Regulation of Constitutive Interferon-Stimulated Genes (Isgs) in Tumor Cells Contributes to Enhanced Antitumor Response of Newcastle Disease Virus-Infected Tumor Vaccines


Abstract
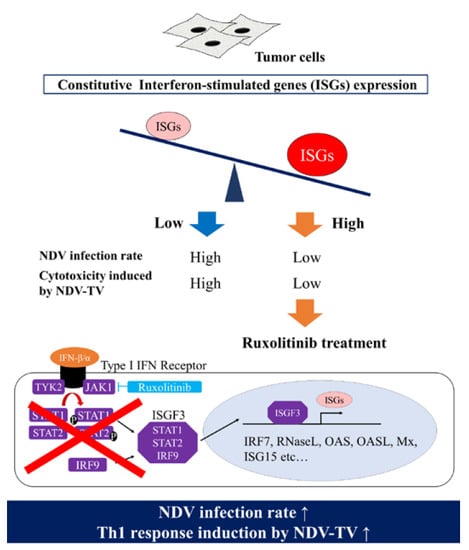
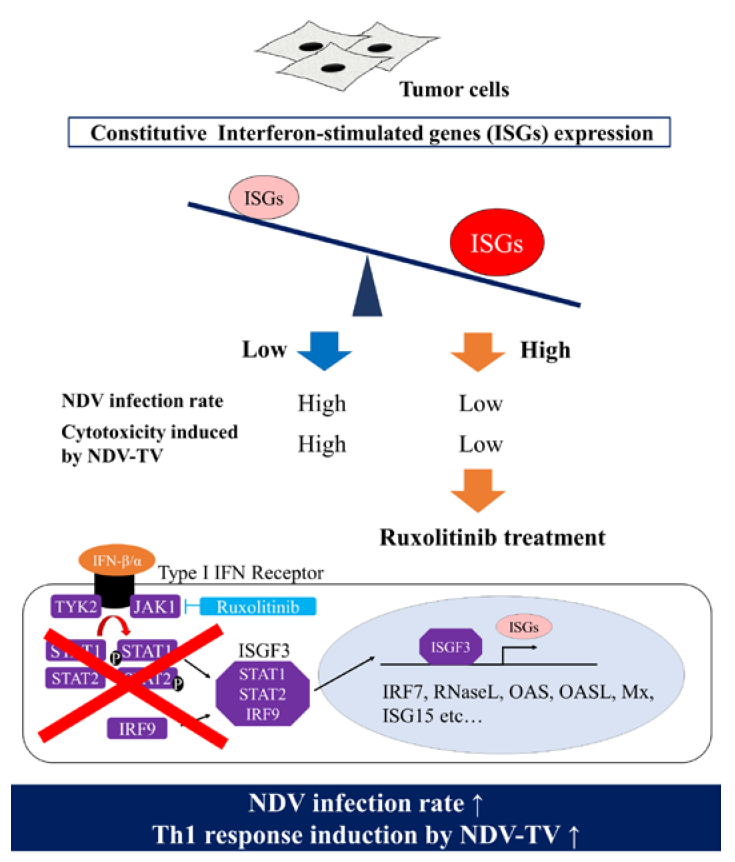
1. Introduction
2. Materials and Methods
2.1. Animals
2.2. Cell Culture and Virus Production
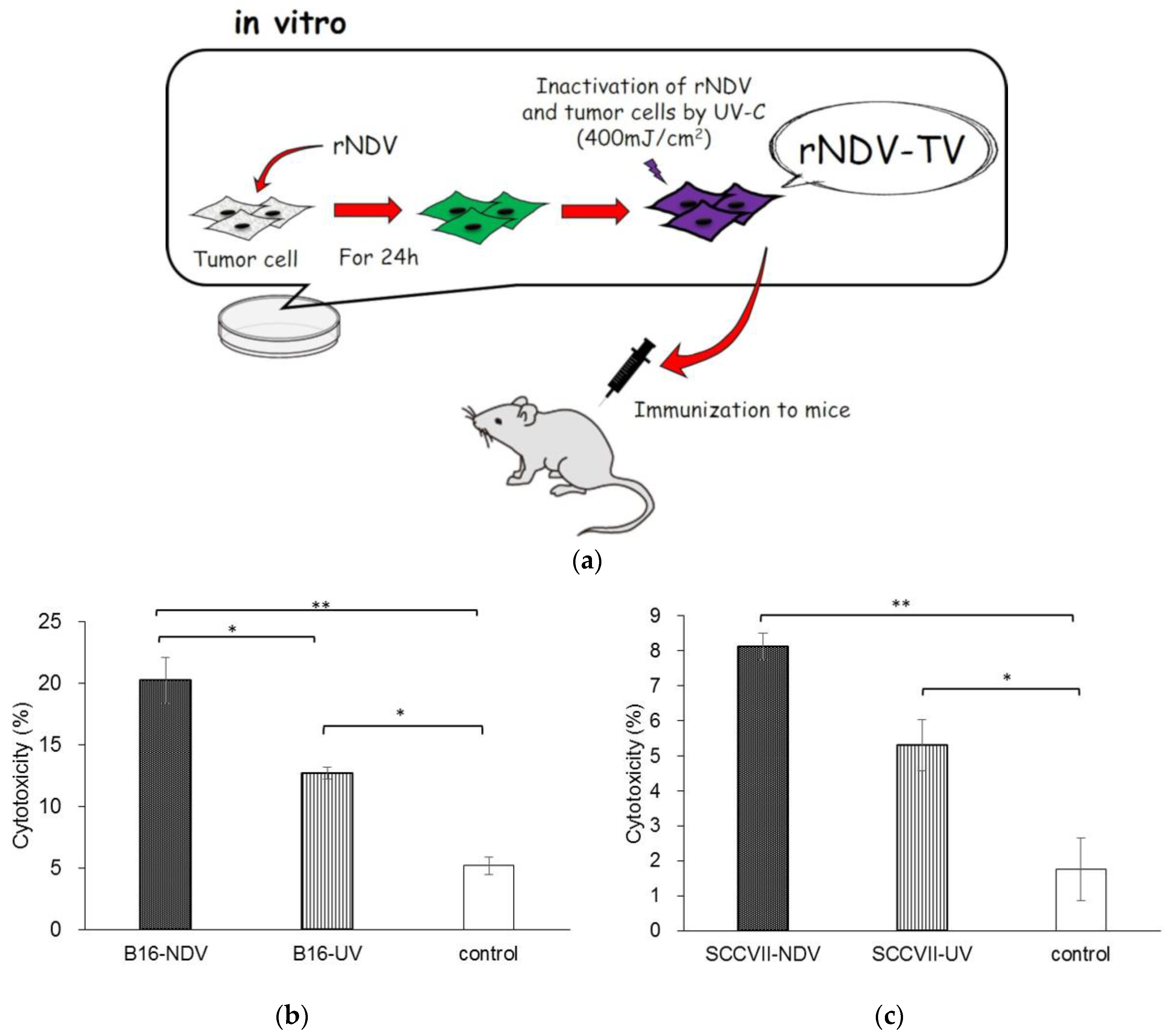
2.3. Preparation of rNDV-TV, Ultraviolet Irradiated Tumor Vaccine (UV-TV) and Ruxolitinib-Treated rNDV-TV (rNDV-TV-Rux)
2.4. Immunization Protocol
2.5. Cytotoxicity Assay
2.6. Viral Infection
2.7. RNA Extraction and cDNA Synthesis
2.8. Quantitative Polymerase Chain Reaction (qPCR)
2.9. JAK Inhibition
2.10. Cytokine Expression
2.11. Flow Cytometry
2.12. Statistical Analyses
3. Result
3.1. Induction of Anti-Tumor Response with rNDV-TV
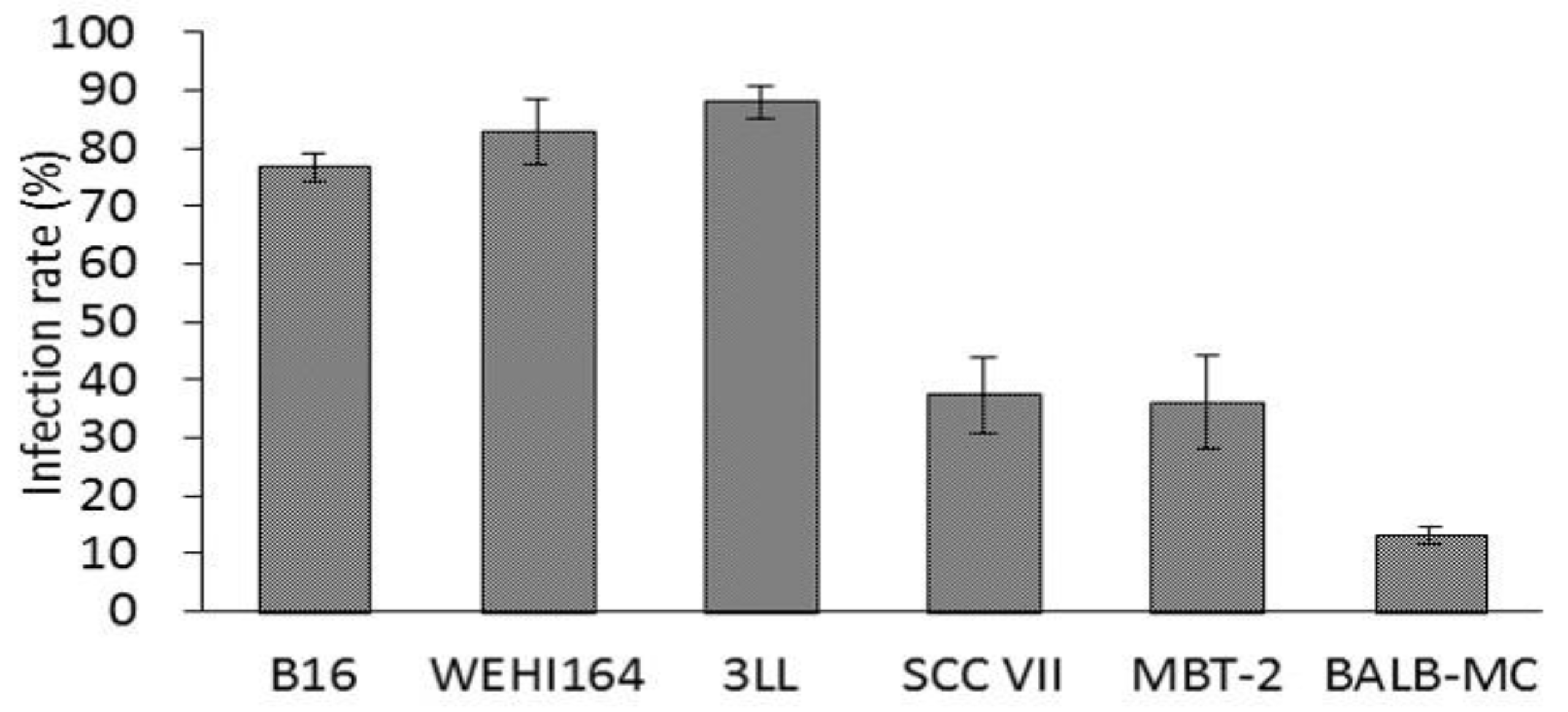
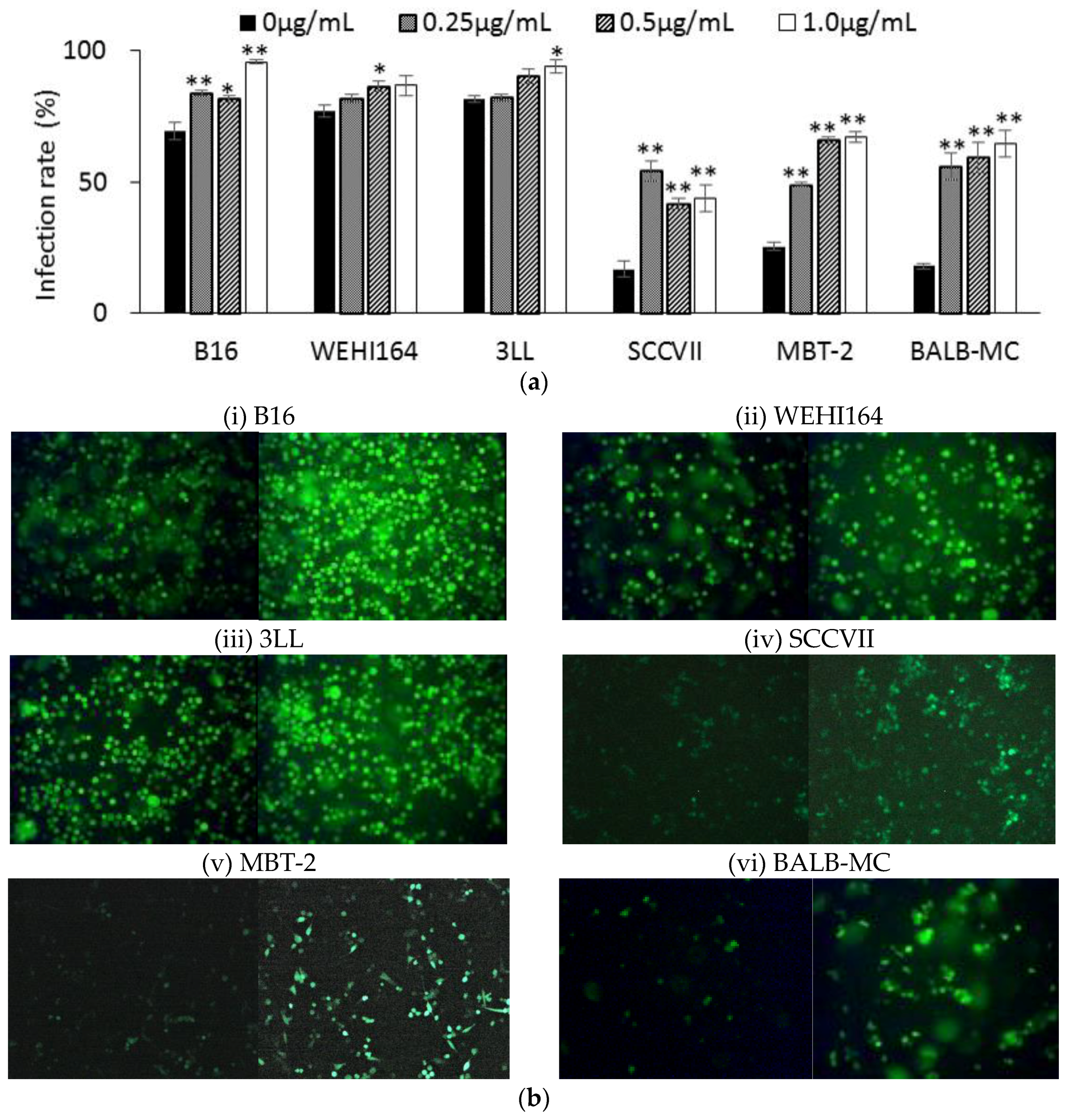
3.2. Infection Rate in Murine Tumor Cell Lines
3.3. The Expression of Type I IFN Related Genes in Tumor Cell Lines
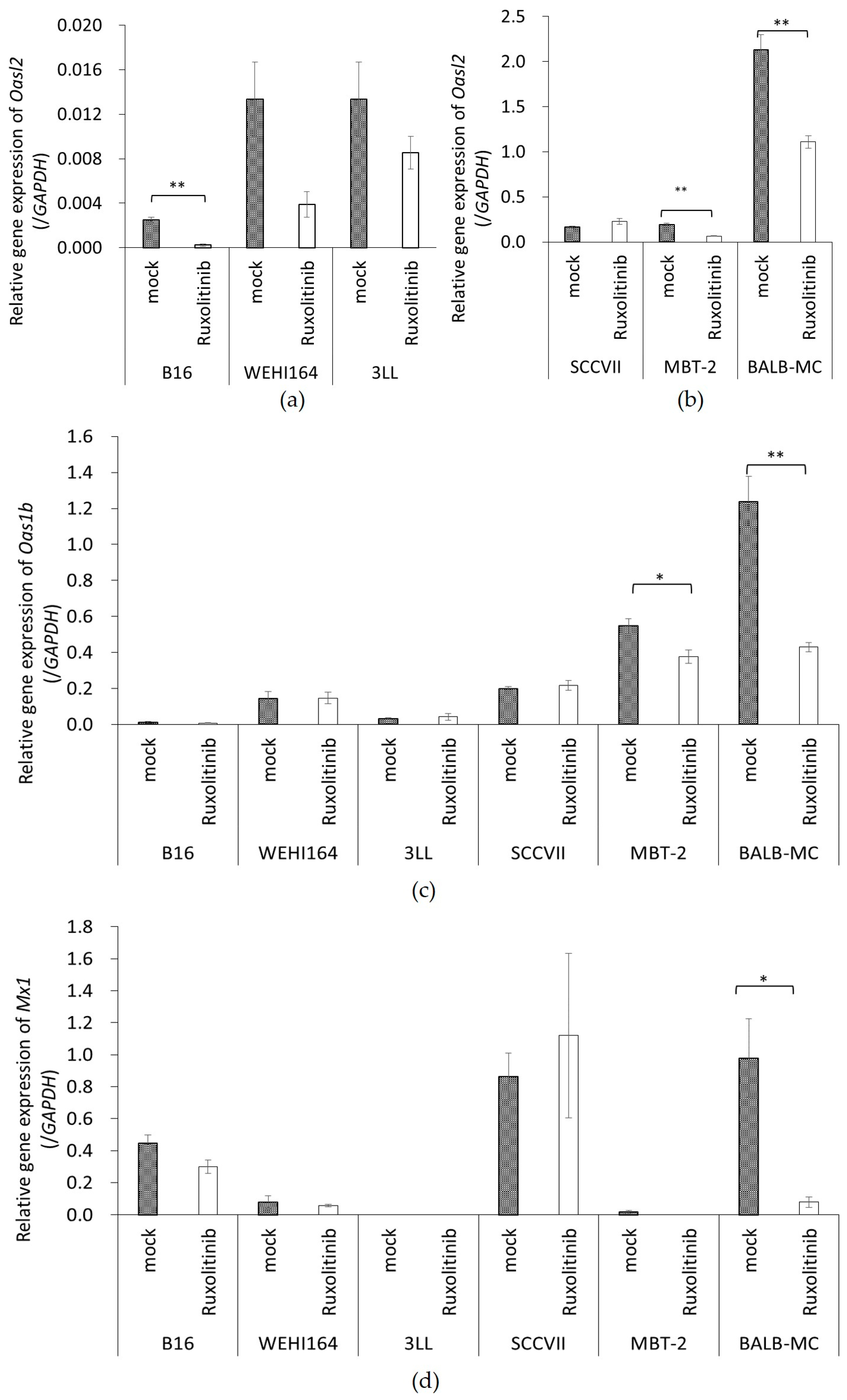
3.4. rNDV Infection Rate in Ruxolitinib (Rux) Treated Tumor Cell Lines
3.5. The Expression of ISGs in Tumor Cell Lines Treated with Rux
3.6. Lymphocyte Subset in Murine SMC Immunized with MBT-NDV-Rux
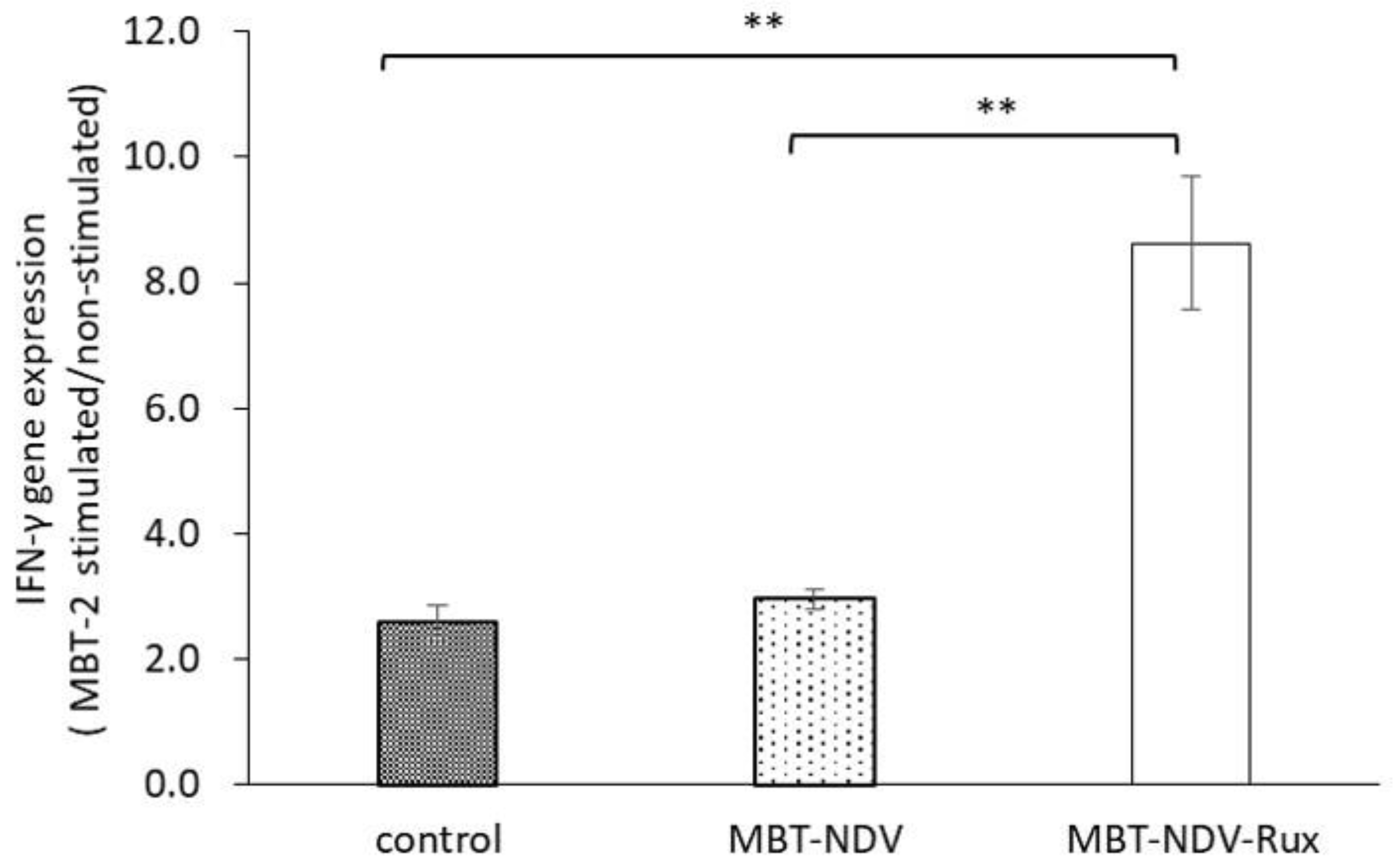
3.7. IFN-γ Gene Expression in Immunized SMC with MBT-2 Stimulation
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Fiola, C.; Peeters, B.; Fournier, P.; Arnold, A.; Bucur, M.; Schirrmacher, V. Tumor selective replication of Newcastle Disease Virus: Association with defects of tumor cells in antiviral defence. Int. J. Cancer 2006, 119, 328–338. [Google Scholar] [CrossRef] [PubMed]
- Vigil, A.; Park, M.S.; Martinez, O.; Chua, M.A.; Xiao, S.; Cros, J.F.; Martínez-Sobrido, L.; Woo, S.L.C.; García-Sastre, A. Use of reverse genetics to enhance the oncolytic properties of Newcastle disease virus. Cancer Res. 2007, 67, 8285–8292. [Google Scholar] [CrossRef] [PubMed]
- Elankumaran, S.; Chavan, V.; Qiao, D.; Shobana, R.; Moorkanat, G.; Biswas, M.; Samal, S.K. Type I Interferon-Sensitive Recombinant Newcastle Disease Virus for Oncolytic Virotherapy. J. Virol. 2010, 84, 3835–3844. [Google Scholar] [CrossRef] [PubMed]
- Fujita, M.; Scheurer, M.E.; Decker, S.A.; McDonald, H.A.; Kohanbash, G.; Kastenhuber, E.R.; Kato, H.; Bondy, M.L.; Ohlfest, J.R.; Okada, H. Role of type 1 IFNs in antiglioma immunosurveillance—Using mouse studies to guide examination of novel prognostic markers in humans. Clin. Cancer Res. 2010, 16, 3409–3419. [Google Scholar] [CrossRef] [PubMed]
- LeBon, A.; Tough, D.F. Links between innate and adaptive immunity via type I interferon. Curr. Opin. Immunol. 2002, 14, 432–436. [Google Scholar] [CrossRef]
- Jarahian, M.; Watzl, C.; Fournier, P.; Arnold, A.; Djandji, D.; Zahedi, S.; Cerwenka, A.; Paschen, A.; Schirrmacher, V.; Momburg, F. Activation of Natural Killer Cells by Newcastle Disease Virus Hemagglutinin-Neuraminidase. J. Virol. 2009, 83, 8108–8121. [Google Scholar] [CrossRef] [PubMed]
- Washburn, B.; Schirrmacher, V. Human tumor cell infection by Newcastle Disease Virus leads to upregulation of HLA and cell adhesion molecules and to induction of interferons, chemokines and finally apoptosis. Int. J. Oncol. 2002, 21, 85–93. [Google Scholar] [CrossRef] [PubMed]
- Ahlert, T.; Sauerbrei, W.; Bastert, G.; Ruhland, S.; Bartik, B.; Simiantonaki, N.; Schumacher, J.; Hacker, B.; Schumacher, M.; Schirrmacher, V. Tumor-cell number and viability as quality and efficacy parameters of autologous virus-modified cancer vaccines in patients with breast or ovarian cancer. J. Clin. Oncol. 1997, 15, 1354–1366. [Google Scholar] [CrossRef] [PubMed]
- Ockert, D.; Schirrmacher, V.; Beck, N.; Stoelben, E.; Ahlert, T.; Flechtenmacher, J.; Hagmuller, E.; Buchcik, R.; Nagel, M.; Saeger, H.D. Newcastle disease virus infected intact autologous tumor cell vaccine for adjuvant active specific immunotherapy of resected colorectal carcinoma. Clin. Cancer Res. 1996, 2, 21–28. [Google Scholar] [PubMed]
- Karcher, J.; Dyckhoff, G.; Beckhove, P.; Reisser, C.; Brysch, M.; Ziouta, Y.; Helmke, B.H.; Weidauer, H.; Schirrmacher, V.; Herold-Mende, C. Antitumor vaccination in patients with head and neck squamous cell carcinomas with autologous virus-modified tumor cells. Cancer Res. 2004, 64, 8057–8061. [Google Scholar] [CrossRef] [PubMed]
- Steiner, H.H. Antitumor Vaccination of Patients with Glioblastoma Multiforme: A Pilot Study to Assess Feasibility, Safety, and Clinical Benefit. J. Clin. Oncol. 2004, 22, 4272–4281. [Google Scholar] [CrossRef] [PubMed]
- Hagiwara, K.; Kadosawa, T.; Nakaya, T. Tumorcidal Effect of Recombinant Newcastle Disease Virus. Open Vet. Sci. J. 2008, 2, 11–15. [Google Scholar] [CrossRef]
- Kishida, S.; Nakaya, T.; Hagiwara, K. Vaccination with Melanoma Cells Infected with Recombinant Newcastle Disease Virus Suppresses Tumor Metastasis. SOJ Immunol. 2017, 5, 1–6. [Google Scholar]
- Takamura-Ishii, M.; Miura, T.; Nakaya, T.; Hagiwara, K. Induction of antitumor response to fibrosarcoma by Newcastle disease virus-infected tumor vaccine. Med. Oncol. 2017, 34. [Google Scholar] [CrossRef] [PubMed]
- Krishnamurthy, S.; Takimoto, T.; Scroggs, R.A.; Portner, A. Differentially regulated interferon response determines the outcome of Newcastle disease virus infection in normal and tumor cell lines. J. Virol. 2006, 80, 5145–5155. [Google Scholar] [CrossRef] [PubMed]
- Wilden, H.; Fournier, P.; Zawatzky, R.; Schirrmacher, V. Expression of RIG-I, IRF3, IFN-ß and IRF7 determines resistance or susceptibility of cells to infection by Newcastle Disease Virus. Int. J. Oncol. 2009, 34, 971–982. [Google Scholar] [PubMed]
- Fournier, P.; Wilden, H.; Schirrmacher, V. Importance of retinoic acid-inducible gene I and of receptor for type I interferon for cellular resistance to infection by Newcastle disease virus. Int. J. Oncol. 2012, 40, 287–298. [Google Scholar] [CrossRef] [PubMed]
- Youngner, J.S.; Scott, A.W.; Hallum, J.V.; Stinebring, W.R. Interferon production by inactivated Newcastle disease virus in cell cultures and in mice. J. Bacteriol. 1966, 92, 862–868. [Google Scholar] [PubMed]
- Ertel, C.; Millar, N.S.; Emmerson, P.T.; Schirrmacher, V.; Von Hoegen, P. Viral hemagglutinin augments peptide-specific cytotoxic T cell responses. Eur. J. Immunol. 1993, 23, 2592–2596. [Google Scholar] [CrossRef] [PubMed]
- Zeng, J.; Fournier, P.; Schirrmacher, V. Induction of Interferon-α and Tumor Necrosis Factor-Related Apoptosis-Inducing Ligand in Human Blood Mononuclear Cells by Hemagglutinin-Neuraminidase but Not F Protein of Newcastle Disease Virus. Virology 2002, 297, 19–30. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | Pre-Infection | Post-Infection (8 h) |
|---|---|---|
| RIG-I | −0.480 * | −0.44 |
| TLR3 | −0.840 ** | −0.486 * |
| TLR7 | −0.342 | −0.057 |
| IFN-β | −0.139 | −0.352 |
| IFN-α | 0.161 | −0.028 |
| IRF-3 | −0.579 * | −0.347 |
| IRF-7 | −0.359 | −0.594 * |
| Mx1 | −0.518 * | −0.371 |
| Mx2 | −0.678 ** | −0.304 |
| OAS1a | −0.574 * | −0.373 |
| OAS1b | −0.685 ** | −0.411 |
| OAS2 | −0.612 ** | −0.480 |
| OAS3 | −0.530 * | −0.373 |
| OASL1 | −0.417 | −0.724 ** |
| OASL2 | −0.672 ** | −0.637 ** |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Takamura-Ishii, M.; Nakaya, T.; Hagiwara, K. Regulation of Constitutive Interferon-Stimulated Genes (Isgs) in Tumor Cells Contributes to Enhanced Antitumor Response of Newcastle Disease Virus-Infected Tumor Vaccines. Cancers 2018, 10, 186. https://doi.org/10.3390/cancers10060186
Takamura-Ishii M, Nakaya T, Hagiwara K. Regulation of Constitutive Interferon-Stimulated Genes (Isgs) in Tumor Cells Contributes to Enhanced Antitumor Response of Newcastle Disease Virus-Infected Tumor Vaccines. Cancers. 2018; 10(6):186. https://doi.org/10.3390/cancers10060186
Chicago/Turabian StyleTakamura-Ishii, Mai, Takaaki Nakaya, and Katsuro Hagiwara. 2018. "Regulation of Constitutive Interferon-Stimulated Genes (Isgs) in Tumor Cells Contributes to Enhanced Antitumor Response of Newcastle Disease Virus-Infected Tumor Vaccines" Cancers 10, no. 6: 186. https://doi.org/10.3390/cancers10060186
APA StyleTakamura-Ishii, M., Nakaya, T., & Hagiwara, K. (2018). Regulation of Constitutive Interferon-Stimulated Genes (Isgs) in Tumor Cells Contributes to Enhanced Antitumor Response of Newcastle Disease Virus-Infected Tumor Vaccines. Cancers, 10(6), 186. https://doi.org/10.3390/cancers10060186