p53-Dependent Apoptotic Effect of Puromycin via Binding of Ribosomal Protein L5 and L11 to MDM2 and Its Combination Effect with RITA or Doxorubicin

 ,
,  ,
,  ,
,  and
and 
Abstract
1. Introduction
2. Results
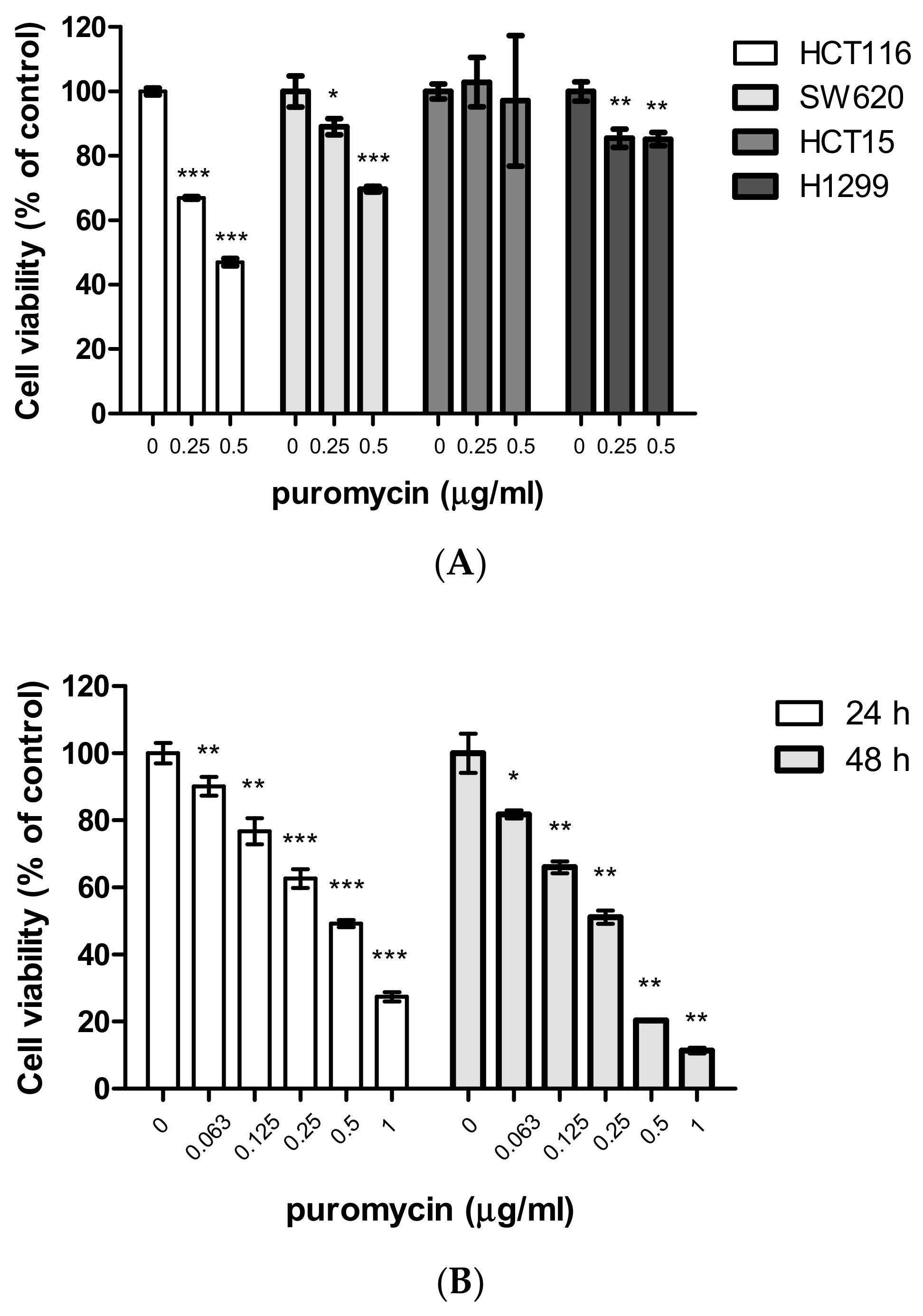
2.1. Puromycin Exerts Cytotoxic and Antiproliferative Effects in Cancer Cells
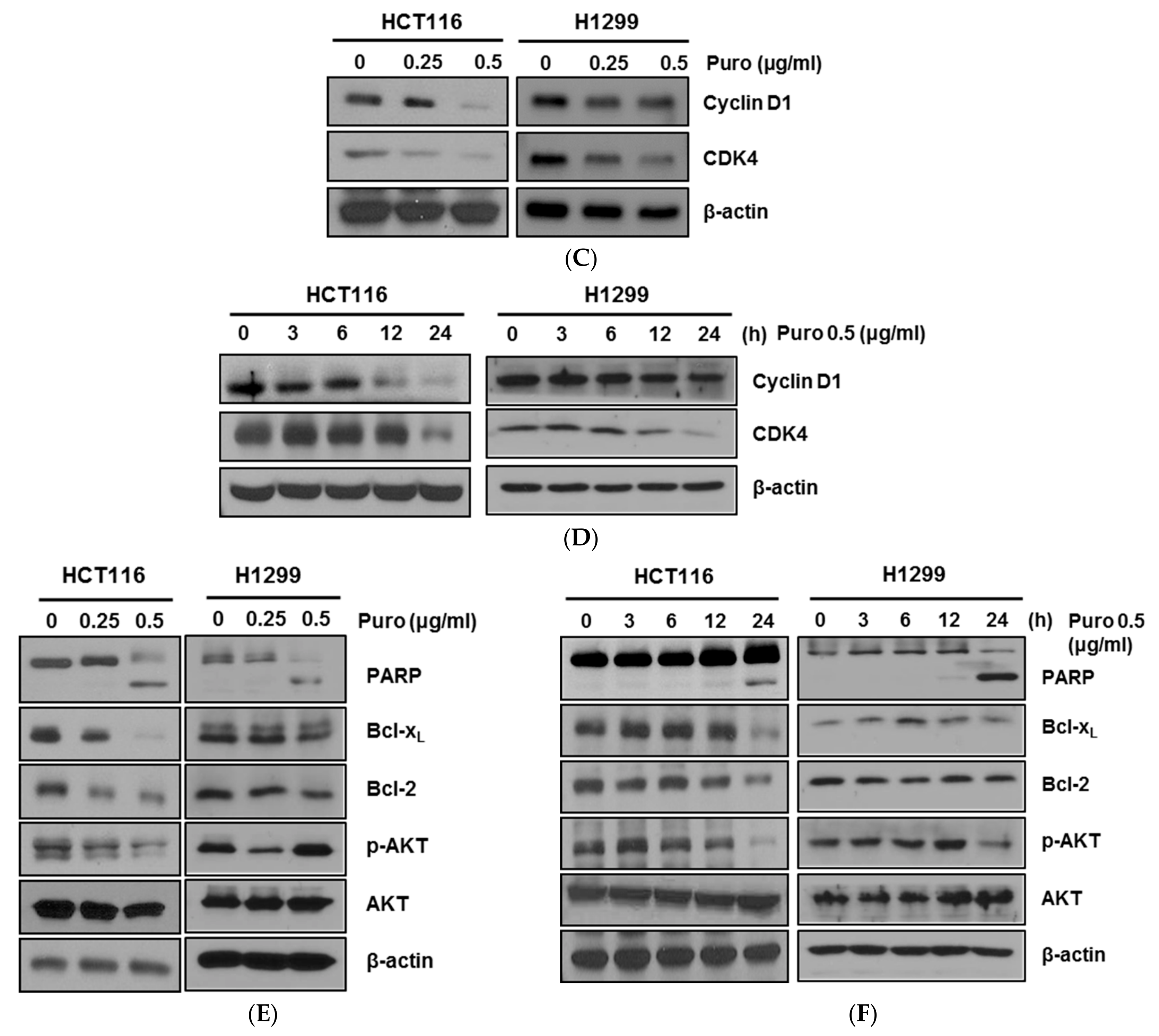
2.2. Puromycin Regulates Apoptosis-Related Proteins along with Increase of Sub-G1 Population
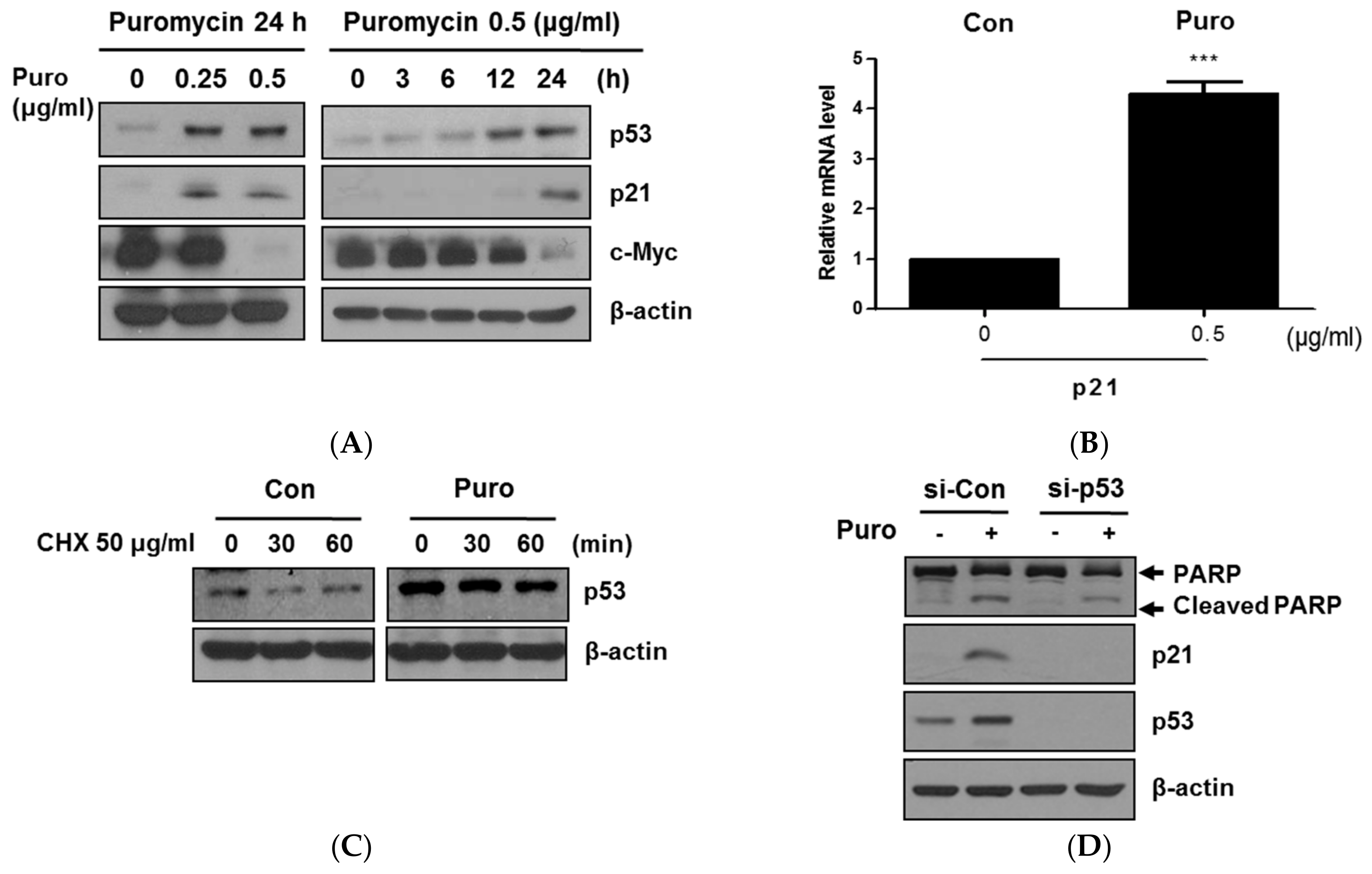
2.3. Puromycin Enhances p53 Stability along with Inhibition of c-Myc in HCT116 Cells
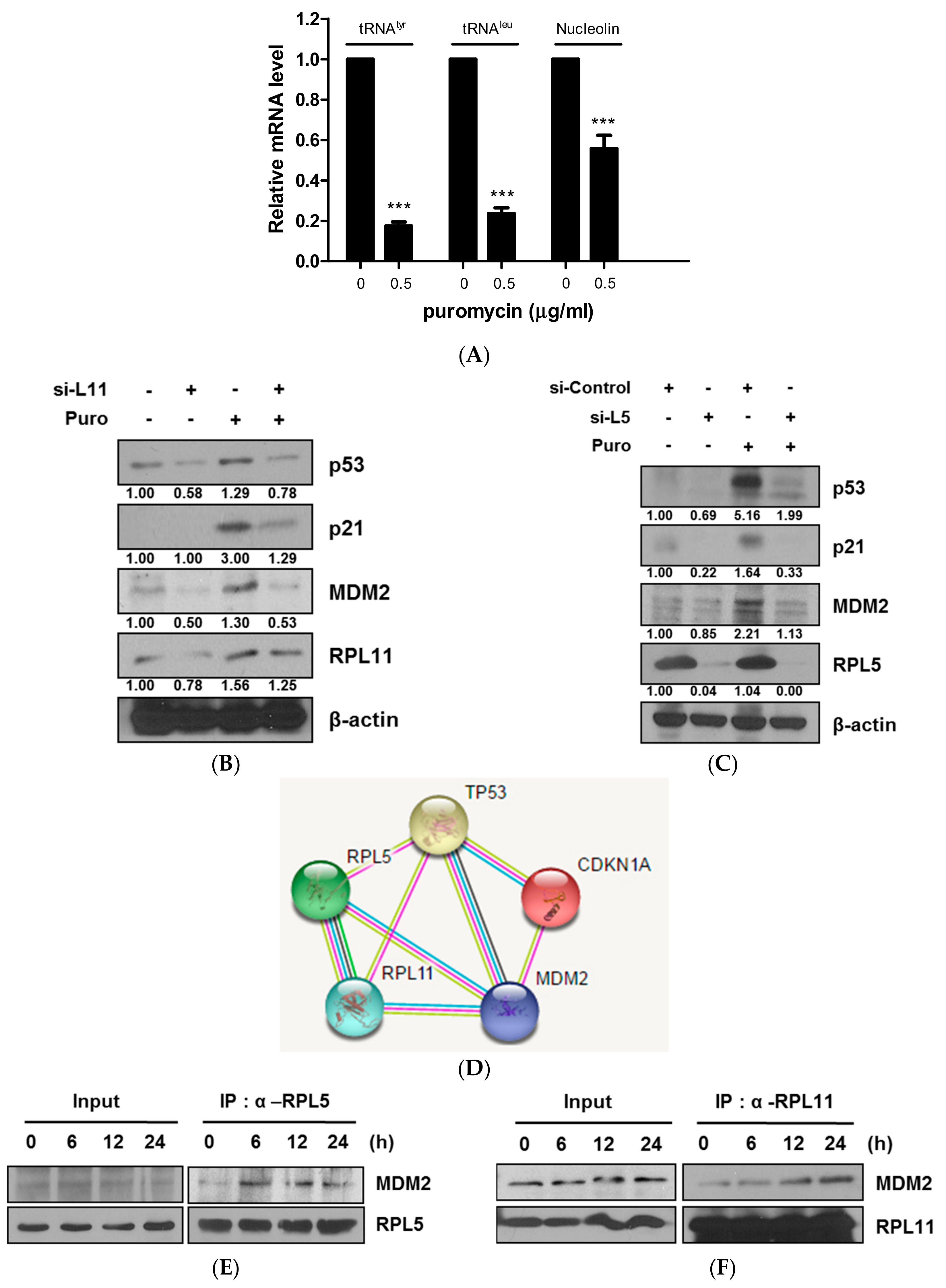
2.4. Ribosomal Protein L5 and L11 Mediate p53 Activation via Direct Binding to MDM2 in HCT116 Cells—An Example of an Equation
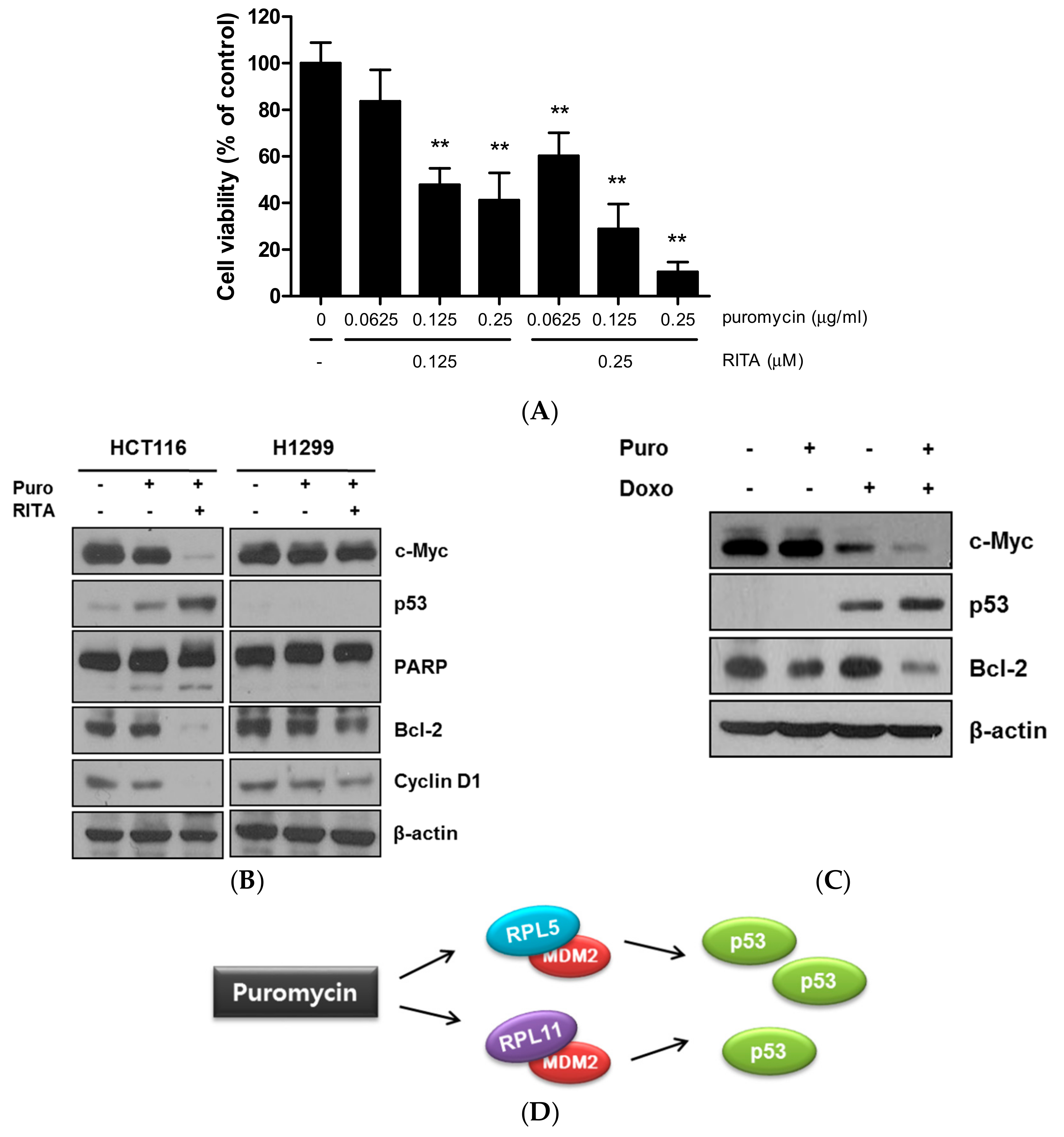
2.5. Combination Effect of Puromycin and Reactivating p53 and Inducing Tumor Apoptosis (RITA) in HCT116 Cells as Antitumor Agents
3. Discussion
4. Materials and Methods
4.1. Cell Culture
4.2. Cytotoxicity Assay
4.3. Cell Cycle Analysis
4.4. RNA Interference and Inhibitor Study
4.5. Colony Formation Assay
4.6. Western Blot and Co-Immunoprecipitation Analysis
4.7. Real-Time Quantitative PCR Analysis (RT-qPCR)
4.8. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Giege, R.; Frugier, M.; Rudinger, J. tRNA mimics. Curr. Opin. Struct. Biol. 1998, 8, 286–293. [Google Scholar] [CrossRef]
- Soderlund, G.; Haarhaus, M.; Chisalita, S.; Arnqvist, H.J. Inhibition of puromycin-induced apoptosis in breast cancer cells by IGF-I occurs simultaneously with increased protein synthesis. Neoplasma 2004, 51, 1–11. [Google Scholar]
- Jung, J.H.; Sohn, E.J.; Shin, E.A.; Lee, D.; Kim, B.; Jung, D.B.; Kim, J.H.; Yun, M.; Lee, H.J.; Park, Y.K.; Kim, S.H. Melatonin Suppresses the Expression of 45S Preribosomal RNA and Upstream Binding Factor and Enhances the Antitumor Activity of Puromycin in MDA-MB-231 Breast Cancer Cells. Evid. Based Complement. Altern. Med. eCAM 2013, 2013, 879746. [Google Scholar] [CrossRef] [PubMed]
- Lacsina, J.R.; Marks, O.A.; Liu, X.; Reid, D.W.; Jagannathan, S.; Nicchitta, C.V. Premature translational termination products are rapidly degraded substrates for MHC class I presentation. PLoS ONE 2012, 7, e51968. [Google Scholar] [CrossRef] [PubMed]
- Betts, H.M.; Milicevic Sephton, S.; Tong, C.; Awais, R.O.; Hill, P.J.; Perkins, A.C.; Aigbirhio, F.I. Synthesis, in Vitro Evaluation, and Radiolabeling of Fluorinated Puromycin Analogues: Potential Candidates for PET Imaging of Protein Synthesis. J. Med. Chem. 2016, 59, 9422–9430. [Google Scholar] [CrossRef] [PubMed]
- Lee, E.Y.; Muller, W.J. Oncogenes and tumor suppressor genes. Cold Spring Harb. Perspect. Biol. 2010, 2, a003236. [Google Scholar] [CrossRef] [PubMed]
- Qin, G.; Meng, Z. Expression of oncogenes and tumor suppressor genes in lungs of rats exposed to sulfur dioxide and benzo(a)pyrene. Inhal. Toxicol. 2010, 22, 322–329. [Google Scholar] [CrossRef] [PubMed]
- Strasser, A.; Harris, A.W.; Jacks, T.; Cory, S. DNA damage can induce apoptosis in proliferating lymphoid cells via p53-independent mechanisms inhibitable by Bcl-2. Cell 1994, 79, 329–339. [Google Scholar] [CrossRef]
- Liu, Y.; He, Y.; Jin, A.; Tikunov, A.P.; Zhou, L.; Tollini, L.A.; Leslie, P.; Kim, T.H.; Li, L.O.; Coleman, R.A.; et al. Ribosomal protein-Mdm2-p53 pathway coordinates nutrient stress with lipid metabolism by regulating MCD and promoting fatty acid oxidation. Proc. Natl. Acad. Sci. USA 2014, 111, E2414–E2422. [Google Scholar] [CrossRef]
- Hernandez-Monge, J.; Rousset-Roman, A.B.; Medina-Medina, I.; Olivares-Illana, V. Dual function of MDM2 and MDMX toward the tumor suppressors p53 and RB. Genes Cancer 2016, 7, 278–287. [Google Scholar] [PubMed]
- Karni-Schmidt, O.; Lokshin, M.; Prives, C. The Roles of MDM2 and MDMX in Cancer. Annu. Rev. Pathol. 2016, 11, 617–644. [Google Scholar] [CrossRef] [PubMed]
- Bursac, S.; Brdovcak, M.C.; Donati, G.; Volarevic, S. Activation of the tumor suppressor p53 upon impairment of ribosome biogenesis. Biochim. Biophys. Acta 2014, 1842, 817–830. [Google Scholar] [CrossRef] [PubMed]
- Stedman, A.; Beck-Cormier, S.; Le Bouteiller, M.; Raveux, A.; Vandormael-Pournin, S.; Coqueran, S.; Lejour, V.; Jarzebowski, L.; Toledo, F.; Robine, S.; et al. Ribosome biogenesis dysfunction leads to p53-mediated apoptosis and goblet cell differentiation of mouse intestinal stem/progenitor cells. Cell Death Differ. 2015, 22, 1865–1876. [Google Scholar] [CrossRef]
- Zhang, Y.; Lu, H. Signaling to p53: Ribosomal proteins find their way. Cancer Cell 2009, 16, 369–377. [Google Scholar] [CrossRef]
- Sun, X.X.; Dai, M.S.; Lu, H. Mycophenolic acid activation of p53 requires ribosomal proteins L5 and L11. J. Biol. Chem. 2008, 283, 12387–12392. [Google Scholar] [CrossRef] [PubMed]
- Mahata, B.; Sundqvist, A.; Xirodimas, D.P. Recruitment of RPL11 at promoter sites of p53-regulated genes upon nucleolar stress through NEDD8 and in an Mdm2-dependent manner. Oncogene 2012, 31, 3060–3071. [Google Scholar] [CrossRef]
- Zhou, X.; Hao, Q.; Liao, J.; Zhang, Q.; Lu, H. Ribosomal protein S14 unties the MDM2-p53 loop upon ribosomal stress. Oncogene 2013, 32, 388–396. [Google Scholar] [CrossRef] [PubMed]
- Sun, X.X.; DeVine, T.; Challagundla, K.B.; Dai, M.S. Interplay between ribosomal protein S27a and MDM2 protein in p53 activation in response to ribosomal stress. J. Biol. Chem. 2011, 286, 22730–22741. [Google Scholar] [CrossRef]
- James, A.; Wang, Y.; Raje, H.; Rosby, R.; DiMario, P. Nucleolar stress with and without p53. Nucleus 2014, 5, 402–426. [Google Scholar] [CrossRef]
- Golomb, L.; Volarevic, S.; Oren, M. p53 and ribosome biogenesis stress: The essentials. FEBS Lett. 2014, 588, 2571–2579. [Google Scholar] [CrossRef]
- Kruse, J.P.; Gu, W. Modes of p53 regulation. Cell 2009, 137, 609–622. [Google Scholar] [CrossRef] [PubMed]
- Ho, J.S.; Ma, W.; Mao, D.Y.; Benchimol, S. p53-Dependent transcriptional repression of c-myc is required for G1 cell cycle arrest. Mol. Cell. Biol. 2005, 25, 7423–7431. [Google Scholar] [CrossRef] [PubMed]
- Dai, M.S.; Sun, X.X.; Lu, H. Ribosomal protein L11 associates with c-Myc at 5 S rRNA and tRNA genes and regulates their expression. J. Biol. Chem. 2010, 285, 12587–12594. [Google Scholar] [CrossRef] [PubMed]
- Xie, S.Y.; Li, Y.J.; Wang, P.Y.; Jiao, F.; Zhang, S.; Zhang, W.J. miRNA-regulated expression of oncogenes and tumor suppressor genes in the cisplatin-inhibited growth of K562 cells. Oncol. Rep. 2010, 23, 1693–1700. [Google Scholar] [CrossRef]
- Singh, R.; Williams, J.; Vince, R. Puromycin based inhibitors of aminopeptidases for the potential treatment of hematologic malignancies. Eur. J. Med. Chem. 2017, 139, 325–336. [Google Scholar] [CrossRef]
- Davidoff, A.N.; Mendelow, B.V. Puromycin is a potent and specific inhibitor of tyrosine kinase activity in HL-60 cells. Anticancer Res. 1992, 12, 1761–1766. [Google Scholar]
- Ueki, N.; Wang, W.; Swenson, C.; McNaughton, C.; Sampson, N.S.; Hayman, M.J. Synthesis and Preclinical Evaluation of a Highly Improved Anticancer Prodrug Activated by Histone Deacetylases and Cathepsin L. Theranostics 2016, 6, 808–816. [Google Scholar] [CrossRef] [PubMed]
- Carr, M.I.; Jones, S.N. Regulation of the Mdm2-p53 signaling axis in the DNA damage response and tumorigenesis. Transl. Cancer Res. 2016, 5, 707–724. [Google Scholar] [CrossRef]
- Trino, S.; De Luca, L.; Laurenzana, I.; Caivano, A.; Del Vecchio, L.; Martinelli, G.; Musto, P. P53-MDM2 Pathway: Evidences for A New Targeted Therapeutic Approach in B-Acute Lymphoblastic Leukemia. Front. Pharmacol. 2016, 7, 491. [Google Scholar] [CrossRef]
- Khoury, K.; Popowicz, G.M.; Holak, T.A.; Domling, A. The p53-MDM2/MDMX axis—A chemotype perspective. MedChemComm 2011, 2, 246–260. [Google Scholar] [CrossRef][Green Version]
- Sipos, F.; Firneisz, G.; Muzes, G. Therapeutic aspects of c-MYC signaling in inflammatory and cancerous colonic diseases. World J. Gastroenterol. 2016, 22, 7938–7950. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Wolf, G.W.; Bhat, K.; Jin, A.; Allio, T.; Burkhart, W.A.; Xiong, Y. Ribosomal protein L11 negatively regulates oncoprotein MDM2 and mediates a p53-dependent ribosomal-stress checkpoint pathway. Mol. Cell. Biol. 2003, 23, 8902–8912. [Google Scholar] [CrossRef] [PubMed]
- Fancello, L.; Kampen, K.R.; Hofman, I.J.; Verbeeck, J.; De Keersmaecker, K. The ribosomal protein gene RPL5 is a haploinsufficient tumor suppressor in multiple cancer types. Oncotarget 2017, 8, 14462–14478. [Google Scholar] [CrossRef] [PubMed]
- Lohrum, M.A.; Ludwig, R.L.; Kubbutat, M.H.; Hanlon, M.; Vousden, K.H. Regulation of HDM2 activity by the ribosomal protein L11. Cancer Cell 2003, 3, 577–587. [Google Scholar] [CrossRef]
- Jin, A.; Itahana, K.; O’Keefe, K.; Zhang, Y. Inhibition of HDM2 and activation of p53 by ribosomal protein L23. Mol. Cell. Biol. 2004, 24, 7669–7680. [Google Scholar] [CrossRef]
- Ranjan, A.; Iwakuma, T. Non-Canonical Cell Death Induced by p53. Int. J. Mol. Sci. 2016, 17, 2068. [Google Scholar] [CrossRef]
- Aylon, Y.; Oren, M. The Paradox of p53: What, How, and Why? Cold Spring Harb. Perspect Med. 2016, 6, a026328. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Primer Name | Sequences 5′→3′ |
|---|---|
| p21-Forward | TCCAGGTTCAACCCACAGCTACTT |
| p21-Reverse | TCAGATGACTCTGGGAAACGCCAA |
| tRNATyr-Forward | CCTTCGATAGCTCAGCTGGTAGAGCGGAGG |
| tRNATyr-Reverse | CGGAATCGGAACCAGCGACCTAAGGATGTCC |
| tRNALeu-Forward | GTCAGGATGGCCGAGTGGTCTAAG |
| tRNALeu-Reverse | CCACGCCTCCATACGGAGAACCAGAAGACCC |
| Nucleolin-Forward | AAGCAGCACCTGGAAAACG |
| Nucleolin-reverse | TCTGAGCCTTCTACTTTCTGTTTCTTG |
| GAPDH-Forward | CACAATGCCGAAGTGGTCGT |
| GAPDH-Reverse | CACAATGCCGAAGTGGTCGT |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Jung, J.H.; Lee, H.; Kim, J.-H.; Sim, D.Y.; Ahn, H.; Kim, B.; Chang, S.; Kim, S.-H. p53-Dependent Apoptotic Effect of Puromycin via Binding of Ribosomal Protein L5 and L11 to MDM2 and Its Combination Effect with RITA or Doxorubicin. Cancers 2019, 11, 582. https://doi.org/10.3390/cancers11040582
Jung JH, Lee H, Kim J-H, Sim DY, Ahn H, Kim B, Chang S, Kim S-H. p53-Dependent Apoptotic Effect of Puromycin via Binding of Ribosomal Protein L5 and L11 to MDM2 and Its Combination Effect with RITA or Doxorubicin. Cancers. 2019; 11(4):582. https://doi.org/10.3390/cancers11040582
Chicago/Turabian StyleJung, Ji Hoon, Hyemin Lee, Ju-Ha Kim, Deok Yong Sim, Hyojin Ahn, Bonglee Kim, Suhwan Chang, and Sung-Hoon Kim. 2019. "p53-Dependent Apoptotic Effect of Puromycin via Binding of Ribosomal Protein L5 and L11 to MDM2 and Its Combination Effect with RITA or Doxorubicin" Cancers 11, no. 4: 582. https://doi.org/10.3390/cancers11040582
APA StyleJung, J. H., Lee, H., Kim, J.-H., Sim, D. Y., Ahn, H., Kim, B., Chang, S., & Kim, S.-H. (2019). p53-Dependent Apoptotic Effect of Puromycin via Binding of Ribosomal Protein L5 and L11 to MDM2 and Its Combination Effect with RITA or Doxorubicin. Cancers, 11(4), 582. https://doi.org/10.3390/cancers11040582