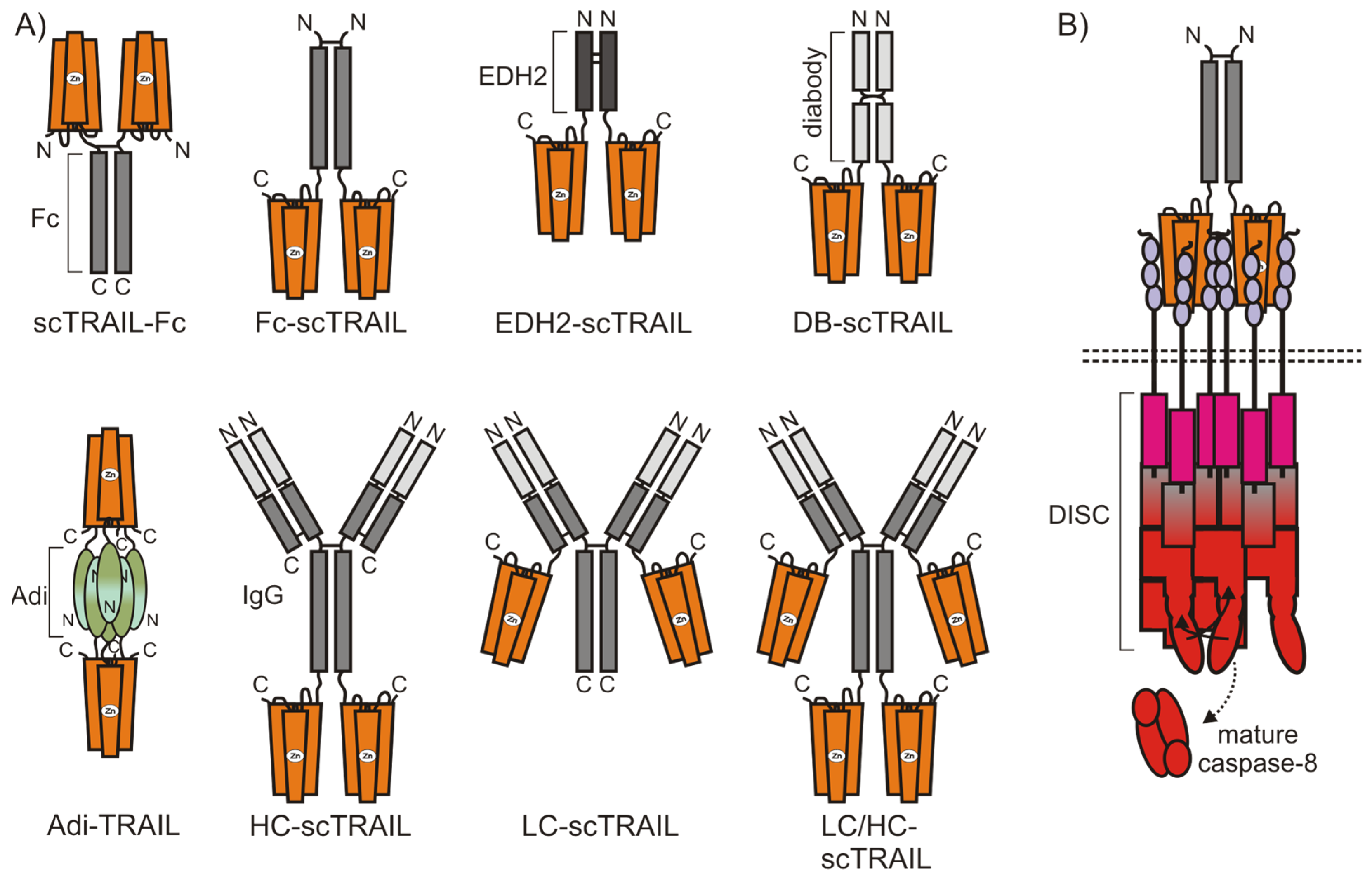
4.2.1. TRAIL Fusion Proteins Containing Two or More TRAIL Trimers
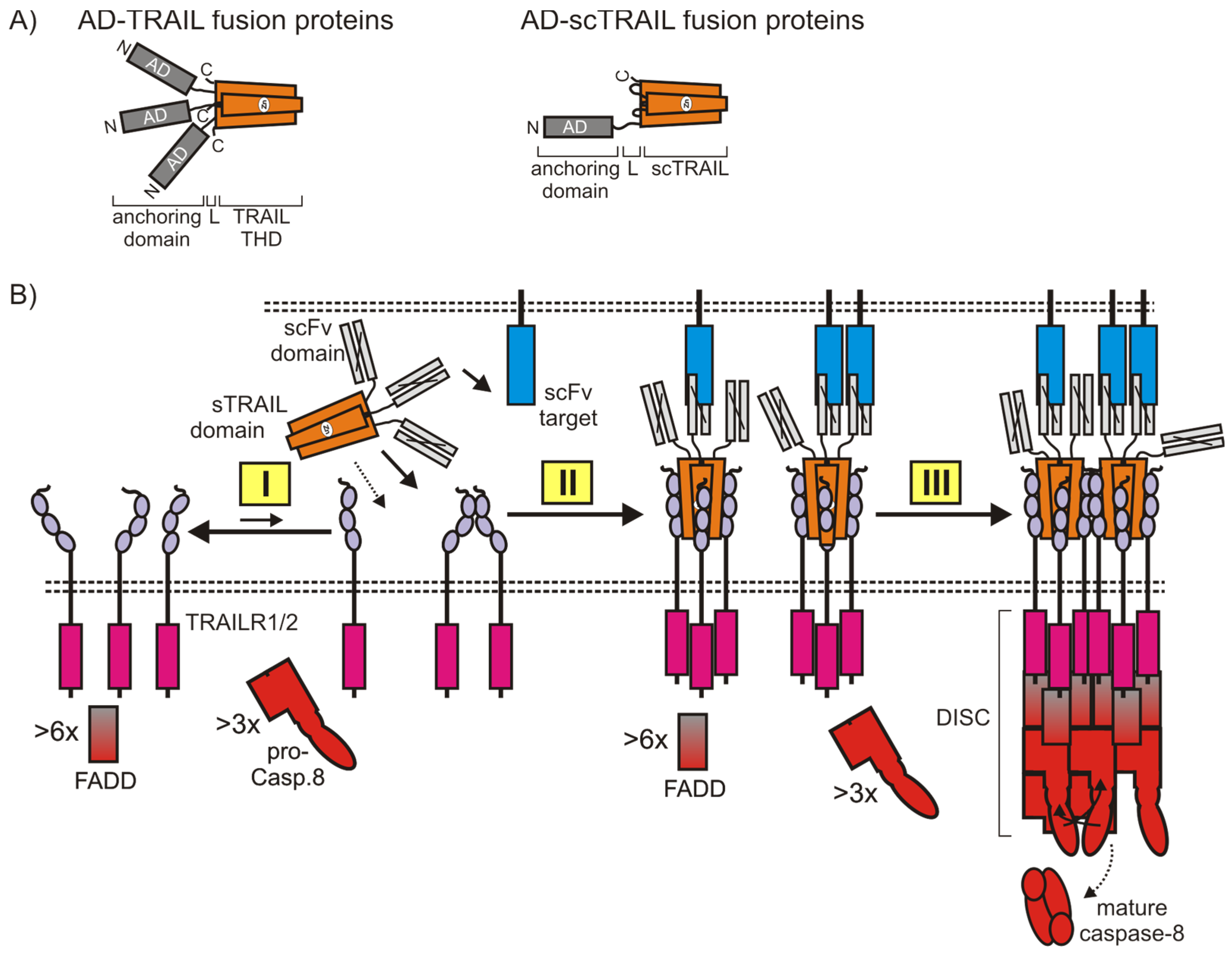
4.2.2. Cell Surface Anchoring TRAIL Fusion Proteins
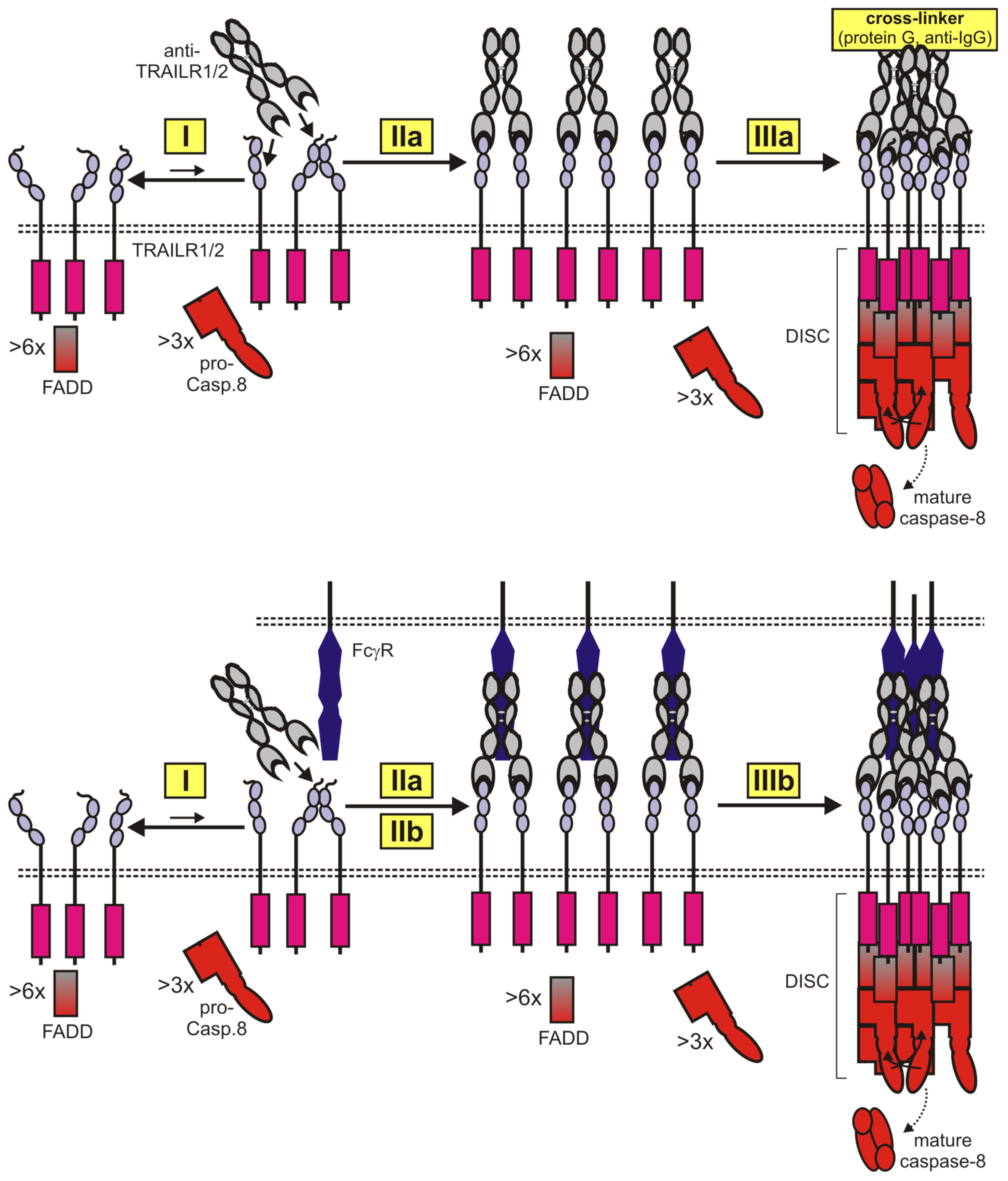
5. Next Generation TRAIL Death Receptor Agonists Based on Antibodies and Related Molecules
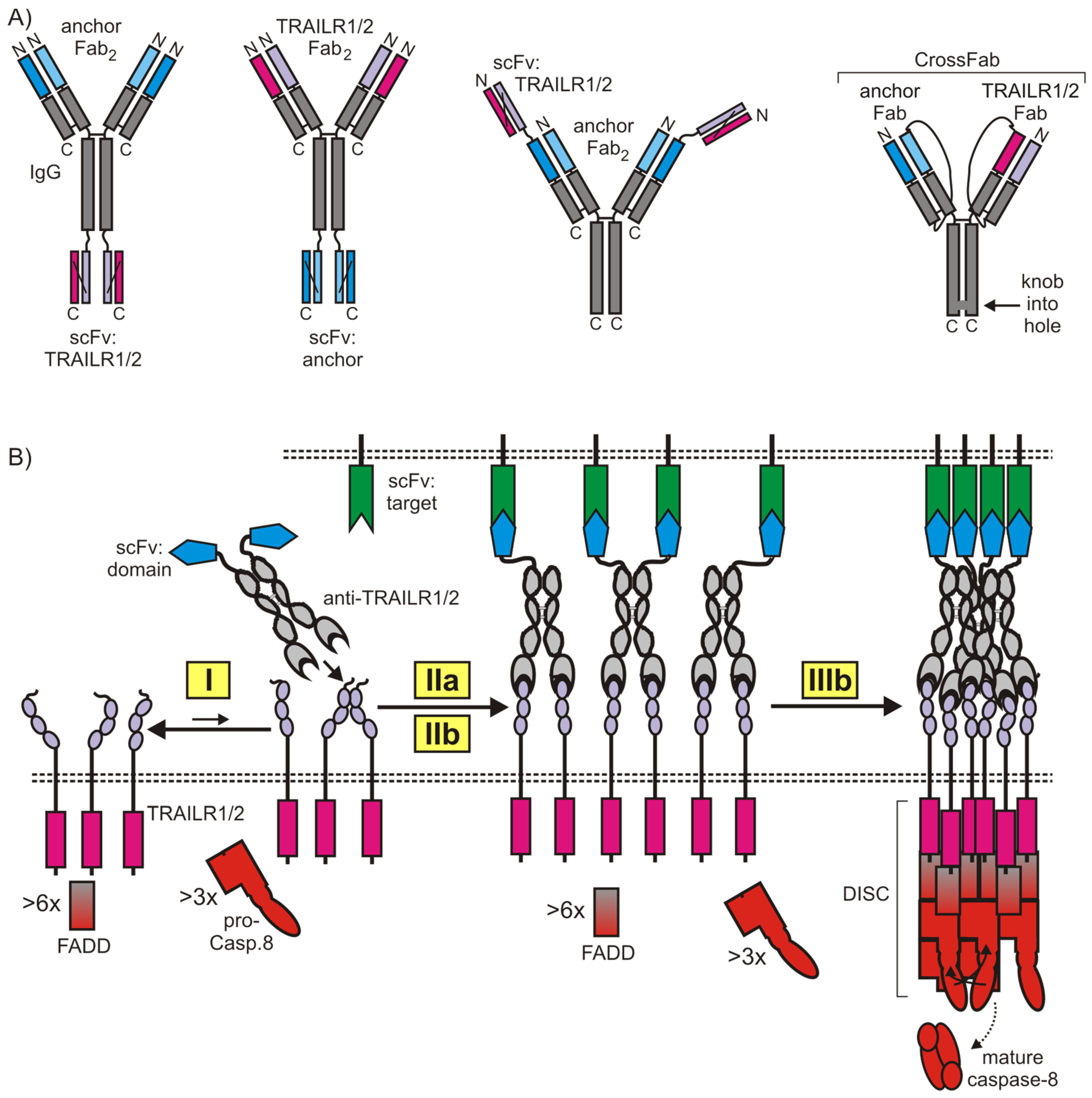
5.1. Anti-TRAIL Death Receptor Variants with Increased Valency
5.2. Cell Surface Anchored Anti-TRAIL Death Receptor Antibody Variants
6. Conclusions
Funding
Acknowledgments
Conflicts of Interest
References
- Bodmer, J.L.; Schneider, P.; Tschopp, J. The molecular architecture of the TNF superfamily. Trends Biochem. Sci. 2002, 27, 19–26. [Google Scholar] [CrossRef]
- Berg, D.; Lehne, M.; Muller, N.; Siegmund, D.; Munkel, S.; Sebald, W.; Pfizenmaier, K.; Wajant, H. Enforced covalent trimerization increases the activity of the TNF ligand family members TRAIL and CD95L. Cell Death Differ. 2007, 14, 2021–2034. [Google Scholar] [CrossRef] [PubMed]
- Wajant, H.; Moosmayer, D.; Wuest, T.; Bartke, T.; Gerlach, E.; Schonherr, U.; Peters, N.; Scheurich, P.; Pfizenmaier, K. Differential activation of TRAIL-R1 and -2 by soluble and membrane TRAIL allows selective surface antigen-directed activation of TRAIL-R2 by a soluble TRAIL derivative. Oncogene 2001, 20, 4101–4106. [Google Scholar] [CrossRef] [PubMed]
- Mariani, S.M.; Krammer, P.H. Differential regulation of TRAIL and CD95 ligand in transformed cells of the T and B lymphocyte lineage. Eur. J. Immunol. 1998, 28, 973–982. [Google Scholar] [CrossRef]
- Kemp, T.J.; Ludwig, A.T.; Earel, J.K.; Moore, J.M.; Vanoosten, R.L.; Moses, B.; Leidal, K.; Nauseef, W.M.; Griffith, T.S. Neutrophil stimulation with Mycobacterium bovis bacillus Calmette-Guerin (BCG) results in the release of functional soluble TRAIL/Apo-2L. Blood 2005, 106, 3474–3482. [Google Scholar] [CrossRef]
- Dokouhaki, P.; Schuh, N.W.; Joe, B.; Allen, C.A.; Der, S.D.; Tsao, M.S.; Zhang, L. NKG2D regulates production of soluble TRAIL by ex vivo expanded human gammadelta T cells. Eur. J. Immunol. 2013, 43, 3175–3182. [Google Scholar] [CrossRef] [PubMed]
- Han, L.H.; Sun, W.S.; Ma, C.H.; Zhang, L.N.; Liu, S.X.; Zhang, Q.; Gao, L.F.; Chen, Y.H. Detection of soluble TRAIL in HBV infected patients and its clinical implications. World J. Gastroenterol. 2002, 8, 1077–1080. [Google Scholar] [CrossRef]
- Lub-de Hooge, M.N.; de Vries, E.G.; de Jong, S.; Bijl, M. Soluble TRAIL concentrations are raised in patients with systemic lupus erythematosus. Ann. Rheum. Dis. 2005, 64, 854–858. [Google Scholar] [CrossRef]
- Iłzecka, J. Decreased serum-soluble TRAIL levels in patients with amyotrophic lateral sclerosis. Acta Neurol. Scand. 2008, 117, 343–346. [Google Scholar] [CrossRef]
- Liabeuf, S.; Barreto, D.V.; Barreto, F.C.; Chasseraud, M.; Brazier, M.; Choukroun, G.; Kamel, S.; Massy, Z.A. The circulating soluble TRAIL is a negative marker for inflammation inversely associated with the mortality risk in chronic kidney disease patients. Nephrol. Dial. Transplant. 2010, 25, 2596–2602. [Google Scholar] [CrossRef]
- Azab, N.A.; Rady, H.M.; Marzouk, S.A. Elevated serum TRAIL levels in scleroderma patients and its possible association with pulmonary involvement. Clin. Rheumatol. 2012, 31, 1359–1364. [Google Scholar] [CrossRef] [PubMed]
- Agostinis, C.; Bulla, R.; Tisato, V.; De Seta, F.; Alberico, S.; Secchiero, P.; Zauli, G. Soluble TRAIL is elevated in recurrent miscarriage and inhibits the in vitro adhesion and migration of HTR8 trophoblastic cells. Hum. Reprod. 2012, 27, 2941–2947. [Google Scholar] [CrossRef] [PubMed]
- Chaemsaithong, P.; Romero, R.; Korzeniewski, S.J.; Schwartz, A.G.; Stampalija, T.; Dong, Z.; Yeo, L.; Hernandez-Andrade, E.; Hassan, S.S.; Chaiworapongsa, T. Soluble TRAIL in normal pregnancy and acute pyelonephritis: A potential explanation for the susceptibility of pregnant women to microbial products and infection. J. Matern. Fetal Neonatal Med. 2013, 26, 1568–1575. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Tawdy, M.H.; Abd El Nasser, M.M.; Abd El Shafy, S.S.; Nada, M.A.; El Sirafy, M.N.; Magd, A.H. Role of serum TRAIL level and TRAIL apoptosis gene expression in multiple sclerosis and relation to brain atrophy. J. Clin. Neurosci. 2014, 21, 1606–1611. [Google Scholar] [CrossRef] [PubMed]
- Tornese, G.; Iafusco, D.; Monasta, L.; Agnoletto, C.; Tisato, V.; Ventura, A.; Zauli, G.; Secchiero, P. The levels of circulating TRAIL at the onset of type 1 diabetes are markedly decreased in patients with ketoacidosis and with the highest insulin requirement. Acta Diabetol. 2014, 51, 239–246. [Google Scholar] [CrossRef] [PubMed]
- Cheng, W.; Liu, F.; Wang, Z.; Zhang, Y.; Zhao, Y.X.; Zhang, Q.; Jiang, F. Soluble TRAIL Concentration in Serum Is Elevated in People with Hypercholesterolemia. PLoS ONE 2015, 10, e0144015. [Google Scholar] [CrossRef] [PubMed]
- Zhou, H.; Wang, Y.; Bi, K.; Qi, H.; Song, S.; Zhou, M.; Chen, L.; Wang, G.; Duan, T. Serum-soluble TRAIL: A potential biomarker for disease activity in myositis patients. Clin. Rheumatol. 2019, 38, 1425–1431. [Google Scholar] [CrossRef] [PubMed]
- Zou, J.; Zhu, F.; Liu, J.; Wang, W.; Zhang, R.; Garlisi, C.G.; Liu, Y.H.; Wang, S.; Shah, H.; Wan, Y.; et al. Catalytic activity of human ADAM33. J. Biol. Chem. 2004, 279, 9818–9830. [Google Scholar] [CrossRef]
- Kawakubo, T.; Okamoto, K.; Iwata, J.; Shin, M.; Okamoto, Y.; Yasukochi, A.; Nakayama, K.I.; Kadowaki, T.; Tsukuba, T.; Yamamoto, K. Cathepsin E prevents tumor growth and metastasis by catalyzing the proteolytic release of soluble TRAIL from tumor cell surface. Cancer Res. 2007, 67, 10869–10878. [Google Scholar] [CrossRef]
- Bodmer, J.L.; Meier, P.; Tschopp, J.; Schneider, P. Cysteine 230 is essential for the structure and activity of the cytotoxic ligand TRAIL. J. Biol. Chem. 2000, 275, 20632–20637. [Google Scholar] [CrossRef]
- Hymowitz, S.G.; O’Connell, M.P.; Ultsch, M.H.; Hurst, A.; Totpal, K.; Ashkenazi, A.; de Vos, A.M.; Kelley, R.F. A unique zinc-binding site revealed by a high-resolution X-ray structure of homotrimeric Apo2L/TRAIL. Biochemistry 2000, 39, 633–640. [Google Scholar] [CrossRef] [PubMed]
- Trabzuni, D.; Famulski, K.S.; Ahmad, M. Functional analysis of tumour necrosis factor-alpha-related apoptosis-inducing ligand (TRAIL): Cysteine-230 plays a critical role in the homotrimerization and biological activity of this novel tumoricidal cytokine. Biochem. J. 2000, 350, 505–510. [Google Scholar] [CrossRef] [PubMed]
- Kayagaki, N.; Yamaguchi, N.; Nakayama, M.; Eto, H.; Okumura, K.; Yagita, H. Type I interferons (IFNs) regulate tumor necrosis factor-related apoptosis-inducing ligand (TRAIL) expression on human T cells: A novel mechanism for the antitumor effects of type I IFNs. J. Exp. Med. 1999, 189, 1451–1460. [Google Scholar] [CrossRef] [PubMed]
- Fanger, N.A.; Maliszewski, C.R.; Schooley, K.; Griffith, T.S. Human dendritic cells mediate cellular apoptosis via tumor necrosis factor-related apoptosis-inducing ligand (TRAIL). J. Exp. Med. 1999, 190, 1155–1164. [Google Scholar] [CrossRef] [PubMed]
- Chaperot, L.; Blum, A.; Manches, O.; Lui, G.; Angel, J.; Molens, J.P.; Plumas, J. Virus or TLR agonists induce TRAIL-mediated cytotoxic activity of plasmacytoid dendritic cells. J. Immunol. 2006, 176, 248–255. [Google Scholar] [CrossRef] [PubMed]
- Hardy, A.W.; Graham, D.R.; Shearer, G.M.; Herbeuval, J.P. HIV turns plasmacytoid dendritic cells (pDC) into TRAIL-expressing killer pDC and down-regulates HIV coreceptors by Toll-like receptor 7-induced IFN-alpha. Proc. Natl. Acad. Sci. USA 2007, 104, 17453–17458. [Google Scholar] [CrossRef]
- Johnsen, A.C.; Haux, J.; Steinkjer, B.; Nonstad, U.; Egeberg, K.; Sundan, A.; Ashkenazi, A.; Espevik, T. Regulation of APO-2 ligand/trail expression in NK cells-involvement in NK cell-mediated cytotoxicity. Cytokine 1999, 11, 664–672. [Google Scholar] [CrossRef]
- Nieda, M.; Nicol, A.; Koezuka, Y.; Kikuchi, A.; Lapteva, N.; Tanaka, Y.; Tokunaga, K.; Suzuki, K.; Kayagaki, N.; Yagita, H.; et al. TRAIL expression by activated human CD4(+)V alpha 24NKT cells induces in vitro and in vivo apoptosis of human acute myeloid leukemia cells. Blood 2001, 97, 2067–2074. [Google Scholar] [CrossRef]
- Dorothee, G.; Vergnon, I.; Menez, J.; Echchakir, H.; Grunenwald, D.; Kubin, M.; Chouaib, S.; Mami-Chouaib, F. Tumor-infiltrating CD4+ T lymphocytes express APO2 ligand (APO2L)/TRAIL upon specific stimulation with autologous lung carcinoma cells: Role of IFN-alpha on APO2L/TRAIL expression and -mediated cytotoxicity. J. Immunol. 2002, 169, 809–817. [Google Scholar] [CrossRef]
- Turchinovich, G.; Ganter, S.; Barenwaldt, A.; Finke, D. NKp46 Calibrates Tumoricidal Potential of Type 1 Innate Lymphocytes by Regulating TRAIL Expression. J. Immunol. 2018, 200, 3762–3768. [Google Scholar] [CrossRef]
- von Karstedt, S.; Montinaro, A.; Walczak, H. Exploring the TRAILs less travelled: TRAIL in cancer biology and therapy. Nat. Rev. Cancer 2017, 17, 352–366. [Google Scholar] [CrossRef] [PubMed]
- Gochuico, B.R.; Zhang, J.; Ma, B.Y.; Marshak-Rothstein, A.; Fine, A. TRAIL expression in vascular smooth muscle. Am. J. Physiol. Lung Cell. Mol. Physiol. 2000, 278, L1045–L1050. [Google Scholar] [CrossRef] [PubMed]
- Sambandam, Y.; Reddy, S.V.; Mulligan, J.L.; Voelkel-Johnson, C.; Wagner, C.L. Vitamin D Modulation of TRAIL Expression in Human Milk and Mammary Epithelial Cells. Sci. Rep. 2017, 7, 4362. [Google Scholar] [CrossRef] [PubMed]
- Wang, Q.; Zhou, Y.; Weiss, H.L.; Chow, C.W.; Evers, B.M. NFATc1 regulation of TRAIL expression in human intestinal cells. PLoS ONE 2011, 6, e19882. [Google Scholar] [CrossRef] [PubMed]
- Zahn, S.; Rehkamper, C.; Ferring-Schmitt, S.; Bieber, T.; Tuting, T.; Wenzel, J. Interferon-alpha stimulates TRAIL expression in human keratinocytes and peripheral blood mononuclear cells: Implications for the pathogenesis of cutaneous lupus erythematosus. Br. J. Dermatol. 2011, 165, 1118–1123. [Google Scholar] [CrossRef] [PubMed]
- Locksley, R.M.; Killeen, N.; Lenardo, M.J. The TNF and TNF receptor superfamilies: Integrating mammalian biology. Cell 2001, 104, 487–501. [Google Scholar] [CrossRef]
- Yuan, X.; Gajan, A.; Chu, Q.; Xiong, H.; Wu, K.; Wu, G.S. Developing TRAIL/TRAIL death receptor-based cancer therapies. Cancer Metastasis Rev. 2018, 37, 733–748. [Google Scholar] [CrossRef]
- Degli-Esposti, M.A.; Dougall, W.C.; Smolak, P.J.; Waugh, J.Y.; Smith, C.A.; Goodwin, R.G. The novel receptor TRAIL-R4 induces NF-kappaB and protects against TRAIL-mediated apoptosis, yet retains an incomplete death domain. Immunity 1997, 7, 813–820. [Google Scholar] [CrossRef]
- Lalaoui, N.; Morle, A.; Merino, D.; Jacquemin, G.; Iessi, E.; Morizot, A.; Shirley, S.; Robert, B.; Solary, E.; Garrido, C.; et al. TRAIL-R4 promotes tumor growth and resistance to apoptosis in cervical carcinoma HeLa cells through AKT. PLoS ONE 2011, 6, e19679. [Google Scholar] [CrossRef]
- Lamoureux, F.; Moriceau, G.; Picarda, G.; Rousseau, J.; Trichet, V.; Redini, F. Regulation of osteoprotegerin pro- or anti-tumoral activity by bone tumor microenvironment. Biochim. Biophys. Acta 2010, 1805, 17–24. [Google Scholar] [CrossRef]
- Lee, H.W.; Lee, S.H.; Lee, H.W.; Ryu, Y.W.; Kwon, M.H.; Kim, Y.S. Homomeric and heteromeric interactions of the extracellular domains of death receptors and death decoy receptors. Biochem. Biophys. Res. Commun. 2005, 330, 1205–1212. [Google Scholar] [CrossRef] [PubMed]
- Lang, I.; Fullsack, S.; Wyzgol, A.; Fick, A.; Trebing, J.; Arana, J.A.; Schafer, V.; Weisenberger, D.; Wajant, H. Binding Studies of TNF Receptor Superfamily (TNFRSF) Receptors on Intact Cells. J. Biol. Chem. 2016, 291, 5022–5037. [Google Scholar] [CrossRef] [PubMed]
- Siegmund, D.; Lang, I.; Wajant, H. Cell death-independent activities of the death receptors CD95, TRAILR1, and TRAILR2. FEBS J. 2017, 284, 1131–1159. [Google Scholar] [CrossRef] [PubMed]
- Chan, F.K.; Chun, H.J.; Zheng, L.; Siegel, R.M.; Bui, K.L.; Lenardo, M.J. A domain in TNF receptors that mediates ligand-independent receptor assembly and signaling. Science 2000, 288, 2351–2354. [Google Scholar] [CrossRef] [PubMed]
- Clancy, L.; Mruk, K.; Archer, K.; Woelfel, M.; Mongkolsapaya, J.; Screaton, G.; Lenardo, M.J.; Chan, F.K. Preligand assembly domain-mediated ligand-independent association between TRAIL receptor 4 (TR4) and TR2 regulates TRAIL-induced apoptosis. Proc. Natl. Acad. Sci. USA 2005, 102, 18099–18104. [Google Scholar] [CrossRef] [PubMed]
- Neumann, S.; Hasenauer, J.; Pollak, N.; Scheurich, P. Dominant negative effects of tumor necrosis factor (TNF)-related apoptosis-inducing ligand (TRAIL) receptor 4 on TRAIL receptor 1 signaling by formation of heteromeric complexes. J. Biol. Chem. 2014, 289, 16576–16587. [Google Scholar] [CrossRef]
- Merino, D.; Lalaoui, N.; Morizot, A.; Schneider, P.; Solary, E.; Micheau, O. Differential inhibition of TRAIL-mediated DR5-DISC formation by decoy receptors 1 and 2. Mol. Cell. Biol. 2006, 26, 7046–7055. [Google Scholar] [CrossRef]
- Reis, C.R.; van der Sloot, A.M.; Natoni, A.; Szegezdi, E.; Setroikromo, R.; Meijer, M.; Sjollema, K.; Stricher, F.; Cool, R.H.; Samali, A.; et al. Rapid and efficient cancer cell killing mediated by high-affinity death receptor homotrimerizing TRAIL variants. Cell Death Dis. 2010, 1, e83. [Google Scholar] [CrossRef]
- Szegezdi, E.; van der Sloot, A.M.; Mahalingam, D.; O’Leary, L.; Cool, R.H.; Munoz, I.G.; Montoya, G.; Quax, W.J.; de Jong, S.; Samali, A.; et al. Kinetics in signal transduction pathways involving promiscuous oligomerizing receptors can be determined by receptor specificity: Apoptosis induction by TRAIL. Mol. Cell. Proteom. 2012, 11, M111.013730. [Google Scholar] [CrossRef]
- Dufour, F.; Rattier, T.; Shirley, S.; Picarda, G.; Constantinescu, A.A.; Morle, A.; Zakaria, A.B.; Marcion, G.; Causse, S.; Szegezdi, E.; et al. N-glycosylation of mouse TRAIL-R and human TRAIL-R1 enhances TRAIL-induced death. Cell Death Differ. 2017, 24, 500–510. [Google Scholar] [CrossRef]
- Wagner, K.W.; Punnoose, E.A.; Januario, T.; Lawrence, D.A.; Pitti, R.M.; Lancaster, K.; Lee, D.; von Goetz, M.; Yee, S.F.; Totpal, K.; et al. Death-receptor O-glycosylation controls tumor-cell sensitivity to the proapoptotic ligand Apo2L/TRAIL. Nat. Med. 2007, 13, 1070–1077. [Google Scholar] [CrossRef] [PubMed]
- Estornes, Y.; Dondelinger, Y.; Weber, K.; Bruggeman, I.; Peall, A.; MacFarlane, M.; Lebecque, S.; Vandenabeele, P.; Bertrand, M.J.M. N-glycosylation of mouse TRAIL-R restrains TRAIL-induced apoptosis. Cell Death Dis. 2018, 9, 494. [Google Scholar] [CrossRef] [PubMed]
- Pan, L.; Fu, T.M.; Zhao, W.; Zhao, L.; Chen, W.; Qiu, C.; Liu, W.; Liu, Z.; Piai, A.; Fu, Q.; et al. Higher-Order Clustering of the Transmembrane Anchor of DR5 Drives Signaling. Cell 2019, 176, 1477–1489. [Google Scholar] [CrossRef] [PubMed]
- Cha, S.S.; Sung, B.J.; Kim, Y.A.; Song, Y.L.; Kim, H.J.; Kim, S.; Lee, M.S.; Oh, B.H. Crystal structure of TRAIL-DR5 complex identifies a critical role of the unique frame insertion in conferring recognition specificity. J. Biol. Chem. 2000, 275, 31171–31177. [Google Scholar] [CrossRef] [PubMed]
- Hymowitz, S.G.; Christinger, H.W.; Fuh, G.; Ultsch, M.; O’Connell, M.; Kelley, R.F.; Ashkenazi, A.; de Vos, A.M. Triggering cell death: The crystal structure of Apo2L/TRAIL in a complex with death receptor 5. Mol. Cell 1999, 4, 563–571. [Google Scholar] [CrossRef]
- Mongkolsapaya, J.; Grimes, J.M.; Chen, N.; Xu, X.N.; Stuart, D.I.; Jones, E.Y.; Screaton, G.R. Structure of the TRAIL-DR5 complex reveals mechanisms conferring specificity in apoptotic initiation. Nat. Struct. Biol. 1999, 6, 1048–1053. [Google Scholar] [CrossRef] [PubMed]
- Valley, C.C.; Lewis, A.K.; Mudaliar, D.J.; Perlmutter, J.D.; Braun, A.R.; Karim, C.B.; Thomas, D.D.; Brody, J.R.; Sachs, J.N. Tumor necrosis factor-related apoptosis-inducing ligand (TRAIL) induces death receptor 5 networks that are highly organized. J. Biol. Chem. 2012, 287, 21265–21278. [Google Scholar] [CrossRef] [PubMed]
- Lewis, A.K.; James, Z.M.; McCaffrey, J.E.; Braun, A.R.; Karim, C.B.; Thomas, D.D.; Sachs, J.N. Open and closed conformations of the isolated transmembrane domain of death receptor 5 support a new model of activation. Biophys. J. 2014, 106, L21–L24. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Fu, T.M.; Li, Y.; Lu, A.; Li, Z.; Vajjhala, P.R.; Cruz, A.C.; Srivastava, D.B.; DiMaio, F.; Penczek, P.A.; Siegel, R.M.; et al. Cryo-EM Structure of Caspase-8 Tandem DED Filament Reveals Assembly and Regulation Mechanisms of the Death-Inducing Signaling Complex. Mol. Cell 2016, 64, 236–250. [Google Scholar] [CrossRef] [PubMed]
- Schleich, K.; Warnken, U.; Fricker, N.; Ozturk, S.; Richter, P.; Kammerer, K.; Schnolzer, M.; Krammer, P.H.; Lavrik, I.N. Stoichiometry of the CD95 death-inducing signaling complex: Experimental and modeling evidence for a death effector domain chain model. Mol. Cell 2012, 47, 306–319. [Google Scholar] [CrossRef]
- Dickens, L.S.; Boyd, R.S.; Jukes-Jones, R.; Hughes, M.A.; Robinson, G.L.; Fairall, L.; Schwabe, J.W.; Cain, K.; Macfarlane, M. A death effector domain chain DISC model reveals a crucial role for caspase-8 chain assembly in mediating apoptotic cell death. Mol. Cell 2012, 47, 291–305. [Google Scholar] [CrossRef] [PubMed]
- Graves, J.D.; Kordich, J.J.; Huang, T.H.; Piasecki, J.; Bush, T.L.; Sullivan, T.; Foltz, I.N.; Chang, W.; Douangpanya, H.; Dang, T.; et al. Apo2L/TRAIL and the death receptor 5 agonist antibody AMG 655 cooperate to promote receptor clustering and antitumor activity. Cancer Cell 2014, 26, 177–189. [Google Scholar] [CrossRef] [PubMed]
- Tuthill, M.H.; Montinaro, A.; Zinngrebe, J.; Prieske, K.; Draber, P.; Prieske, S.; Newsom-Davis, T.; von Karstedt, S.; Graves, J.; Walczak, H. TRAIL-R2-specific antibodies and recombinant TRAIL can synergise to kill cancer cells. Oncogene 2015, 34, 2138–2144. [Google Scholar] [CrossRef] [PubMed]
- Trauzold, A.; Siegmund, D.; Schniewind, B.; Sipos, B.; Egberts, J.; Zorenkov, D.; Emme, D.; Roder, C.; Kalthoff, H.; Wajant, H. TRAIL promotes metastasis of human pancreatic ductal adenocarcinoma. Oncogene 2006, 25, 7434–7439. [Google Scholar] [CrossRef] [PubMed]
- Hoogwater, F.J.; Nijkamp, M.W.; Smakman, N.; Steller, E.J.; Emmink, B.L.; Westendorp, B.F.; Raats, D.A.; Sprick, M.R.; Schaefer, U.; Van Houdt, W.J.; et al. Oncogenic K-Ras turns death receptors into metastasis-promoting receptors in human and mouse colorectal cancer cells. Gastroenterology 2010, 138, 2357–2367. [Google Scholar] [CrossRef] [PubMed]
- von Karstedt, S.; Conti, A.; Nobis, M.; Montinaro, A.; Hartwig, T.; Lemke, J.; Legler, K.; Annewanter, F.; Campbell, A.D.; Taraborrelli, L.; et al. Cancer cell-autonomous TRAIL-R signaling promotes KRAS-driven cancer progression, invasion, and metastasis. Cancer Cell 2015, 27, 561–573. [Google Scholar] [CrossRef] [PubMed]
- Hartwig, T.; Montinaro, A.; von Karstedt, S.; Sevko, A.; Surinova, S.; Chakravarthy, A.; Taraborrelli, L.; Draber, P.; Lafont, E.; Arce Vargas, F.; et al. The TRAIL-Induced Cancer Secretome Promotes a Tumor-Supportive Immune Microenvironment via CCR2. Mol. Cell 2017, 65, 730–742. [Google Scholar] [CrossRef]
- Ouyang, X.; Shi, M.; Jie, F.; Bai, Y.; Shen, P.; Yu, Z.; Wang, X.; Huang, C.; Tao, M.; Wang, Z.; et al. Phase III study of dulanermin (recombinant human tumor necrosis factor-related apoptosis-inducing ligand/Apo2 ligand) combined with vinorelbine and cisplatin in patients with advanced non-small-cell lung cancer. Investig. New Drugs 2018, 36, 315–322. [Google Scholar] [CrossRef]
- Nair, P.M.; Flores, H.; Gogineni, A.; Marsters, S.; Lawrence, D.A.; Kelley, R.F.; Ngu, H.; Sagolla, M.; Komuves, L.; Bourgon, R.; et al. Enhancing the antitumor efficacy of a cell-surface death ligand by covalent membrane display. Proc. Natl. Acad. Sci. USA 2015, 112, 5679–5684. [Google Scholar] [CrossRef]
- Geng, C.; Hou, J.; Zhao, Y.; Ke, X.; Wang, Z.; Qiu, L.; Xi, H.; Wang, F.; Wei, N.; Liu, Y.; et al. A multicenter, open-label phase II study of recombinant CPT (Circularly Permuted TRAIL) plus thalidomide in patients with relapsed and refractory multiple myeloma. Am. J. Hematol. 2014, 89, 1037–1042. [Google Scholar] [CrossRef]
- Kelley, S.K.; Harris, L.A.; Xie, D.; Deforge, L.; Totpal, K.; Bussiere, J.; Fox, J.A. Preclinical studies to predict the disposition of Apo2L/tumor necrosis factor-related apoptosis-inducing ligand in humans: Characterization of in vivo efficacy, pharmacokinetics, and safety. J. Pharmacol. Exp. Ther. 2001, 299, 31–38. [Google Scholar] [PubMed]
- Chuntharapai, A.; Dodge, K.; Grimmer, K.; Schroeder, K.; Marsters, S.A.; Koeppen, H.; Ashkenazi, A.; Kim, K.J. Isotype-dependent inhibition of tumor growth in vivo by monoclonal antibodies to death receptor 4. J. Immunol. 2001, 166, 4891–4898. [Google Scholar] [CrossRef] [PubMed]
- Natoni, A.; MacFarlane, M.; Inoue, S.; Walewska, R.; Majid, A.; Knee, D.; Stover, D.R.; Dyer, M.J.; Cohen, G.M. TRAIL signals to apoptosis in chronic lymphocytic leukaemia cells primarily through TRAIL-R1 whereas cross-linked agonistic TRAIL-R2 antibodies facilitate signalling via TRAIL-R2. Br. J. Haematol. 2007, 139, 568–577. [Google Scholar] [CrossRef] [PubMed]
- Pukac, L.; Kanakaraj, P.; Humphreys, R.; Alderson, R.; Bloom, M.; Sung, C.; Riccobene, T.; Johnson, R.; Fiscella, M.; Mahoney, A.; et al. HGS-ETR1, a fully human TRAIL-receptor 1 monoclonal antibody, induces cell death in multiple tumour types in vitro and in vivo. Br. J. Cancer 2005, 92, 1430–1441. [Google Scholar] [CrossRef] [PubMed]
- Medler, J.; Nelke, J.; Weisenberger, D.; Steinfatt, T.; Rothaug, M.; Berr, S.; Hunig, T.; Beilhack, A.; Wajant, H. TNFRSF receptor-specific antibody fusion proteins with targeting controlled FcgammaR-independent agonistic activity. Cell Death Dis. 2019, 10, 224. [Google Scholar] [CrossRef] [PubMed]
- Yada, A.; Yazawa, M.; Ishida, S.; Yoshida, H.; Ichikawa, K.; Kurakata, S.; Fujiwara, K. A novel humanized anti-human death receptor 5 antibody CS-1008 induces apoptosis in tumor cells without toxicity in hepatocytes. Ann. Oncol. 2008, 19, 1060–1067. [Google Scholar] [CrossRef] [PubMed]
- Motoki, K.; Mori, E.; Matsumoto, A.; Thomas, M.; Tomura, T.; Humphreys, R.; Albert, V.; Muto, M.; Yoshida, H.; Aoki, M.; et al. Enhanced apoptosis and tumor regression induced by a direct agonist antibody to tumor necrosis factor-related apoptosis-inducing ligand receptor 2. Clin. Cancer Res. 2005, 11, 3126–3135. [Google Scholar] [CrossRef]
- Kaplan-Lefko, P.J.; Graves, J.D.; Zoog, S.J.; Pan, Y.; Wall, J.; Branstetter, D.G.; Moriguchi, J.; Coxon, A.; Huard, J.N.; Xu, R.; et al. Conatumumab, a fully human agonist antibody to death receptor 5, induces apoptosis via caspase activation in multiple tumor types. Cancer Biol. Ther. 2010, 9, 618–631. [Google Scholar] [CrossRef]
- Adams, C.; Totpal, K.; Lawrence, D.; Marsters, S.; Pitti, R.; Yee, S.; Ross, S.; Deforge, L.; Koeppen, H.; Sagolla, M.; et al. Structural and functional analysis of the interaction between the agonistic monoclonal antibody Apomab and the proapoptotic receptor DR5. Cell Death Differ. 2008, 15, 751–761. [Google Scholar] [CrossRef]
- Wilson, N.S.; Yang, B.; Yang, A.; Loeser, S.; Marsters, S.; Lawrence, D.; Li, Y.; Pitti, R.; Totpal, K.; Yee, S.; et al. An Fcgamma receptor-dependent mechanism drives antibody-mediated target-receptor signaling in cancer cells. Cancer Cell 2011, 19, 101–113. [Google Scholar] [CrossRef]
- Zeng, Y.; Wu, X.X.; Fiscella, M.; Shimada, O.; Humphreys, R.; Albert, V.; Kakehi, Y. Monoclonal antibody to tumor necrosis factor-related apoptosis-inducing ligand receptor 2 (TRAIL-R2) induces apoptosis in primary renal cell carcinoma cells in vitro and inhibits tumor growth in vivo. Int. J. Oncol. 2006, 28, 421–430. [Google Scholar] [CrossRef] [PubMed]
- Guo, Y.; Chen, C.; Zheng, Y.; Zhang, J.; Tao, X.; Liu, S.; Zheng, D.; Liu, Y. A novel anti-human DR5 monoclonal antibody with tumoricidal activity induces caspase-dependent and caspase-independent cell death. J. Biol. Chem. 2005, 280, 41940–41952. [Google Scholar] [CrossRef] [PubMed]
- Haynes, N.M.; Hawkins, E.D.; Li, M.; McLaughlin, N.M.; Hammerling, G.J.; Schwendener, R.; Winoto, A.; Wensky, A.; Yagita, H.; Takeda, K.; et al. CD11c+ dendritic cells and B cells contribute to the tumoricidal activity of anti-DR5 antibody therapy in established tumors. J. Immunol. 2010, 185, 532–541. [Google Scholar] [CrossRef] [PubMed]
- Li, F.; Ravetch, J.V. Apoptotic and antitumor activity of death receptor antibodies require inhibitory Fcγ receptor engagement. Proc. Natl. Acad. Sci. USA 2012, 109, 10966–10971. [Google Scholar] [CrossRef] [PubMed]
- Li, F.; Ravetch, J.V. Antitumor activities of agonistic anti-TNFR antibodies require differential FcgammaRIIB coengagement in vivo. Proc. Natl. Acad. Sci. USA 2013, 110, 19501–19506. [Google Scholar] [CrossRef] [PubMed]
- Dobson, C.L.; Main, S.; Newton, P.; Chodorge, M.; Cadwallader, K.; Humphreys, R.; Albert, V.; Vaughan, T.J.; Minter, R.R.; Edwards, B.M. Human monomeric antibody fragments to TRAIL-R1 and TRAIL-R2 that display potent in vitro agonism. MAbs 2009, 1, 552–562. [Google Scholar] [CrossRef] [PubMed]
- Park, K.J.; Lee, S.H.; Kim, T.I.; Lee, H.W.; Lee, C.H.; Kim, E.H.; Jang, J.Y.; Choi, K.S.; Kwon, M.H.; Kim, Y.S. A human scFv antibody against TRAIL receptor 2 induces autophagic cell death in both TRAIL-sensitive and TRAIL-resistant cancer cells. Cancer Res. 2007, 67, 7327–7334. [Google Scholar] [CrossRef]
- Secchiero, P.; Sblattero, D.; Chiaruttini, C.; Melloni, E.; Macor, P.; Zorzet, S.; Tripodo, C.; Tedesco, F.; Marzari, R.; Zauli, G. Selection and characterization of a novel agonistic human recombinant anti-TRAIL-R2 minibody with anti-leukemic activity. Int. J. Immunopathol. Pharmacol. 2009, 22, 73–83. [Google Scholar] [CrossRef]
- Shi, J.; Liu, Y.; Zheng, Y.; Guo, Y.; Zhang, J.; Cheung, P.T.; Xu, R.; Zheng, D. Therapeutic expression of an anti-death receptor 5 single-chain fixed-variable region prevents tumor growth in mice. Cancer Res. 2006, 66, 11946–11953. [Google Scholar] [CrossRef]
- Zauli, G.; Corallini, F.; Zorzet, S.; Grill, V.; Marzari, R.; Secchiero, P. In vivo anti-lymphoma activity of an agonistic human recombinant anti-TRAIL-R2 minibody. Investig. New Drugs 2012, 30, 405–407. [Google Scholar] [CrossRef]
- Tamada, T.; Shinmi, D.; Ikeda, M.; Yonezawa, Y.; Kataoka, S.; Kuroki, R.; Mori, E.; Motoki, K. TRAIL-R2 Superoligomerization Induced by Human Monoclonal Agonistic Antibody KMTR2. Sci. Rep. 2015, 5, 17936. [Google Scholar] [CrossRef] [PubMed]
- Qiu, Y.; Zhang, Z.; Shi, J.; Liu, S.; Liu, Y.; Zheng, D. A novel anti-DR5 chimeric antibody and epirubicin synergistically suppress tumor growth. IUBMB Life 2012, 64, 757–765. [Google Scholar] [CrossRef] [PubMed]
- Wu, G.S.; Burns, T.F.; McDonald, E.R., 3rd; Jiang, W.; Meng, R.; Krantz, I.D.; Kao, G.; Gan, D.D.; Zhou, J.Y.; Muschel, R.; et al. KILLER/DR5 is a DNA damage-inducible p53-regulated death receptor gene. Nat. Genet. 1997, 17, 141–143. [Google Scholar] [CrossRef] [PubMed]
- Papadopoulos, K.P.; Isaacs, R.; Bilic, S.; Kentsch, K.; Huet, H.A.; Hofmann, M.; Rasco, D.; Kundamal, N.; Tang, Z.; Cooksey, J.; et al. Unexpected hepatotoxicity in a phase I study of TAS266, a novel tetravalent agonistic Nanobody(R) targeting the DR5 receptor. Cancer Chemother. Pharmacol. 2015, 75, 887–895. [Google Scholar] [CrossRef] [PubMed]
- Zuch de Zafra, C.L.; Ashkenazi, A.; Darbonne, W.C.; Cheu, M.; Totpal, K.; Ortega, S.; Flores, H.; Walker, M.D.; Kabakoff, B.; Lum, B.L.; et al. Antitherapeutic antibody-mediated hepatotoxicity of recombinant human Apo2L/TRAIL in the cynomolgus monkey. Cell Death Dis. 2016, 7, e2338. [Google Scholar] [CrossRef] [PubMed]
- Wu, X.; Li, P.; Qian, C.; Li, O.; Zhou, Y. Trimeric coiled-coil domain of human pulmonary surfactant protein D enhances zinc-binding ability and biologic activity of soluble TRAIL. Mol. Immunol. 2009, 46, 2381–2388. [Google Scholar] [CrossRef] [PubMed]
- Schneider, B.; Munkel, S.; Krippner-Heidenreich, A.; Grunwald, I.; Wels, W.S.; Wajant, H.; Pfizenmaier, K.; Gerspach, J. Potent antitumoral activity of TRAIL through generation of tumor-targeted single-chain fusion proteins. Cell Death Dis. 2010, 1, e68. [Google Scholar] [CrossRef] [PubMed]
- Walczak, H.; Miller, R.E.; Ariail, K.; Gliniak, B.; Griffith, T.S.; Kubin, M.; Chin, W.; Jones, J.; Woodward, A.; Le, T.; et al. Tumoricidal activity of tumor necrosis factor-related apoptosis-inducing ligand in vivo. Nat. Med. 1999, 5, 157–163. [Google Scholar] [CrossRef] [PubMed]
- Fang, F.; Wang, A.P.; Yang, S.F. Antitumor activity of a novel recombinant mutant human tumor necrosis factor-related apoptosis-inducing ligand. Acta Pharmacol. Sin. 2005, 26, 1373–1381. [Google Scholar] [CrossRef]
- Yan, J.; Wang, L.; Wang, Z.; Wang, Z.; Wang, B.; Zhu, R.; Bi, J.; Wu, J.; Zhang, H.; Wu, H.; et al. Engineered adenovirus fiber shaft fusion homotrimer of soluble TRAIL with enhanced stability and antitumor activity. Cell Death Dis. 2016, 7, e2274. [Google Scholar] [CrossRef]
- Melendez, M.E.; Silva-Oliveira, R.J.; Silva Almeida Vicente, A.L.; Rebolho Batista Arantes, L.M.; Carolina de Carvalho, A.; Epstein, A.L.; Reis, R.M.; Carvalho, A.L. Construction and characterization of a new TRAIL soluble form, active at picomolar concentrations. Oncotarget 2018, 9, 27233–27241. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Muller, N.; Schneider, B.; Pfizenmaier, K.; Wajant, H. Superior serum half life of albumin tagged TNF ligands. Biochem. Biophys. Res. Commun. 2010, 396, 793–799. [Google Scholar] [CrossRef] [PubMed]
- Holler, N.; Tardivel, A.; Kovacsovics-Bankowski, M.; Hertig, S.; Gaide, O.; Martinon, F.; Tinel, A.; Deperthes, D.; Calderara, S.; Schulthess, T.; et al. Two adjacent trimeric Fas ligands are required for Fas signaling and formation of a death-inducing signaling complex. Mol. Cell. Biol. 2003, 23, 1428–1440. [Google Scholar] [CrossRef] [PubMed]
- Bossen, C.; Ingold, K.; Tardivel, A.; Bodmer, J.L.; Gaide, O.; Hertig, S.; Ambrose, C.; Tschopp, J.; Schneider, P. Interactions of tumor necrosis factor (TNF) and TNF receptor family members in the mouse and human. J. Biol. Chem. 2006, 281, 13964–13971. [Google Scholar] [CrossRef] [PubMed]
- Gieffers, C.; Kluge, M.; Merz, C.; Sykora, J.; Thiemann, M.; Schaal, R.; Fischer, C.; Branschadel, M.; Abhari, B.A.; Hohenberger, P.; et al. APG350 induces superior clustering of TRAIL receptors and shows therapeutic antitumor efficacy independent of cross-linking via Fcgamma receptors. Mol. Cancer Ther. 2013, 12, 2735–2747. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.; Davis, J.S.; Wu, X. Immunoglobulin Fc domain fusion to TRAIL significantly prolongs its plasma half-life and enhances its antitumor activity. Mol. Cancer Ther. 2014, 13, 643–650. [Google Scholar] [CrossRef] [PubMed]
- Brin, E.; Wu, K.; Dagostino, E.; Meng-Chiang Kuo, M.; He, Y.; Shia, W.J.; Chen, L.C.; Stempniak, M.; Hickey, R.; Almassy, R.; et al. TRAIL stabilization and cancer cell sensitization to its pro-apoptotic activity achieved through genetic fusion with arginine deiminase. Oncotarget 2018, 9, 36914–36928. [Google Scholar] [CrossRef]
- Hutt, M.; Marquardt, L.; Seifert, O.; Siegemund, M.; Muller, I.; Kulms, D.; Pfizenmaier, K.; Kontermann, R.E. Superior Properties of Fc-comprising scTRAIL Fusion Proteins. Mol. Cancer Ther. 2017, 16, 2792–2802. [Google Scholar] [CrossRef]
- Seifert, O.; Plappert, A.; Fellermeier, S.; Siegemund, M.; Pfizenmaier, K.; Kontermann, R.E. Tetravalent antibody-scTRAIL fusion proteins with improved properties. Mol. Cancer Ther. 2014, 13, 101–111. [Google Scholar] [CrossRef]
- Siegemund, M.; Schneider, F.; Hutt, M.; Seifert, O.; Muller, I.; Kulms, D.; Pfizenmaier, K.; Kontermann, R.E. IgG-single-chain TRAIL fusion proteins for tumour therapy. Sci. Rep. 2018, 8, 7808. [Google Scholar] [CrossRef]
- Madhumathi, J.; Sridevi, S.; Verma, R.S. Novel TNF-related Apoptotic-inducing Ligand-based Immunotoxin for Therapeutic Targeting of CD25 Positive Leukemia. Target. Oncol. 2016, 11, 535–547. [Google Scholar] [CrossRef] [PubMed]
- Liu, R.; Ma, X.; Wang, H.; Xi, Y.; Qian, M.; Yang, W.; Luo, D.; Fan, L.; Xia, X.; Zhou, J.; et al. The novel fusion protein sTRAIL-TMTP1 exhibits a targeted inhibition of primary tumors and metastases. J. Mol. Med. (Berl.) 2014, 92, 165–175. [Google Scholar] [CrossRef] [PubMed]
- Bremer, E.; Kuijlen, J.; Samplonius, D.; Walczak, H.; de Leij, L.; Helfrich, W. Target cell-restricted and -enhanced apoptosis induction by a scFv:sTRAIL fusion protein with specificity for the pancarcinoma-associated antigen EGP2. Int. J. Cancer 2004, 109, 281–290. [Google Scholar] [CrossRef] [PubMed]
- Bremer, E.; Samplonius, D.F.; van Genne, L.; Dijkstra, M.H.; Kroesen, B.J.; de Leij, L.F.; Helfrich, W. Simultaneous inhibition of epidermal growth factor receptor (EGFR) signaling and enhanced activation of tumor necrosis factor-related apoptosis-inducing ligand (TRAIL) receptor-mediated apoptosis induction by an scFv:sTRAIL fusion protein with specificity for human EGFR. J. Biol. Chem. 2005, 280, 10025–10033. [Google Scholar] [CrossRef] [PubMed]
- Bremer, E.; Samplonius, D.F.; Peipp, M.; van Genne, L.; Kroesen, B.J.; Fey, G.H.; Gramatzki, M.; de Leij, L.F.; Helfrich, W. Target cell-restricted apoptosis induction of acute leukemic T cells by a recombinant tumor necrosis factor-related apoptosis-inducing ligand fusion protein with specificity for human CD7. Cancer Res. 2005, 65, 3380–3388. [Google Scholar] [CrossRef] [PubMed]
- Bremer, E.; de Bruyn, M.; Samplonius, D.F.; Bijma, T.; ten Cate, B.; de Leij, L.F.; Helfrich, W. Targeted delivery of a designed sTRAIL mutant results in superior apoptotic activity towards EGFR-positive tumor cells. J. Mol. Med. (Berl.) 2008, 86, 909–924. [Google Scholar] [CrossRef] [PubMed]
- Stieglmaier, J.; Bremer, E.; Kellner, C.; Liebig, T.M.; ten Cate, B.; Peipp, M.; Schulze-Koops, H.; Pfeiffer, M.; Buhring, H.J.; Greil, J.; et al. Selective induction of apoptosis in leukemic B-lymphoid cells by a CD19-specific TRAIL fusion protein. Cancer Immunol. Immunother. 2008, 57, 233–246. [Google Scholar] [CrossRef]
- ten Cate, B.; Bremer, E.; de Bruyn, M.; Bijma, T.; Samplonius, D.; Schwemmlein, M.; Huls, G.; Fey, G.; Helfrich, W. A novel AML-selective TRAIL fusion protein that is superior to Gemtuzumab Ozogamicin in terms of in vitro selectivity, activity and stability. Leukemia 2009, 23, 1389–1397. [Google Scholar] [CrossRef]
- de Bruyn, M.; Rybczynska, A.A.; Wei, Y.; Schwenkert, M.; Fey, G.H.; Dierckx, R.A.; van Waarde, A.; Helfrich, W.; Bremer, E. Melanoma-associated Chondroitin Sulfate Proteoglycan (MCSP)-targeted delivery of soluble TRAIL potently inhibits melanoma outgrowth in vitro and in vivo. Mol. Cancer 2010, 9, 301. [Google Scholar] [CrossRef]
- de Bruyn, M.; Wei, Y.; Wiersma, V.R.; Samplonius, D.F.; Klip, H.G.; van der Zee, A.G.; Yang, B.; Helfrich, W.; Bremer, E. Cell surface delivery of TRAIL strongly augments the tumoricidal activity of T cells. Clin. Cancer Res. 2011, 17, 5626–5637. [Google Scholar] [CrossRef]
- Hartung, F.; Stuhmer, W.; Pardo, L.A. Tumor cell-selective apoptosis induction through targeting of K(V)10.1 via bifunctional TRAIL antibody. Mol. Cancer 2011, 10, 109. [Google Scholar] [CrossRef] [PubMed]
- Siegemund, M.; Pollak, N.; Seifert, O.; Wahl, K.; Hanak, K.; Vogel, A.; Nussler, A.K.; Gottsch, D.; Munkel, S.; Bantel, H.; et al. Superior antitumoral activity of dimerized targeted single-chain TRAIL fusion proteins under retention of tumor selectivity. Cell Death Dis. 2012, 3, e295. [Google Scholar] [CrossRef] [PubMed]
- El-Mesery, M.; Trebing, J.; Schafer, V.; Weisenberger, D.; Siegmund, D.; Wajant, H. CD40-directed scFv-TRAIL fusion proteins induce CD40-restricted tumor cell death and activate dendritic cells. Cell Death Dis. 2013, 4, e916. [Google Scholar] [CrossRef] [PubMed]
- Yan, C.; Li, S.; Li, Z.; Peng, H.; Yuan, X.; Jiang, L.; Zhang, Y.; Fan, D.; Hu, X.; Yang, M.; et al. Human umbilical cord mesenchymal stem cells as vehicles of CD20-specific TRAIL fusion protein delivery: A double-target therapy against non-Hodgkin’s lymphoma. Mol. Pharm. 2013, 10, 142–151. [Google Scholar] [CrossRef] [PubMed]
- Kouno, M.; Lin, C.; Schechter, N.M.; Siegel, D.; Yang, X.; Seykora, J.T.; Stanley, J.R. Targeted delivery of tumor necrosis factor-related apoptosis-inducing ligand to keratinocytes with a pemphigus mAb. J. Investig. Dermatol. 2013, 133, 2212–2220. [Google Scholar] [CrossRef] [PubMed]
- Trebing, J.; El-Mesery, M.; Schafer, V.; Weisenberger, D.; Siegmund, D.; Silence, K.; Wajant, H. CD70-restricted specific activation of TRAILR1 or TRAILR2 using scFv-targeted TRAIL mutants. Cell Death Dis. 2014, 5, e1035. [Google Scholar] [CrossRef]
- Wiersma, V.R.; He, Y.; Samplonius, D.F.; van Ginkel, R.J.; Gerssen, J.; Eggleton, P.; Zhou, J.; Bremer, E.; Helfrich, W. A CD47-blocking TRAIL fusion protein with dual pro-phagocytic and pro-apoptotic anticancer activity. Br. J. Haematol. 2014, 164, 304–307. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.H.; Ni, C.W.; Lin, Y.Z.; Yin, L.; Jiang, C.B.; Lv, C.T.; Le, Y.; Lang, Y.; Zhao, C.Y.; Yang, K.; et al. Targeted induction of apoptosis in glioblastoma multiforme cells by an MRP3-specific TRAIL fusion protein in vitro. Tumour Biol. 2014, 35, 1157–1168. [Google Scholar] [CrossRef]
- Wiersma, V.R.; de Bruyn, M.; Shi, C.; Gooden, M.J.; Wouters, M.C.; Samplonius, D.F.; Hendriks, D.; Nijman, H.W.; Wei, Y.; Zhou, J.; et al. C-type lectin-like molecule-1 (CLL1)-targeted TRAIL augments the tumoricidal activity of granulocytes and potentiates therapeutic antibody-dependent cell-mediated cytotoxicity. MAbs 2015, 7, 321–330. [Google Scholar] [CrossRef]
- Hendriks, D.; He, Y.; Koopmans, I.; Wiersma, V.R.; van Ginkel, R.J.; Samplonius, D.F.; Helfrich, W.; Bremer, E. Programmed Death Ligand 1 (PD-L1)-targeted TRAIL combines PD-L1-mediated checkpoint inhibition with TRAIL-mediated apoptosis induction. Oncoimmunology 2016, 5, e1202390. [Google Scholar] [CrossRef]
- Assohou-Luty, C.; Gerspach, J.; Siegmund, D.; Muller, N.; Huard, B.; Tiegs, G.; Pfizenmaier, K.; Wajant, H. A CD40-CD95L fusion protein interferes with CD40L-induced prosurvival signaling and allows membrane CD40L-restricted activation of CD95. J. Mol. Med. (Berl.) 2006, 84, 785–797. [Google Scholar] [CrossRef] [PubMed]
- Cao, L.; Du, P.; Jiang, S.H.; Jin, G.H.; Huang, Q.L.; Hua, Z.C. Enhancement of antitumor properties of TRAIL by targeted delivery to the tumor neovasculature. Mol. Cancer Ther. 2008, 7, 851–861. [Google Scholar] [CrossRef] [PubMed]
- Garg, G.; Gibbs, J.; Belt, B.; Powell, M.A.; Mutch, D.G.; Goedegebuure, P.; Collins, L.; Piwnica-Worms, D.; Hawkins, W.G.; Spitzer, D. Novel treatment option for MUC16-positive malignancies with the targeted TRAIL-based fusion protein Meso-TR3. BMC Cancer 2014, 14, 35. [Google Scholar] [CrossRef] [PubMed]
- Su, Y.; Tatzel, K.; Wang, X.; Belt, B.; Binder, P.; Kuroki, L.; Powell, M.A.; Mutch, D.G.; Hawkins, W.G.; Spitzer, D. Mesothelin’s minimal MUC16 binding moiety converts TR3 into a potent cancer therapeutic via hierarchical binding events at the plasma membrane. Oncotarget 2016, 7, 31534–31549. [Google Scholar] [CrossRef] [PubMed]
- Uckun, F.M.; Myers, D.E.; Qazi, S.; Ozer, Z.; Rose, R.; D’Cruz, O.J.; Ma, H. Recombinant human CD19L-sTRAIL effectively targets B cell precursor acute lymphoblastic leukemia. J. Clin. Investig. 2015, 125, 1006–1018. [Google Scholar] [CrossRef] [PubMed]
- Zhu, Y.; Bassoff, N.; Reinshagen, C.; Bhere, D.; Nowicki, M.O.; Lawler, S.E.; Roux, J.; Shah, K. Bi-specific molecule against EGFR and death receptors simultaneously targets proliferation and death pathways in tumors. Sci. Rep. 2017, 7, 2602. [Google Scholar] [CrossRef] [PubMed]
- Tao, Z.; Yang, H.; Shi, Q.; Fan, Q.; Wan, L.; Lu, X. Targeted Delivery to Tumor-associated Pericytes via an Affibody with High Affinity for PDGFRbeta Enhances the in vivo Antitumor Effects of Human TRAIL. Theranostics 2017, 7, 2261–2276. [Google Scholar] [CrossRef]
- Prigozhina, T.B.; Szafer, F.; Aronin, A.; Tzdaka, K.; Amsili, S.; Makdasi, E.; Shani, N.; Dranitzki Elhalel, M. Fn14.TRAIL fusion protein is oligomerized by TWEAK into a superefficient TRAIL analog. Cancer Lett. 2017, 400, 99–109. [Google Scholar] [CrossRef]
- Wajant, H. The TWEAK-Fn14 system as a potential drug target. Br. J. Pharmacol. 2013, 170, 748–764. [Google Scholar] [CrossRef]
- Bremer, E.; Samplonius, D.; Kroesen, B.J.; van Genne, L.; de Leij, L.; Helfrich, W. Exceptionally potent anti-tumor bystander activity of an scFv:sTRAIL fusion protein with specificity for EGP2 toward target antigen-negative tumor cells. Neoplasia 2004, 6, 636–645. [Google Scholar] [CrossRef]
- Allen, J.E.; Kline, C.L.; Prabhu, V.V.; Wagner, J.; Ishizawa, J.; Madhukar, N.; Lev, A.; Baumeister, M.; Zhou, L.; Lulla, A.; et al. Discovery and clinical introduction of first-in-class imipridone ONC201. Oncotarget 2016, 7, 74380–74392. [Google Scholar] [CrossRef] [PubMed]
- Lee, S.H.; Park, D.W.; Sung, E.S.; Park, H.R.; Kim, J.K.; Kim, Y.S. Humanization of an agonistic anti-death receptor 4 single chain variable fragment antibody and avidity-mediated enhancement of its cell death-inducing activity. Mol. Immunol. 2010, 47, 816–824. [Google Scholar] [CrossRef] [PubMed]
- Huet, H.A.; Growney, J.D.; Johnson, J.A.; Li, J.; Bilic, S.; Ostrom, L.; Zafari, M.; Kowal, C.; Yang, G.; Royo, A.; et al. Multivalent nanobodies targeting death receptor 5 elicit superior tumor cell killing through efficient caspase induction. MAbs 2014, 6, 1560–1570. [Google Scholar] [CrossRef] [PubMed]
- Liu, F.; Si, Y.; Liu, G.; Li, S.; Zhang, J.; Ma, Y. The tetravalent anti-DR5 antibody without cross-linking direct induces apoptosis of cancer cells. Biomed. Pharmacother. 2015, 70, 41–45. [Google Scholar] [CrossRef] [PubMed]
- Clore, G.M.; Omichinski, J.G.; Sakaguchi, K.; Zambrano, N.; Sakamoto, H.; Appella, E.; Gronenborn, A.M. High-resolution structure of the oligomerization domain of p53 by multidimensional NMR. Science 1994, 265, 386–391. [Google Scholar] [CrossRef] [PubMed]
- Swers, J.S.; Grinberg, L.; Wang, L.; Feng, H.; Lekstrom, K.; Carrasco, R.; Xiao, Z.; Inigo, I.; Leow, C.C.; Wu, H.; et al. Multivalent scaffold proteins as superagonists of TRAIL receptor 2-induced apoptosis. Mol. Cancer Ther. 2013, 12, 1235–1244. [Google Scholar] [CrossRef] [PubMed]
- Greer, Y.E.; Gilbert, S.F.; Gril, B.; Narwal, R.; Peacock Brooks, D.L.; Tice, D.A.; Steeg, P.S.; Lipkowitz, S. MEDI3039, a novel highly potent tumor necrosis factor (TNF)-related apoptosis-inducing ligand (TRAIL) receptor 2 agonist, causes regression of orthotopic tumors and inhibits outgrowth of metastatic triple-negative breast cancer. Breast Cancer Res. 2019, 21, 27. [Google Scholar] [CrossRef]
- Valldorf, B.; Fittler, H.; Deweid, L.; Ebenig, A.; Dickgiesser, S.; Sellmann, C.; Becker, J.; Zielonka, S.; Empting, M.; Avrutina, O.; et al. An Apoptosis-Inducing Peptidic Heptad That Efficiently Clusters Death Receptor 5. Angew. Chem. Int. Ed. Engl. 2016, 55, 5085–5089. [Google Scholar] [CrossRef]
- Michaelson, J.S.; Demarest, S.J.; Miller, B.; Amatucci, A.; Snyder, W.B.; Wu, X.; Huang, F.; Phan, S.; Gao, S.; Doern, A.; et al. Anti-tumor activity of stability-engineered IgG-like bispecific antibodies targeting TRAIL-R2 and LTbetaR. MAbs 2009, 1, 128–141. [Google Scholar] [CrossRef]
- He, Y.; Hendriks, D.; van Ginkel, R.; Samplonius, D.; Bremer, E.; Helfrich, W. Melanoma-Directed Activation of Apoptosis Using a Bispecific Antibody Directed at MCSP and TRAIL Receptor-2/Death Receptor-5. J. Investig. Dermatol. 2016, 136, 541–544. [Google Scholar] [CrossRef]
- Shivange, G.; Urbanek, K.; Przanowski, P.; Perry, J.S.A.; Jones, J.; Haggart, R.; Kostka, C.; Patki, T.; Stelow, E.; Petrova, Y.; et al. A Single-Agent Dual-Specificity Targeting of FOLR1 and DR5 as an Effective Strategy for Ovarian Cancer. Cancer Cell 2018, 34, 331–345. [Google Scholar] [CrossRef] [PubMed]
- Brunker, P.; Wartha, K.; Friess, T.; Grau-Richards, S.; Waldhauer, I.; Koller, C.F.; Weiser, B.; Majety, M.; Runza, V.; Niu, H.; et al. RG7386, a Novel Tetravalent FAP-DR5 Antibody, Effectively Triggers FAP-Dependent, Avidity-Driven DR5 Hyperclustering and Tumor Cell Apoptosis. Mol. Cancer Ther. 2016, 15, 946–957. [Google Scholar] [CrossRef] [PubMed]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Disease | Soluble TRAIL Level | Correlation with Clinical Relevant Factors | Reference |
|---|---|---|---|
| Hepatitis B virus infection | Increased | Yes, liver damage | [7] |
| Systemic lupus erythematosus | Increased | No | [8] |
| Amyotrophic lateral sclerosis | Reduced | No | [9] |
| Chronic kidney disease | Reduced | Yes, inverse association with mortality risk | [10] |
| Systemic sclerosis | Increased | Yes, associated with pulmonary involvement | [11] |
| Recurrent miscarriage | Increased | Not investigated | [12] |
| Preeclampsia 1 | Decreased | Not investigated | [13] |
| Multiple sclerosis | Reduced | No | [14] |
| Type I diabetes | Reduced | Not investigated | [15] |
| Hypercholesterolemia | Increase | Yes, low-density lipoprotein | [16] |
| Polymyositis and Dermatomyocitie | Increased | Yes, disease activity | [17] |
| Name | Isotype | Increase of Agonistic Activity | Reference | |
|---|---|---|---|---|
| by Crosslinking 2 | by FcγR-Binding 3 | |||
| TRAILR1/DR4 | ||||
| 4H6 | mIgG1 | >1000 | [72] | |
| 4G7 | mIgG2a | >1000 | [72] | |
| DR4-A | >100 | [73] | ||
| Mapatumumab (HGS-ETR1) | hIgG1 | Highly active w/o crosslinking | [74] | |
| DJR1 | mIgG1 | Considerable | [75] | |
| HS101 | mIgG1 | Considerable | [75] | |
| TRAILR2/DR5 | ||||
| CS-1008 (from TRA-8, Tigatuzumab) | hIgG1 | 10 to >1000 | [76] | |
| KMTR2 | hIgG1 | ~3; already highly active w/o crosslinking | [77] | |
| LBY135 | hIgG1 | >100 | [73] | |
| Conatumumab (AMG655) | hIgG1 | 100 to >1000 | [78] | |
| Drozitumab Apomab | hIgG1 | >100 | Considerable | [79,80] |
| LexatumumabHGS-ETR2 | hIgG1 | >10 | [81] | |
| Zaptuzumab (AD5.10) | hIgG1 | Highly active w/o crosslinking | [82] | |
| DJR2 | mIgG1 | Considerable | [75] | |
| D-6 | mIgG1 | Considerable | [75] | |
| Anti-mDR5 (MD5-1) | Hamster IgG | Considerable | [83,84,85] | |
| Name | Stabilization Strategy | Activity Increase 1 | Oligomerization Effect 2 | Reference |
|---|---|---|---|---|
| LZ-TRAIL | N-terminal leucine zipper | 3 to >100 (cell-type dependent) | n.d. 3 | [98] |
| CPT | 135-281-L-121-134 | 20–60 | n.d. | [60,99] |
| TNC-TRAIL | N-terminal tenascin-C trimerization domain | 5–10 | 250–1000 | [2] |
| ST | Coiled-coil of SP-D | ~10 | n.d. | [96] |
| scTRAIL | Peptide linker connected TRAIL protomers | scTRAIL has only been published as part of scTRAIL fusion proteins | [97] | |
| HA5FT | Ad5 fiber | 3–5 | n.d. | [100] |
| HA5ST | Ad5 shaft | 3–5 | ||
| sfTRAIL | N-terminal foldon domain 4 | n.d. | n.d. | [101] |
| Name | Oligomerization Strategy | Number of TRAIL Domains 1 | Activity Increase 2 | Reference |
|---|---|---|---|---|
| APG350 3 | C-terminal IgG1 Fc domain | 2 | >100 | [105] |
| Fc-scTRAIL | N-terminal IgG1 Fc domain | 2 | 10 | [108] |
| Db-scTRAIL | 2 | 10–30 | [108] | |
| EDH2-scTRAIL | N-terminal heavy chain domain 2 of IgE | 2 | 10 | [108,109] |
| LC-scTRAIL | Fused to light chain of IgG1 | 2 | n.d. 4 | [110] |
| HC-scTRAIL | Fused to heavy chain of IgG1 | 2 | n.d. 4 | [110] |
| LC/HC-scTRAIL | Fused to light and heavy chain of IgG1 | 4 | n.d. 4 | [110] |
| Adi-TRAIL | Fusion to arginine deiminase | 2 | n.d. 4 | [111] |
| Anchoring Domain | Anchor Target | Activity Increase 1 | Effect of Anchoring Domain | Reference |
|---|---|---|---|---|
| scFv:FAP | FAP | ~20 | [3] | |
| scFv:C54 | EpCAM (EGP2) | >100 | [113] | |
| scFv:425 | EGFR | - 2 | [114] | |
| scFv:CD7 | CD7 | >100 | [115] | |
| scFv:425 | EGFR | ~50 3 | [116] | |
| scFv:CD19 | CD19 | - 2 | [117] | |
| scFv:CD33 | CD33 | >50 | [118] | |
| scFv:MCSP | MCSP | >100 | [119] | |
| scFv:ErbB2 | ErbB2 4 | ~10 | [97] | |
| K12 | CD7 5 | >>100 | [120] | |
| svFv:CD3 | CD3 5 | >>100 | [120] | |
| scFv:62 | Kv10.1 | [121] | ||
| scFv:hu225 | EGFR 4 | ~10 | [122] | |
| scFv:G28 | CD40 | ~100 | Activates CD40 and thus stimulates DC maturation. | [123] |
| scFv:CD20 | CD20 | - 2 | [124] | |
| scFv:Px44 | DSG | [125] | ||
| scFv:CD70 | CD70 6 | 10–100 | Inhibition of CD70–CD27 interaction. | [126] |
| scFv:CD47 | CD47 | >50 | Blocks CD47-SIRPα interaction and abrogates inhibition phagocytosis. | [127] |
| scFv:M58 | MRP-3 | [106,128] | ||
| scFv:hu225-EHD | EGFR | 10–20 | [106] | |
| scFv:CLL-1 | CLL1 5 | >>100 | [129] | |
| scFv:PD-L1 | PD1 | >100 | Blocks PD1–PD–L1 interaction. | [130] |
| scFv-EHD | EGFR | 3–5 | [108] | |
| CD40ed | memCD40L | >100 | Blocks antiapoptotic CD40L signaling. | [131] |
| RGD | αVβ3, αVβ5 | - 2 | [132] | |
| Mesothelin | Muc16 | >10 | [133] | |
| Meso(1-64) | Muc16 | >10 | [134] | |
| CD19L | CD19 | - 2 | [135] | |
| ENb | EGFR | - 2 | Blocks EGFR signaling. | [136] |
| Z | PDGFRß | ~4 | [137] |
© 2019 by the author. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wajant, H. Molecular Mode of Action of TRAIL Receptor Agonists—Common Principles and Their Translational Exploitation. Cancers 2019, 11, 954. https://doi.org/10.3390/cancers11070954
Wajant H. Molecular Mode of Action of TRAIL Receptor Agonists—Common Principles and Their Translational Exploitation. Cancers. 2019; 11(7):954. https://doi.org/10.3390/cancers11070954
Chicago/Turabian StyleWajant, Harald. 2019. "Molecular Mode of Action of TRAIL Receptor Agonists—Common Principles and Their Translational Exploitation" Cancers 11, no. 7: 954. https://doi.org/10.3390/cancers11070954
APA StyleWajant, H. (2019). Molecular Mode of Action of TRAIL Receptor Agonists—Common Principles and Their Translational Exploitation. Cancers, 11(7), 954. https://doi.org/10.3390/cancers11070954