Abstract
Epstein-Barr virus (EBV) is an etiological agent for gastric cancer with significant worldwide variations. Molecular characterizations of EBV have shown phylogeographical variations among healthy populations and in EBV-associated diseases, particularly the cosegregated BamHI-I fragment and XhoI restriction site of exon 1 of the LMP-1 gene. In the Americas, both cosegregated variants are present in EBV carriers, which aligns with the history of Asian and European human migration to this continent. Furthermore, novel recombinant variants have been found, reflecting the genetic makeup of this continent. However, in the case of EBV-associated gastric cancer (EBV-associated GC), the cosegregated European BamHI-“i” fragment and XhoI restriction site strain prevails. Thus, we propose that a disrupted coevolution between viral phylogeographical strains and mixed human ancestry in the Americas might explain the high prevalence of this particular gastric cancer subtype. This cosegregated region contains two relevant transcripts for EBV-associated GC, the BARF-1 and miR-BARTs. Thus, genome-wide association studies (GWAS) or targeted sequencing of both transcripts may be required to clarify their role as a potential source of this disrupted coevolution.
1. Introduction
Since the recognition of the Epstein-Barr virus (EBV) as an etiological agent for gastric cancer (GC) [1], an explosion of research has taken place, focused on the pathogenesis and novel therapeutic developments for this particular subtype (for a review in this series see Reference [2]). In this scenario, meta-analyses and aggregated individual-level studies have shown significant variations in the worldwide rate of EBV-associated GC with a particularly high prevalence in the Americas compared with those reported in the Asian, European, and African continents [3,4,5,6,7]. Furthermore, studies in the so-called New World indicate that European (mostly Spanish) and Amerindian admixture (i.e., Hispanic ancestry) is clearly associated with higher rates of EBV infection in GC when compared with rates from those with White/non-Hispanic heritage in the US or Brazilians with Japanese ancestry (Table 1) [8,9,10,11]. Thus, the Americas provide a unique opportunity for uncovering the molecular basis of this particular subtype of GC. Here, we propose that a disrupted EBV-human co-evolution based on the combination of phylogeographic polymorphisms of EBV and mixed ancestries in the Americas might explain the high prevalence of this particular disease subtype.

Table 1.
Ancestry and Epstein-Barr virus-associated gastric cancer in the Americas.
2. Phylogenetic Classification of EBV
The molecular characterization of EBV has been facilitated by whole genome sequences, restriction fragment length polymorphisms (RFLP), genome-wide association studies (GWAS) from healthy donors, the study of benign and malignant lesions, and naturally infected GC SNU-719 cell lines [12,13,14,15,16] (for a review see Reference [17]). EBV can be classified as type 1 or 2, based on the substitution of 1.8 kb in the C-terminal domain of the EBNA-2 gene. Nucleotide differences at EBNA-3A, -3B, and -3C genes also contribute to this classification [18]. These two types of EBVs display phylogeographical differences as the EBV type-1 is the most common strain in the Asian, European, and American continents, whereas type-2 is frequently found in Africa [19,20]. These subtypes differ in their capacity to transform B-lymphocytes into a proliferative state [21]. A second classification of EBV has been elaborated by the RFLP map of the prototype EBV B95-8 genome after digestion with the BamHI restriction enzyme (Figure 1). The BamHI-F fragment is found in the majority of the healthy population and in EBV-associated diseases in Europe, Africa, and the Americas, including EBV-associated GC [22,23]. The presence of an extra BamHI restriction site within this fragment is the ‘‘f’’ variant. Although initially identified in nasopharyngeal carcinomas (NPC) (an undifferentiated epithelial-like tumor originating in the pharynx) [24,25], subsequent reports have found it in a low frequency among the healthy population and in EBV-associated diseases worldwide [23,26]. The “f” variant is located in the promoter region of the EBNA-1 gene, however, the functional significance of this polymorphism is currently unknown.
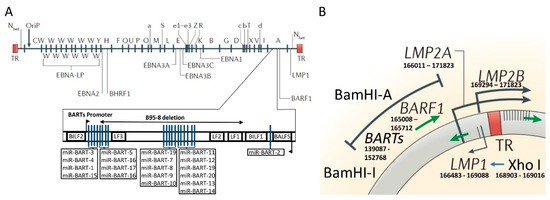
Figure 1.
Genomic map of Epstein-Barr virus (EBV) genome with major phylogeographic variants. (A) Diagram shows the location of open reading frames for EBV latent proteins on the BamHI restriction map of the prototype EBV B95-8 genome. The BamHI fragments are named according to size and are indicated by capital letters, with A being the largest. Lowercase letters indicate variations in size within these fragments. TR refers to the terminal repeats at each end of the genome. Below is a schematic representation of viral miR-BARTs in sequence order with deletion indicated within the BamHI-A fragment of the EBV genome in the B95-8 strain. Nhet is used to indicate heterogeneity in this region according to the number of TRs within different virus isolates. (B) Schematic representation of the genomic location of BamHI-I and BamHI-A fragments with TR. Within the exon 1 of LMP1 gene is the XhoI restriction site. The region containing the BamHI-I and XhoI restriction site is 21,277 nucleotides long and contains several relevant transcripts (miR-BARTS and BARF-1) for EBV transformation abilities. Figure is adapted from References [39] (with permission to use part of the figure) and [40] (localization of miR-BARTs).
The BamHI–I fragment harbors one of the greatest ranges of phylogeographical variations among healthy donors and EBV-associated diseases [23,26,27]. This fragment predominates in Asia, while the presence of an extra BamHI site defines the type “i”, which prevails in Europe and Africa [25,28,29,30,31]. Another relevant phylogeographical variation in the EBV chromosome is the polymorphism of the XhoI restriction site of exon 1 of LMP-1 gene. In Asia, the predominant viral strain lacks this restriction site (i.e., XhoI loss) [32]. However, the presence of this site (i.e., XhoI) defines the European and African subtypes [33].
Since these two sites (the BamHI–I fragment and the XhoI restriction sites) are closely located in the viral genome (see Figure 1), cosegregation is found in the Asian as well as the European and African strains. In the Americas, these cosegregations are also present among EBV carriers at a population level aligned with the waves of human migration [23,34]. In addition, novel recombinant variants have been found in this continent [23,35]. The discovery of these strains in the Americas should be understood as an amalgam of fragments from ancestral EBV sequences reflecting the genetic makeup of this continent’s population [36,37,38].
3. The Molecular Structure of the Cosegregated BamHI–I Fragment and XhoI Region of the EBV
GWAS analysis found that latent genes were the most diverse regions of the viral genome with the EBNA-3A, -3B, -3C, BPLF-1 and LMP-1 genes harboring the most abundant non-synonymous variants [41]. Interestingly, the LMP-1 gene region which contains the XhoI polymorphisms involved in the phylogeographical classification of EBV, may contribute to the variations in the prevalence of EBV-associated GC throughout the world. Additionally, two relevant transcript regions are located between both cosegregated variants (the BamHI–I fragment and the XhoI restriction site) (Figure 1), the BamHI-A rightward transcripts (BARTs) and BamHI-A rightward frame-1 (BARF-1). BARTs transcripts have several distinct spliced forms [42,43], whereas BARF-1 is located downstream of BART and encodes 221 amino acids [43,44] which are translated into a protein of 31–33 kDa [45].
EBV-encoded BARF-1 is a putative viral oncogene (oncogenic initiator or oncogenic cofactor) in EBV-associated GC [46,47]. BARF-1 was shown to be expressed in tissues of various EBV-associated epithelioid malignancies, but not in those of lymphoid malignancies. Using a specialized BARF-1-nucleic acid sequence-based amplification assay (NASBA), it has been demonstrated that BARF-1 exists in all EBV-associated GC tissues [46,48]. Furthermore, it was shown that recombinant expression of BARF-1 induced tumorigenic transformation of mouse fibroblasts and tumor formation in newborn rats [49]. Reconstitution of an NPC-type EBV infection using NPC-derived cell lines demonstrated that BARF-1 contributes to the tumorigenicity of NPC cells [50]. BARF-1 enhances the tumorigenicity of EBV-negative B-lymphocyte-derived cell lines [51,52], inhibits apoptosis by activating bcl-2 [53], and induces cell cycle activation [54,55]. BARF-1 has sequence homology with the human colony stimulating factor-1 receptor (CSF-1), which is the gene product of the human proto-oncogene, c-fms [56]. It has been suggested that CSF-1 and its receptor are involved in the tumorigenicity of epithelial cells, as increased expression is observed in GC as well as other carcinomas [50,57,58]. BARF-1 is secreted by EBV-carrying B cells upon the induction of lytic infection and it binds to CSF-1, inhibiting the binding of CSF-1 to the CSF-1 receptor. This leads to inhibition of IFN-α secretion and modulates the fates of immune-related cells such as macrophages [56,59,60]. Secreted BARF-1 can upregulate nuclear factor κB (NFκB) in an autocrine and paracrine manner in GC [47]. BARF-1-expressing GC cells displayed a high rate of proliferation, high levels of NFκB, and miR-146a, which can be reversed by NFκB knockdown [47]. Silencing BARF-1 upregulates the expression of pro-apoptotic proteins and downregulates the expression of anti-apoptotic proteins [61].
In the BART region, a cluster of 22 miRNAs precursors have been described [62]. These miRNAs generate 40 out of 44 mature miRNAs encoded by EBV (Figure 1) [62,63,64]. miR-BARTs show higher overall expression levels in EBV-infected epithelial cancers in comparison to EBV-infected lymphoblastoid cell lines and Burkitt’s lymphoma [65]. It is well established that these viral miRNAs modulate the host inflammatory response and favor EBV evasion, facilitating the maintenance of the latent infection and contributing to carcinogenesis [66,67]. These findings propose that miR-BARTs are key players in epithelial malignancies such as EBV-associated GC [65]. A comprehensive review of miR-BARTs and their function in EBV-associated GC has recently been published [68]. Taken together, it seems plausible that not only coding genes such as BARF-1, but also noncoding genes (i.e., miR-BARTs), may act as a potential source of variability in EBV-associated GC. Therefore, GWAS studies of the EBV genome, as well as targeted sequencing of the BARF-1 and BARTs transcripts in EBV-associated GC and the healthy population, will be essential for expanding our understanding of viral diversity.
4. Human Ancestry in the Americas and EBV-Associated Gastric Carcinoma
The complex demographic diversity of the Americas stems from many different waves of migration. In this regard, three major ethnicities contributed to the genetic makeup of this population: Amerindians, Europeans, and Africans [34,69,70]. A variety of approaches have been used to estimate the complexity of genetic ancestry in the Americas tracing back 15,000 years to the first waves of Asian-derived Amerindian migrations across Beringia [34,71,72]. This was followed by European migration during the mid-sixteenth century, initiating the genetic mixing of these populations [73]. The African component was primarily introduced through the slave trade during the seventeenth century, adding more complexity to the demographic diversity of the Americas [34].
Thus, the so-called New World provides a unique opportunity for uncovering the genetic basis of diseases [37,70]. In particular, differences in the incidences and mortality rates of GC according to ancestry have shown a disproportionate burden within the Amerindian populations [74,75]. Specific examples are the indigenous Inuit and Mapuche populations [76] who inhabit the Northern Arctic regions and Patagonia, respectively. In the case of the Inuit, the incidence of GC has increased significantly in recent decades despite a decline in the global mortality rate during the same time period [77,78,79,80,81]. Mapuche ancestry appears to be a major risk factor for GC in Chile, a country with one of the highest age-standardized incidence rates of the disease worldwide (23/100,000 inhabitants) [82]. Other examples include Peru and Colombia, where a positive association between Amerindian ancestry and GC has also been described [34,83,84]. In Brazil, da Silva and colleagues [85] reported the occurrence of GC in the Amazon region, which features an admixed population. Using a case-control design and multiple logistic regression analysis, these authors found that for every 10% increase in European ancestry, there is a 20% decrease in the probability of developing GC (p = 0.01; OR = 0.81; 95% CI 0.54–0.88), suggesting that European ancestry may be a protective factor for this disease. This information correlates with the fact that European countries tend to report lower incidence rates of GC than Asia or the Americas [82]. Thus, these examples highlight the major role of Amerindian ancestry in the occurrence of GC.
In the case of EBV-associated GC, the combination of phylogeographic polymorphisms of EBV and mixed ancestries in the Americas requires further exploration. As shown in Figure 2, in a healthy population, the human admixed heritage in Chile reflects European (48.5%) and Asian-derived Amerindian (49.9%) with a minor African component (1.6%) [37].
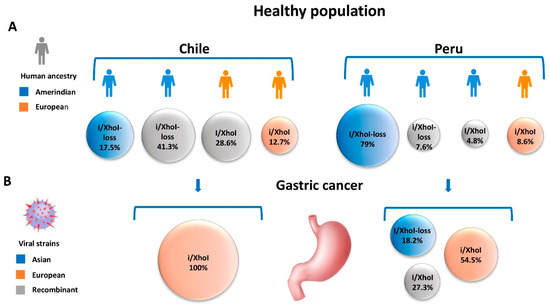
Figure 2.
Human heritage and EBV strains in the healthy population and EBV-associated GC in Chile and Peru. (A) Human ancestries and Asian, European and recombinant viral strains identified in the healthy population of Chile and Peru. (B) Distribution and geographical origin of viral strains among EBV-associated GC patients in Chile and Peru, showing the predominance of the European BamHI-”i” fragment and XhoI restriction site strain. In the case of the healthy population, EBV strains were examined from throat washing specimens, and in the case of gastric cancer patients, from paraffin-embedded tumor specimens.
Accordingly, the EBV recombinant strains prevail (41.3% and 28.6%) and the cosegregated ancestral strains are found at a lower frequency (17.5% Asian and 12.7% European) [23]. However, in Peru, a country with a predominantly Amerindian population (75% Amerindian vs. ~25% European) [34], the Asian strain predominates (79.0%) with a small proportion of European and recombinants variants (8.6%, 7.6% and 4.8%, respectively) [35]. In the case of EBV-associated GC, the cosegregated European BamHI-”i” fragment and XhoI restriction site strain prevails in Chile and Peru (100% and 54.5%, respectively) [23,35]. The predominance of the European strain could be likely due to particular recombinations of genes located within the BamHI fragment and XhoI region–as is the case of BARF-1 and BARTs in an Amerindian ancestry host. The lack of ancestral coadaptation supports the proposal of a disrupted co-evolution in the case of EBV-associated GC.
5. Other Examples of “Disrupted Co-Evolution” in Cancer-Related Infectious Agents
The interaction between the phylogeography of EBV and human ancestry is not a unique feature of this virus. A disrupted co-evolution has been previously proposed for H. pylori and human papillomavirus (HPV) and their associated tumors (GC and cervical cancer, respectively) (for a review see [86]). H. pylori, a bacterium that chronically colonizes the gastric mucosa, coevolved with human migration patterns [87]. In the Americas, genomic data has revealed the rapid evolution of H. pylori over the last 500 years. Phylogenetic studies based on genomic strain data have shown that most of the strains cluster according to their country of origin, suggesting that subpopulations of H. pylori have evolved at an accelerated rate, in order to adapt to particular human ancestries [88,89]. This rapid adaptation has been associated with three genes encoding outer membrane proteins which are important for the attachment of the bacterium to the gastric mucosa [88]. This scenario might account for the regional variability of GC [88,89]. This observation was originally assessed by de Sablet et al. [90]. These authors analyzed the phylogeographic origin of H. pylori isolates from two locations in Colombia with strikingly different incidences of GC: Tuquerres, in the Andes mountains (150/100,000 inhabitants) and Tumaco, on the coast (6/100,000 inhabitants). Using Multi-Locus Sequence Typing (MLST), these authors found that 100% of the isolates from Tuquerres but only 34% from Tumaco were classified as hpEurope. In the latter location, the remaining 66% isolates were hpAfrica1. This finding mirrored the ethnic composition of the host in both locations, being admixture European-Amerindian ancestry in Tuquerres and African-European ancestry in Tumaco. Functional studies have shown that differential expression between the European and African strains is observed in virulence factors, such as cagA, vacA, and babB and were associated with increased gastric histologic lesions in human gastric samples [90].
HPV is a naked circular double-stranded DNA virus with more than 200 genotypes based on the genome sequence of L1 gene [91]. High-risk (HR) strains, HPV-16 and -18 are the most frequent HPV detected in cervical, anogenital, and some head and neck cancers. Subtypes and variants of HPV-16 cluster into five major branches of a phylogenetic tree: European (E), Asian/American (AA), East Asian (As), and two African (Af1 and Af2) [92,93]. In the case of HPV-18, subtypes and variants cluster into three major branches: African (Af), European (E), as well as Asian and American Indian (As + AI) [93]. A study of HR-HPV in an Italian cohort demonstrated that non-European variants of HPV-16, Af1 and AA, were found at an increased frequency in invasive lesions [94]. A separate study of female university students in the US showed that those infected with non-European HPV-16 variants were 6.5 times more likely to develop high-grade cervical intraepithelial neoplasia than those with European variants [95]. Based on the aforementioned observations found in H. pylori and HPV, Kodaman et al., [86] proposed the concept of “disrupted co-evolution” between the pathogen and its host as a contributor to the phylogeographic origin of disease. Here, we propose that this may also be the case for EBV-associated GC.
6. Are Phylogeographic Variations of Epstein–Barr Virus Relevant to Other EBV-Associated Diseases?
Disparities in the incidence of EBV-associated diseases, besides GC, vary greatly in different parts of the world [96]. EBV-associated epithelial cancers represent 80% of all EBV-associated malignancies; among these is NPC with an incidence of >120,000 new cases and >70,000 deaths [82]. NPC has a characterized geographical distribution, with a higher incidence in Southern China, Southeast Asia, and to a lesser extent, the Maghrebi Arabic regions of North Africa and the Northern Arctic [97]. Differences in the prevalence of different EBV type-1 and -2 strains, as well as BamHI-F region, have been observed in NPC [17]. Studies in Portugal, Hong Kong, and China reported that EBV type-1 and prototype F were the most prevalent [98,99,100]. Of note, the study from Portugal showed that type-2 and variant “f” were significantly associated with NPC (p = 0.019; RR = 8.90). The XhoI loss variant was present in most of the NPC cases [17]. It proved particularly high in countries such as China, Malaysia, and Taiwan with more than 80% of the cases presenting this variant [17,101,102,103]. On the contrary, the presence of the XhoI site is found in NPCs from North Africa [104], confirming the phylogeographic distribution of this polymorphism. Another variant of the LMP-1 gene is the 30 bp-deletion (del-LMP-1) which seems to be more prevalent in NPC patients than healthy individuals in China, Malaysia, Hong-Kong, Taiwan, Tunisia, and Morocco [17,99,102,104,105,106,107]. However, in Portugal, 100% of NPC patients exhibited the wt-LPM1 variant [17]. A recent meta-analysis by da Costa et al. [105] confirmed the association between the 30-bp del-LMP-1 and XhoI loss with NPC susceptibility, although they found no association when analyzing the cosegregation of these variants in NPC patients.
Hodgkin lymphoma (HL) is a disease that can also exhibit the presence of EBV. It presents two distinct disease entities, the commonly diagnosed classical Hodgkin lymphoma (CHL) and the uncommon nodular lymphocyte-predominant Hodgkin lymphoma [108,109]. EBV is found in only a proportion of CHL cases, but in tropical regions, up to 100% of the population is EBV-positive [108]. HL accounts for ~80,000 new cases and >26,000 deaths, according to a recent global report [82]. The prevalence of EBV in CHL differs according to age, sex, region, histologic subtype, and clinical stage, as confirmed by a meta-analysis [110]. According to this study, the reported prevalence of EBV infection in CHL was 47.9%, with a significantly higher rate in Africa, Central America, and South America. EBV-positive CHL showed a higher incidence in children than in adults and was also significantly related to male gender (OR = 1.8, 95% CI: 1.510–2.038; p < 0.001) [110]. Regarding the common EBV variants distribution for HL among populations, it appears that EBV type-1 is the most prevalent, which is similar to what was described for NPC [17]. Studies in China, Korea, Spain, Denmark, and Australia showed that EBV type-1 was the most prevalent type among HL patients [17,111,112,113,114,115]. Moreover, type-1 was also the predominant strain in EBV variants in South American CHL, accounting for 78% Argentine and 86% Brazilian cases [116]. In the case of the XhoI loss, this variant was found to be predominant in EBV-associated Hodgkin’s disease cases and in the healthy Chinese population [111] confirming the phylogeographic distribution of this polymorphism. For del-LMP-1 variant distribution among populations, these are similar between China and Korea with more than 80% of the cases harboring the variant [17,111,113]. Similarly, in the case of South America, a higher frequency of the del-LMP-1 variant was observed in lymphomas (65%) than in non-neoplastic controls (27%) (OR 4.97, CI 95% 1.53–16.79; p = 0.005) [116]. Although phylogeographic variants of EBV seems to be relevant also in other EBV-associated diseases, such as NPC and HL, the del-LMP-1 variant has shown no differences in the case of EBV-associated GC [117].
7. Conclusions
Ancestral and recombinant strains of EBV in the Americas mirror the human genetic ancestry among the healthy population. However, there is a predominance of the European origin strain, based on the cosegregation of BamHI- I fragment and XhoI restriction site strain, in the case of EBV-associated GC. This observation proposes that a “disrupted co-evolution” might explain the high prevalence of EBV-associated GC in the Americas. Variations of two relevant transcripts, the BARF-1 and the miR-BARTs in this region might be associated with this high prevalence. Further studies are essential to expand our understanding of the phylogeographical diversity of EBV.
Funding
This research was funded by CONICYT-FONDAP 15130011 and Fondecyt 1151411, 1161219, 3160592 and 11181330.
Acknowledgments
We would like to thank Bree Johnson for English proofreading and editing the manuscript.
Conflicts of Interest
The authors declare no conflict of interest. The funders had no role in the design of the study; in the collection, analyses, or interpretation of data; in the writing of the manuscript, or in the decision to publish the results.
References
- The Cancer Genome Atlas Consortium. Comprehensive molecular characterization of gastric adenocarcinoma. Nature 2014, 513, 202–209. [Google Scholar] [CrossRef] [PubMed]
- Nishikawa, J.; Iizasa, H.; Yoshiyama, H.; Shimokuri, K.; Kobayashi, Y.; Sasaki, S.; Nakamura, M.; Yanai, H.; Sakai, K.; Suehiro, Y.; et al. Clinical Importance of Epstein(-)Barr Virus-Associated Gastric Cancer. Cancers 2018, 10, 167. [Google Scholar] [CrossRef] [PubMed]
- Sousa, H.; Pinto-Correia, A.L.; Medeiros, R.; Dinis-Ribeiro, M. Epstein-Barr virus is associated with gastric carcinoma: The question is what is the significance? World J. Gastroenterol. 2008, 14, 4347–4351. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.H.; Kim, S.H.; Han, S.H.; An, J.S.; Lee, E.S.; Kim, Y.S. Clinicopathological and molecular characteristics of Epstein-Barr virus-associated gastric carcinoma: A meta-analysis. J. Gastroenterol. Hepatol. 2009, 24, 354–365. [Google Scholar] [CrossRef] [PubMed]
- Murphy, G.; Pfeiffer, R.; Camargo, M.C.; Rabkin, C.S. Meta-analysis shows that prevalence of Epstein-Barr virus-positive gastric cancer differs based on sex and anatomic location. Gastroenterology 2009, 137, 824–833. [Google Scholar] [CrossRef] [PubMed]
- Li, S.; Du, H.; Wang, Z.; Zhou, L.; Zhao, X.; Zeng, Y. Meta-analysis of the relationship between Epstein-Barr virus infection and clinicopathological features of patients with gastric carcinoma. Science China. Life Sci. 2010, 53, 524–530. [Google Scholar] [CrossRef] [PubMed]
- Camargo, M.C.; Murphy, G.; Koriyama, C.; Pfeiffer, R.M.; Kim, W.H.; Herrera-Goepfert, R.; Corvalan, A.H.; Carrascal, E.; Abdirad, A.; Anwar, M.; et al. Determinants of Epstein-Barr virus-positive gastric cancer: An international pooled analysis. Br. J. Cancer 2011, 105, 38–43. [Google Scholar] [CrossRef] [PubMed]
- Koriyama, C.; Akiba, S.; Iriya, K.; Yamaguti, T.; Hamada, G.S.; Itoh, T.; Eizuru, Y.; Aikou, T.; Watanabe, S.; Tsugane, S.; et al. Epstein-Barr virus-associated gastric carcinoma in Japanese Brazilians and non-Japanese Brazilians in Sao Paulo. Jpn. J. Cancer res. 2001, 92, 911–917. [Google Scholar] [CrossRef]
- Gulley, M.L.; Pulitzer, D.R.; Eagan, P.A.; Schneider, B.G. Epstein-Barr virus infection is an early event in gastric carcinogenesis and is independent of bcl-2 expression and p53 accumulation. Hum. Pathol. 1996, 27, 20–27. [Google Scholar] [CrossRef]
- Vo, Q.N.; Geradts, J.; Gulley, M.L.; Boudreau, D.A.; Bravo, J.C.; Schneider, B.G. Epstein-Barr virus in gastric adenocarcinomas: Association with ethnicity and CDKN2A promoter methylation. J. Clin. Pathol. 2002, 55, 669–675. [Google Scholar] [CrossRef]
- Alarcon, A.; Figueroa, U.; Espinoza, B.; Sandoval, A.; Carrasco-Aviño, G.; Aguayo, F.R.; Corvalan, A.H. Epstein-Barr Virus–Associated Gastric Carcinoma: The Americas’ Perspective; Lunet, N., Ed.; Intech: London, UK, 2017. [Google Scholar] [CrossRef]
- Baer, R.; Bankier, A.T.; Biggin, M.D.; Deininger, P.L.; Farrell, P.J.; Gibson, T.J.; Hatfull, G.; Hudson, G.S.; Satchwell, S.C.; Seguin, C.; et al. DNA sequence and expression of the B95-8 Epstein-Barr virus genome. Nature 1984, 310, 207–211. [Google Scholar] [CrossRef] [PubMed]
- de Jesus, O.; Smith, P.R.; Spender, L.C.; Elgueta Karstegl, C.; Niller, H.H.; Huang, D.; Farrell, P.J. Updated Epstein-Barr virus (EBV) DNA sequence and analysis of a promoter for the BART (CST, BARF0) RNAs of EBV. J. Gen. Virol. 2003, 84, 1443–1450. [Google Scholar] [CrossRef] [PubMed]
- Lei, H.; Li, T.; Hung, G.C.; Li, B.; Tsai, S.; Lo, S.C. Identification and characterization of EBV genomes in spontaneously immortalized human peripheral blood B lymphocytes by NGS technology. BMC Genom. 2013, 14, 804. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Yang, W.; Pan, Y.; Ji, J.; Lu, Z.; Ke, Y. Genome-wide analysis of Epstein-Barr virus (EBV) isolated from EBV-associated gastric carcinoma (EBVaGC). Oncotarget 2015, 7, 4903. [Google Scholar] [CrossRef] [PubMed]
- Santpere, G.; Darre, F.; Blanco, S.; Alcami, A.; Villoslada, P.; Mar Alba, M.; Navarro, A. Genome-wide analysis of wild-type Epstein-Barr virus genomes derived from healthy individuals of the 1,000 Genomes Project. Genome Biol. Evol. 2014, 6, 846–860. [Google Scholar] [CrossRef]
- Neves, M.; Marinho-Dias, J.; Ribeiro, J.; Sousa, H. Epstein-Barr virus strains and variations: Geographic or disease-specific variants? J. Med. Virol. 2017, 89, 373–387. [Google Scholar] [CrossRef] [PubMed]
- Sample, J.; Young, L.; Martin, B.; Chatman, T.; Kieff, E.; Rickinson, A.; Kieff, E. Epstein-Barr virus types 1 and 2 differ in their EBNA-3A, EBNA-3B, and EBNA-3C genes. J. Virol. 1990, 64, 4084–4092. [Google Scholar]
- Zimber, U.; Adldinger, H.K.; Lenoir, G.M.; Vuillaume, M.; Knebel-Doeberitz, M.V.; Laux, G.; Desgranges, C.; Wittmann, P.; Freese, U.K.; Schneider, U.; et al. Geographical prevalence of two types of Epstein-Barr virus. Virology 1986, 154, 56–66. [Google Scholar] [CrossRef]
- Young, L.S.; Yao, Q.Y.; Rooney, C.M.; Sculley, T.B.; Moss, D.J.; Rupani, H.; Laux, G.; Bornkamm, G.W.; Rickinson, A.B. New type B isolates of Epstein-Barr virus from Burkitt’s lymphoma and from normal individuals in endemic areas. J. Gen. Virol. 1987, 68 Pt 11, 2853–2862. [Google Scholar] [CrossRef]
- Young, L.S.; Murray, P.G. Epstein-Barr virus and oncogenesis: From latent genes to tumours. Oncogene 2003, 22, 5108–5121. [Google Scholar] [CrossRef]
- Hudson, G.S.; Gibson, T.J.; Barrell, B.G. The BamHI F region of the B95-8 Epstein-Barr virus genome. Virology 1985, 147, 99–109. [Google Scholar] [CrossRef]
- Corvalan, A.H.; Ding, S.; Koriyama, C.; Carrascal, E.; Carrasquilla, G.; Backhouse, C.; Urzua, L.; Argandona, J.; Palma, M.; Eizuru, Y.; et al. Association of a distinctive strain of Epstein-Barr virus with gastric cancer. Int. J. Cancer 2006, 118, 1736–1742. [Google Scholar] [CrossRef] [PubMed]
- Lung, M.L.; Lam, W.P.; Sham, J.; Choy, D.; Yong-Sheng, Z.; Guo, H.Y.; Ng, M.H. Detection and prevalence of the “f” variant of Epstein-Barr virus in southern China. Virology 1991, 185, 67–71. [Google Scholar] [CrossRef]
- Lung, M.L.; Chang, R.S.; Huang, M.L.; Guo, H.Y.; Choy, D.; Sham, J.; Tsao, S.Y.; Cheng, P.; Ng, M.H. Epstein-Barr virus genotypes associated with nasopharyngeal carcinoma in southern China. Virology 1990, 177, 44–53. [Google Scholar] [CrossRef]
- Chen, J.N.; Ding, Y.G.; Feng, Z.Y.; Li, H.G.; He, D.; Du, H.; Wu, B.; Shao, C.K. Association of distinctive Epstein-Barr virus variants with gastric carcinoma in Guangzhou, southern China. J. Med. Virol. 2010, 82, 658–667. [Google Scholar] [CrossRef] [PubMed]
- Cui, Y.; Wang, Y.; Liu, X.; Chao, Y.; Xing, X.; Zhao, C.; Liu, C.; Luo, B. Genotypic analysis of Epstein-Barr virus isolates associated with nasopharyngeal carcinoma in Northern China. Intervirology 2011, 54, 131–138. [Google Scholar] [CrossRef]
- Lung, M.L.; Chang, G.C. Detection of distinct Epstein-Barr virus genotypes in NPC biopsies from southern Chinese and Caucasians. Int. J. Cancer 1992, 52, 34–37. [Google Scholar] [CrossRef]
- Lung, M.L.; Chang, R.S.; Jones, J.H. Genetic polymorphism of natural Epstein-Barr virus isolates from infectious mononucleosis patients and healthy carriers. J. Virol. 1988, 62, 3862–3866. [Google Scholar]
- Sidagis, J.; Ueno, K.; Tokunaga, M.; Ohyama, M.; Eizuru, Y. Molecular epidemiology of Epstein-Barr virus (EBV) in EBV-related malignancies. Int. J. Cancer 1997, 72, 72–76. [Google Scholar] [CrossRef]
- Abdel-Hamid, M.; Chen, J.J.; Constantine, N.; Massoud, M.; Raab-Traub, N. EBV strain variation: Geographical distribution and relation to disease state. Virology 1992, 190, 168–175. [Google Scholar] [CrossRef]
- Hu, L.F.; Zabarovsky, E.R.; Chen, F.; Cao, S.L.; Ernberg, I.; Klein, G.; Winberg, G. Isolation and sequencing of the Epstein-Barr virus BNLF-1 gene (LMP1) from a Chinese nasopharyngeal carcinoma. J. Gen. Virol. 1991, 72 Pt 10, 2399–2409. [Google Scholar] [CrossRef]
- Khanim, F.; Yao, Q.Y.; Niedobitek, G.; Sihota, S.; Rickinson, A.B.; Young, L.S. Analysis of Epstein-Barr virus gene polymorphisms in normal donors and in virus-associated tumors from different geographic locations. Blood 1996, 88, 3491–3501. [Google Scholar] [PubMed]
- Adhikari, K.; Chacon-Duque, J.C.; Mendoza-Revilla, J.; Fuentes-Guajardo, M.; Ruiz-Linares, A. The Genetic Diversity of the Americas. Annu. Rev. Genom. Hum. Genet. 2017, 18, 277–296. [Google Scholar] [CrossRef] [PubMed]
- Ordonez, P.; Koriyama, C.; Ding, S.; Yoshiwara, E.; Corvalan, A.H.; Takano, J.; Chirinos, J.L.; Watanabe, J.; Miyagui, J.; Hidalgo, H.; et al. Identification of the distinctive type i/XhoI+ strain of Epstein-Barr virus in gastric carcinoma in Peru. Anticancer Res. 2011, 31, 3607–3613. [Google Scholar] [PubMed]
- Sans, M. Admixture studies in Latin America: From the 20th to the 21st century. Hum. Boil. 2000, 72, 155–177. [Google Scholar]
- Eyheramendy, S.; Martinez, F.I.; Manevy, F.; Vial, C.; Repetto, G.M. Genetic structure characterization of Chileans reflects historical immigration patterns. Nat. Commun. 2015, 6, 6472. [Google Scholar] [CrossRef] [PubMed]
- Halliburton, I.W. Intertypic recombinants of herpes simplex viruses. J. Gen. Virol. 1980, 48, 1–23. [Google Scholar] [CrossRef]
- Young, L.S.; Yap, L.F.; Murray, P.G. Epstein-Barr virus: More than 50 years old and still providing surprises. Nature reviews. Cancer 2016, 16, 789–802. [Google Scholar] [CrossRef]
- Barth, S.; Pfuhl, T.; Mamiani, A.; Ehses, C.; Roemer, K.; Kremmer, E.; Jaker, C.; Hock, J.; Meister, G.; Grasser, F.A. Epstein-Barr virus-encoded microRNA miR-BART2 down-regulates the viral DNA polymerase BALF5. Nucleic Acids Res. 2008, 36, 666–675. [Google Scholar] [CrossRef]
- Yao, Y.; Xu, M.; Liang, L.; Zhang, H.; Xu, R.; Feng, Q.; Feng, L.; Luo, B.; Zeng, Y.X. Genome-wide analysis of Epstein-Barr virus identifies variants and genes associated with gastric carcinoma and population structure. Tumour Boil. 2017, 39, 1010428317714195. [Google Scholar] [CrossRef]
- Price, A.M.; Luftig, M.A. To be or not IIb: A multi-step process for Epstein-Barr virus latency establishment and consequences for B cell tumorigenesis. PLoS Pathog. 2015, 11, e1004656. [Google Scholar] [CrossRef] [PubMed]
- Gilligan, K.; Sato, H.; Rajadurai, P.; Busson, P.; Young, L.; Rickinson, A.; Tursz, T.; Raab-Traub, N. Novel transcription from the Epstein-Barr virus terminal EcoRI fragment, DIJhet, in a nasopharyngeal carcinoma. J. Virol. 1990, 64, 4948–4956. [Google Scholar] [PubMed]
- Sadler, R.H.; Raab-Traub, N. Structural analyses of the Epstein-Barr virus BamHI A transcripts. J. Virol. 1995, 69, 1132–1141. [Google Scholar] [PubMed]
- Hayes, D.P.; Brink, A.A.; Vervoort, M.B.; Middeldorp, J.M.; Meijer, C.J.; van den Brule, A.J. Expression of Epstein-Barr virus (EBV) transcripts encoding homologues to important human proteins in diverse EBV associated diseases. Mol. Pathol. 1999, 52, 97–103. [Google Scholar] [CrossRef] [PubMed]
- zur Hausen, A.; Brink, A.A.; Craanen, M.E.; Middeldorp, J.M.; Meijer, C.J.; van den Brule, A.J. Unique transcription pattern of Epstein-Barr virus (EBV) in EBV-carrying gastric adenocarcinomas: Expression of the transforming BARF1 gene. Cancer Res. 2000, 60, 2745–2748. [Google Scholar] [PubMed]
- Chang, M.S.; Kim, D.H.; Roh, J.K.; Middeldorp, J.M.; Kim, Y.S.; Kim, S.; Han, S.; Kim, C.W.; Lee, B.L.; Kim, W.H.; et al. Epstein-Barr virus-encoded BARF1 promotes proliferation of gastric carcinoma cells through regulation of NF-kappaB. J. Virol. 2013, 87, 10515–10523. [Google Scholar] [CrossRef] [PubMed]
- Brink, A.A.; Vervoort, M.B.; Middeldorp, J.M.; Meijer, C.J.; van den Brule, A.J. Nucleic acid sequence-based amplification, a new method for analysis of spliced and unspliced Epstein-Barr virus latent transcripts, and its comparison with reverse transcriptase PCR. J. Clin. Microbial. 1998, 36, 3164–3169. [Google Scholar]
- Wei, M.X.; Ooka, T. A transforming function of the BARF1 gene encoded by Epstein-Barr virus. EMBO J. 1989, 8, 2897–2903. [Google Scholar] [CrossRef]
- Takada, K. Role of EBER and BARF1 in nasopharyngeal carcinoma (NPC) tumorigenesis. Semin. Cancer Biol. 2012, 22, 162–165. [Google Scholar] [CrossRef]
- Wei, M.X.; Moulin, J.C.; Decaussin, G.; Berger, F.; Ooka, T. Expression and tumorigenicity of the Epstein-Barr virus BARF1 gene in human Louckes B-lymphocyte cell line. Cancer Res. 1994, 54, 1843–1848. [Google Scholar]
- Sheng, W.; Decaussin, G.; Ligout, A.; Takada, K.; Ooka, T. Malignant transformation of Epstein-Barr virus-negative Akata cells by introduction of the BARF1 gene carried by Epstein-Barr virus. J. Virol. 2003, 77, 3859–3865. [Google Scholar] [CrossRef] [PubMed]
- Wang, Q.; Tsao, S.W.; Ooka, T.; Nicholls, J.M.; Cheung, H.W.; Fu, S.; Wong, Y.C.; Wang, X. Anti-apoptotic role of BARF1 in gastric cancer cells. Cancer Lett. 2006, 238, 90–103. [Google Scholar] [CrossRef] [PubMed]
- Sall, A.; Caserta, S.; Jolicoeur, P.; Franqueville, L.; de Turenne-Tessier, M.; Ooka, T. Mitogenic activity of Epstein-Barr virus-encoded BARF1 protein. Oncogene 2004, 23, 4938–4944. [Google Scholar] [CrossRef] [PubMed]
- Wiech, T.; Nikolopoulos, E.; Lassman, S.; Heidt, T.; Schopflin, A.; Sarbia, M.; Werner, M.; Shimizu, Y.; Sakka, E.; Ooka, T.; et al. Cyclin D1 expression is induced by viral BARF1 and is overexpressed in EBV-associated gastric cancer. Virchows Arch. 2008, 452, 621–627. [Google Scholar] [CrossRef] [PubMed]
- Strockbine, L.D.; Cohen, J.I.; Farrah, T.; Lyman, S.D.; Wagener, F.; DuBose, R.F.; Armitage, R.J.; Spriggs, M.K. The Epstein-Barr virus BARF1 gene encodes a novel, soluble colony-stimulating factor-1 receptor. J. Virol. 1998, 72, 4015–4021. [Google Scholar] [PubMed]
- Sapi, E.; Flick, M.B.; Gilmore-Hebert, M.; Rodov, S.; Kacinski, B.M. Transcriptional regulation of the c-fms (CSF-1R) proto-oncogene in human breast carcinoma cells by glucocorticoids. Oncogene 1995, 10, 529–542. [Google Scholar] [PubMed]
- Lin, E.Y.; Nguyen, A.V.; Russell, R.G.; Pollard, J.W. Colony-stimulating factor 1 promotes progression of mammary tumors to malignancy. J. Exp. Med. 2001, 193, 727–740. [Google Scholar] [CrossRef]
- Hoebe, E.K.; Le Large, T.Y.; Tarbouriech, N.; Oosterhoff, D.; De Gruijl, T.D.; Middeldorp, J.M.; Greijer, A.E. Epstein-Barr virus-encoded BARF1 protein is a decoy receptor for macrophage colony stimulating factor and interferes with macrophage differentiation and activation. Viral Immunol. 2012, 25, 461–470. [Google Scholar] [CrossRef]
- Hoebe, E.K.; Le Large, T.Y.; Greijer, A.E.; Middeldorp, J.M. BamHI-A rightward frame 1, an Epstein-Barr virus-encoded oncogene and immune modulator. Rev. Med. Virol. 2013, 23, 367–383. [Google Scholar] [CrossRef]
- Mohidin, T.B.; Ng, C.C. BARF1 gene silencing triggers caspase-dependent mitochondrial apoptosis in Epstein-Barr virus-positive malignant cells. J. Biosci. 2015, 40, 41–51. [Google Scholar] [CrossRef]
- Cai, X.; Schafer, A.; Lu, S.; Bilello, J.P.; Desrosiers, R.C.; Edwards, R.; Raab-Traub, N.; Cullen, B.R. Epstein-Barr virus microRNAs are evolutionarily conserved and differentially expressed. PLoS Pathog. 2006, 2, e23. [Google Scholar] [CrossRef] [PubMed]
- Grundhoff, A.; Sullivan, C.S.; Ganem, D. A combined computational and microarray-based approach identifies novel microRNAs encoded by human gamma-herpesviruses. RNA 2006, 12, 733–750. [Google Scholar] [CrossRef] [PubMed]
- Zhu, J.Y.; Pfuhl, T.; Motsch, N.; Barth, S.; Nicholls, J.; Grasser, F.; Meister, G. Identification of novel Epstein-Barr virus microRNA genes from nasopharyngeal carcinomas. J. Virol. 2009, 83, 3333–3341. [Google Scholar] [CrossRef] [PubMed]
- Qiu, J.; Cosmopoulos, K.; Pegtel, M.; Hopmans, E.; Murray, P.; Middeldorp, J.; Shapiro, M.; Thorley-Lawson, D.A. A novel persistence associated EBV miRNA expression profile is disrupted in neoplasia. PLoS Pathog. 2011, 7, e1002193. [Google Scholar] [CrossRef] [PubMed]
- Seto, E.; Moosmann, A.; Gromminger, S.; Walz, N.; Grundhoff, A.; Hammerschmidt, W. Micro RNAs of Epstein-Barr virus promote cell cycle progression and prevent apoptosis of primary human B cells. PLoS Pathog. 2010, 6, e1001063. [Google Scholar] [CrossRef] [PubMed]
- Albanese, M.; Tagawa, T. MicroRNAs of Epstein-Barr Virus Control Innate and Adaptive Antiviral Immunity. J. Virol. 2017, 91, e01667-16. [Google Scholar] [CrossRef] [PubMed]
- Polakovicova, I.; Jerez, S.; Wichmann, I.A.; Sandoval-Borquez, A.; Carrasco-Veliz, N.; Corvalan, A.H. Role of microRNAs and Exosomes in Helicobacter pylori and Epstein-Barr Virus Associated Gastric Cancers. Front. Microbiol. 2018, 9, 636. [Google Scholar] [CrossRef]
- Homburger, J.R.; Moreno-Estrada, A.; Gignoux, C.R.; Nelson, D.; Sanchez, E.; Ortiz-Tello, P.; Pons-Estel, B.A.; Acevedo-Vasquez, E.; Miranda, P.; Langefeld, C.D.; et al. Genomic Insights into the Ancestry and Demographic History of South America. PLoS Genet. 2015, 11, e1005602. [Google Scholar] [CrossRef]
- Moreno-Estrada, A.; Gignoux, C.R.; Fernandez-Lopez, J.C.; Zakharia, F.; Sikora, M.; Contreras, A.V.; Acuna-Alonzo, V.; Sandoval, K.; Eng, C.; Romero-Hidalgo, S.; et al. Human genetics. The genetics of Mexico recapitulates Native American substructure and affects biomedical traits. Science 2014, 344, 1280–1285. [Google Scholar] [CrossRef]
- Shriner, D. Overview of Admixture Mapping. Curr. Protoc. Hum. Genet. 2017, 94, 1–23. [Google Scholar] [CrossRef]
- Moreno-Mayar, J.V.; Vinner, L.; de Barros Damgaard, P.; de la Fuente, C.; Chan, J.; Spence, J.P.; Allentoft, M.E.; Vimala, T.; Racimo, F.; Pinotti, T.; et al. Early human dispersals within the Americas. Science 2018. [Google Scholar] [CrossRef] [PubMed]
- Bryc, K.; Velez, C.; Karafet, T.; Moreno-Estrada, A.; Reynolds, A.; Auton, A.; Hammer, M.; Bustamante, C.D.; Ostrer, H. Colloquium paper: Genome-wide patterns of population structure and admixture among Hispanic/Latino populations. Proc. Natl. Acad. Sci. USA 2010, 107 (Suppl. 2), 8954–8961. [Google Scholar] [CrossRef] [PubMed]
- Ashktorab, H.; Kupfer, S.S.; Brim, H.; Carethers, J.M. Racial Disparity in Gastrointestinal Cancer Risk. Gastroenterology 2017, 153, 910–923. [Google Scholar] [CrossRef] [PubMed]
- Dong, E.; Duan, L.; Wu, B.U. Racial and Ethnic Minorities at Increased Risk for Gastric Cancer in a Regional US Population Study. Clin. Gastroenterol. Hepatol. 2017, 15, 511–517. [Google Scholar] [CrossRef] [PubMed]
- Arnold, M.; Moore, S.P.; Hassler, S.; Ellison-Loschmann, L.; Forman, D.; Bray, F. The burden of stomach cancer in indigenous populations: A systematic review and global assessment. Gut 2014, 63, 64–71. [Google Scholar] [CrossRef] [PubMed]
- Friborg, J.T.; Melbye, M. Cancer patterns in Inuit populations. Lancet Oncol. 2008, 9, 892–900. [Google Scholar] [CrossRef]
- Circumpolar Inuit Cancer Review Working Group; Kelly, J.; Lanier, A.; Santos, M.; Healey, S.; Louchini, R.; Friborg, J.; Young, K.; Ng, C. Cancer among the circumpolar Inuit, 1989-2003. II. Patterns and trends. Int. J. Circumpolar Health 2008, 67, 408–420. [Google Scholar]
- Amiri, M.; Janssen, F.; Kunst, A.E. The decline in stomach cancer mortality: Exploration of future trends in seven European countries. Eur. J. Epidemiol. 2011, 26, 23–28. [Google Scholar] [CrossRef]
- Siegel, R.; Naishadham, D.; Jemal, A. Cancer statistics, 2013. CA: A Cancer J. Clin. 2013, 63, 11–30. [Google Scholar] [CrossRef]
- Mathias, R.A.; Taub, M.A.; Gignoux, C.R.; Fu, W.; Musharoff, S.; O’Connor, T.D.; Vergara, C.; Torgerson, D.G.; Pino-Yanes, M.; Shringarpure, S.S.; et al. A continuum of admixture in the Western Hemisphere revealed by the African Diaspora genome. Nat. Commun. 2016, 7, 12522. [Google Scholar] [CrossRef]
- Bray, F.; Ferlay, J.; Soerjomataram, I.; Siegel, R.L.; Torre, L.A.; Jemal, A. Global cancer statistics 2018: GLOBOCAN estimates of incidence and mortality worldwide for 36 cancers in 185 countries. CA: A Cancer J. Clin. 2018, 68, 394–424. [Google Scholar] [CrossRef] [PubMed]
- Moore, S.P.; Forman, D.; Pineros, M.; Fernandez, S.M.; de Oliveira Santos, M.; Bray, F. Cancer in indigenous people in Latin America and the Caribbean: A review. Cancer Med. 2014, 3, 70–80. [Google Scholar] [CrossRef] [PubMed]
- Pineros, M.; Ferlay, J.; Murillo, R. Cancer incidence estimates at the national and district levels in Colombia. Salud Publica Mex. 2006, 48, 455–465. [Google Scholar] [CrossRef] [PubMed]
- da Silva, E.M.; Fernandes, M.R.; de Carvalho, D.C.; Leitao, L.P.C.; Cavalcante, G.C.; Pereira, E.E.B.; Modesto, A.A.C.; Guerreiro, J.F.; de Assumpcao, P.P.; Dos Santos, S.E.B.; et al. Correction to: Effect of genetic ancestry to the risk of susceptibility to gastric cancer in a mixed population of the Brazilian Amazon. BMC Res. Notes 2017, 10, 761. [Google Scholar] [CrossRef] [PubMed]
- Kodaman, N.; Sobota, R.S.; Mera, R.; Schneider, B.G.; Williams, S.M. Disrupted human–pathogen co-evolution: A model for disease. Front. Genet. 2014, 5, 290. [Google Scholar] [CrossRef] [PubMed]
- Linz, B.; Balloux, F.; Moodley, Y.; Manica, A.; Liu, H.; Roumagnac, P.; Falush, D.; Stamer, C.; Prugnolle, F.; van der Merwe, S.W.; et al. An African origin for the intimate association between humans and Helicobacter pylori. Nature 2007, 445, 915–918. [Google Scholar] [CrossRef] [PubMed]
- Thorell, K.; Yahara, K.; Berthenet, E.; Lawson, D.J.; Mikhail, J.; Kato, I.; Mendez, A.; Rizzato, C.; Bravo, M.M.; Suzuki, R.; et al. Rapid evolution of distinct Helicobacter pylori subpopulations in the Americas. PLoS Genet. 2017, 13, e1006546. [Google Scholar] [CrossRef]
- Munoz-Ramirez, Z.Y.; Mendez-Tenorio, A.; Kato, I.; Bravo, M.M.; Rizzato, C.; Thorell, K.; Torres, R.; Aviles-Jimenez, F.; Camorlinga, M.; Canzian, F.; et al. Whole Genome Sequence and Phylogenetic Analysis Show Helicobacter pylori Strains from Latin America Have Followed a Unique Evolution Pathway. Front. Cell. Infect. Microbiol. 2017, 7, 50. [Google Scholar] [CrossRef]
- de Sablet, T.; Piazuelo, M.B.; Shaffer, C.L.; Schneider, B.G.; Asim, M.; Chaturvedi, R.; Bravo, L.E.; Sicinschi, L.A.; Delgado, A.G.; Mera, R.M.; et al. Phylogeographic origin of Helicobacter pylori is a determinant of gastric cancer risk. Gut 2011, 60, 1189–1195. [Google Scholar] [CrossRef]
- Van Doorslaer, K.; Chen, Z.; Bernard, H.U.; Chan, P.K.S.; DeSalle, R.; Dillner, J.; Forslund, O.; Haga, T.; McBride, A.A.; Villa, L.L.; et al. ICTV Virus Taxonomy Profile: Papillomaviridae. J. Gen. Virol. 2018, 99, 989–990. [Google Scholar] [CrossRef]
- Ho, L.; Chan, S.Y.; Burk, R.D.; Das, B.C.; Fujinaga, K.; Icenogle, J.P.; Kahn, T.; Kiviat, N.; Lancaster, W.; Mavromara-Nazos, P.; et al. The genetic drift of human papillomavirus type 16 is a means of reconstructing prehistoric viral spread and the movement of ancient human populations. J. Virol. 1993, 67, 6413–6423. [Google Scholar] [PubMed]
- Ong, C.K.; Chan, S.Y.; Campo, M.S.; Fujinaga, K.; Mavromara-Nazos, P.; Labropoulou, V.; Pfister, H.; Tay, S.K.; ter Meulen, J.; Villa, L.L.; et al. Evolution of human papillomavirus type 18: An ancient phylogenetic root in Africa and intratype diversity reflect coevolution with human ethnic groups. J. Virol. 1993, 67, 6424–6431. [Google Scholar] [PubMed]
- Tornesello, M.L.; Duraturo, M.L.; Salatiello, I.; Buonaguro, L.; Losito, S.; Botti, G.; Stellato, G.; Greggi, S.; Piccoli, R.; Pilotti, S.; et al. Analysis of human papillomavirus type-16 variants in Italian women with cervical intraepithelial neoplasia and cervical cancer. J. Med. Virol. 2004, 74, 117–126. [Google Scholar] [CrossRef] [PubMed]
- Xi, L.F.; Koutsky, L.A.; Galloway, D.A.; Kuypers, J.; Hughes, J.P.; Wheeler, C.M.; Holmes, K.K.; Kiviat, N.B. Genomic variation of human papillomavirus type 16 and risk for high grade cervical intraepithelial neoplasia. J. Natl. Cancer Inst. 1997, 89, 796–802. [Google Scholar] [CrossRef] [PubMed]
- Correia, S.; Palser, A.; Elgueta Karstegl, C.; Middeldorp, J.M.; Ramayanti, O.; Cohen, J.I.; Hildesheim, A.; Fellner, M.D.; Wiels, J.; White, R.E.; et al. Natural Variation of Epstein-Barr Virus Genes, Proteins, and Primary MicroRNA. J. Virol. 2017, 91. [Google Scholar] [CrossRef] [PubMed]
- Chang, C.M.; Yu, K.J.; Mbulaiteye, S.M.; Hildesheim, A.; Bhatia, K. The extent of genetic diversity of Epstein-Barr virus and its geographic and disease patterns: A need for reappraisal. Virus Res. 2009, 143, 209–221. [Google Scholar] [CrossRef] [PubMed]
- Neves, M.; Marinho-Dias, J.; Ribeiro, J.; Esteves, M.; Maltez, E.; Baldaque, I.; Breda, E.; Monteiro, E.; Medeiros, R.; Sousa, H. Characterization of Epstein-Barr virus strains and LMP1-deletion variants in Portugal. J. Med. Virol. 2015, 87, 1382–1388. [Google Scholar] [CrossRef] [PubMed]
- Cheung, S.T.; Leung, S.F.; Lo, K.W.; Chiu, K.W.; Tam, J.S.; Fok, T.F.; Johnson, P.J.; Lee, J.C.; Huang, D.P. Specific latent membrane protein 1 gene sequences in type 1 and type 2 Epstein-Barr virus from nasopharyngeal carcinoma in Hong Kong. Int. J. Cancer 1998, 76, 399–406. [Google Scholar] [CrossRef]
- Wu, S.J.; Lay, J.D.; Chen, C.L.; Chen, J.Y.; Liu, M.Y.; Su, I.J. Genomic analysis of Epstein-Barr virus in nasal and peripheral T-cell lymphoma: A comparison with nasopharyngeal carcinoma in an endemic area. J. Med. Virol. 1996, 50, 314–321. [Google Scholar] [CrossRef]
- Lin, S.X.; Zong, Y.S.; Zhang, M.; Han, A.J.; Zhong, B.L.; Liang, Y.J. Study of sequence variations of Epstein-Barr virus LMP1 gene in nasopharyngeal carcinoma. Zhonghua Bing Li Xue Za Zhi = Chin. J. Pathol. 2005, 34, 791–795. [Google Scholar]
- Tan, E.L.; Peh, S.C.; Sam, C.K. Analyses of Epstein-Barr virus latent membrane protein-1 in Malaysian nasopharyngeal carcinoma: High prevalence of 30-bp deletion, Xho1 polymorphism and evidence of dual infections. J. Med. Virol. 2003, 69, 251–257. [Google Scholar] [CrossRef] [PubMed]
- Chang, Y.S.; Su, I.J.; Chung, P.J.; Shu, C.H.; Ng, C.K.; Wu, S.J.; Liu, S.T. Detection of an Epstein-Barr-virus variant in T-cell-lymphoma tissues identical to the distinct strain observed in nasopharyngeal carcinoma in the Taiwanese population. Int. J. Cancer 1995, 62, 673–677. [Google Scholar] [CrossRef] [PubMed]
- Ayadi, W.; Feki, L.; Khabir, A.; Boudawara, T.; Ghorbel, A.; Charfeddine, I.; Daoud, J.; Frikha, M.; Hammami, A.; Karray-Hakim, H. Polymorphism analysis of Epstein-Barr virus isolates of nasopharyngeal carcinoma biopsies from Tunisian patients. Virus Genes 2007, 34, 137–145. [Google Scholar] [CrossRef] [PubMed]
- da Costa, V.G.; Marques-Silva, A.C.; Moreli, M.L. The Epstein-Barr virus latent membrane protein-1 (LMP1) 30-bp deletion and XhoI-polymorphism in nasopharyngeal carcinoma: A meta-analysis of observational studies. Syst. Rev. 2015, 4, 46. [Google Scholar] [CrossRef] [PubMed]
- Cheung, S.T.; Lo, K.W.; Leung, S.F.; Chan, W.Y.; Choi, P.H.; Johnson, P.J.; Lee, J.C.; Huang, D.P. Prevalence of LMP1 deletion variant of Epstein-Barr virus in nasopharyngeal carcinoma and gastric tumors in Hong Kong. Int. J. Cancer 1996, 66, 711–712. [Google Scholar] [CrossRef]
- Chang, K.P.; Hao, S.P.; Lin, S.Y.; Ueng, S.H.; Pai, P.C.; Tseng, C.K.; Hsueh, C.; Hsieh, M.S.; Yu, J.S.; Tsang, N.M. The 30-bp deletion of Epstein-Barr virus latent membrane protein-1 gene has no effect in nasopharyngeal carcinoma. Laryngoscope 2006, 116, 541–546. [Google Scholar] [CrossRef] [PubMed]
- Swerdlow, S.H.; Campo, E.; N.L., H.; Jaffe, E.S.; Pileri, S.A.; Stein, H.; Thiele, J.; Vardiman, J.W. WHO Classification of Tumours of Haematopoietic and Lymphoid Tissues; IARC: Lyon, France, 2008. [Google Scholar]
- Ansell, S.M. Hodgkin Lymphoma: Diagnosis and Treatment. Mayo Clin. Proc. 2015, 90, 1574–1583. [Google Scholar] [CrossRef]
- Lee, J.H.; Kim, Y.; Choi, J.W.; Kim, Y.S. Prevalence and prognostic significance of Epstein-Barr virus infection in classical Hodgkin’s lymphoma: A meta-analysis. Arch. Med. Res. 2014, 45, 417–431. [Google Scholar] [CrossRef]
- Zhou, X.G.; Sandvej, K.; Li, P.J.; Ji, X.L.; Yan, Q.H.; Zhang, X.P.; Da, J.P.; Hamilton-Dutoit, S.J. Epstein--Barr virus gene polymorphisms in Chinese Hodgkin’s disease cases and healthy donors: Identification of three distinct virus variants. J. Gen. Virol. 2001, 82, 1157–1167. [Google Scholar] [CrossRef]
- Garcia-Cosio, M.; Santon, A.; Martin, P.; Reguero, M.E.; Cristobal, E.; Bellas, C. Analysis of Epstein-Barr virus strains and variants in classical Hodgkin’s lymphoma by laser microdissection. Histol. Histopathol. 2008, 23, 209–217. [Google Scholar] [CrossRef]
- Kim, I.; Park, E.R.; Park, S.H.; Lin, Z.; Kim, Y.S. Characteristics of Epstein-Barr virus isolated from the malignant lymphomas in Korea. J. Med. Virol. 2002, 67, 59–66. [Google Scholar] [CrossRef] [PubMed]
- Sandvej, K.; Peh, S.C.; Andresen, B.S.; Pallesen, G. Identification of potential hot spots in the carboxy-terminal part of the Epstein-Barr virus (EBV) BNLF-1 gene in both malignant and benign EBV-associated diseases: High frequency of a 30-bp deletion in Malaysian and Danish peripheral T-cell lymphomas. Blood 1994, 84, 4053–4060. [Google Scholar] [PubMed]
- Boyle, M.J.; Vasak, E.; Tschuchnigg, M.; Turner, J.J.; Sculley, T.; Penny, R.; Cooper, D.A.; Tindall, B.; Sewell, W.A. Subtypes of Epstein-Barr virus (EBV) in Hodgkin’s disease: Association between B-type EBV and immunocompromise. Blood 1993, 81, 468–474. [Google Scholar] [PubMed]
- Guiretti, D.M.; Chabay, P.A.; Valva, P.; Stefanoff, C.G.; Barros, M.H.; De Matteo, E.; Renault, I.Z.; Preciado, M.V.; Hassan, R. Structural variability of the carboxy-terminus of Epstein-Barr virus encoded latent membrane protein 1 gene in Hodgkin’s lymphomas. J. Med. Virol. 2007, 79. [Google Scholar] [CrossRef] [PubMed]
- Hayashi, K.; Chen, W.G.; Chen, Y.Y.; Murakami, I.; Chen, H.L.; Ohara, N.; Nose, S.; Hamaya, K.; Matsui, S.; Bacchi, M.M.; et al. Deletion of Epstein-Barr virus latent membrane protein 1 gene in Japanese and Brazilian gastric carcinomas, metastatic lesions, and reactive lymphocytes. Am. J. Pathol. 1998, 152, 191–198. [Google Scholar] [PubMed]


© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).