Epstein Barr Virus-Associated Hodgkin Lymphoma



Abstract
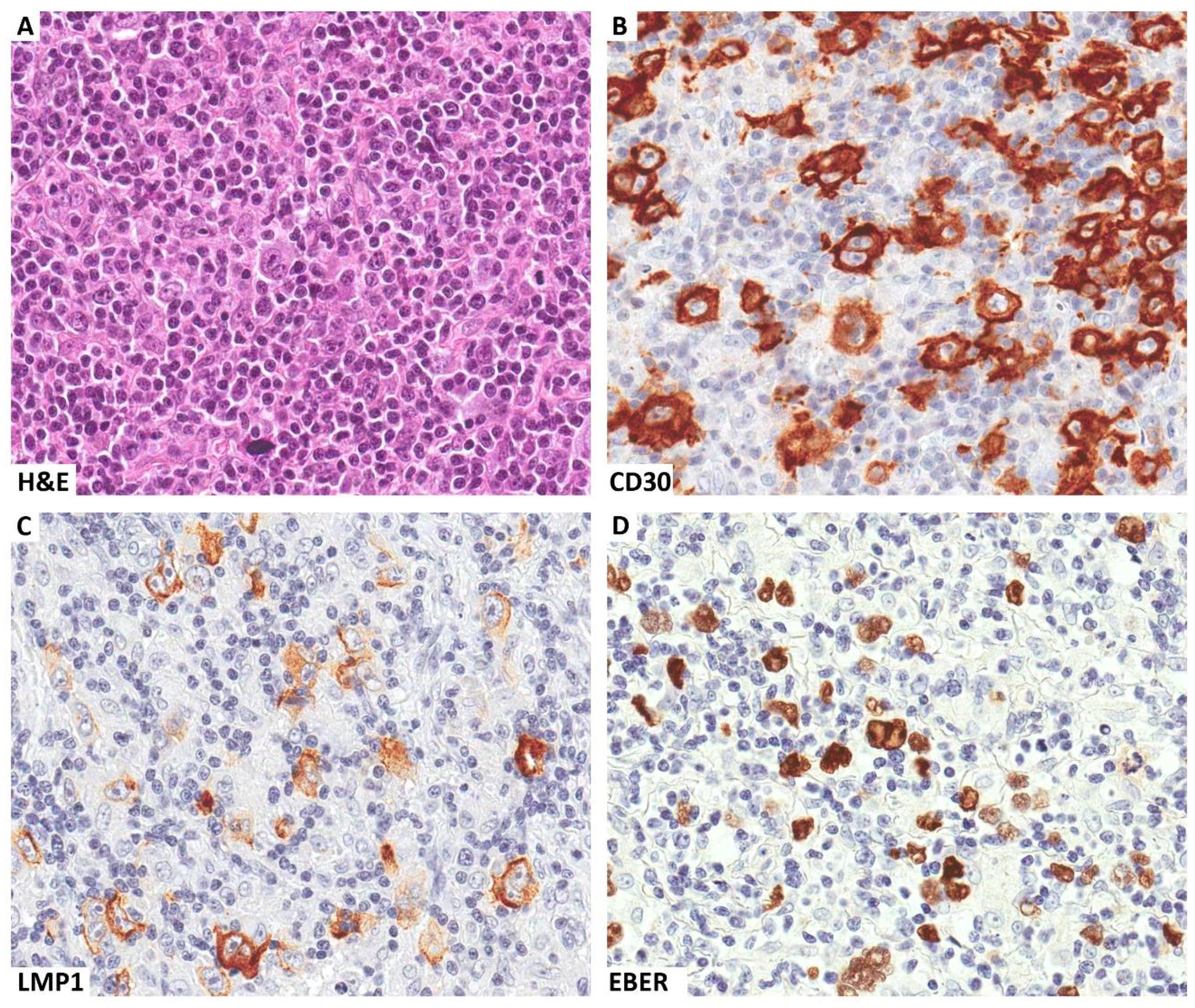
:1. Introduction
2. Epstein–Barr Virus and cHL
3. Tumour Microenvironment in EBV-Related cHL
4. HIV-Related Hodgkin Lymphoma
5. Implications of Immune Evasion for Immunotherapy in EBV-Associated cHL
6. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Jackson, H.; Parker, F. Hodgkin’s disease. I. General considerations. N. Engl. J. Med. 1944, 230, 1–8. [Google Scholar] [CrossRef]
- Lukes, R.J.; Butler, J.J. The pathology and nomenclature of Hodgkin’s disease. Cancer Res. 1966, 26, 1063–1083. [Google Scholar] [PubMed]
- Stein, H.; Pileri, S.A.; Weiss, L.M. Classical Hodgkin lymphoma, introduction. In WHO Classification of Tumours of Haematopoietic and Lymphoid Tissues, Revised 4th ed.; Swerdlow, S.H., Campo, E., Harris, N.L., Jaffe, E.S., Pileri, S.A., Stein, H., Thiele, J., Eds.; IARC: Lyon, France, 2017; pp. 424–430. [Google Scholar]
- Lukes, R.J.; Craver, L.F.; Hall, T.C.; Rappaport, H.; Ruben, P. Report of the nomenclature committee. Cancer Res. 1966, 26, 1311. [Google Scholar]
- Younes, A.; Carbone, A.; Johnson, P.; Dabaja, B.; Ansell, S.; Kuruvilla, J. Hodgkin’s lymphoma. In De Vita, Hellman, and Rosenberg’s Cancer: Principles & Practice of Oncology; De Vita, V.T.J., Lawrrence, T.S., Rosemberg, S.A., Eds.; Wolters Kluwer Health; Lippincott Williams & Wilkins: Philadelphia, PA, USA, 2014. [Google Scholar]
- Carbone, A.; Gloghini, A. Hodgkin lymphoma classification: Are we at a crossroads? Cancer 2017, 123, 3654–3655. [Google Scholar] [CrossRef] [PubMed]
- Boudová, L.; Torlakovic, E.; Delabie, J.; Reimer, P.; Pfistner, B.; Wiedenmann, S.; Diehl, V.; Müller-Hermelink, H.K.; Rüdiger, T. Nodular lymphocyte-predominant Hodgkin lymphoma with nodules resembling T-cell/histiocyte-rich B-cell lymphoma: Differential diagnosis between nodular lymphocyte-predominant Hodgkin lymphoma and T-cell/histiocyte-rich B-cell lymphoma. Blood 2003, 102, 3753–3758. [Google Scholar] [PubMed]
- Brune, V.; Tiacci, E.; Pfeil, I.; Döring, C.; Eckerle, S.; van Noesel, C.J.; Klapper, W.; Falini, B.; von Heydebreck, A.; Metzler, D.; et al. Origin and pathogenesis of nodular lymphocyte-predominant Hodgkin lymphoma as revealed by global gene expression analysis. J. Exp. Med. 2008, 205, 2251–2268. [Google Scholar] [CrossRef] [PubMed]
- De Wolf-Peeters, C.; Delabie, J.; Campo, E.; Jaffe, E.S.; Delsol, G. T cell/histiocyte-rich large B-cell lymphoma. In World Health Organization Classification of Tumours, Pathology and Genetics of Tumours of Haematopoietic and Lymphoid Tissues; Swerdlow, S.H., Campo, E., Harris, N.L., Jaffe, E.S., Pileri, S.A., Stein, H., Thiele, J., Vardiman, J.W., Eds.; IARC Press: Lyon, France, 2008; pp. 238–239. [Google Scholar]
- Swerdlow, S.H.; Campo, E.; Pileri, S.A.; Harris, N.L.; Stein, H.; Siebert, R.; Advani, R.; Ghielmini, M.; Salles, G.A.; Zelenetz, A.D.; et al. The 2016 revision of the World Health Organization classification of lymphoid neoplasms. Blood 2016, 127, 2375–2390. [Google Scholar] [CrossRef] [PubMed]
- IARC Monograph on the Evaluation of Carcinogenic Risk to Humans. A Review of Human Carcinogens. Part B: Biological Agents; IARC: Lyon, France, 2012; Volume 100. [Google Scholar]
- Dolcetti, R.; Gloghini, A.; Caruso, A.; Carbone, A. A lymphomagenic role for HIV beyond immune suppression? Blood 2016, 127, 1403–1409. [Google Scholar] [CrossRef] [PubMed]
- Shannon-Lowe, C.; Rickinson, A.B.; Bell, A.I. Epstein-Barr virus-associated lymphomas. Philos. Trans. R. Soc. Lond. B Biol. Sci. 2017, 372, 1732. [Google Scholar] [CrossRef] [PubMed]
- Grywalska, E.; Rolinski, J. Epstein-Barr virus-associated lymphomas. Semin. Oncol. 2015, 42, 291–303. [Google Scholar] [CrossRef] [PubMed]
- Chetaille, B.; Bertucci, F.; Finetti, P.; Esterni, B.; Stamatoullas, A.; Picquenot, J.M.; Copin, M.C.; Morschhauser, F.; Casasnovas, O.; Petrella, T.; et al. Molecular profiling of classical Hodgkin lymphoma tissues uncovers variations in the tumor microenvironment and correlations with EBV infection and outcome. Blood 2009, 113, 2765–2775. [Google Scholar] [CrossRef] [PubMed]
- Aldinucci, D.; Gloghini, A.; Pinto, A.; De Filippi, R.; Carbone, A. The classical Hodgkin’s lymphoma microenvironment and its role in promoting tumour growth and immune escape. J. Pathol. 2010, 221, 248–263. [Google Scholar] [CrossRef] [PubMed]
- Steidl, C.; Connors, J.M.; Gascoyne, R.D. Molecular pathogenesis of Hodgkin’s lymphoma: Increasing evidence of the importance of the microenvironment. J. Clin. Oncol. 2011, 29, 1812–1826. [Google Scholar] [CrossRef] [PubMed]
- Carbone, A.; Gloghini, A.; Carlo-Stella, C. Are EBV-related and EBV-unrelated Hodgkin lymphomas different with regard to susceptibility to checkpoint blockade? Blood 2018. [Google Scholar] [CrossRef] [PubMed]
- Mills, C.D. Anatomy of a discovery: M1 and m2 macrophages. Front. Immunol. 2015, 6, 212. [Google Scholar] [CrossRef] [PubMed]
- Barros, M.H.; Segges, P.; Vera-Lozada, G.; Hassan, R.; Niedobitek, G. Macrophage polarization reflects T cell composition of tumor microenvironment in pediatric classical Hodgkin lymphoma and has impact on survival. PLoS ONE 2015, 10, e0124531. [Google Scholar] [CrossRef] [PubMed]
- Eliopoulos, A.G.; Stack, M.; Dawson, C.W.; Kaye, K.M.; Hodgkin, L.; Sihota, S.; Rowe, M.; Young, L.S. Epstein-Barr virus-encoded LMP1 and CD40 mediate IL-6 production in epithelial cells via an NF-kappaB pathway involving TNF receptor-associated factors. Oncogene 1997, 14, 2899–2916. [Google Scholar] [CrossRef] [PubMed]
- Nakagomi, H.; Dolcetti, R.; Bejarano, M.T.; Pisa, P.; Kiessling, R.; Masucci, M.G. The Epstein-Barr virus latent membrane protein-1 (LMP1) induces interleukin-10 production in Burkitt lymphoma lines. Int. J. Cancer 1994, 57, 240–244. [Google Scholar] [CrossRef] [PubMed]
- Eliopoulos, A.G.; Gallagher, N.J.; Blake, S.M.; Dawson, C.W.; Young, L.S. Activation of the p38 mitogen-activated protein kinase pathway by Epstein-Barr virus-encoded latent membrane protein 1 coregulates interleukin-6 and interleukin-8 production. J. Biol. Chem. 1999, 274, 16085–16096. [Google Scholar] [CrossRef] [PubMed]
- Carbone, A.; Vaccher, E.; Gloghini, A.; Pantanowitz, L.; Abayomi, A.; de Paoli, P.; Franceschi, S. Diagnosis and management of lymphomas and other cancers in HIV-infected patients. Nat. Rev. Clin. Oncol. 2014, 11, 223–238. [Google Scholar] [CrossRef] [PubMed]
- Carbone, A.; Volpi, C.C.; Gualeni, A.V.; Gloghini, A. Epstein-Barr virus associated lymphomas in people with HIV. Curr. Opin. HIV AIDS 2017, 12, 39–46. [Google Scholar] [CrossRef] [PubMed]
- Hartmann, S.; Jakobus, C.; Rengstl, B.; Döring, C.; Newrzela, S.; Brodt, H.R.; Wolf, T.; Hansmann, M.L. Spindle-shaped CD163+ rosetting macrophages replace CD4+ T-cells in HIV-related classical Hodgkin lymphoma. Mod. Pathol. 2013, 26, 648–657. [Google Scholar] [CrossRef] [PubMed]
- Cassol, E.; Cassetta, L.; Rizzi, C.; Alfano, M.; Poli, G. M1 and M2a polarization of human monocyte-derived macrophages inhibits HIV-1 replication by distinct mechanisms. J. Immunol. 2009, 182, 6237–6346. [Google Scholar] [CrossRef] [PubMed]
- Biggar, R.J.; Jaffe, E.S.; Goedert, J.J.; Chaturvedi, A.; Pfeiffer, R.; Engels, E.A. Hodgkin lymphoma and immunodeficiency in persons with HIV/AIDS. Blood 2006, 108, 3786–3791. [Google Scholar] [CrossRef] [PubMed]
- Clifford, G.M.; Rickenbach, M.; Lise, M.; Dal Maso, L.; Battegay, M.; Bohlius, J.; Boffi, E.A.E.; Karrer, U.; Jundt, G.; Bordoni, A.; et al. Swiss HIV Cohort Study. Hodgkin lymphoma in the Swiss HIV Cohort Study. Blood 2009, 113, 5737–5742. [Google Scholar] [CrossRef] [PubMed]
- Tsirigotis, P.; Savani, B.N.; Nagler, A. Programmed death-1 immune checkpoint blockade in the treatment of hematological malignancies. Ann. Med. 2016, 48, 428–439. [Google Scholar] [CrossRef] [PubMed]
- Ansell, S.M.; Lesokhin, A.M.; Borrello, I.; Halwani, A.; Scott, E.C.; Gutierrez, M.; Schuster, S.J.; Millenson, M.M.; Cattry, D.; Freeman, G.J.; et al. PD-1 blockade with nivolumab in relapsed or refractory Hodgkin’s lymphoma. N. Engl. J. Med. 2015, 372, 311–319. [Google Scholar] [CrossRef] [PubMed]
- Younes, A.; Ansell, S.M. Novel agents in the treatment of Hodgkin lymphoma: Biological basis and clinical results. Semin. Hematol. 2016, 53, 186–189. [Google Scholar] [CrossRef] [PubMed]
- Bartlett, N.L. Emerging role of novel therapies in Hodgkin lymphoma: Proceed with caution. Hematol. Am. Soc. Hematol. Educ. Program. 2017, 2017, 317–323. [Google Scholar]
- Shanbhag, S.; Ambinder, R.F. Hodgkin lymphoma: A review and update on recent progress. CA Cancer J. Clin. 2018, 68, 116–132. [Google Scholar] [CrossRef] [PubMed]
- Carbone, A.; Gloghini, A.; Castagna, L.; Santoro, A.; Carlo-Stella, C. Primary refractory and early-relapsed Hodgkin’s lymphoma: Strategies for therapeutic targeting based on the tumour microenvironment. J. Pathol. 2015, 237, 4–13. [Google Scholar] [CrossRef] [PubMed]
- Green, M.R.; Monti, S.M.; Rodig, S.J.; Juszczynski, P.; Currie, T.; O’Donnell, E.; Chapuy, B.; Takeyama, K.; Neuberg, D.; Golub, T.R.; et al. Integrative analysis reveals selective 9p24.1 amplification, increased PD-1 ligand expression, and further induction via JAK2 in nodular sclerosing Hodgkin lymphoma and primary mediastinal large B-cell lymphoma. Blood 2010, 116, 3268–3277. [Google Scholar] [CrossRef] [PubMed]
- Green, M.R.; Rodig, S.; Juszczynski, P.; Ouyang, J.; Sinha, P.; O’Donnell, E.; Neuberg, D.; Shipp, M.A. Constitutive AP-1 activity and EBV infection induce PD-L1 in Hodgkin lymphomas and posttransplant lymphoproliferative disorders: Implications for targeted therapy. Clin. Cancer Res. 2012, 18, 1611–1618. [Google Scholar] [CrossRef] [PubMed]
- Roemer, M.G.; Advani, R.H.; Ligon, A.H.; Natkunam, Y.; Redd, R.A.; Homer, H.; Connelly, C.F.; Sun, H.H.; Daadi, S.E.; Freeman, G.J.; et al. PD-L1 and PD-L2 Genetic Alterations Define Classical Hodgkin Lymphoma and Predict Outcome. J. Clin. Oncol. 2016, 34, 2690–2697. [Google Scholar] [CrossRef] [PubMed]
- Koh, Y.W.; Jeon, Y.K.; Yoon, D.H.; Suh, C.; Huh, J. Programmed death 1 expression in the peritumoral microenvironment is associated with a poorer prognosis in classical Hodgkin lymphoma. Tumour Biol. 2016, 37, 7507–7514. [Google Scholar] [CrossRef] [PubMed]
- Greenough, T.C.; Campellone, S.C.; Brody, R.; Jain, S.; Sanchez-Merino, V.; Somasundaran, M.; Luzuriaga, K. Programmed Death-1 expression on Epstein Barr virus specific CD8+ T cells varies by stage of infection, epitope specificity, and T-cell receptor usage. PLoS ONE 2010, 5, e12926. [Google Scholar] [CrossRef] [PubMed]
- Duffield, A.S.; Ascierto, M.L.; Anders, R.A.; Taube, J.M.; Meeker, A.K.; Chen, S.; McMiller, T.L.; Phillips, N.A.; Xu, H.; Ogurtsova, A.; et al. Th17 immune microenvironment in Epstein-Barr virus-negative Hodgkin lymphoma: Implications for immunotherapy. Blood Adv. 2017, 1, 1324–1334. [Google Scholar] [CrossRef] [PubMed]
- Di Napoli, A.; Al-Jadiri, M.F.; Talerico, C.; Duranti, E.; Pilozzi, E.; Trivedi, P.; Anastasiadou, E.; Alsaadawi, A.R.; Al-Darraji, A.F.; Al-Hadad, S.A.; et al. Epstein-Barr virus (EBV) positive classical Hodgkin lymphoma of Iraqi children: An immunophenotypic and molecular characterization of Hodgkin/Reed-Sternberg cells. Pediatr. Blood Cancer 2013, 60, 2068–2072. [Google Scholar] [CrossRef] [PubMed]


{kind=link}
| Host | Hodgkin Lymphoma | EBV Infection |
|---|---|---|
| Without known immunosuppression | NLPHL | Usually absent |
| cHL, nodular sclerosis | Variably present | |
| cHL, mixed cellularity | Usually present | |
| Rare cHL subtypes | Variably present | |
| With acquired Immunodeficiency | HIV-associated cHL | Present |
| Post-transplant, cHL type PTLD | Present | |
| Iatrogenic (methotrexate) | Variably present |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Carbone, A.; Gloghini, A. Epstein Barr Virus-Associated Hodgkin Lymphoma. Cancers 2018, 10, 163. https://doi.org/10.3390/cancers10060163
Carbone A, Gloghini A. Epstein Barr Virus-Associated Hodgkin Lymphoma. Cancers. 2018; 10(6):163. https://doi.org/10.3390/cancers10060163
Chicago/Turabian StyleCarbone, Antonino, and Annunziata Gloghini. 2018. "Epstein Barr Virus-Associated Hodgkin Lymphoma" Cancers 10, no. 6: 163. https://doi.org/10.3390/cancers10060163
APA StyleCarbone, A., & Gloghini, A. (2018). Epstein Barr Virus-Associated Hodgkin Lymphoma. Cancers, 10(6), 163. https://doi.org/10.3390/cancers10060163