Deletions of Chromosome 7q Affect Nuclear Organization and HLXB9Gene Expression in Hematological Disorders

 ,
,  , , ,
, , ,  and
and 
Abstract
1. Introduction
2. Results
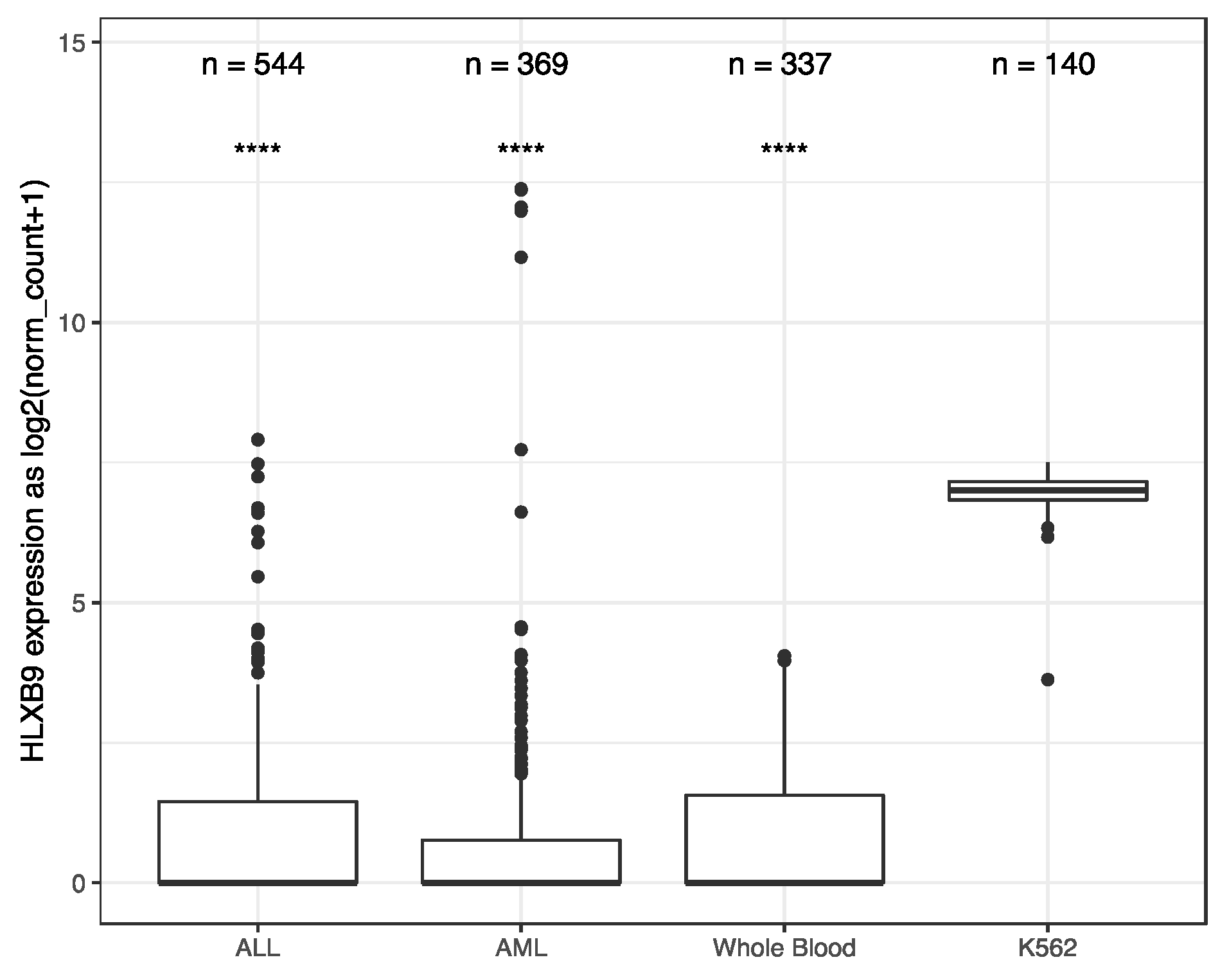
2.1. Data Mining of HLXB9 Expression
2.2. Classification of Large Series of Patients Based on HLXB9Expression Patterns
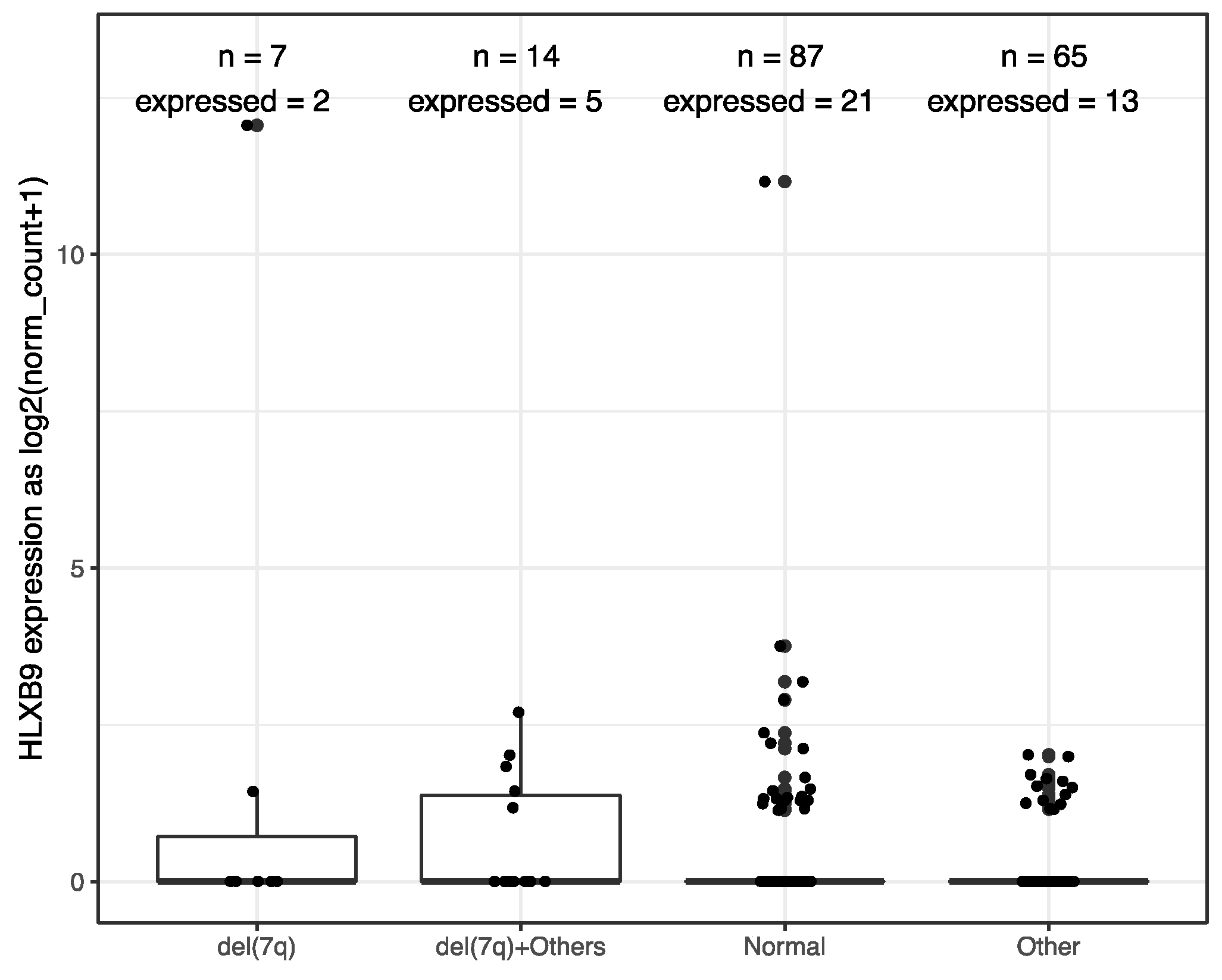
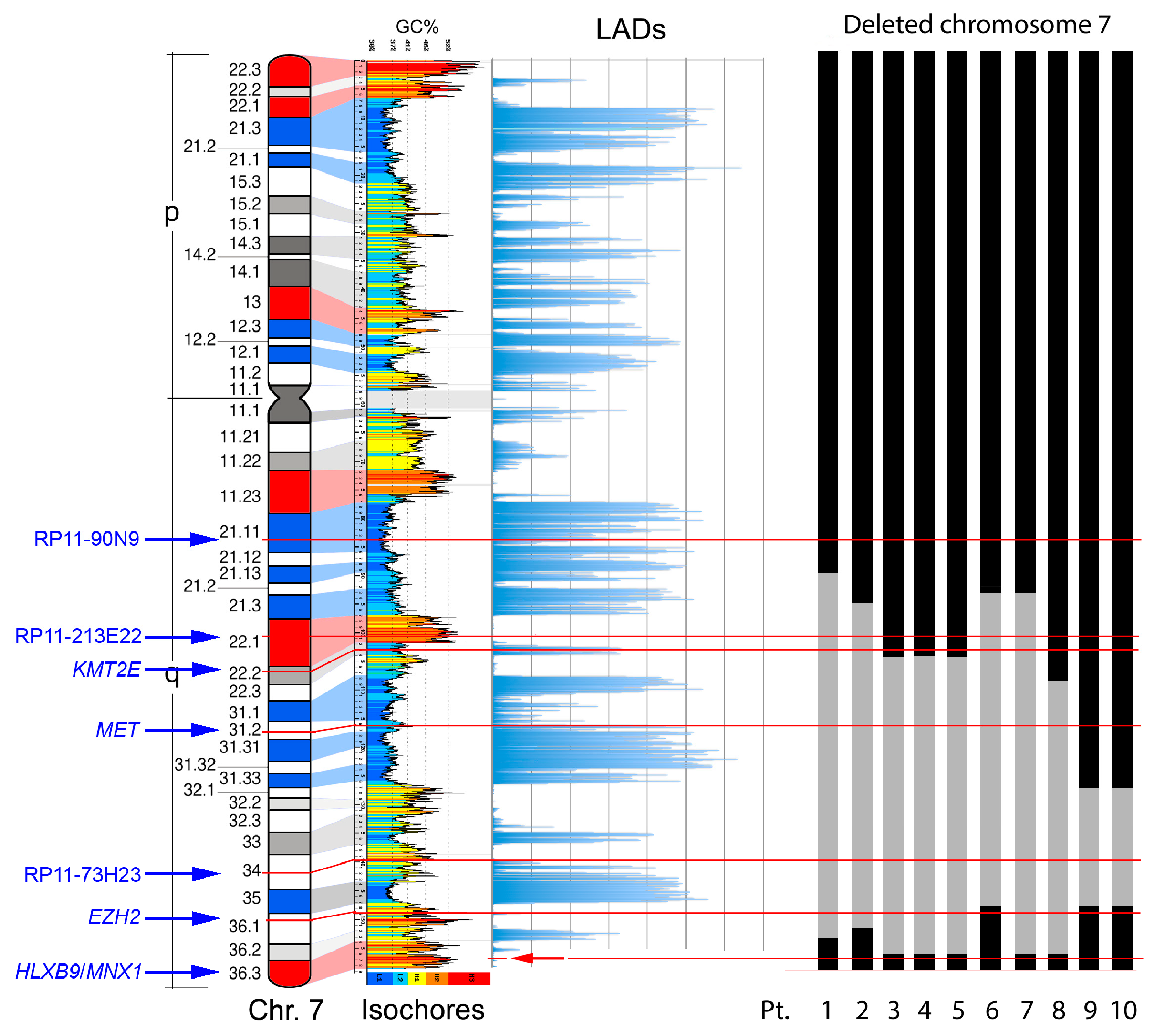
2.3. Deletion Mapping of Breakpoints, Radial Nuclear Location and Expression Analyzes in a Selected Series of Patients
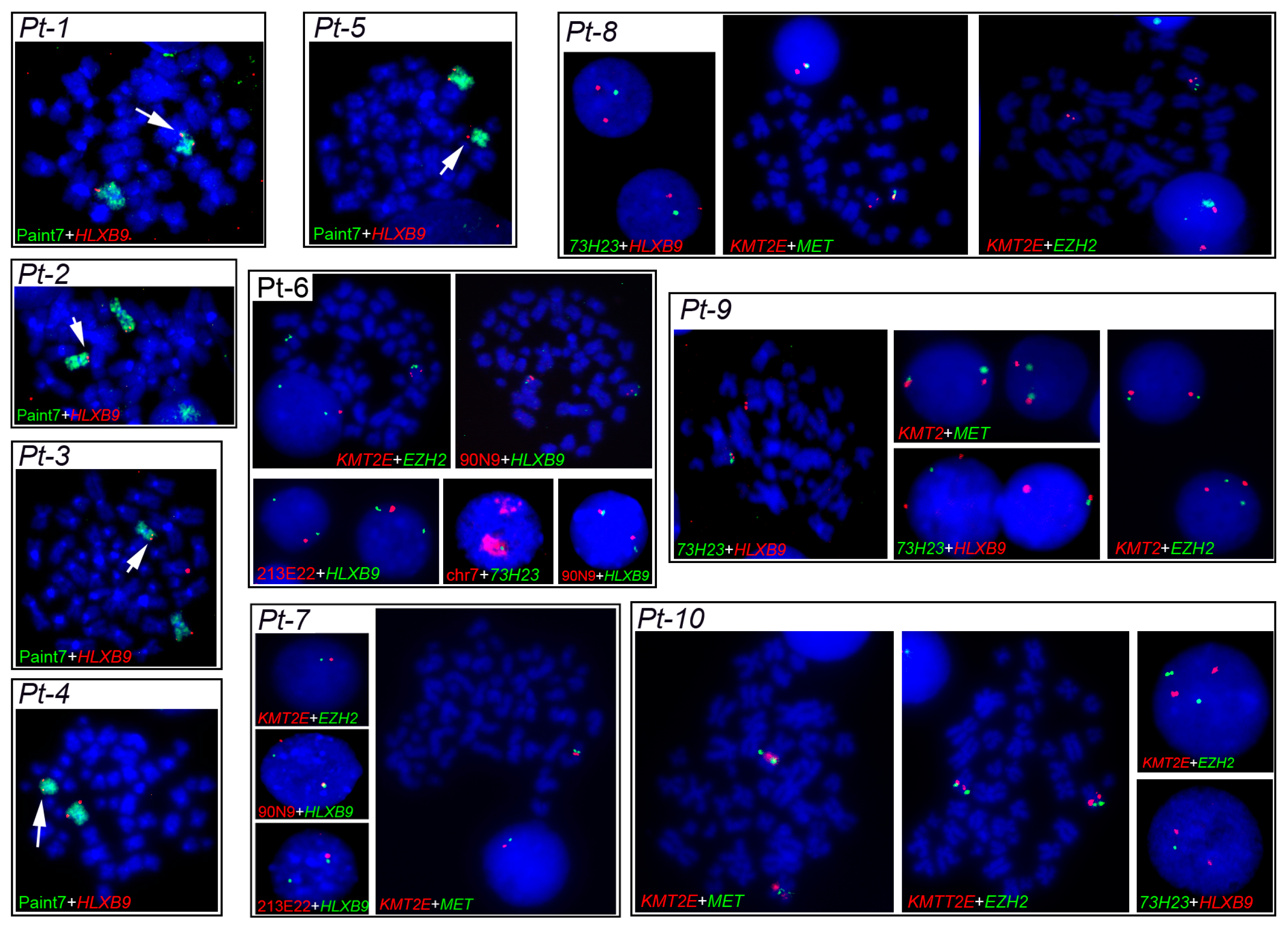
2.3.1. Characterization of del(7q) Breakpoints
2.3.2. HLXB9 Radial Nuclear Location
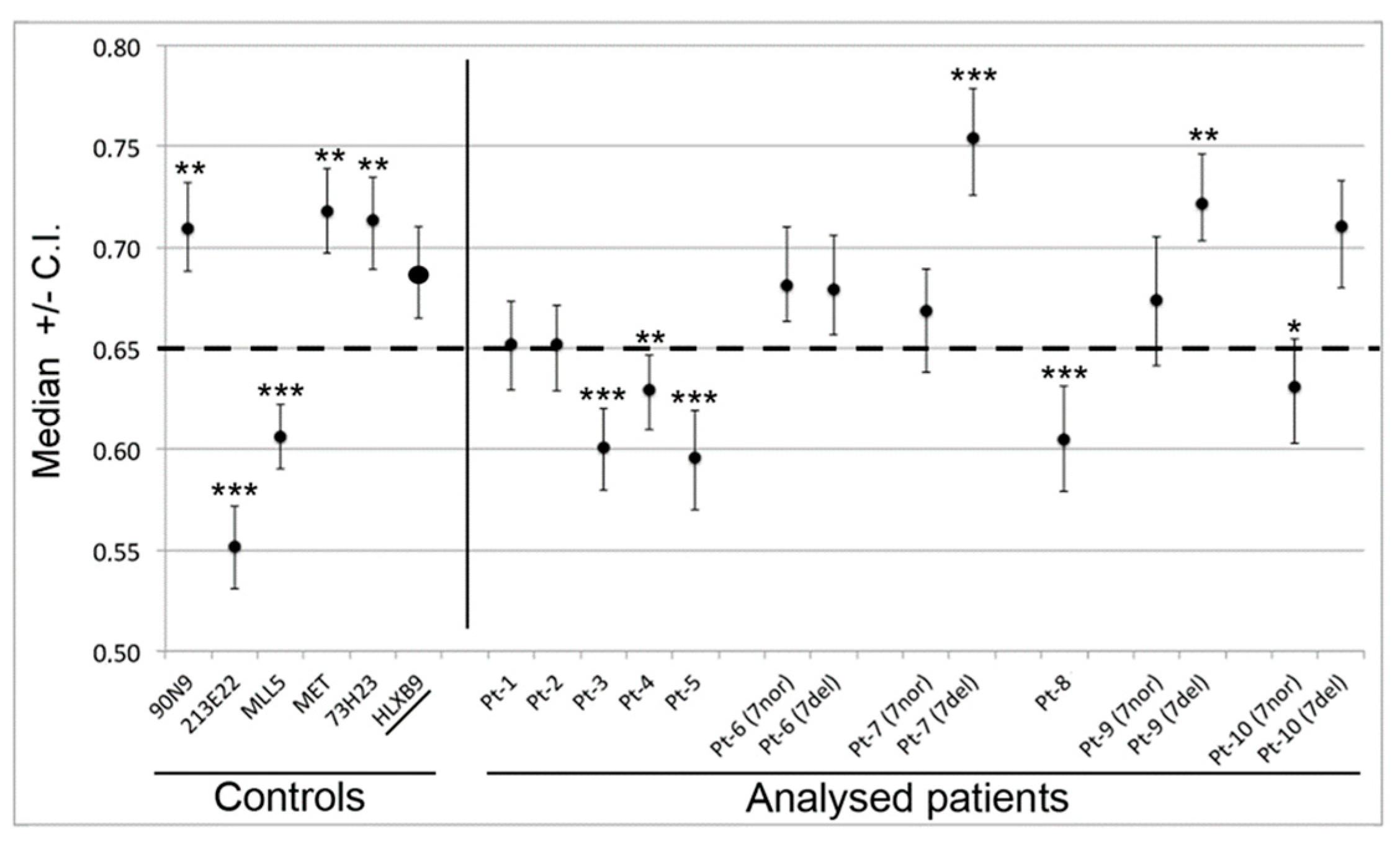
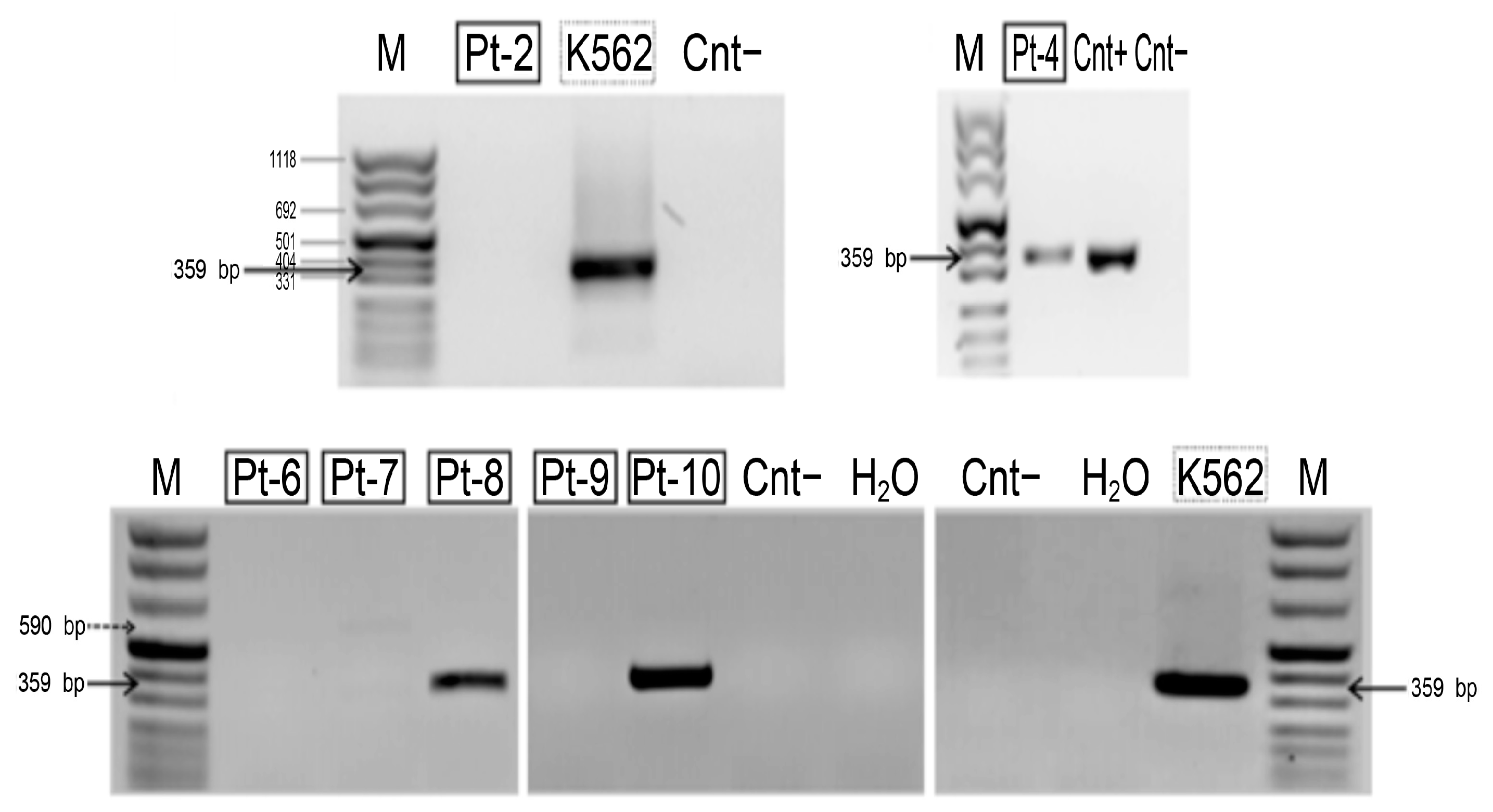
2.3.3. HLXB9Expression
3. Discussion
3.1. HLXB9 Expression is Not Associated with Any Specific Leukemia Subtype
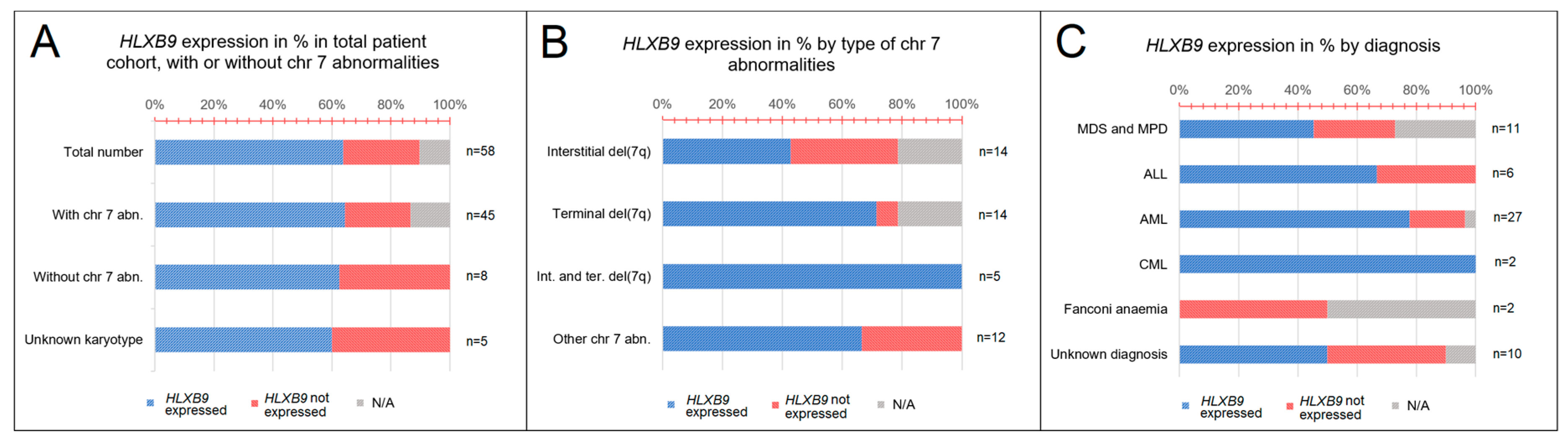
3.2. HLXB9 Expression is Not Necessarily Associated with the Presence of Chromosome 7 Abnormalities
3.3. HLXB9 is Expressed When del(7q) Proximal Breakpoint Lies in a GC-rich Genomic Region
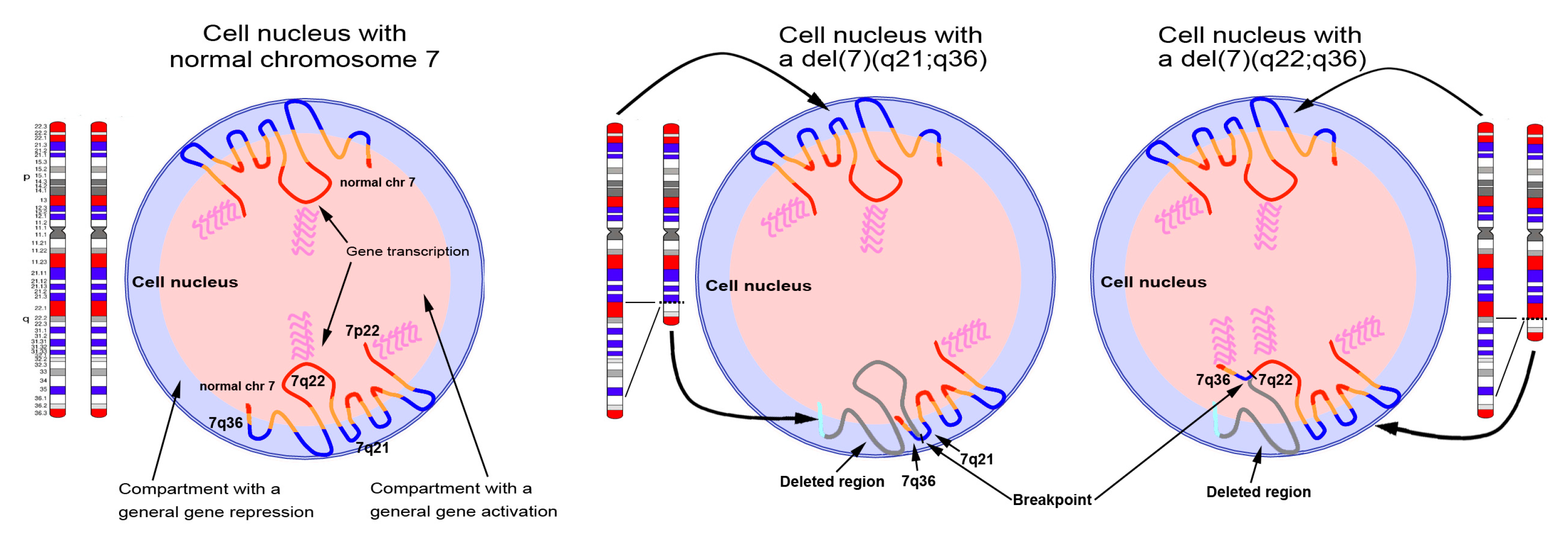
3.4. Mechanisms Leading to HLXB9 Re-positioning in the Nucleus
3.5. Gene Expression in 3D Chromatin Architecture
4. Materials and Methods
4.1. Patient Samples
4.2. Control Samples
4.3. Reverse Transcription Polymerase Chain Reaction (RT-PCR)
4.4. Data Mining
4.5. Fluorescence in Situ Hybridization (FISH) and Image Capture
4.6. Radial Nuclear Positioning Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Pombo, A.; Dillon, N. Three-dimensional genome architecture: players and mechanisms. Nat. Rev. Mol. Cell. Biol. 2015, 16, 245–257. [Google Scholar] [CrossRef]
- Brickner, J. Genetic and epigenetic control of the spatial organization of the genome. Mol. Biol. Cell 2017, 28, 364–369. [Google Scholar] [CrossRef] [PubMed]
- Sadoni, N.; Langer, S.; Fauth, C.; Bernardi, G.; Cremer, T.; Turner, B.M.; Zink, D. Nuclear organization of mammalian genomes: polar chromosome territories build up functionally distinct higher order compartments. J. Cell. Biol. 1999, 146, 1211–1226. [Google Scholar] [CrossRef] [PubMed]
- Croft, J.A.; Bridger, J.M.; Boyle, S.; Perry, P.; Teague, P.; Bickmore, W.A. Differences in the localization and morphology of chromosomes in the human nucleus. J. Cell. Biol. 1999, 145, 1119–1131. [Google Scholar] [CrossRef] [PubMed]
- Boyle, S.; Gilchrist, S.; Bridger, J.M.; Mahy, N.L.; Ellis, J.A.; Bickmore, W.A. The spatial organization of human chromosomes within the nuclei of normal and emerin-mutant cells. Hum. Mol. Genet. 2001, 10, 211–219. [Google Scholar] [CrossRef] [PubMed]
- Cremer, T.; Cremer, C. Chromosome territories, nuclear architecture and gene regulation in mammalian cells. Nat. Rev. Genet. 2001, 2, 292–301. [Google Scholar] [CrossRef] [PubMed]
- Saccone, S.; Federico, C.; Bernardi, G. Localization of the gene-richest and the gene-poorest isochores in the interphase nuclei of mammals and birds. Gene 2002, 300, 169–178. [Google Scholar] [CrossRef]
- Federico, C.; Scavo, C.; Cantarella, C.D.; Motta, S.; Saccone, S.; Bernardi, G. Gene-rich and gene-poor chromosomal regions have different locations in the interphase nuclei of cold-blooded vertebrates. Chromosoma 2006, 115, 123–128. [Google Scholar] [CrossRef] [PubMed]
- Meuleman, W.; Peric-Hupkes, D.; Kind, J.; Beaudry, J.B.; Pagie, L.; Kellis, M.; Reinders, M.; Wessels, L.; van Steensel, B. Constitutive nuclear lamina–genome interactions are highly conserved and associated with A/T-rich sequence. Genome Res. 2013, 23, 270–280. [Google Scholar] [CrossRef] [PubMed]
- Federico, C.; Cantarella, C.D.; Di Mare, P.; Tosi, S.; Saccone, S. The radial arrangement of the human chromosome 7 in the lymphocyte cell nucleus is associated with chromosomal band gene density. Chromosoma 2008, 117, 399–410. [Google Scholar] [CrossRef] [PubMed]
- Misteli, T. Beyond the sequence: cellular organization of genome function. Cell 2007, 128, 787–800. [Google Scholar] [CrossRef]
- Morey, C.; Da Silva, N.R.; Kmita, M.; Duboule, D.; Bickmore, W.A. Ectopic nuclear reorganization driven by a Hoxb1 transgene transported into Hoxd. J. Cell. Sci. 2008, 121, 571–577. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Finlan, L.E.; Sproul, D.; Thomson, I.; Boyle, S.; Kerr, E.; Perry, P.; Ylstra, B.; Chubb, J.R.; Bickmore, W.A. Recruitment to the nuclear periphery can alter expression of genes in human cells. PLoS Genet. 2008, 4, e1000039. [Google Scholar] [CrossRef] [PubMed]
- Szczerbal, I.; Foster, H.A.; Bridger, J.M. The spatial repositioning of adipogenesis genes is correlated with their expression status in a porcine mesenchymal stem cell adipogenesis model system. Chromosoma 2009, 118, 647–663. [Google Scholar] [CrossRef] [PubMed]
- Elcock, L.S.; Bridger, J.M. Exploring the relationship between interphase gene positioning, transcriptional regulation and the nuclear matrix. Biochem. Soc. Trans. 2010, 38, 263–267. [Google Scholar] [CrossRef] [PubMed]
- Harewood, L.; Schutz, F.; Boyle, S.; Perry, P.; Delorenzi, M.; Bickmore, W.A.; Reymond, A. The effect of translocation-induced nuclear reorganization on gene expression. Genome Res. 2010, 20, 554–564. [Google Scholar] [CrossRef] [PubMed]
- Pindyurin, A.V. van Steensel, B. Hox in space. Nucleus 2012, 3, 118–122. [Google Scholar] [CrossRef][Green Version]
- Fraser, J.; Williamson, I.; Bickmore, W.A.; Dostie, J. An overview of genome organization and how we got there: from FISH to Hi-C. Microbiol. Mol. Biol. Rev. 2015, 79, 347–372. [Google Scholar] [CrossRef]
- Lieberman Aiden, E.; van Berkum, N.L.; Williams, L.; Imakaev, M.; Ragoczy, T.; Telling, A.; Amit, I.; Lajoie, B.R.; Sabo, P.J.; Dorschner, M.O.; et al. Comprehensive mapping of long-range interactions reveals folding principles of the human genome. Science 2009, 326, 289–293. [Google Scholar] [CrossRef]
- Jabbari, K.; Bernardi, G. An isochore framework underlies chromatin architecture. PLoS ONE 2017, 12, e0168023. [Google Scholar] [CrossRef]
- Federico, C.; Pappalardo, A.M.; Ferrito, V.; Tosi, S.; Saccone, S. Genomic properties of chromosomal bands are linked to evolutionary rearrangements and new centromere formation in primates. Chromosome Res. 2017, 25, 261–276. [Google Scholar] [CrossRef]
- Dixon, J.R.; Selvaraj, S.; Yue, F.; Kim, A.; Li, Y.; Shen, Y.; Hu, M.; Liu, J.S.; Ren, B. Topological domains in mammalian genomes identified by analysis of chromatin interactions. Nature 2012, 485, 376. [Google Scholar] [CrossRef] [PubMed]
- Rao, S.S.; Huntley, M.H.; Durand, N.C.; Stamenova, E.K.; Bochkov, I.D.; Robinson, J.T.; Sanborn, A.L.; Machol, I.; Omer, A.D.; Lander, E.S.; et al. A 3D map of the human genome at kilobase resolution reveals principles of chromatin looping. Cell 2014, 159, 1665–1680. [Google Scholar] [CrossRef]
- Akhtar, W.; de Jong, J.; Pindyurin, A.V.; Pagie, L.; Meuleman, W.; de Ridder, J.; Berns, A.; Wessels, L.F.; van Lohuizen, M.; van Steensel, B. Chromatin position effects assayed by thousands of reporters integrated in parallel. Cell 2013, 154, 914–927. [Google Scholar] [CrossRef]
- Kind, J.; Pagie, L.; Ortabozkoyun, H.; Boyle, S.; de Vries, S.S.; Janssen, H.; Amendola, M.; Nolen, L.D.; Bickmore, W.A.; van Steensel, B. Single-cell dynamics of genome-nuclear lamina interactions. Cell 2013, 153, 178–192. [Google Scholar] [CrossRef]
- Van Steensel, B.; Belmont, A.S. Lamina-associated domains: links with chromosome architecture, heterochromatin, and gene repression. Cell 2017, 169, 780–791. [Google Scholar] [CrossRef]
- Szczerbal, I.; Bridger, J.M. Association of adipogenic genes with SC-35 domains during porcine adipogenesis. Chromosome Res. 2010, 18, 887–895. [Google Scholar] [CrossRef] [PubMed]
- Leotta, C.G.; Federico, C.; Brundo, M.V.; Tosi, S.; Saccone, S. HLXB9 gene expression, and nuclear location during in vitro neuronal differentiation in the SK-N-BE neuroblastoma cell line. PLoS ONE 2104, 9, e105481. [Google Scholar] [CrossRef] [PubMed]
- Bridger, J.M.; Arican-Gotkas, H.D.; Foster, H.A.; Godwin, L.S.; Harvey, A.; Kill, I.R.; Knight, M.; Mehta, I.S.; Ahmed, M.H. The non-random repositioning of whole chromosomes and individual gene loci in interphase nuclei and its relevance in disease, infection, aging, and cancer. Springer 2014, 263–279. [Google Scholar]
- Meaburn, K.J.; Misteli, T. Locus-specific and activity-independent gene repositioning during early tumorigenesis. J. Cell. Biol. 2008, 180, 39–50. [Google Scholar] [CrossRef]
- Meaburn, K.J.; Gudla, P.R.; Khan, S.; Lockett, S.J.; Misteli, T. Disease-specific gene repositioning in breast cancer. J. Cell. Biol. 2009, 187, 801–812. [Google Scholar] [CrossRef] [PubMed]
- Federico, C.; Leotta, C.G.; Bruno, F.; Longo, A.M.; Owoka, T.; Tosi, S.; Saccone, S. GDM-1. Nuclear repositioning of the non-translocated HLXB9 allele in the leukemia cell line GDM-1 harboring a t(6;7)(q23;q36). Cytogenet. Genome Res 2017, 153, 10–17. [Google Scholar] [CrossRef]
- Kozubek, S.; Lukasova, E.; Mareckova, A.; Skalníková, M.; Kozubek, M.; Bártová, E.; Kroha, V.; Krahulcová, E.; Slotová, J. The topological organization of chromosomes 9 and 22 in cell nuclei has a determinative role in the induction of t(9,22) translocations and in the pathogenesis of t(9,22) leukemias. Chromosoma 1999, 108, 426–435. [Google Scholar] [CrossRef] [PubMed]
- Lukášová, E.; Kozubek, S.; Kozubek, M.; Falk, M.; Amrichová, J. The 3D structure of human chromosomes in cell nuclei. Chromosom. Res. 2002, 10, 535–548. [Google Scholar] [CrossRef]
- Taslerova, R.; Kozubek, S.; Lukasova, E.; Jirsová, P.; Bártová, E.; Kozubek, M. Arrangement of chromosome 11 and 22 territories, EWSR1 and FLI1 genes, and other genetic elements of these chromosomes in human lymphocytes and Ewing sarcoma cells. Hum. Genet. 2003, 112, 143–155. [Google Scholar] [PubMed]
- Barki-Celli, L.; Lefebvre, C.; Le Baccon, P.; Nadeau, G.; Bonnefoix, T.; Usson, Y.; Vourc’h, C.; Khochbin, S.; Leroux, D.; Callanan, M. Differences in nuclear positioning of 1q12 pericentric heterochromatin in normal and tumor B lymphocytes with 1q rearrangements. Genes, Chromosomes Cancer 2005, 43, 339–349. [Google Scholar] [CrossRef]
- Grasser, F.; Neusser, M.; Fiegler, H.; Thormeyer, T.; Cremer, M.; Carter, N.P.; Cremer, T.; Müller, S. Replication-timing-correlated spatial chromatin arrangements in cancer and in primate interphase nuclei. J. Cell. Sci. 2008, 121, 1876–1886. [Google Scholar] [CrossRef]
- Ballabio, E.; Cantarella, C.D.; Federico, C.; Di Mare, P.; Hall, G.; Harbott, J.; Hughes, J.; Saccone, S.; Tosi, S. Ectopic expression of the HLXB9 gene is associated with an altered nuclear position in t(7;12). Leukemia 2009, 23, 1179–1182. [Google Scholar] [CrossRef]
- Tosi, S.; Mostafa Kamel, Y.; Owoka, T.; Federico, C.; Truong, T.H.; Saccone, S. Pediatric acute myeloid leukemia with the t(7;12)(q36;p13) rearrangement: a review of the biological and clinical management aspects. Biomark Res. 2015, 3, 21. [Google Scholar] [CrossRef]
- Harrison, K.A.; Thaler, J.; Pfaff, S.L.; Gu, H.; Kehrl, J.H. Pancreas dorsal lobe agenesis and abnormal islets of Langerhans in HLXB9-deficient mice. Nature genetics 1999, 23, 71. [Google Scholar] [CrossRef]
- Thaler, J.; Harrison, K.; Sharma, K.; Lettieri, K.; Kehrl, J.; Pfaff, S.L. Active suppression of interneuron programs within developing motor neurons revealed by analysis of homeodomain factor HB9. Neuron 1999, 23, 675–687. [Google Scholar] [CrossRef]
- Hagan, D.M.; Ross, A.J.; Strachan, T.A.; Lynch, S.A.; Ruiz-Perez, V.; Wang, Y.M.; Scambler, P.; Custard, E.; Reardon, W.; Hassan, S.; et al. Mutation analysis and embryonic expression of the HLXB9 Currarino syndrome gene. Am. J. Hum. Genet. 2000, 66, 1504–1515. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Neufing, P.J.; Kalionis, B.; Horsfall, D.J.; Ricciardelli, C.; Stahl, J.; Vivekanandan, S.; Raymond, W.; Tilley, W.D. Expression and localization of homeodomain proteins DLX4/HB9 in normal and malignant human breast tissues. Anticancer Res. 2003, 23, 1479–1488. [Google Scholar]
- Tian, T.; Wang, M.; Zhu, Y.; Zhu, W.; Yang, T.; Li, H.; Lin, S.; Dai, C.; Deng, Y.; Song, D.; et al. Expression, clinical significance, and functional prediction of MNX1 in breast cancer. Mol. Ther. Nucleic Acids 2018, 13, 399–406. [Google Scholar] [CrossRef]
- Zhang, L.; Wang, J.; Wang, Y.; Zhang, Y.; Castro, P.; Shao, L.; Sreekumar, A.; Putluri, N.; Guha, N.; Deepak, S.; et al. MNX1 is oncogenically upregulated in African-American prostate cancer. Cancer Res. 2016, 76, 6290–6298. [Google Scholar] [CrossRef] [PubMed]
- Chen, M.; Wu, R.; Li, G.; Liu, C.; Tan, L.; Xiao, K.; Ye, Y.; Qin, Z. Motor neuron and pancreas homeobox 1/HLXB9 promotes sustained proliferation in bladder cancer by upregulating CCNE1/2. J. Exp. Clin. Cancer Res. 2018, 37, 154. [Google Scholar] [CrossRef]
- Hollington, P.; Neufing, P.; Kalionis, B.; Waring, P.; Bentel, J.; Wattchow, D.; Tilley, W.D. Expression and localization of homeodomain proteins DLX4, HB9, and HB24 in malignant and benign human colorectal tissues. Anticancer Res. 2004, 24, 955–962. [Google Scholar]
- Wilkens, L.; Jaggi, R.; Hammer, C.; Inderbitzin, D.; Giger, O. von Neuhoff, N. The homeobox gene HLXB9 is upregulated in a morphological subset of poorly differentiated hepatocellular carcinoma. Virchows Arch. 2011, 458, 697–708. [Google Scholar] [CrossRef]
- Desai, S.S.; Kharade, S.S.; Parekh, V.I.; Iyer, S.; Agarwal, S.K. Pro-oncogenic roles of HLXB9 in insulinoma cells through interaction with Nono and downregulation of the c-Met inhibitor Cblb. J. Biol. Chem. 2015, 290, 25595–25608. [Google Scholar] [CrossRef]
- Harrison, K.A.; Druey, K.M.; Deguchi, Y.; Tuscano, J.M.; Kehrl, J.H. A novel human homeobox gene distantly related to proboscipedia is expressed in lymphoid and pancreatic tissues. J. Biol. Chem. 1994, 269, 19968–19975. [Google Scholar]
- Deguchi, Y.; Kehrl, J.H. Selective expression of two homeobox genes in CD34-positive cells from human bone marrow. Blood 1991, 78, 323–328. [Google Scholar]
- Von Bergh, A.R.; van Drunen, E.; van Wering, E.R.; van Zutven, L.J.; Hainmann, I.; Lönnerholm, G.; Meijerink, J.P.; Pieters, R.; Beverloo, H.B. High incidence of t(7;12)(q36;p13) in infant AML but not in infant ALL, with a dismal outcome and ectopic expression of HLXB9. Genes Chromosomes Cancer 2006, 45, 731–739. [Google Scholar] [CrossRef] [PubMed]
- Wildenhain, S.; Ingenhag, D.; Ruckert, C.; Degistirici, Ö.; Dugas, M.; Meisel, R.; Hauer, J.; Borkhardt, A. Homeobox protein HB9 binds to the prostaglandin E receptor, 2 promoter and inhibits intracellular cAMP mobilization in leukemic cells. J. Biol. Chem. 2012, 287, 40703–40712. [Google Scholar] [CrossRef] [PubMed]
- Ingenhag, D.; Reister, S.; Auer, F.; Bhatia, S.; Wildenhain, S.; Picard, D.; Remke, M.; Hoell, J.I.; Kloetgen, A.; Sohn, D.; et al. The homeobox transcription factor HB9 induces senescence and blocks differentiation in hematopoietic stem and progenitor cells. Hematologica 2018, 104, 35–46. [Google Scholar] [CrossRef] [PubMed]
- Gupta, R.; Harankhedkar, S.; Rahman, K.; Singh, M.K.; Chandra, D.; Mittal, N.; Gupta, A.; Nityanand, S. Prevalence of chromosome 7 abnormalities in myelodysplastic syndrome and acute myeloid leukemia: A single center study and brief literature review. Indian J. Hematol. Blood Transfus. 2018, 34, 602–611. [Google Scholar] [CrossRef]
- Le Beau, M.M.; Espinosa, R.; Davis, E.M.; Eisenbart, J.D.; Larson, R.A.; Green, E.D. Cytogenetic and molecular delineation of a region of chromosome 7 commonly deleted in malignant myeloid diseases. Blood 1996, 88, 1930–1935. [Google Scholar] [CrossRef]
- Fischer, K.; Fröhling, S.; Scherer, S.W.; McAllister Brown, J.; Scholl, C.; Stilgenbauer, S.; Tsui, L.C.; Lichter, P.; Döhner, H. Molecular cytogenetic delineation of deletions and translocations involving chromosome band 7q22 in myeloid leukemias. Blood 1997, 89, 2036–2041. [Google Scholar]
- Tosi, S.; Scherer, S.W.; Giudici, G.; Czepulkowski, B.; Biondi, A.; Kearney, L. Delineation of multiple deleted regions in 7q in myeloid disorders. Genes Chromosomes Cancer 1999, 25, 384–392. [Google Scholar] [CrossRef]
- Honda, H.; Nagamachi, A.; Inaba, T. -7/7q- syndrome in myeloid-lineage hematopoietic malignancies: attempts to understand this complex disease entity. Oncogene 2015, 34, 2413–2425. [Google Scholar] [CrossRef]
- Mehta, P.A.; Harris, R.E.; Davies, S.M.; Kim, M.O.; Mueller, R.; Lampkin, B.; Mo, J.; Myers, K.; Smolarek, T.A. Numerical chromosomal changes and risk of development of myelodysplastic syndrome–acute myeloid leukemia in patients with Fanconi anemia. Cancer Genet. Cytogenet. 2010, 203, 180–186. [Google Scholar] [CrossRef]
- Tosi, S.; Harbott, J.; Haas, O.A.; Douglas, A.; Hughes, D.M.; Ross, F.M.; Biondi, A.; Scherer, S.W.; Kearney, L. Classification of deletions and identification of cryptic translocations involving 7q by fluoresence in situ hybridization. Leukemia 1996, 10, 644–649. [Google Scholar]
- Costantini, M.; Clay, O.; Federico, C.; Saccone, S.; Auletta, F.; Bernardi, G. Human chromosomal bands: nested structure, high definition map and molecular basis. Chromosoma 2007, 116, 29–40. [Google Scholar] [CrossRef]
- Kind, J.; Pagie, L.; de Vries, S.S.; Nahidiazar, L.; Dey, S.S.; Bienko, M.; Zhan, Y.; Lajoie, B.; de Graaf, C.A.; Amendola, M.; et al. Genome-wide maps of nuclear lamina interactions in single human cells. Cell 2015, 163, 134–147. [Google Scholar] [CrossRef]
- Tosi, S.; Hughes, J.; Scherer, S.W.; Nakabayashi, K.; Harbott, J.; Haas, O.A.; Cazzaniga, G.; Biondi, A.; Kempski, H.; Kearney, L. Heterogeneity of the 7q36 breakpoints in the t(7;12) involving ETV6 in infant leukemia. Genes Chromosomes Cancer 2003, 38, 191–200. [Google Scholar] [CrossRef]
- Ferguson, S.; Gautrey, H.E.; Strathdee, G. The dual role of HLXB9 in leukemia. Pediatr. Blood Cancer 2011, 56, 349–352. [Google Scholar] [CrossRef]
- González, M.B.; Gutiérrez, N.C.; García, J.L.; Schoenmakers, E.F.; Solé, F.; Calasanz, M.J.; San Miguel, J.F.; Hernández, J.M. Heterogeneity of structural abnormalities in the 7q31. 3∼ q34 region in myeloid malignancies. Cancer Genet. Cytogenet. 2004, 150, 136–143. [Google Scholar] [CrossRef]
- McNerney, M.E.; Brown, C.D.; Wang, X.; Bartom, E.T.; Karmakar, S.; Bandlamudi, C.; Yu, S.; Ko, J.; Sandall, B.P.; Stricker, T.; et al. CUX1 is a haploinsufficient tumor suppressor gene on chromosome 7 frequently inactivated in acute myeloid leukemia. Blood 2013, 121, 975–983. [Google Scholar] [CrossRef]
- Huang, R.; Liao, X.; Li, Q. Identification of key pathways and genes in TP53 mutation acute myeloid leukemia: evidence from bioinformatics analysis. Onco. Targets Ther. 2017, 11, 163–173. [Google Scholar] [CrossRef]
- Inaba, T.; Honda, H.; Matsui, H. The enigma of monosomy 7. Blood 2017, 131, 2891–2898. [Google Scholar] [CrossRef]
- Jost, K.L.; Bertulat, B.; Rapp, A.; Brero, A.; Hardt, T.; Domaing, P.; Gösele, C.; Schulz, H.; Hübner, N.; Cardoso, M.C. Gene repositioning within the cell nucleus is not random and is determined by its genomic neighborhood. Epigenetics Chromatin 2015, 8, 36. [Google Scholar] [CrossRef]
- Guelen, L.; Pagie, L.; Brasset, E.; Meuleman, W.; Faza, M.B.; Talhout, W.; Eussen, B.H.; de Klein, A.; Wessels, L.; de Laat, W.; et al. Domain organization of human chromosomes revealed by mapping of nuclear lamina interactions. Nature 2008, 453, 948–951. [Google Scholar] [CrossRef] [PubMed]
- Gonzalez-Sandoval, A.; Towbin, B.D.; Kalck, V.; Cabianca, D.S.; Gaidatzis, D.; Hauer, M.H.; Geng, L.; Wang, L.; Yang, T.; Wang, X.; et al. Perinuclear anchoring of H3K9-methylated chromatin stabilizes induced cell fate in C. elegans embryos. Cell 2015, 163, 1333–1347. [Google Scholar] [CrossRef]
- Egecioglu, D.; Brickner, J.H. Gene positioning and expression. Curr. Opin. Cell. Biol. 2011, 23, 338–345. [Google Scholar] [CrossRef] [PubMed]
- Remeseiro, S.; Hörnblad, A.; Spitz, F. Gene regulation during development in the light of topologically associating domains. Wiley Interdiscip. Rev. Dev. Biol. 2015, 5, 169–185. [Google Scholar] [CrossRef]
- Rodríguez-Carballo, E.; Lopez-Delisle, L.; Zhan, Y.; Fabre, P.J.; Beccari, L.; El-Idrissi, I.; Huynh, T.H.N.; Ozadam, H.; Dekker, J.; Duboule, D. The HoxD cluster is a dynamic and resilient TAD boundary controlling the segregation of antagonistic regulatory landscapes. Genes Dev. 2017, 31, 2264–2281. [Google Scholar] [CrossRef] [PubMed]
- Leshner, M.; Devine, M.; Roloff, G.W.; True, L.D.; Misteli, T.; Meaburn, K.J. Locus-specific gene repositioning in prostate cancer. Mol. Biol. Cell. 2016, 27, 236–246. [Google Scholar] [CrossRef] [PubMed]
- Meaburn, K.J.; Agunloye, O.; Devine, M.; Leshner, M.; Roloff, G.W.; True, L.D.; Misteli, T. Tissue-of-origin-specific gene repositioning in breast and prostate cancer. Histochem. Cell Biol. 2016, 145, 433–446. [Google Scholar] [CrossRef] [PubMed]
- Taberlay, P.C.; Achinger-Kawecka, J.; Lun, A.T.; Buske, F.A.; Sabir, K.; Gould, C.M.; Zotenko, E.; Bert, S.A.; Giles, K.A.; Bauer, D.C.; et al. Three-dimensional disorganization of the cancer genome occurs coincident with long range genetic and epigenetic alterations. Genome Res. 2016, 26, 719–731. [Google Scholar] [CrossRef] [PubMed]
- Tian, X.; Wang, Y.; Zhao, F.; Liu, J.; Yin, J.; Chen, D.; Ma, W.; Ke, X. A new classification of interphase nuclei based on spatial organization of chromosome 8 and 21 for t(8;21)(q22q22) acute myeloid leukemia by three-dimensional fluorescence in situ hybridization. Leuk. Res. 2015, 39, 1414–1420. [Google Scholar] [CrossRef] [PubMed]
- Beverloo, H.B.; Panagopoulos, I.; Isaksson, M.; van Wering, E.; van Drunen, E.; de Klein, A.; Johansson, B.; Slater, R. Fusion of the homeobox gene HLXB9 and the ETV6 gene in infant acute myeloid leukemias with the t(7;12)(q36;p13). Cancer Res. 2001, 61, 5374–5377. [Google Scholar]
- Vivian, J.; Rao, A.A.; Nothaft, F.A.; Ketchum, C.; Armstrong, J.; Novak, A.; Pfeil, J.; Narkizian, J.; Deran, A.D.; Musselman-Brown, A.; et al. Toil enables reproducible, open source, big biomedical data analyzes. Nat. Biotechnol. 2017, 35, 314–316. [Google Scholar] [CrossRef] [PubMed]
- Garimberti, E.; Tosi, S. Fluorescence in situ hybridization (FISH), basic principles and methodology. Methods Mol. Biol. 2010, 659, 3–20. [Google Scholar] [PubMed]
- Landau, S.; Rabe-Hesketh, S. StatView for Windows, Version 5.0. Stat. Methods Med. Res. 1999, 8, 337–341. [Google Scholar] [CrossRef] [PubMed]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Pt | Diagnosis | Karyotype | Reference |
|---|---|---|---|
| 1 | CMML | 46,del(7)(q21q36) | [61] |
| 2 | MDS | 46,XY,del(7)(q21.3-22q36) | [58,61] |
| 3 | AML-M5 | 46,XX,del(7)(q22q36) | [58] |
| 4 | MDS | 46,XY,del(7)(q22q36) | [61] |
| 5 | MDS | 45,XX,del(5)(q14),del(7)(q22), del(9)(q22),-17 | This study |
| 6 | CML | 47,XY,del(7)(q21),+21[20] 1 | This study |
| 7 | FA | 46,XY,dup(1)(q24q44),del(7)(q21)[5] 1 | This study |
| 8 | AML-M6 | 44,XY,t(1;3)(p21;?),-5,del(7)(q31.1),-17,-20,+mar[cp18] 1 | This study |
| 9 | t-AML | 46,XX,del(7)(q32.1[4]/46,XX[6] | This study |
| 10 | AML | 47,XY,+8[11]/47,idem,del(7)(q32.1)[9] | This study |
| Probes | Band | Patients | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| Pt.1 | Pt.2 | Pt.3 | Pt.4 | Pt.5 | Pt.6 | Pt.7 | Pt.8 | Pt.9 | Pt.10 | ||
| RP11-90N9 | 7q21.11 | + | + | + | + | + | |||||
| Cos 5.2 | 7q21.3 | + | |||||||||
| RP11-213E22 | 7q22.1 | − | − | + | + | + | |||||
| Cos 7d.2 | 7q22.1 | − | |||||||||
| Cos 8a.2 | 7q22.1 | − | |||||||||
| Cos 1120 | 7q22.1 | − | |||||||||
| PAC 1065 (CUTL1) | 7q22.1 | + | |||||||||
| KMT2E gene 1 | 7q22 | − | − | + | + | + | |||||
| HSC7E485 | 7q22.2 | − | − | ||||||||
| MET gene 1 | 7q31.2 | − | − | − | + | + | |||||
| RP11-73H23 | 7q34 | − | − | − | − | − | |||||
| HSC7E124 | 7q36.1 | − | − | ||||||||
| EZH2 gene 1 | 7q36.1 | + | − | − | + | + | |||||
| HSC7E86 | 7q36.2 | − | + | ||||||||
| HSC7E769 | 7q36.2 | + | |||||||||
| RP5-1121A15 | 7q36.3 | + | + | + | + | + | + | + | + | + | + |
| HLXB9Expression | RNL | GC-level (a) | Pt ID | |
|---|---|---|---|---|
| Not Expressed | 0.754(•) | GC-poor | 7 |  |
| Not Expressed | 0.722(•) | GC-poor | 9 | |
| Expressed | 0.710(•) | GC-poor | 10 | |
| Not Expressed | 0.671(•) | GC-poor | 6 | |
| NA | 0.652(▲) | GC-poor | 1 | |
| Not Expressed | 0.652(▲) | GC-poor | 2 | |
| Expressed | 0.629(▲) | GC-rich | 4 | |
| Expressed | 0.605(▲) | GC-rich | 8 | |
| NA | 0.601(▲) | GC-rich | 3 | |
| NA | 0.593(▲) | GC-rich | 5 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Federico, C.; Owoka, T.; Ragusa, D.; Sturiale, V.; Caponnetto, D.; Leotta, C.G.; Bruno, F.; Foster, H.A.; Rigamonti, S.; Giudici, G.; et al. Deletions of Chromosome 7q Affect Nuclear Organization and HLXB9Gene Expression in Hematological Disorders. Cancers 2019, 11, 585. https://doi.org/10.3390/cancers11040585
Federico C, Owoka T, Ragusa D, Sturiale V, Caponnetto D, Leotta CG, Bruno F, Foster HA, Rigamonti S, Giudici G, et al. Deletions of Chromosome 7q Affect Nuclear Organization and HLXB9Gene Expression in Hematological Disorders. Cancers. 2019; 11(4):585. https://doi.org/10.3390/cancers11040585
Chicago/Turabian StyleFederico, Concetta, Temitayo Owoka, Denise Ragusa, Valentina Sturiale, Domenica Caponnetto, Claudia Giovanna Leotta, Francesca Bruno, Helen A. Foster, Silvia Rigamonti, Giovanni Giudici, and et al. 2019. "Deletions of Chromosome 7q Affect Nuclear Organization and HLXB9Gene Expression in Hematological Disorders" Cancers 11, no. 4: 585. https://doi.org/10.3390/cancers11040585
APA StyleFederico, C., Owoka, T., Ragusa, D., Sturiale, V., Caponnetto, D., Leotta, C. G., Bruno, F., Foster, H. A., Rigamonti, S., Giudici, G., Cazzaniga, G., Bridger, J. M., Sisu, C., Saccone, S., & Tosi, S. (2019). Deletions of Chromosome 7q Affect Nuclear Organization and HLXB9Gene Expression in Hematological Disorders. Cancers, 11(4), 585. https://doi.org/10.3390/cancers11040585