Oncolytic Viruses for Canine Cancer Treatment


Abstract
1. Introduction
2. Brief History of Oncolytic Viruses
3. Oncolytic Viruses for Canine Cancer Treatment
3.1. Paramyxoviridae Family
3.1.1. Measles Virus (MV)
3.1.2. Canine Distemper Virus (CDV)
3.1.3. Newcastle Disease Virus (NDV)
3.1.4. Sendai Virus (SV)
3.2. Rabdoviridae Family
Vesicular Stomatitis Virus
3.3. Togaviridae Family
Semliki Forest Virus (SFV)
3.4. Reoviridae Family
Reovirus Serotype 3
3.5. Parvoviridae Family
Canine Parvovirus Serotype 2 (CPV-2)
3.6. Poxviridae Family
3.6.1. Myxoma Virus (MYXV)
3.6.2. Canarypox Virus
3.6.3. Vaccinia Virus (VACV)
3.7. Adenoviridae Family
3.7.1. Canine Adenovirus Type 2 (CAV-2)
3.7.2. Human Adenovirus Type 5 (Ad5)
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Couzin-Frankel, J. Breakthrough of the year 2013. Cancer immunotherapy. Science (New York, N.Y.) 2013, 342, 1432–1433. [Google Scholar] [CrossRef] [PubMed]
- Topalian, S.L.; Hodi, F.S.; Brahmer, J.R.; Gettinger, S.N.; Smith, D.C.; McDermott, D.F.; Powderly, J.D.; Carvajal, R.D.; Sosman, J.A.; Atkins, M.B.; et al. Safety, activity, and immune correlates of anti-pd-1 antibody in cancer. N. Engl. J. Med. 2012, 366, 2443–2454. [Google Scholar] [CrossRef] [PubMed]
- Hodi, F.S.; O‘Day, S.J.; McDermott, D.F.; Weber, R.W.; Sosman, J.A.; Haanen, J.B.; Gonzalez, R.; Robert, C.; Schadendorf, D.; Hassel, J.C.; et al. Improved survival with ipilimumab in patients with metastatic melanoma. N. Engl. J. Med. 2010, 363, 711–723. [Google Scholar] [CrossRef] [PubMed]
- Wolchok, J.D.; Kluger, H.; Callahan, M.K.; Postow, M.A.; Rizvi, N.A.; Lesokhin, A.M.; Segal, N.H.; Ariyan, C.E.; Gordon, R.A.; Reed, K.; et al. Nivolumab plus ipilimumab in advanced melanoma. N. Engl. J. Med. 2013, 369, 122–133. [Google Scholar] [CrossRef] [PubMed]
- White, C.L.; Twigger, K.R.; Vidal, L.; De Bono, J.S.; Coffey, M.; Heinemann, L.; Morgan, R.; Merrick, A.; Errington, F.; Vile, R.G.; et al. Characterization of the adaptive and innate immune response to intravenous oncolytic reovirus (dearing type 3) during a phase i clinical trial. Gene Ther. 2008, 15, 911–920. [Google Scholar] [CrossRef] [PubMed]
- Thomas, E.D.; Meza-Perez, S.; Bevis, K.S.; Randall, T.D.; Gillespie, G.Y.; Langford, C.; Alvarez, R.D. Il-12 expressing oncolytic herpes simplex virus promotes anti-tumor activity and immunologic control of metastatic ovarian cancer in mice. J. Ovarian Res. 2016, 9, 70. [Google Scholar] [CrossRef] [PubMed]
- Khanna, C.; Lindblad-Toh, K.; Vail, D.; London, C.; Bergman, P.; Barber, L.; Breen, M.; Kitchell, B.; McNeil, E.; Modiano, J.F.; et al. The dog as a cancer model. Nat. Biotechnol. 2006, 24, 1065–1066. [Google Scholar] [CrossRef] [PubMed]
- Baioni, E.; Scanziani, E.; Vincenti, M.C.; Leschiera, M.; Bozzetta, E.; Pezzolato, M.; Desiato, R.; Bertolini, S.; Maurella, C.; Ru, G. Estimating canine cancer incidence: Findings from a population-based tumour registry in northwestern italy. BMC Vet. Res. 2017, 13, 203. [Google Scholar] [CrossRef] [PubMed]
- Kidd, C. The many challenges of veterinary oncology. Can. Vet. J. 2008, 49, 1132–1135. [Google Scholar] [PubMed]
- Bronson, R.T. Variation in age at death of dogs of different sexes and breeds. Am. J. Vet. Res. 1982, 43, 2057–2059. [Google Scholar] [PubMed]
- Fleming, J.M.; Creevy, K.E.; Promislow, D.E. Mortality in north american dogs from 1984 to 2004: An investigation into age-, size-, and breed-related causes of death. J. Vet. Intern. Med. 2011, 25, 187–198. [Google Scholar] [CrossRef] [PubMed]
- Spangler, W.L.; Culbertson, M.R. Prevalence, type, and importance of splenic diseases in dogs: 1,480 cases (1985–1989). J. Am. Vet. Med. Assoc. 1992, 200, 829–834. [Google Scholar] [PubMed]
- Grabarevic, Z.; Spoljar, J.B.; Kurilj, A.G.; Sostaric-Zuckermann, I.C.; Artukovic, B.; Hohsteter, M.; Beck, A.; Dzaja, P.; Strmecki, N.M. Mast cell tumor in dogs--incidence and histopathological characterization. Coll. Antropol. 2009, 33, 253–258. [Google Scholar] [PubMed]
- Ramos-Vara, J.A.; Beissenherz, M.E.; Miller, M.A.; Johnson, G.C.; Pace, L.W.; Fard, A.; Kottler, S.J. Retrospective study of 338 canine oral melanomas with clinical, histologic, and immunohistochemical review of 129 cases. Vet. Pathol. 2000, 37, 597–608. [Google Scholar] [CrossRef] [PubMed]
- Egenvall, A.; Nodtvedt, A.; von Euler, H. Bone tumors in a population of 400 000 insured swedish dogs up to 10 y of age: Incidence and survival. Can. J. Vet. Res. 2007, 71, 292–299. [Google Scholar] [PubMed]
- Salas, Y.; Marquez, A.; Diaz, D.; Romero, L. Epidemiological study of mammary tumors in female dogs diagnosed during the period 2002-2012: A growing animal health problem. PLoS ONE 2015, 10, e0127381. [Google Scholar] [CrossRef] [PubMed]
- Innes, J.R.; Parry, H.B.; Berger, J. Leukaemia in dogs; including a record of a case treated by urethane. Br. Vet. J. 1946, 102, 383–393. [Google Scholar] [PubMed]
- Brick, J.O.; Roenigk, W.J.; Wilson, G.P. Chemotherapy of malignant lymphoma in dogs and cats. J. Am. Vet. Med. Assoc. 1968, 153, 47–52. [Google Scholar] [PubMed]
- Wang, S.L.; Lee, J.J.; Liao, A.T. Comparison of efficacy and toxicity of doxorubicin and mitoxantrone in combination chemotherapy for canine lymphoma. Can. Vet. J. 2016, 57, 271–276. [Google Scholar] [PubMed]
- Kiupel, M.; Webster, J.D.; Bailey, K.L.; Best, S.; DeLay, J.; Detrisac, C.J.; Fitzgerald, S.D.; Gamble, D.; Ginn, P.E.; Goldschmidt, M.H.; et al. Proposal of a 2-tier histologic grading system for canine cutaneous mast cell tumors to more accurately predict biological behavior. Vet. Pathol. 2011, 48, 147–155. [Google Scholar] [CrossRef] [PubMed]
- Shearin, A.L.; Hedan, B.; Cadieu, E.; Erich, S.A.; Schmidt, E.V.; Faden, D.L.; Cullen, J.; Abadie, J.; Kwon, E.M.; Grone, A.; et al. The mtap-cdkn2a locus confers susceptibility to a naturally occurring canine cancer. Cancer Epidemiol. Biomarkers Prev. 2012, 21, 1019–1027. [Google Scholar] [CrossRef] [PubMed]
- Im, K.S.; Kim, N.H.; Lim, H.Y.; Kim, H.W.; Shin, J.I.; Sur, J.H. Analysis of a new histological and molecular-based classification of canine mammary neoplasia. Vet. Pathol. 2014, 51, 549–559. [Google Scholar] [CrossRef] [PubMed]
- Bushell, K.R.; Kim, Y.; Chan, F.C.; Ben-Neriah, S.; Jenks, A.; Alcaide, M.; Fornika, D.; Grande, B.M.; Arthur, S.; Gascoyne, R.D.; et al. Genetic inactivation of traf3 in canine and human b-cell lymphoma. Blood 2015, 125, 999–1005. [Google Scholar] [CrossRef] [PubMed]
- Yurchenko, K.S.; Zhou, P.; Kovner, A.V.; Zavjalov, E.L.; Shestopalova, L.V.; Shestopalov, A.M. Oncolytic effect of wild-type newcastle disease virus isolates in cancer cell lines in vitro and in vivo on xenograft model. PLoS ONE 2018, 13, e0195425. [Google Scholar] [CrossRef] [PubMed]
- Hwang, C.C.; Umeki, S.; Igase, M.; Coffey, M.; Noguchi, S.; Okuda, M.; Mizuno, T. The effects of oncolytic reovirus in canine lymphoma cell lines. Vet. Comp. Oncol. 2016, 14 (Suppl. 1), 61–73. [Google Scholar] [CrossRef]
- Mostafa, A.A.; Meyers, D.E.; Thirukkumaran, C.M.; Liu, P.J.; Gratton, K.; Spurrell, J.; Shi, Q.; Thakur, S.; Morris, D.G. Oncolytic reovirus and immune checkpoint inhibition as a novel immunotherapeutic strategy for breast cancer. Cancers 2018, 10, E205. [Google Scholar] [CrossRef] [PubMed]
- Bierman, H.R.; Crile, D.M.; Dod, K.S.; Kelly, K.H.; Petrakis, N.L.; White, L.P.; Shimkin, M.B. Remissions in leukemia of childhood following acute infectious disease: Staphylococcus and streptococcus, varicella, and feline panleukopenia. Cancer 1953, 6, 591–605. [Google Scholar] [CrossRef]
- Pelner, L.; Fowler, G.A.; Nauts, H.C. Effects of concurrent infections and their toxins on the course of leukemia. Acta Med. Scand. 1958, 338, 1–47. [Google Scholar] [CrossRef]
- Bluming, A.Z.; Ziegler, J.L. Regression of burkitt‘s lymphoma in association with measles infection. Lancet 1971, 2, 105–106. [Google Scholar] [CrossRef]
- Gross, S. Measles and leukaemia. Lancet 1971, 1, 397–398. [Google Scholar] [CrossRef]
- Huebner, R.J.; Rowe, W.P.; Schatten, W.E.; Smith, R.R.; Thomas, L.B. Studies on the use of viruses in the treatment of carcinoma of the cervix. Cancer 1956, 9, 1211–1218. [Google Scholar] [PubMed]
- Pack, G.T. Note on the experimental use of rabies vaccine for melanomatosis. AMA Arch. Derm. Syphilol. 1950, 62, 694–695. [Google Scholar] [CrossRef] [PubMed]
- Southam, C.M.; Moore, A.E. Clinical studies of viruses as antineoplastic agents with particular reference to egypt 101 virus. Cancer 1952, 5, 1025–1034. [Google Scholar] [CrossRef]
- Lerner, B.H. Sins of omission--cancer research without informed consent. N. Engl. J. Med. 2004, 351, 628–630. [Google Scholar] [CrossRef] [PubMed]
- Garber, K. China approves worlds first oncolytic virus therapy for cancer treatment. J. Natl. Cancer Inst. 2006, 98, 298–300. [Google Scholar] [CrossRef] [PubMed]
- Dolgin, E. Oncolytic viruses get a boost with first fda-approval recommendation. Nat. Rev. Drug Discov. 2015, 14, 369–371. [Google Scholar] [CrossRef] [PubMed]
- Fields, B.N.; Knipe, D.M.; Howley, P.M. Fields Virology; Wolters Kluwer Health/Lippincott Williams & Wilkins: Philadelphia, PA, USA, 2013. [Google Scholar]
- Farzan, S.F.; Palermo, L.M.; Yokoyama, C.C.; Orefice, G.; Fornabaio, M.; Sarkar, A.; Kellogg, G.E.; Greengard, O.; Porotto, M.; Moscona, A. Premature activation of the paramyxovirus fusion protein before target cell attachment with corruption of the viral fusion machinery. J. Biol. Chem. 2011, 286, 37945–37954. [Google Scholar] [CrossRef] [PubMed]
- Cantin, C.; Holguera, J.; Ferreira, L.; Villar, E.; Munoz-Barroso, I. Newcastle disease virus may enter cells by caveolae-mediated endocytosis. J. Gen. Virol. 2007, 88, 559–569. [Google Scholar] [CrossRef] [PubMed]
- Goncalves-Carneiro, D.; McKeating, J.A.; Bailey, D. The measles virus receptor slamf1 can mediate particle endocytosis. J. Virol. 2017, 91, e02255-16. [Google Scholar] [CrossRef] [PubMed]
- Mansour, M.; Palese, P.; Zamarin, D. Oncolytic specificity of newcastle disease virus is mediated by selectivity for apoptosis-resistant cells. J. Virol. 2011, 85, 6015–6023. [Google Scholar] [CrossRef] [PubMed]
- Zygiert, Z. Hodgkin‘s disease: Remissions after measles. Lancet 1971, 1, 593. [Google Scholar] [CrossRef]
- Taqi, A.M.; Abdurrahman, M.B.; Yakubu, A.M.; Fleming, A.F. Regression of hodgkin‘s disease after measles. Lancet 1981, 1, 1112. [Google Scholar] [CrossRef]
- Beck, M.; Smerdel, S.; Dedic, I.; Delimar, N.; Rajninger-Miholic, M.; Juzbasic, M.; Manhalter, T.; Vlatkovic, R.; Borcic, B.; Mihajic, Z. Immune response to edmonston-zagreb measles virus strain in monovalent and combined mmr vaccine. Dev. Biol. Stand. 1986, 65, 95–100. [Google Scholar] [PubMed]
- Grote, D.; Russell, S.J.; Cornu, T.I.; Cattaneo, R.; Vile, R.; Poland, G.A.; Fielding, A.K. Live attenuated measles virus induces regression of human lymphoma xenografts in immunodeficient mice. Blood 2001, 97, 3746–3754. [Google Scholar] [CrossRef] [PubMed]
- Noyce, R.S.; Richardson, C.D. Nectin 4 is the epithelial cell receptor for measles virus. Trends Microbiol. 2012, 20, 429–439. [Google Scholar] [CrossRef] [PubMed]
- Muhlebach, M.D.; Mateo, M.; Sinn, P.L.; Prufer, S.; Uhlig, K.M.; Leonard, V.H.; Navaratnarajah, C.K.; Frenzke, M.; Wong, X.X.; Sawatsky, B.; et al. Adherens junction protein nectin-4 is the epithelial receptor for measles virus. Nature 2011, 480, 530–533. [Google Scholar] [CrossRef] [PubMed]
- Derycke, M.S.; Pambuccian, S.E.; Gilks, C.B.; Kalloger, S.E.; Ghidouche, A.; Lopez, M.; Bliss, R.L.; Geller, M.A.; Argenta, P.A.; Harrington, K.M.; et al. Nectin 4 overexpression in ovarian cancer tissues and serum: Potential role as a serum biomarker. Am. J. Clin. Pathol. 2010, 134, 835–845. [Google Scholar] [CrossRef] [PubMed]
- Fabre-Lafay, S.; Monville, F.; Garrido-Urbani, S.; Berruyer-Pouyet, C.; Ginestier, C.; Reymond, N.; Finetti, P.; Sauvan, R.; Adelaide, J.; Geneix, J.; et al. Nectin-4 is a new histological and serological tumor associated marker for breast cancer. BMC Cancer 2007, 7, 73. [Google Scholar] [CrossRef] [PubMed]
- Takano, A.; Ishikawa, N.; Nishino, R.; Masuda, K.; Yasui, W.; Inai, K.; Nishimura, H.; Ito, H.; Nakayama, H.; Miyagi, Y.; et al. Identification of nectin-4 oncoprotein as a diagnostic and therapeutic target for lung cancer. Cancer Res. 2009, 69, 6694–6703. [Google Scholar] [CrossRef] [PubMed]
- Sugiyama, T.; Yoneda, M.; Kuraishi, T.; Hattori, S.; Inoue, Y.; Sato, H.; Kai, C. Measles virus selectively blind to signaling lymphocyte activation molecule as a novel oncolytic virus for breast cancer treatment. Gene Ther. 2013, 20, 338–347. [Google Scholar] [CrossRef] [PubMed]
- Shoji, K.; Yoneda, M.; Fujiyuki, T.; Amagai, Y.; Tanaka, A.; Matsuda, A.; Ogihara, K.; Naya, Y.; Ikeda, F.; Matsuda, H.; et al. Development of new therapy for canine mammary cancer with recombinant measles virus. Mol. Ther. Oncolytics 2016, 3, 15022. [Google Scholar] [CrossRef] [PubMed]
- Betz, D.; Schoenrock, D.; Mischke, R.; Baumgartner, W.; Nolte, I. Postoperative treatment outcome in canine mammary tumors. Multivariate analysis of the prognostic value of pre- and postoperatively available information. Tierarztl. Prax. Ausg. K Kleintiere Heimtiere 2012, 40, 235–242. [Google Scholar] [PubMed]
- Pratakpiriya, W.; Seki, F.; Otsuki, N.; Sakai, K.; Fukuhara, H.; Katamoto, H.; Hirai, T.; Maenaka, K.; Techangamsuwan, S.; Lan, N.T.; et al. Nectin4 is an epithelial cell receptor for canine distemper virus and involved in neurovirulence. J. Virol. 2012, 86, 10207–10210. [Google Scholar] [CrossRef] [PubMed]
- Sawatsky, B.; Wong, X.X.; Hinkelmann, S.; Cattaneo, R.; von Messling, V. Canine distemper virus epithelial cell infection is required for clinical disease but not for immunosuppression. J. Virol. 2012, 86, 3658–3666. [Google Scholar] [CrossRef] [PubMed]
- Qeska, V.; Barthel, Y.; Herder, V.; Stein, V.M.; Tipold, A.; Urhausen, C.; Gunzel-Apel, A.R.; Rohn, K.; Baumgartner, W.; Beineke, A. Canine distemper virus infection leads to an inhibitory phenotype of monocyte-derived dendritic cells in vitro with reduced expression of co-stimulatory molecules and increased interleukin-10 transcription. PLoS ONE 2014, 9, e96121. [Google Scholar] [CrossRef] [PubMed]
- Schobesberger, M.; Summerfield, A.; Doherr, M.G.; Zurbriggen, A.; Griot, C. Canine distemper virus-induced depletion of uninfected lymphocytes is associated with apoptosis. Vet. Immunol. Immunopathol. 2005, 104, 33–44. [Google Scholar] [CrossRef] [PubMed]
- Khosravi, M.; Bringolf, F.; Röthlisberger, S.; Bieringer, M.; Schneider-Schaulies, J.; Zurbriggen, A.; Origgi, F.; Plattet, P. Canine distemper virus fusion activation: Critical role of residue e123 of cd150/slam. J. Virol. 2016, 90, 1622–1637. [Google Scholar] [CrossRef] [PubMed]
- Del Puerto, H.L.; Martins, A.S.; Milsted, A.; Souza-Fagundes, E.M.; Braz, G.F.; Hissa, B.; Andrade, L.O.; Alves, F.; Rajao, D.S.; Leite, R.C.; et al. Canine distemper virus induces apoptosis in cervical tumor derived cell lines. Virol. J. 2011, 8, 334. [Google Scholar] [CrossRef] [PubMed]
- Suter, S.E.; Chein, M.B.; von Messling, V.; Yip, B.; Cattaneo, R.; Vernau, W.; Madewell, B.R.; London, C.A. In vitro canine distemper virus infection of canine lymphoid cells: A prelude to oncolytic therapy for lymphoma. Clin. Cancer Res. 2005, 11, 1579–1587. [Google Scholar] [CrossRef] [PubMed]
- Grone, A.; Fonfara, S.; Baumgartner, W. Cell type-dependent cytokine expression after canine distemper virus infection. Viral Immunol. 2002, 15, 493–505. [Google Scholar] [CrossRef] [PubMed]
- Yamaguchi, R.; Kojimoto, A.; Sakai, H.; Uchida, K.; Sugano, S.; Tateyama, S. Growth characteristics of canine distemper virus in a new cell line cct cells originated from canine malignant histiocytosis. J. Vet. Med. Sci. 2005, 67, 203–206. [Google Scholar] [CrossRef] [PubMed]
- Puff, C.; Krudewig, C.; Imbschweiler, I.; Baumgartner, W.; Alldinger, S. Influence of persistent canine distemper virus infection on expression of reck, matrix-metalloproteinases and their inhibitors in a canine macrophage/monocytic tumour cell line (dh82). Vet. J. 2009, 182, 100–107. [Google Scholar] [CrossRef] [PubMed]
- Dervisis, N.G.; Kiupel, M.; Qin, Q.; Cesario, L. Clinical prognostic factors in canine histiocytic sarcoma. Vet. Comp. Oncol. 2017, 15, 1171–1180. [Google Scholar] [CrossRef] [PubMed]
- Henson, M.S.; Suter, S.E.; von Messling, V.A.; Cattaneo, R.; Fielding, A.K. 803. The effects of intratumoral injection of a replicating morbillivirus in a canine model of naturally occurring lymphoma. Mol. Ther. 2005, 11, 312. [Google Scholar]
- Ford, R.B.; Larson, L.J.; McClure, K.D.; Schultz, R.D.; Welborn, L.V. 2017 aaha canine vaccination guidelines. J. Am. Anim. Hosp. Assoc. 2017, 53, 243–251. [Google Scholar] [CrossRef] [PubMed]
- Nelson, C.B.; Pomeroy, B.S.; Schrall, K.; Park, W.E.; Lindeman, R.J. An outbreak of conjunctivitis due to newcastle disease virus (ndv) occurring in poultry workers. Am. J. Public Health Nations Health 1952, 42, 672–678. [Google Scholar] [CrossRef] [PubMed]
- Zulkifli, M.M.; Ibrahim, R.; Ali, A.M.; Aini, I.; Jaafar, H.; Hilda, S.S.; Alitheen, N.B.; Abdullah, J.M. Newcastle diseases virus strain v4upm displayed oncolytic ability against experimental human malignant glioma. Neurol. Res. 2009, 31, 3–10. [Google Scholar] [CrossRef] [PubMed]
- Bian, J.; Wang, K.; Kong, X.; Liu, H.; Chen, F.; Hu, M.; Zhang, X.; Jiao, X.; Ge, B.; Wu, Y.; et al. Caspase- and p38-mapk-dependent induction of apoptosis in a549 lung cancer cells by newcastle disease virus. Arch. Virol. 2011, 156, 1335–1344. [Google Scholar] [CrossRef] [PubMed]
- Buijs, P.R.; van Eijck, C.H.; Hofland, L.J.; Fouchier, R.A.; van den Hoogen, B.G. Different responses of human pancreatic adenocarcinoma cell lines to oncolytic newcastle disease virus infection. Cancer Gene Ther. 2014, 21, 24–30. [Google Scholar] [CrossRef] [PubMed]
- Sanchez, D.; Pelayo, R.; Sarmiento, R.E.; Medina, L.A.; Cesarman-Maus, G.N.; Nuñez, L.; Carrillo, N.; de Jesus Paredes, J.; Corona, H.; Vadillo, E. In Vitro and in Vivo oncolytic Activity of Lasota Strain of Newcastle Disease Virus on a Lymphoma B-Cell Line and a Canine Cutaneous T-Cell Lymphoma. Blood 2014, 124, 5504. [Google Scholar]
- Sanchez, D.; Pelayo, R.; Medina, L.A.; Vadillo, E.; Sanchez, R.; Nunez, L.; Cesarman-Maus, G.; Sarmiento-Silva, R.E. Newcastle disease virus: Potential therapeutic application for human and canine lymphoma. Viruses 2016, 8, E3. [Google Scholar] [CrossRef] [PubMed]
- Jarahian, M.; Watzl, C.; Fournier, P.; Arnold, A.; Djandji, D.; Zahedi, S.; Cerwenka, A.; Paschen, A.; Schirrmacher, V.; Momburg, F. Activation of natural killer cells by newcastle disease virus hemagglutinin-neuraminidase. J. Virol. 2009, 83, 8108–8121. [Google Scholar] [CrossRef] [PubMed]
- Koks, C.A.; Garg, A.D.; Ehrhardt, M.; Riva, M.; Vandenberk, L.; Boon, L.; De Vleeschouwer, S.; Agostinis, P.; Graf, N.; Van Gool, S.W. Newcastle disease virotherapy induces long-term survival and tumor-specific immune memory in orthotopic glioma through the induction of immunogenic cell death. Int. J. Cancer 2015, 136, E313–E325. [Google Scholar] [CrossRef] [PubMed]
- Termeer, C.C.; Schirrmacher, V.; Brocker, E.B.; Becker, J.C. Newcastle disease virus infection induces b7-1/b7-2-independent t-cell costimulatory activity in human melanoma cells. Cancer Gene Ther. 2000, 7, 316–323. [Google Scholar] [CrossRef] [PubMed]
- Chang, C.Y.; Tai, J.A.; Li, S.; Nishikawa, T.; Kaneda, Y. Virus-stimulated neutrophils in the tumor microenvironment enhance t cell-mediated anti-tumor immunity. Oncotarget 2016, 7, 42195–42207. [Google Scholar] [CrossRef] [PubMed]
- Kurooka, M.; Kaneda, Y. Inactivated sendai virus particles eradicate tumors by inducing immune responses through blocking regulatory t cells. Cancer Res. 2007, 67, 227–236. [Google Scholar] [CrossRef] [PubMed]
- Tanaka, M.; Shimbo, T.; Kikuchi, Y.; Matsuda, M.; Kaneda, Y. Sterile alpha motif containing domain 9 is involved in death signaling of malignant glioma treated with inactivated sendai virus particle (hvj-e) or type i interferon. Int. J. Cancer 2010, 126, 1982–1991. [Google Scholar] [CrossRef] [PubMed]
- Ilyinskaya, G.V.; Mukhina, E.V.; Soboleva, A.V.; Matveeva, O.V.; Chumakov, P.M. Oncolytic sendai virus therapy of canine mast cell tumors (a pilot study). Front. Vet. Sci. 2018, 5, 116. [Google Scholar] [CrossRef] [PubMed]
- Donnelly, L.; Mullin, C.; Balko, J.; Goldschmidt, M.; Krick, E.; Hume, C.; Brown, D.C.; Sorenmo, K. Evaluation of histological grade and histologically tumour-free margins as predictors of local recurrence in completely excised canine mast cell tumours. Vet. Comp. Oncol. 2015, 13, 70–76. [Google Scholar] [CrossRef] [PubMed]
- Fujihara, A.; Kurooka, M.; Miki, T.; Kaneda, Y. Intratumoral injection of inactivated sendai virus particles elicits strong antitumor activity by enhancing local cxcl10 expression and systemic nk cell activation. Cancer Immunol. Immunother. 2008, 57, 73–84. [Google Scholar] [CrossRef] [PubMed]
- Piccinotti, S.; Kirchhausen, T.; Whelan, S.P. Uptake of rabies virus into epithelial cells by clathrin-mediated endocytosis depends upon actin. J. Virol. 2013, 87, 11637–11647. [Google Scholar] [CrossRef] [PubMed]
- Scherer, C.F.; O‘Donnell, V.; Golde, W.T.; Gregg, D.; Estes, D.M.; Rodriguez, L.L. Vesicular stomatitis new jersey virus (vsnjv) infects keratinocytes and is restricted to lesion sites and local lymph nodes in the bovine, a natural host. Vet. Res. 2007, 38, 375–390. [Google Scholar] [CrossRef] [PubMed]
- Quiroz, E.; Moreno, N.; Peralta, P.H.; Tesh, R.B. A human case of encephalitis associated with vesicular stomatitis virus (indiana serotype) infection. Am. J. Trop. Med. Hyg. 1988, 39, 312–314. [Google Scholar] [CrossRef] [PubMed]
- LeBlanc, A.K.; Naik, S.; Galyon, G.D.; Jenks, N.; Steele, M.; Peng, K.W.; Federspiel, M.J.; Donnell, R.; Russell, S.J. Safety studies on intravenous administration of oncolytic recombinant vesicular stomatitis virus in purpose-bred beagle dogs. Hum. Gene Ther. Clin. Dev. 2013, 24, 174–181. [Google Scholar] [CrossRef] [PubMed]
- Shen, W.; Patnaik, M.M.; Ruiz, A.; Russell, S.J.; Peng, K.W. Immunovirotherapy with vesicular stomatitis virus and pd-l1 blockade enhances therapeutic outcome in murine acute myeloid leukemia. Blood 2016, 127, 1449–1458. [Google Scholar] [CrossRef] [PubMed]
- Naik, S.; Galyon, G.D.; Jenks, N.J.; Steele, M.B.; Miller, A.C.; Allstadt, S.D.; Suksanpaisan, L.; Peng, K.W.; Federspiel, M.J.; Russell, S.J.; et al. Comparative oncology evaluation of intravenous recombinant oncolytic vesicular stomatitis virus therapy in spontaneous canine cancer. Mol. Cancer Ther. 2018, 17, 316–326. [Google Scholar] [CrossRef] [PubMed]
- Jenks, N.; Myers, R.; Greiner, S.M.; Thompson, J.; Mader, E.K.; Greenslade, A.; Griesmann, G.E.; Federspiel, M.J.; Rakela, J.; Borad, M.J.; et al. Safety studies on intrahepatic or intratumoral injection of oncolytic vesicular stomatitis virus expressing interferon-β in rodents and nonhuman primates. Hum. Gene Ther. 2010, 21, 451–462. [Google Scholar] [CrossRef] [PubMed]
- Muik, A.; Kneiske, I.; Werbizki, M.; Wilflingseder, D.; Giroglou, T.; Ebert, O.; Kraft, A.; Dietrich, U.; Zimmer, G.; Momma, S.; et al. Pseudotyping vesicular stomatitis virus with lymphocytic choriomeningitis virus glycoproteins enhances infectivity for glioma cells and minimizes neurotropism. J. Virol. 2011, 85, 5679–5684. [Google Scholar] [CrossRef] [PubMed]
- Kimpel, J.; Urbiola, C.; Koske, I.; Tober, R.; Banki, Z.; Wollmann, G.; von Laer, D. The oncolytic virus vsv-gp is effective against malignant melanoma. Viruses 2018, 10, E108. [Google Scholar] [CrossRef] [PubMed]
- Vaha-Koskela, M.J.; Tuittila, M.T.; Nygardas, P.T.; Nyman, J.K.; Ehrengruber, M.U.; Renggli, M.; Hinkkanen, A.E. A novel neurotropic expression vector based on the avirulent a7(74) strain of semliki forest virus. J. Neurovirol. 2003, 9, 1–15. [Google Scholar] [CrossRef] [PubMed]
- Fros, J.J.; Pijlman, G.P. Alphavirus infection: Host cell shut-off and inhibition of antiviral responses. Viruses 2016, 8, E166. [Google Scholar] [CrossRef] [PubMed]
- Mathiot, C.C.; Grimaud, G.; Garry, P.; Bouquety, J.C.; Mada, A.; Daguisy, A.M.; Georges, A.J. An outbreak of human semliki forest virus infections in central african republic. Am. J. Trop. Med. Hyg. 1990, 42, 386–393. [Google Scholar] [CrossRef] [PubMed]
- Willems, W.R.; Kaluza, G.; Boschek, C.B.; Bauer, H.; Hager, H.; Schutz, H.J.; Feistner, H. Semliki forest virus: Cause of a fatal case of human encephalitis. Science 1979, 203, 1127–1129. [Google Scholar] [CrossRef] [PubMed]
- Bradish, C.J.; Allner, K.; Maber, H.B. The virulence of original and derived strains of semliki forest virus for mice, guinea-pigs and rabbits. J. Gen. Virol. 1971, 12, 141–160. [Google Scholar] [CrossRef] [PubMed]
- Reagan, R.L.; Strand, N.; Brueckner, A.L. Bwamba fever virus and semliki forest virus in young dogs. Proc. Soc. Exp. Biol. Med. 1953, 82, 642–643. [Google Scholar] [CrossRef] [PubMed]
- Vaha-Koskela, M.J.; Kallio, J.P.; Jansson, L.C.; Heikkila, J.E.; Zakhartchenko, V.A.; Kallajoki, M.A.; Kahari, V.M.; Hinkkanen, A.E. Oncolytic capacity of attenuated replicative semliki forest virus in human melanoma xenografts in severe combined immunodeficient mice. Cancer Res. 2006, 66, 7185–7194. [Google Scholar] [CrossRef] [PubMed]
- Heikkila, J.E.; Vaha-Koskela, M.J.; Ruotsalainen, J.J.; Martikainen, M.W.; Stanford, M.M.; McCart, J.A.; Bell, J.C.; Hinkkanen, A.E. Intravenously administered alphavirus vector va7 eradicates orthotopic human glioma xenografts in nude mice. PLoS ONE 2010, 5, e8603. [Google Scholar] [CrossRef] [PubMed]
- Ruotsalainen, J.J.; Kaikkonen, M.U.; Niittykoski, M.; Martikainen, M.W.; Lemay, C.G.; Cox, J.; De Silva, N.S.; Kus, A.; Falls, T.J.; Diallo, J.S.; et al. Clonal variation in interferon response determines the outcome of oncolytic virotherapy in mouse ct26 colon carcinoma model. Gene Ther. 2015, 22, 65–75. [Google Scholar] [CrossRef] [PubMed]
- Autio, K.P.; Ruotsalainen, J.J.; Anttila, M.O.; Niittykoski, M.; Waris, M.; Hemminki, A.; Vaha-Koskela, M.J.; Hinkkanen, A.E. Attenuated semliki forest virus for cancer treatment in dogs: Safety assessment in two laboratory beagles. BMC Vet. Res. 2015, 11, 170. [Google Scholar] [CrossRef] [PubMed]
- Nishiya, A.T.; Massoco, C.O.; Felizzola, C.R.; Perlmann, E.; Batschinski, K.; Tedardi, M.V.; Garcia, J.S.; Mendonça, P.P.; Teixeira, T.F.; Zaidan Dagli, M.L. Comparative aspects of canine melanoma. Vet. Sci. 2016, 3, E7. [Google Scholar] [CrossRef] [PubMed]
- Cameron, D.J.; Bishop, R.F.; Veenstra, A.A.; Barnes, G.L.; Holmes, I.H.; Ruck, B.J. Pattern of shedding of two noncultivable viruses in stools of newborn babies. J. Med. Virol. 1978, 2, 7–13. [Google Scholar] [CrossRef] [PubMed]
- Hwang, C.C.; Mochizuki, M.; Maeda, K.; Okuda, M.; Mizuno, T. Seroepidemiology of reovirus in healthy dogs in six prefectures in japan. J. Vet. Med. Sci. 2014, 76, 471–475. [Google Scholar] [CrossRef] [PubMed]
- Selb, B.; Weber, B. A study of human reovirus igg and iga antibodies by elisa and western blot. J. Virol. Methods 1994, 47, 15–25. [Google Scholar] [CrossRef]
- Tai, J.H.; Williams, J.V.; Edwards, K.M.; Wright, P.F.; Crowe, J.E., Jr.; Dermody, T.S. Prevalence of reovirus-specific antibodies in young children in nashville, tennessee. J. Infect. Dis. 2005, 191, 1221–1224. [Google Scholar] [CrossRef] [PubMed]
- Gong, J.; Sachdev, E.; Mita, A.C.; Mita, M.M. Clinical development of reovirus for cancer therapy: An oncolytic virus with immune-mediated antitumor activity. World J. Methodol. 2016, 6, 25–42. [Google Scholar] [CrossRef] [PubMed]
- Hwang, C.C.; Umeki, S.; Kubo, M.; Hayashi, T.; Shimoda, H.; Mochizuki, M.; Maeda, K.; Baba, K.; Hiraoka, H.; Coffey, M.; et al. Oncolytic reovirus in canine mast cell tumor. PLoS ONE 2013, 8, e73555. [Google Scholar] [CrossRef] [PubMed]
- Norman, K.L.; Hirasawa, K.; Yang, A.D.; Shields, M.A.; Lee, P.W. Reovirus oncolysis: The ras/ralgef/p38 pathway dictates host cell permissiveness to reovirus infection. Proc. Natl. Acad. Sci. USA. 2004, 101, 11099–11104. [Google Scholar] [CrossRef] [PubMed]
- Igase, M.; Hwang, C.C.; Coffey, M.; Okuda, M.; Noguchi, S.; Mizuno, T. The oncolytic effects of reovirus in canine solid tumor cell lines. J. Vet. Med. Sci. 2015, 77, 541–548. [Google Scholar] [CrossRef] [PubMed]
- Hwang, C.C.; Igase, M.; Sakurai, M.; Haraguchi, T.; Tani, K.; Itamoto, K.; Shimokawa, T.; Nakaichi, M.; Nemoto, Y.; Noguchi, S.; et al. Oncolytic reovirus therapy: Pilot study in dogs with spontaneously occurring tumours. Vet. Comp. Oncol. 2017, 16, 229–238. [Google Scholar] [CrossRef] [PubMed]
- Mahalingam, D.; Fountzilas, C.; Moseley, J.; Noronha, N.; Tran, H.; Chakrabarty, R.; Selvaggi, G.; Coffey, M.; Thompson, B.; Sarantopoulos, J. A phase ii study of reolysin((r)) (pelareorep) in combination with carboplatin and paclitaxel for patients with advanced malignant melanoma. Cancer Chemother. Pharmacol. 2017, 79, 697–703. [Google Scholar] [CrossRef] [PubMed]
- Saunders, M.; Anthoney, A.; Coffey, M.; Mettinger, K.; Thompson, B.; Melcher, A.; Nutting, C.M.; Harrington, K. Results of a phase ii study to evaluate the biological effects of intratumoral (itu) reolysin in combination with low dose radiotherapy (rt) in patients (pts) with advanced cancers. J. Clin. Oncol. 2009, 27, e14514. [Google Scholar]
- Jaime-Ramirez, A.C.; Yu, J.G.; Caserta, E.; Yoo, J.Y.; Zhang, J.; Lee, T.J.; Hofmeister, C.; Lee, J.H.; Kumar, B.; Pan, Q.; et al. Reolysin and histone deacetylase inhibition in the treatment of head and neck squamous cell carcinoma. Mol. Ther. Oncolytics 2017, 5, 87–96. [Google Scholar] [CrossRef] [PubMed]
- Kapoor, A.; Simmonds, P.; Lipkin, W.I. Discovery and characterization of mammalian endogenous parvoviruses. J. Virol. 2010, 84, 12628–12635. [Google Scholar] [CrossRef] [PubMed]
- Malerba, M.; Daeffler, L.; Rommelaere, J.; Iggo, R.D. Replicating parvoviruses that target colon cancer cells. J. Virol. 2003, 77, 6683–6691. [Google Scholar] [CrossRef] [PubMed]
- Grekova, S.P.; Raykov, Z.; Zawatzky, R.; Rommelaere, J.; Koch, U. Activation of a glioma-specific immune response by oncolytic parvovirus minute virus of mice infection. Cancer Gene Ther. 2012, 19, 468–475. [Google Scholar] [CrossRef] [PubMed]
- Angelova, A.L.; Barf, M.; Geletneky, K.; Unterberg, A.; Rommelaere, J. Immunotherapeutic potential of oncolytic h-1 parvovirus: Hints of glioblastoma microenvironment conversion towards immunogenicity. Viruses 2017, 9, 382. [Google Scholar] [CrossRef] [PubMed]
- Paglino, J.C.; Ozduman, K.; van den Pol, A.N. Luiii parvovirus selectively and efficiently targets, replicates in, and kills human glioma cells. J. Virol. 2012, 86, 7280–7291. [Google Scholar] [CrossRef] [PubMed]
- Nykky, J.; Tuusa, J.E.; Kirjavainen, S.; Vuento, M.; Gilbert, L. Mechanisms of cell death in canine parvovirus-infected cells provide intuitive insights to developing nanotools for medicine. Int. J. Nanomed. 2010, 5, 417–428. [Google Scholar]
- Singh, P.; Destito, G.; Schneemann, A.; Manchester, M. Canine parvovirus-like particles, a novel nanomaterial for tumor targeting. J. Nanobiotechnol. 2006, 4, 2. [Google Scholar] [CrossRef] [PubMed]
- Cooper, B.J.; Carmichael, L.E.; Appel, M.J.; Greisen, H. Canine viral enteritis. Ii. Morphologic lesions in naturally occurring parvovirus infection. Cornell Vet. 1979, 69, 134–144. [Google Scholar] [PubMed]
- Martin, V.; Najbar, W.; Gueguen, S.; Grousson, D.; Eun, H.M.; Lebreux, B.; Aubert, A. Treatment of canine parvoviral enteritis with interferon-omega in a placebo-controlled challenge trial. Vet. Microbiol. 2002, 89, 115–127. [Google Scholar] [CrossRef]
- Ford, J.; McEndaffer, L.; Renshaw, R.; Molesan, A.; Kelly, K. Parvovirus infection is associated with myocarditis and myocardial fibrosis in young dogs. Vet. Pathol. 2017, 54, 964–971. [Google Scholar] [CrossRef] [PubMed]
- Mech, L.D.; Goyal, S.M.; Paul, W.J.; Newton, W.E. Demographic effects of canine parvovirus on a free-ranging wolf population over 30 years. J. Wildl. Dis. 2008, 44, 824–836. [Google Scholar] [CrossRef] [PubMed]
- Abdelmagid, O.Y.; Larson, L.; Payne, L.; Tubbs, A.; Wasmoen, T.; Schultz, R. Evaluation of the efficacy and duration of immunity of a canine combination vaccine against virulent parvovirus, infectious canine hepatitis virus, and distemper virus experimental challenges. Vet. Ther. 2004, 5, 173–186. [Google Scholar] [PubMed]
- Gore, T.C.; Lakshmanan, N.; Duncan, K.L.; Coyne, M.J.; Lum, M.A.; Sterner, F.J. Three-year duration of immunity in dogs following vaccination against canine adenovirus type-1, canine parvovirus, and canine distemper virus. Vet. Ther. 2005, 6, 5–14. [Google Scholar] [PubMed]
- Bergman, J.G.; Muniz, M.; Sutton, D.; Fensome, R.; Ling, F.; Paul, G. Comparative trial of the canine parvovirus, canine distemper virus and canine adenovirus type 2 fractions of two commercially available modified live vaccines. Vet. Rec. 2006, 159, 733–736. [Google Scholar] [CrossRef] [PubMed]
- Palermo, L.M.; Hueffer, K.; Parrish, C.R. Residues in the apical domain of the feline and canine transferrin receptors control host-specific binding and cell infection of canine and feline parvoviruses. J. Virol. 2003, 77, 8915–8923. [Google Scholar] [CrossRef] [PubMed]
- Miller, M.A.; Towle, H.A.; Heng, H.G.; Greenberg, C.B.; Pool, R.R. Mandibular ossifying fibroma in a dog. Vet. Pathol. 2008, 45, 203–206. [Google Scholar] [CrossRef] [PubMed]
- Miles, C.R.; Bell, C.M.; Pinkerton, M.E.; Soukup, J.W. Maxillary ameloblastic fibroma in a dog. Vet. Pathol. 2011, 48, 823–826. [Google Scholar] [CrossRef] [PubMed]
- Soltero-Rivera, M.; Engiles, J.B.; Reiter, A.M.; Reetz, J.; Lewis, J.R.; Sanchez, M.D. Benign and malignant proliferative fibro-osseous and osseous lesions of the oral cavity of dogs. Vet. Pathol. 2015, 52, 894–902. [Google Scholar] [CrossRef] [PubMed]
- Phattarataratip, E.; Pholjaroen, C.; Tiranon, P. A clinicopathologic analysis of 207 cases of benign fibro-osseous lesions of the jaws. Int. J. Surg. Pathol. 2014, 22, 326–333. [Google Scholar] [CrossRef] [PubMed]
- Fox, L.E.; Geoghegan, S.L.; Davis, L.H.; Hartzel, J.S.; Kubilis, P.; Gruber, L.A. Owner satisfaction with partial mandibulectomy or maxillectomy for treatment of oral tumors in 27 dogs. J. Am. Anim. Hosp. Assoc. 1997, 33, 25–31. [Google Scholar] [CrossRef] [PubMed]
- Martin, S.J. Cell death and inflammation: The case for il-1 family cytokines as the canonical damps of the immune system. FEBS J. 2016, 283, 2599–2615. [Google Scholar] [CrossRef] [PubMed]
- Laliberte, J.P.; Weisberg, A.S.; Moss, B. The membrane fusion step of vaccinia virus entry is cooperatively mediated by multiple viral proteins and host cell components. PLoS Pathog. 2011, 7, e1002446. [Google Scholar] [CrossRef] [PubMed]
- MacNeill, A.L.; Weishaar, K.M.; Seguin, B.; Powers, B.E. Safety of an oncolytic myxoma virus in dogs with soft tissue sarcoma. Viruses 2018, 10, 398. [Google Scholar] [CrossRef] [PubMed]
- Frey, S.E.; Newman, F.K.; Cruz, J.; Shelton, W.B.; Tennant, J.M.; Polach, T.; Rothman, A.L.; Kennedy, J.S.; Wolff, M.; Belshe, R.B.; et al. Dose-related effects of smallpox vaccine. N. Engl. J. Med. 2002, 346, 1275–1280. [Google Scholar] [CrossRef] [PubMed]
- Gentschev, I.; Ehrig, K.; Donat, U.; Hess, M.; Rudolph, S.; Chen, N.; Yu, Y.A.; Zhang, Q.; Bullerdiek, J.; Nolte, I.; et al. Significant growth inhibition of canine mammary carcinoma xenografts following treatment with oncolytic vaccinia virus glv-1h68. J. Oncol. 2010, 2010, 736907. [Google Scholar] [CrossRef] [PubMed]
- Gentschev, I.; Adelfinger, M.; Josupeit, R.; Rudolph, S.; Ehrig, K.; Donat, U.; Weibel, S.; Chen, N.G.; Yu, Y.A.; Zhang, Q.; et al. Preclinical evaluation of oncolytic vaccinia virus for therapy of canine soft tissue sarcoma. PLoS ONE 2012, 7, e37239. [Google Scholar] [CrossRef] [PubMed]
- Patil, S.S.; Gentschev, I.; Adelfinger, M.; Donat, U.; Hess, M.; Weibel, S.; Nolte, I.; Frentzen, A.; Szalay, A.A. Virotherapy of canine tumors with oncolytic vaccinia virus glv-1h109 expressing an anti-vegf single-chain antibody. PLoS ONE 2012, 7, e47472. [Google Scholar] [CrossRef] [PubMed]
- Gentschev, I.; Patil, S.S.; Adelfinger, M.; Weibel, S.; Geissinger, U.; Frentzen, A.; Chen, N.G.; Yu, Y.A.; Zhang, Q.; Ogilvie, G.; et al. Characterization and evaluation of a new oncolytic vaccinia virus strain livp6.1.1 for canine cancer therapy. Bioengineered 2013, 4, 84–89. [Google Scholar] [CrossRef] [PubMed]
- Adelfinger, M.; Bessler, S.; Frentzen, A.; Cecil, A.; Langbein-Laugwitz, J.; Gentschev, I.; Szalay, A.A. Preclinical testing oncolytic vaccinia virus strain glv-5b451 expressing an anti-vegf single-chain antibody for canine cancer therapy. Viruses 2015, 7, 4075–4092. [Google Scholar] [CrossRef] [PubMed]
- Zizzo, N.; Patruno, R.; Zito, F.A.; Di Summa, A.; Tinelli, A.; Troilo, S.; Misino, A.; Ruggieri, E.; Goffredo, V.; Gadaleta, C.D.; et al. Vascular endothelial growth factor concentrations from platelets correlate with tumor angiogenesis and grading in a spontaneous canine non-hodgkin lymphoma model. Leuk. Lymphoma 2010, 51, 291–296. [Google Scholar] [CrossRef] [PubMed]
- Kerr, P.J.; Ghedin, E.; DePasse, J.V.; Fitch, A.; Cattadori, I.M.; Hudson, P.J.; Tscharke, D.C.; Read, A.F.; Holmes, E.C. Evolutionary history and attenuation of myxoma virus on two continents. PLoS Pathog. 2012, 8, e1002950. [Google Scholar] [CrossRef] [PubMed]
- Woo, Y.; Kelly, K.J.; Stanford, M.M.; Galanis, C.; Chun, Y.S.; Fong, Y.; McFadden, G. Myxoma virus is oncolytic for human pancreatic adenocarcinoma cells. Ann. Surg. Oncol. 2008, 15, 2329–2335. [Google Scholar] [CrossRef] [PubMed]
- Correa, R.J.; Komar, M.; Tong, J.G.; Sivapragasam, M.; Rahman, M.M.; McFadden, G.; Dimattia, G.E.; Shepherd, T.G. Myxoma virus-mediated oncolysis of ascites-derived human ovarian cancer cells and spheroids is impacted by differential akt activity. Gynecol. Oncol. 2012, 125, 441–450. [Google Scholar] [CrossRef] [PubMed]
- Lun, X.; Yang, W.; Alain, T.; Shi, Z.Q.; Muzik, H.; Barrett, J.W.; McFadden, G.; Bell, J.; Hamilton, M.G.; Senger, D.L.; et al. Myxoma virus is a novel oncolytic virus with significant antitumor activity against experimental human gliomas. Cancer Res. 2005, 65, 9982–9990. [Google Scholar] [CrossRef] [PubMed]
- Kim, M.; Madlambayan, G.J.; Rahman, M.M.; Smallwood, S.E.; Meacham, A.M.; Hosaka, K.; Scott, E.W.; Cogle, C.R.; McFadden, G. Myxoma virus targets primary human leukemic stem and progenitor cells while sparing normal hematopoietic stem and progenitor cells. Leukemia 2009, 23, 2313–2317. [Google Scholar] [CrossRef] [PubMed]
- Wang, G.; Barrett, J.W.; Stanford, M.; Werden, S.J.; Johnston, J.B.; Gao, X.; Sun, M.; Cheng, J.Q.; McFadden, G. Infection of human cancer cells with myxoma virus requires akt activation via interaction with a viral ankyrin-repeat host range factor. Proc. Natl. Acad. Sci. USA. 2006, 103, 4640–4645. [Google Scholar] [CrossRef] [PubMed]
- Nathaniel, R.; MacNeill, A.L.; Wang, Y.X.; Turner, P.C.; Moyer, R.W. Cowpox virus crma, myxoma virus serp2 and baculovirus p35 are not functionally interchangeable caspase inhibitors in poxvirus infections. J. Gen. Virol. 2004, 85, 1267–1278. [Google Scholar] [CrossRef] [PubMed]
- Urbasic, A.S.; Hynes, S.; Somrak, A.; Contakos, S.; Rahman, M.M.; Liu, J.; MacNeill, A.L. Oncolysis of canine tumor cells by myxoma virus lacking the serp2 gene. Am. J. Vet. Res. 2012, 73, 1252–1261. [Google Scholar] [CrossRef] [PubMed]
- Dennis, M.M.; McSporran, K.D.; Bacon, N.J.; Schulman, F.Y.; Foster, R.A.; Powers, B.E. Prognostic factors for cutaneous and subcutaneous soft tissue sarcomas in dogs. Vet. Pathol. 2011, 48, 73–84. [Google Scholar] [CrossRef] [PubMed]
- Kent, M.S.; Collins, C.J.; Ye, F. Activation of the akt and mammalian target of rapamycin pathways and the inhibitory effects of rapamycin on those pathways in canine malignant melanoma cell lines. Am. J. Vet. Res. 2009, 70, 263–269. [Google Scholar] [CrossRef] [PubMed]
- Rodriguez, S.; Fadlalla, K.; Graham, T.; Tameru, B.; Fermin, C.D.; Samuel, T. Immunohistochemical evaluation of akt protein activation in canine mast cell tumours. J. Comp. Pathol. 2012, 147, 171–176. [Google Scholar] [CrossRef] [PubMed]
- Costers, S.; Lefebvre, D.J.; Delputte, P.L.; Nauwynck, H.J. Porcine reproductive and respiratory syndrome virus modulates apoptosis during replication in alveolar macrophages. Arch. Virol. 2008, 153, 1453–1465. [Google Scholar] [CrossRef] [PubMed]
- Chen, S.P.; Wu, J.L.; Su, Y.C.; Hong, J.R. Anti-bcl-2 family members, zfbcl-x(l) and zfmcl-1a, prevent cytochrome c release from cells undergoing betanodavirus-induced secondary necrotic cell death. Apoptosis 2007, 12, 1043–1060. [Google Scholar] [CrossRef] [PubMed]
- Teigler, J.E.; Phogat, S.; Franchini, G.; Hirsch, V.M.; Michael, N.L.; Barouch, D.H. The canarypox virus vector alvac induces distinct cytokine responses compared to the vaccinia virus-based vectors mva and nyvac in rhesus monkeys. J. Virol. 2014, 88, 1809–1814. [Google Scholar] [CrossRef] [PubMed]
- Jourdier, T.M.; Moste, C.; Bonnet, M.C.; Delisle, F.; Tafani, J.P.; Devauchelle, P.; Tartaglia, J.; Moingeon, P. Local immunotherapy of spontaneous feline fibrosarcomas using recombinant poxviruses expressing interleukin 2 (il2). Gene Ther. 2003, 10, 2126–2132. [Google Scholar] [CrossRef] [PubMed]
- Guo, Z.S.; Naik, A.; O‘Malley, M.E.; Popovic, P.; Demarco, R.; Hu, Y.; Yin, X.; Yang, S.; Zeh, H.J.; Moss, B.; et al. The enhanced tumor selectivity of an oncolytic vaccinia lacking the host range and antiapoptosis genes spi-1 and spi-2. Cancer Res. 2005, 65, 9991–9998. [Google Scholar] [CrossRef] [PubMed]
- Greiner, S.; Humrich, J.Y.; Thuman, P.; Sauter, B.; Schuler, G.; Jenne, L. The highly attenuated vaccinia virus strain modified virus ankara induces apoptosis in melanoma cells and allows bystander dendritic cells to generate a potent anti-tumoral immunity. Clin. Exp. Immunol. 2006, 146, 344–353. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Q.; Yu, Y.A.; Wang, E.; Chen, N.; Danner, R.L.; Munson, P.J.; Marincola, F.M.; Szalay, A.A. Eradication of solid human breast tumors in nude mice with an intravenously injected light-emitting oncolytic vaccinia virus. Cancer Res. 2007, 67, 10038–10046. [Google Scholar] [CrossRef] [PubMed]
- Gentschev, I.; Stritzker, J.; Hofmann, E.; Weibel, S.; Yu, Y.A.; Chen, N.; Zhang, Q.; Bullerdiek, J.; Nolte, I.; Szalay, A.A. Use of an oncolytic vaccinia virus for the treatment of canine breast cancer in nude mice: Preclinical development of a therapeutic agent. Cancer Gene Ther. 2009, 16, 320–328. [Google Scholar] [CrossRef] [PubMed]
- Patruno, R.; Arpaia, N.; Gadaleta, C.D.; Passantino, L.; Zizzo, N.; Misino, A.; Lucarelli, N.M.; Catino, A.; Valerio, P.; Ribatti, D.; et al. Vegf concentration from plasma-activated platelets rich correlates with microvascular density and grading in canine mast cell tumour spontaneous model. J. Cell. Mol. Med. 2009, 13, 555–561. [Google Scholar] [CrossRef] [PubMed]
- Rossmeisl, J.H.; Duncan, R.B.; Huckle, W.R.; Troy, G.C. Expression of vascular endothelial growth factor in tumors and plasma from dogs with primary intracranial neoplasms. Am. J. Vet. Res. 2007, 68, 1239–1245. [Google Scholar] [CrossRef] [PubMed]
- Restucci, B.; Papparella, S.; Maiolino, P.; De Vico, G. Expression of vascular endothelial growth factor in canine mammary tumors. Vet. Pathol. 2002, 39, 488–493. [Google Scholar] [CrossRef] [PubMed]
- Sobczynska-Rak, A.; Polkowska, I.; Silmanowicz, P. Elevated vascular endothelial growth factor (vegf) levels in the blood serum of dogs with malignant neoplasms of the oral cavity. Acta Vet. Hung. 2014, 62, 362–371. [Google Scholar] [CrossRef] [PubMed]
- Balboni, A.; Mollace, C.; Giunti, M.; Dondi, F.; Prosperi, S.; Battilani, M. Investigation of the presence of canine adenovirus (cadv) in owned dogs in northern italy. Res. Vet. Sci. 2014, 97, 631–636. [Google Scholar] [CrossRef] [PubMed]
- Roy, S. Isolation and characterization of adenoviruses persistently shed from the gastrointestinal tract of non-human primates. PLoS Pathog. 2009, 5, e1000503. [Google Scholar] [CrossRef] [PubMed]
- Ternovoi, V.V.; Le, L.P.; Belousova, N.; Smith, B.F.; Siegal, G.P.; Curiel, D.T. Productive replication of human adenovirus type 5 in canine cells. J. Virol. 2005, 79, 1308–1311. [Google Scholar] [CrossRef] [PubMed]
- Abou El Hassan, M.A.I.; van der Meulen-Muileman, I.; Abbas, S.; Kruyt, F.A.E. Conditionally replicating adenoviruses kill tumor cells via a basic apoptotic machinery-independent mechanism that resembles necrosis-like programmed cell death. J. Virol. 2004, 78, 12243–12251. [Google Scholar] [CrossRef] [PubMed]
- Hemminki, A.; Kanerva, A.; Kremer, E.J.; Bauerschmitz, G.J.; Smith, B.F.; Liu, B.; Wang, M.; Desmond, R.A.; Keriel, A.; Barnett, B.; et al. A canine conditionally replicating adenovirus for evaluating oncolytic virotherapy in a syngeneic animal model. Mol. Ther. 2003, 7, 163–173. [Google Scholar] [CrossRef]
- Smith, B.F.; Curiel, D.T.; Ternovoi, V.V.; Borovjagin, A.V.; Baker, H.J.; Cox, N.; Siegal, G.P. Administration of a conditionally replicative oncolytic canine adenovirus in normal dogs. Cancer Biother. Radiopharm. 2006, 21, 601–606. [Google Scholar] [CrossRef] [PubMed]
- Alcayaga-Miranda, F.; Cascallo, M.; Rojas, J.J.; Pastor, J.; Alemany, R. Osteosarcoma cells as carriers to allow antitumor activity of canine oncolytic adenovirus in the presence of neutralizing antibodies. Cancer Gene Ther. 2010, 17, 792–802. [Google Scholar] [CrossRef] [PubMed]
- Le, L.P.; Rivera, A.A.; Glasgow, J.N.; Ternovoi, V.V.; Wu, H.; Wang, M.; Smith, B.F.; Siegal, G.P.; Curiel, D.T. Infectivity enhancement for adenoviral transduction of canine osteosarcoma cells. Gene Ther. 2006, 13, 389–399. [Google Scholar] [CrossRef] [PubMed]
- Laborda, E.; Puig-Saus, C.; Rodriguez-Garcia, A.; Moreno, R.; Cascallo, M.; Pastor, J.; Alemany, R. A prb-responsive, rgd-modified, and hyaluronidase-armed canine oncolytic adenovirus for application in veterinary oncology. Mol. Ther. 2014, 22, 986–998. [Google Scholar] [CrossRef] [PubMed]
- Cejalvo, T.; Perise-Barrios, A.J.; Del Portillo, I.; Laborda, E.; Rodriguez-Milla, M.A.; Cubillo, I.; Vazquez, F.; Sardon, D.; Ramirez, M.; Alemany, R.; et al. Remission of spontaneous canine tumors after systemic cellular viroimmunotherapy. Cancer Res. 2018, 78, 4891–4901. [Google Scholar] [CrossRef] [PubMed]
- von Euler, H.; Sadeghi, A.; Carlsson, B.; Rivera, P.; Loskog, A.; Segall, T.; Korsgren, O.; Totterman, T.H. Efficient adenovector cd40 ligand immunotherapy of canine malignant melanoma. J. Immunother. 2008, 31, 377–384. [Google Scholar] [CrossRef] [PubMed]
- Smedley, R.C.; Spangler, W.L.; Esplin, D.G.; Kitchell, B.E.; Bergman, P.J.; Ho, H.Y.; Bergin, I.L.; Kiupel, M. Prognostic markers for canine melanocytic neoplasms: A comparative review of the literature and goals for future investigation. Vet. Pathol. 2011, 48, 54–72. [Google Scholar] [CrossRef] [PubMed]
- Westberg, S.; Sadeghi, A.; Svensson, E.; Segall, T.; Dimopoulou, M.; Korsgren, O.; Hemminki, A.; Loskog, A.S.; Totterman, T.H.; von Euler, H. Treatment efficacy and immune stimulation by adcd40l gene therapy of spontaneous canine malignant melanoma. J. Immunother. 2013, 36, 350–358. [Google Scholar] [CrossRef] [PubMed]


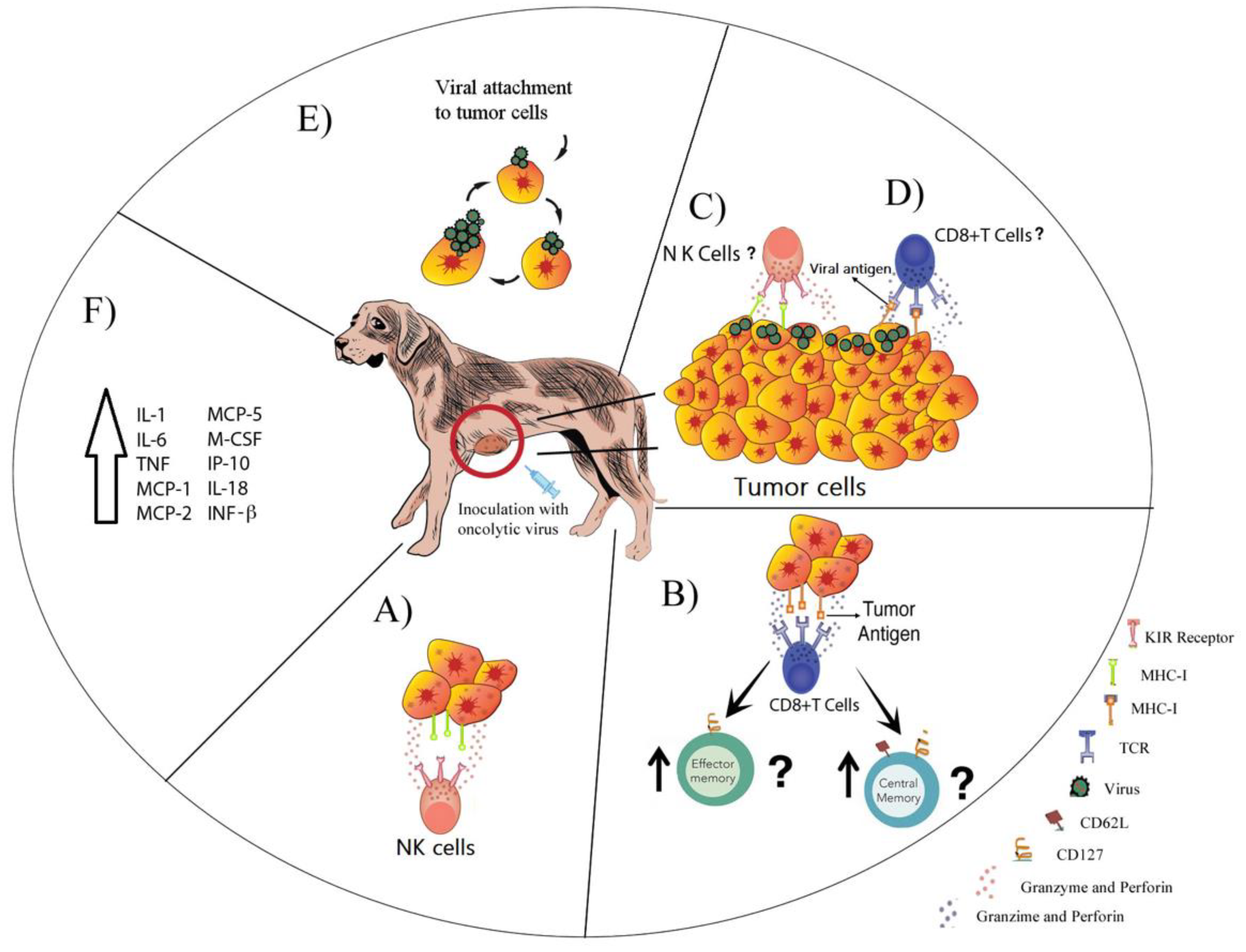{kind=link}
| Family | Genus | Virus |
|---|---|---|
| Paramyxoviridae | Morbilivirus | Measles, canine distemper virus,rinderpest, peste des petits ruminants |
| Avulavirus | Newcastle disease virus | |
| Respirovirus | Sendai | |
| Rubulavirus | Mumps * | |
| Henipavirus | Hendra | |
| Aquaparamyxovirus | Salmon aquaparamyxovirus | |
| Ferlavirus | Reptilian ferlavirus |
| Family | Genus | Species | Virus | Tumor Type Studied | References |
|---|---|---|---|---|---|
| Poxviridae | Leporipoxvirus | Myxoma virus | MYXV wild-type | In vitro in osteosarcoma, transitional cell carcinoma, hemangiosarcoma, mastocytoma, soft tissue sarcoma, stromal gastrointestinal tumor, perianal adenoma, mixed mammary tumor, and renal carcinoma | [136] |
| MYXVΔserp1 | |||||
| Avipoxvirus | Canarypox virus | ALVAC | Canine spontaneous melanoma | [137] | |
| Ortopoxvirus | Vaccinia Virus | GLV-1h68 | Canine mammary adenoma and carcinoma, both in vitro and in vivo in mice | [138,139] | |
| LIVP1.1.1 | Soft tissue sarcoma in vitro and in vivo in mice | [140] | |||
| GLV-1h109 | Soft tissue sarcoma in vitro and in vivo in mice, and in vitro in prostatic carcinoma | [141] | |||
| LIVP6.1.1 | Soft tissue sarcoma and prostatic carcinoma, both in vitro and in vivo in mice, and in vitro in melanoma and osteosarcoma | [142] | |||
| GLV-5b451 | In vitro in mammary carcinoma, mammary adenoma, and prostatic carcinoma, as well as in soft tissue sarcoma in vitro and in vivo in mice | [143] | |||
| NYVAC | Canine spontaneous melanoma | [137] |
| Genome | Family | Genus | Species | Strain Natural (N)/Modified (M) | Canine Model | References |
|---|---|---|---|---|---|---|
| ssRNA | Paramixoviridae | Morbilivirus | Measles | rMV-SLAMblind (M) | - Mammary gland tumor (Cell line CF33 nectine-4+) | [53] |
| Canine distemper virus (CDV) | Onderstepoort (N) | - Histiocytic sarcoma (cell line DH82) | [62] | |||
| FXNO (N), YSA-TC (N), and MD-77 (N) | - Histiocytic sarcoma (cell line CTT) | [63] | ||||
| pCDVeGFPΔN (M) | - Round-cell leukocytic neoplasia (cell line CCL-1390) - T-cell leukemia (cell line CLGL-90) - B-cell lymphoma (cell line 17-70) | [61] | ||||
| CDV (N) | - Canine lymphoma patient | [67] | ||||
| Avulavirus | Newcastle disease virus (NDV) | NDV-MLS (N) | - B-cell lymphoma (primary cells) - Healthy PBMC | [73] | ||
| Respirovirus | Sendai virus (SV) | SV (N) | - Canine spontaneous mastocytoma | [80] | ||
| ssRNA | Rabdoviridae | Vesiculovirus | Stomatitis vesicular virus (VSV) | VSV-IFNβ-NIS (M) | - Healthy dogs - Canine cancer patients | [86,88] |
| ssRNA | Togaviridae | Alfavirus | Semliki Forest virus | VA7 (N) | - OSA (cell lines Abrams and D17) - Healthy dogs | [101] |
| dsRNA | Reoviridae | Ortoreovirus | Reovirus | Dearing (Reolysin®) (N) | - Visceral MCT (cell lines VIMC and CoMS) - Cutaneous MCT (cell lines CM-MC and HRMC) - T-cell lymphoma (cell lines CL-1, UL-1, CLGL-90, Nody-1, Ema, and CLK) - OSA (cell lines D17, Gracie, Abrams, MacKinley, and Moresco) - Mammary gland tumor (cell lines CIP-p, CHM-m, CNM-m, CIP-m, CTB-p, CTB-m, and CHM-p) - Melanoma (cell lines CMGD2, CMeC1, CMeC2, KMeC, and LMeC). - Advanced canine cancer patients | [25,108,109,111] |
| ssDNA | Parvoviridae | Parvovirus | Canine parvovirus | Canine parvovirus (N) | - Fibroma (cell line A27) | [120] |
| dsDNA | Poxviridae | Leporipoxvirus | Mixoma virus (MYXV) | MYXV (N) | - OSA (Abrams and D-17 cell lines) - TCC (Bliley cells) - HSA (Den and Fitz cells) - MCT (2 primary cell lines) - HSA (1 primary cell line) - STS (1 primary cell line) - Stromal gastrointestinal tumor (primary cells) - Perianal adenoma (cell line) - Mixed mammary tumor (cell line) - Renal carcinoma (primary cells) - Healthy fibrous tissue (cell line) | [151] |
| MYXVΔserp1 (M) | ||||||
| Avipoxvirus | Canaripox virus | ALVAC (M) | - Canine spontaneous melanoma - Healthy dog | [137] | ||
| Ortopoxvirus | Vaccinia virus | GLV-1h68 (M) | - Mammary adenoma (cell line ZMTH3 and canine xenograft in mouse) - Mammary carcinoma (cell line MTH52c and canine xenograft in mouse) | [138,139] | ||
| LIVP1.1.1 (M) | - STS (cell line STSA-1 and canine xenograft in mouse) | [140] | ||||
| GLV-1h109 (M) | - STS (cell line STSA-1 and canine xenograft in mouse) - Prostatic carcinoma (cell line DTT08/40) | [141] | ||||
| LIVP6.1.1 (N) | - STS (cell line STSA-1 and xenograft in mouse) - Prostatic carcinoma (cell line DT08/40 and xenograft in mouse)- Melanoma (cell line CHAS) - OSA (cell line D17). | [142] | ||||
| GLV-5b451 (M) | - Mammary carcinoma (cell line MTH52c) - Mammary adenoma (cell line ZMTH3) - Prostatic carcinoma (cell line CT1258) - STS (cell line STSA-1 and xenograft in mouse) | [143] | ||||
| NYVAC (M) | - Canine melanoma patients - Healthy dog | [137] | ||||
| dsDNA | Adenoviridae | Mastadenovirus | Canine adenovirus (CAV) | OC-CAVE1 (M) | - OSA (cell lines D22, D17, and CF11, and xenograft in mouse) - Healthy dogs | [172,173,174,175] |
| ICOCAV17 | - OSA (cell lines Abrams and D17) - Melanoma (cell lines CML1 and 17CM98) - Canine cancer patients | [176,177] | ||||
| Human adenovirus (Ad5) | Ad5CMVGFP (M) | - OSA (primary cells) | [170] | |||
| AdCD40L (M) | - Canine malignant melanoma patients | [178,179] |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sánchez, D.; Cesarman-Maus, G.; Amador-Molina, A.; Lizano, M. Oncolytic Viruses for Canine Cancer Treatment. Cancers 2018, 10, 404. https://doi.org/10.3390/cancers10110404
Sánchez D, Cesarman-Maus G, Amador-Molina A, Lizano M. Oncolytic Viruses for Canine Cancer Treatment. Cancers. 2018; 10(11):404. https://doi.org/10.3390/cancers10110404
Chicago/Turabian StyleSánchez, Diana, Gabriela Cesarman-Maus, Alfredo Amador-Molina, and Marcela Lizano. 2018. "Oncolytic Viruses for Canine Cancer Treatment" Cancers 10, no. 11: 404. https://doi.org/10.3390/cancers10110404
APA StyleSánchez, D., Cesarman-Maus, G., Amador-Molina, A., & Lizano, M. (2018). Oncolytic Viruses for Canine Cancer Treatment. Cancers, 10(11), 404. https://doi.org/10.3390/cancers10110404