The Intrinsic and Extrinsic Implications of RANKL/RANK Signaling in Osteosarcoma: From Tumor Initiation to Lung Metastases



 ,
,  and
and 
Abstract
1. Introduction
2. Results
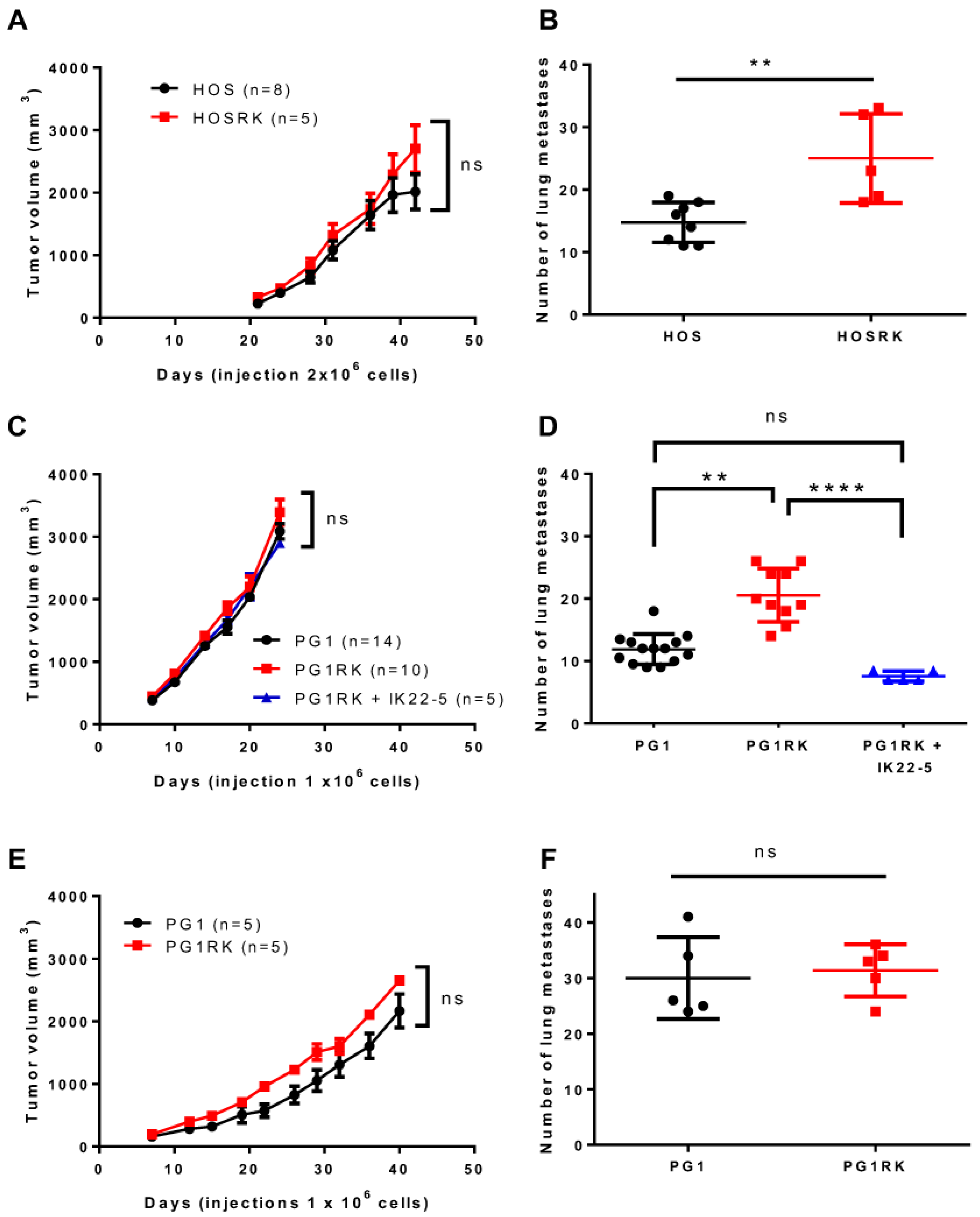
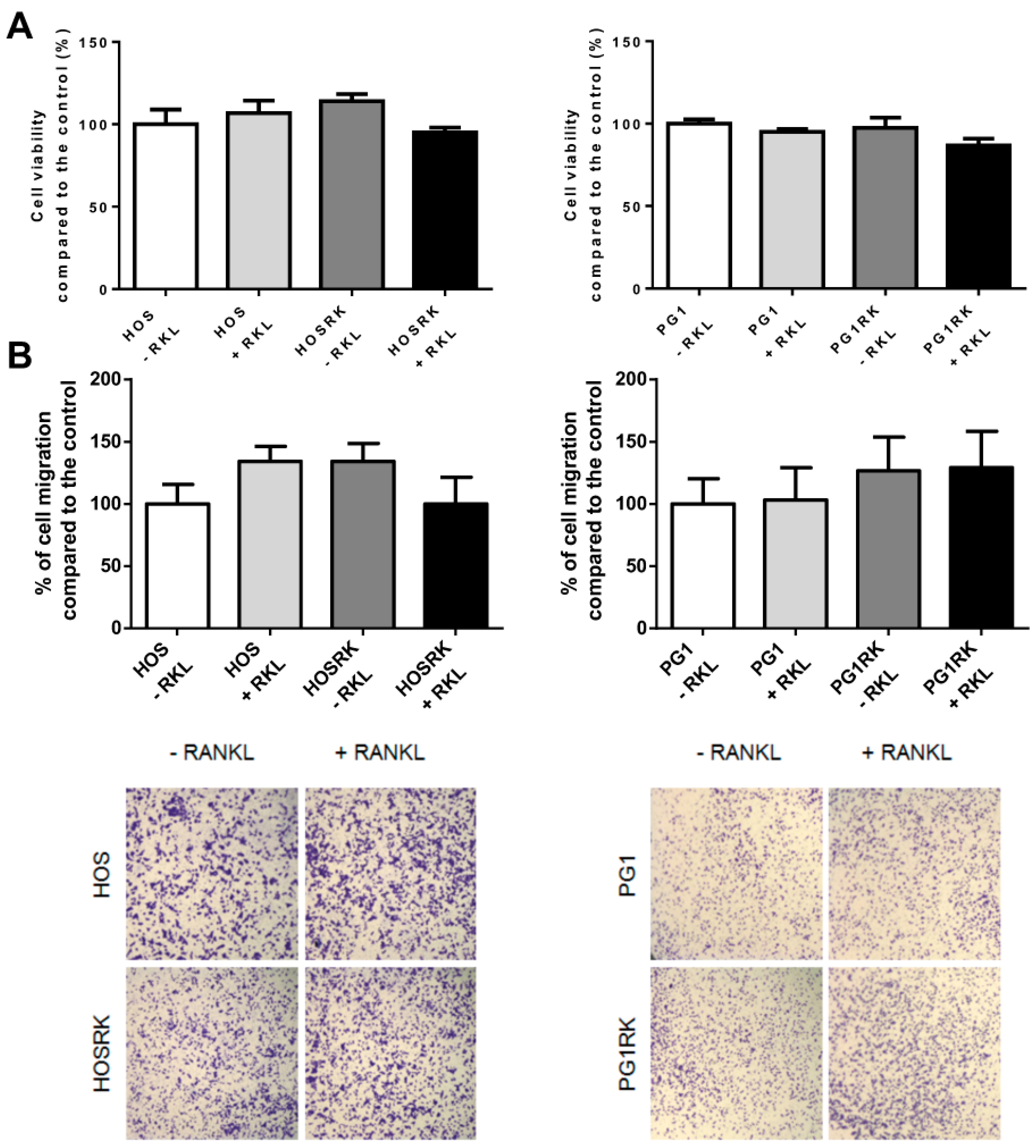
2.1. Intrinsic RANK Expression by Osteosarcoma Cells Does Not Impact Cell Proliferation or Tumor Growth
2.2. RANK Expression by Osteosarcoma Cells Increases the Number of Lung Metastases Rankl-Dependently in Nude Mice
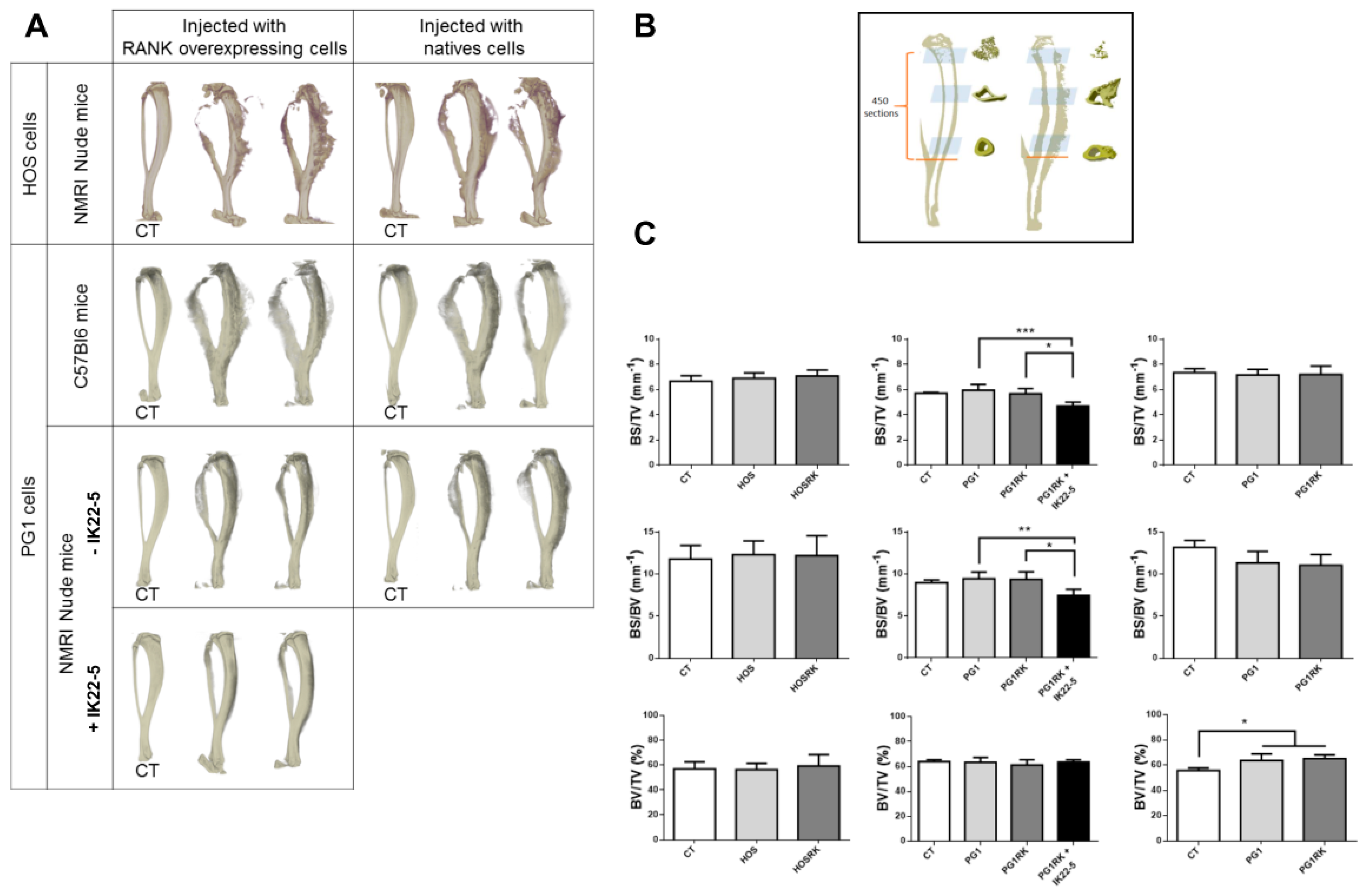
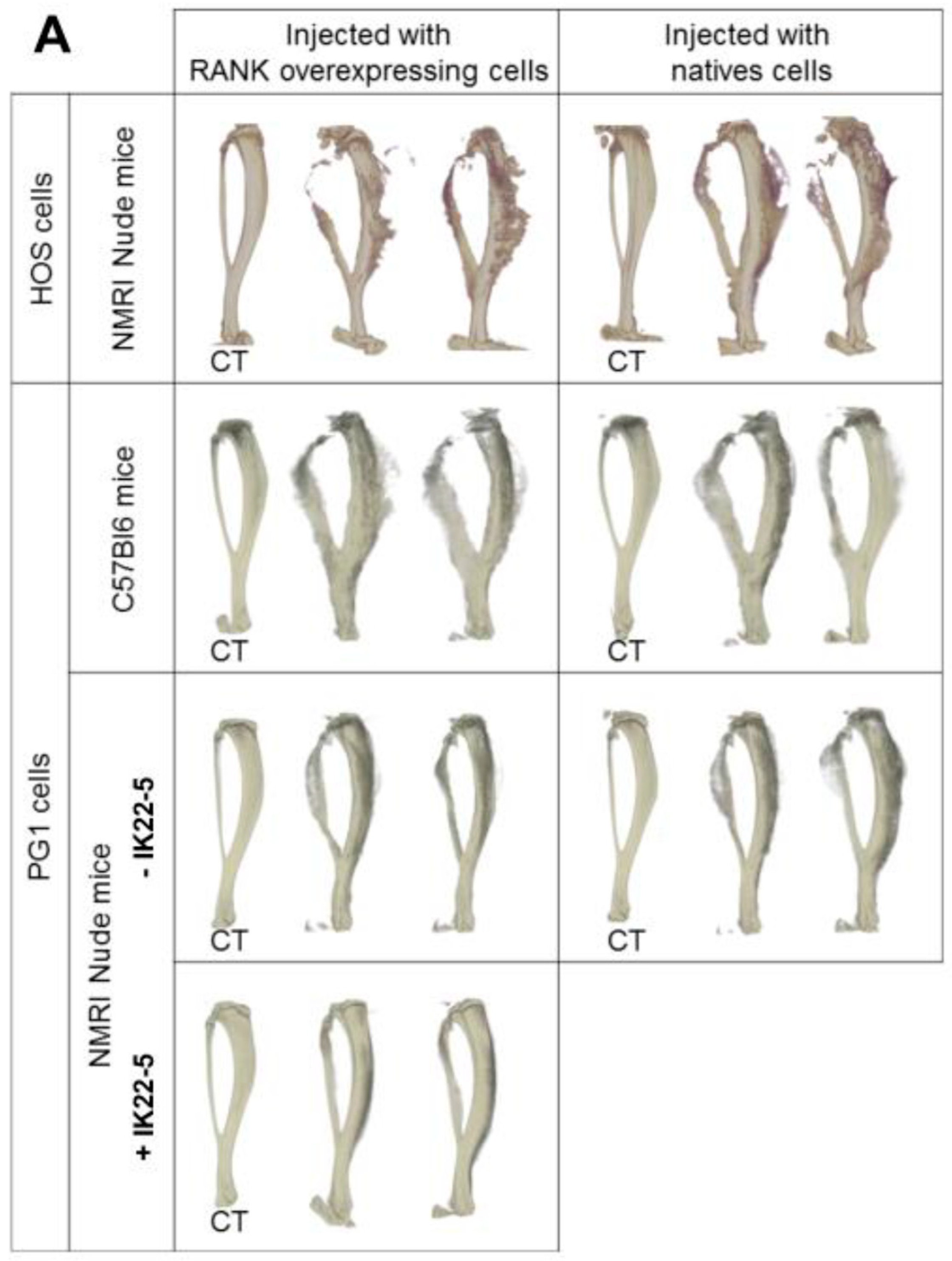
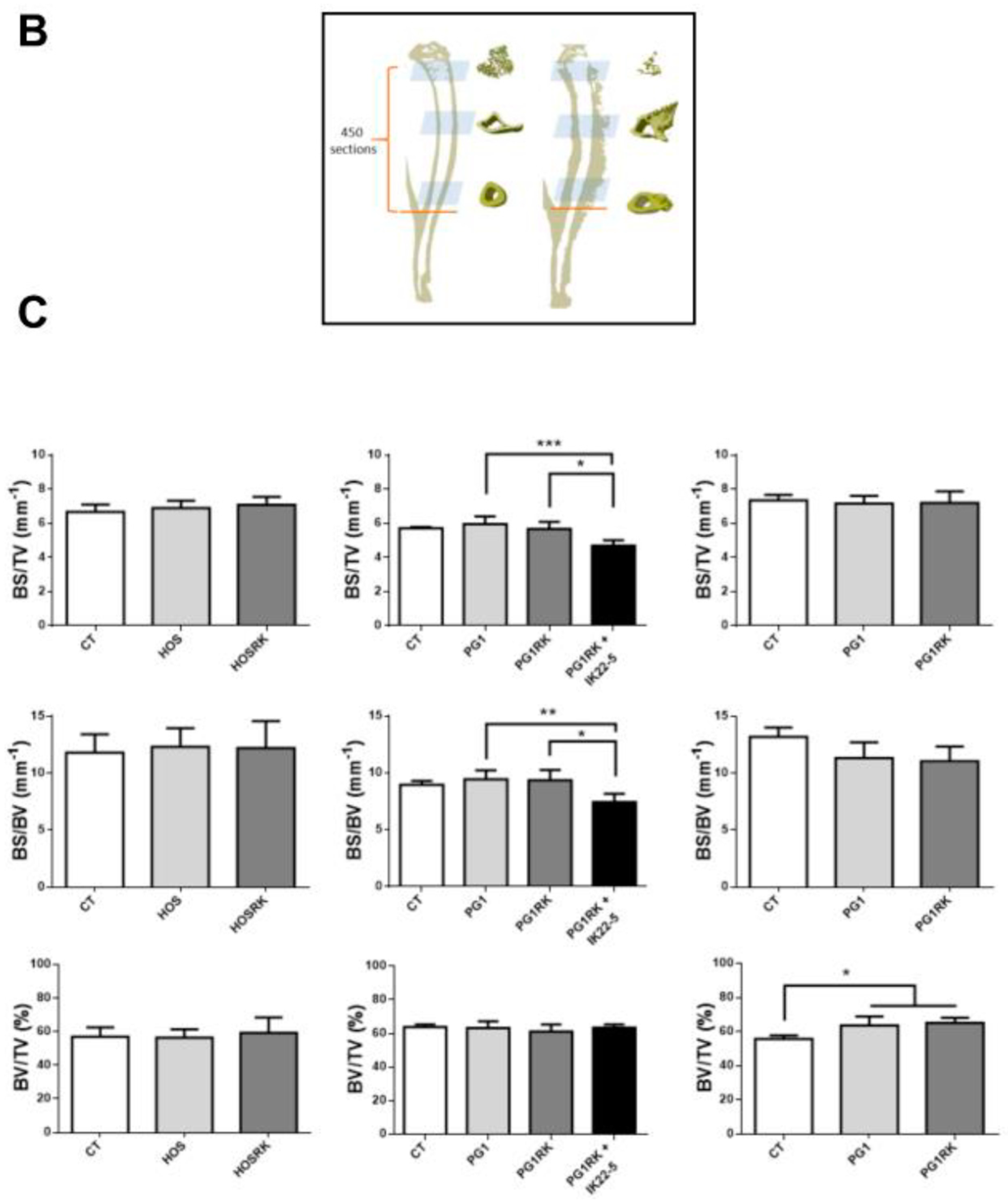
2.3. RANK Expression by Osteosarcoma Cells Has No Impact on Tumor-Associated Bone Destruction
2.4. Consequences of Adding RANKL to RANK Over-Expressing HOS Cells, with Regard to Expression Levels of the Genes Encoding Factors Implicated in the Different Cellular Processes
2.5. Genetically-Achieved RANKL Depletion in T Cells Has No Impact on Tumor Growth or Metastatic Spread
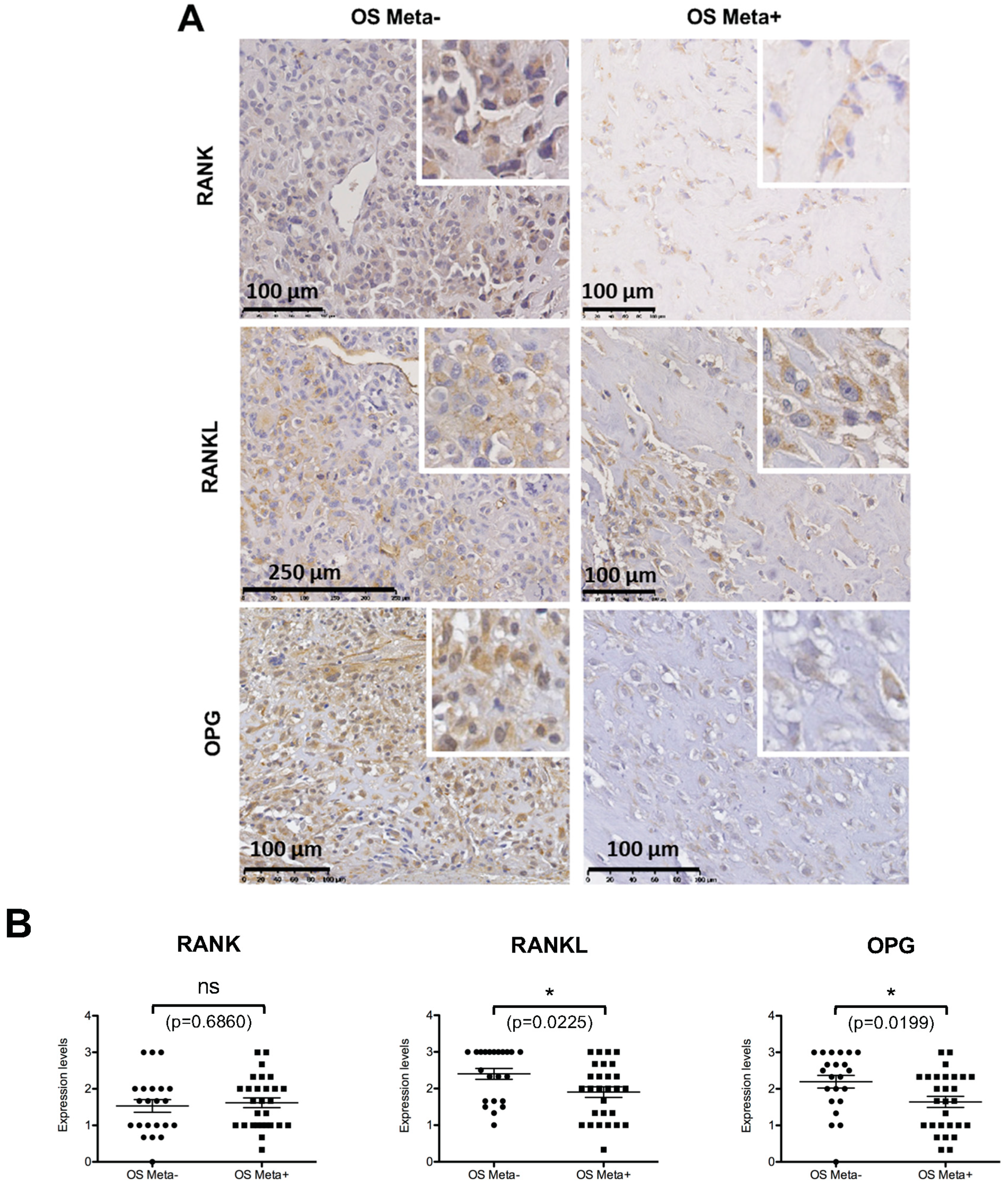
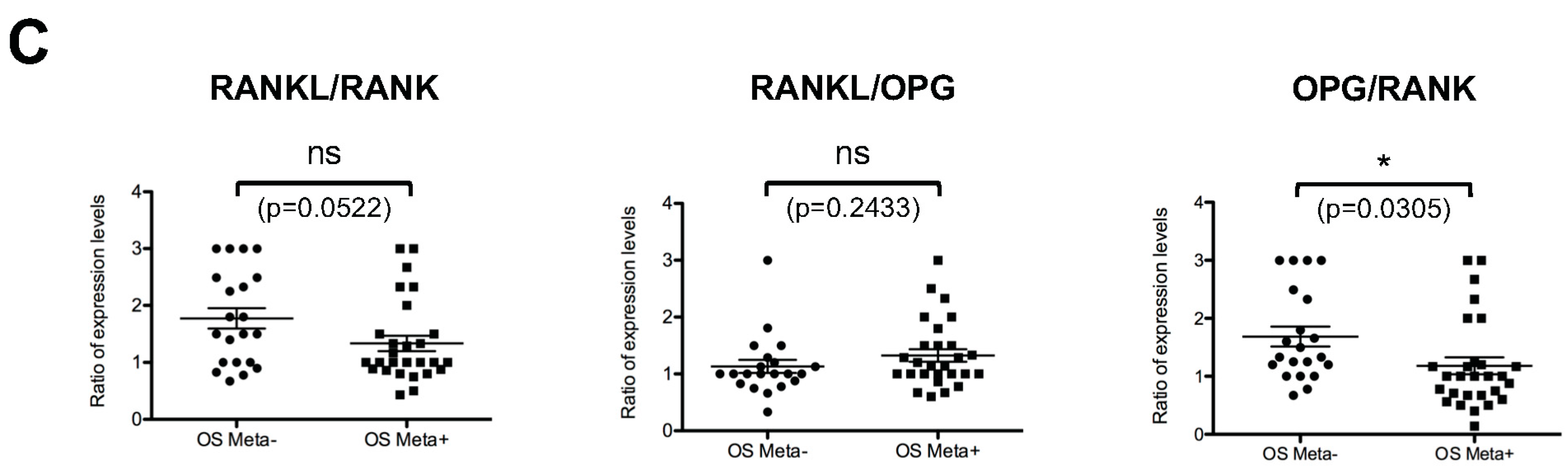
2.6. Comparative Analysis of RANK, RANKL and OPG Expression in Biopsies of Patients with or without Metastases at the Time of Diagnosis
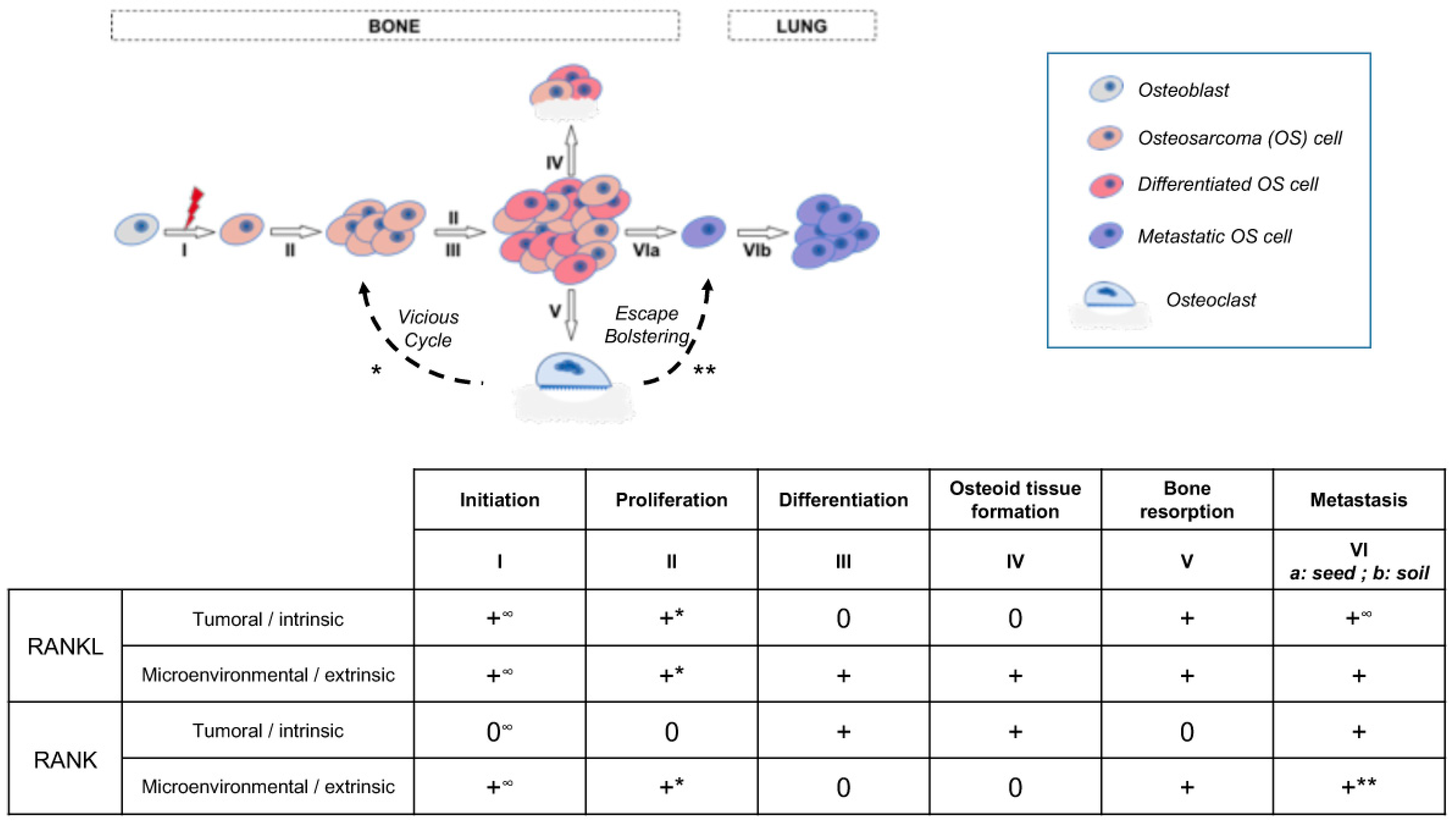
3. Discussion
4. Materials and Methods
4.1. Cell Culture
4.2. Lentivirus Production and Osteosarcoma Cell Transduction
4.3. Validation of Functional Rank Over-Expression in Osteosarcoma Cells
4.4. Cell Viability Assay
4.5. RNA Isolation, Reverse Transcription and Quantitative PCR
4.6. Migration Assay
4.7. Mouse Models of Osteosarcoma
4.8. IK22-5 RANKL Blocking Antibody Injections
4.9. Micro-CT Analysis
4.10. Immunohistochemistry
4.11. Tissue Microarrays
4.12. Statistics
4.13. Western Blot Analysis, Zymography Assay and Flow Cytometry
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Heymann, D. Preface. In Bone Cancer; Academic Press: San Diego, CA, USA, 2010; p. ix. ISBN 978-0-12-374895-9. [Google Scholar]
- Bielack, S.S.; Hecker-Nolting, S.; Blattmann, C.; Kager, L. Advances in the management of osteosarcoma. F1000Research 2016, 5, 2767. [Google Scholar] [CrossRef] [PubMed]
- Lindsey, B.A.; Markel, J.E.; Kleinerman, E.S. Osteosarcoma Overview. Rheumatol. Ther. 2017, 4, 25–43. [Google Scholar] [CrossRef] [PubMed]
- Nataraj, V.; Rastogi, S.; Khan, S.A.; Sharma, M.C.; Agarwala, S.; Vishnubhatla, S.; Bakhshi, S. Prognosticating metastatic osteosarcoma treated with uniform chemotherapy protocol without high dose methotrexate and delayed metastasectomy: A single center experience of 102 patients. Clin. Transl. Oncol. 2016, 18, 937–944. [Google Scholar] [CrossRef] [PubMed]
- Allison, D.C.; Carney, S.C.; Ahlmann, E.R.; Hendifar, A.; Chawla, S.; Fedenko, A.; Angeles, C.; Menendez, L.R. A Meta-Analysis of Osteosarcoma Outcomes in the Modern Medical Era. Sarcoma 2012, 2012, 704872. [Google Scholar] [CrossRef] [PubMed]
- Heymann, M.-F.; Brown, H.K.; Heymann, D. Drugs in early clinical development for the treatment of osteosarcoma. Expert Opin. Investig. Drugs 2016, 25, 1265–1280. [Google Scholar] [CrossRef] [PubMed]
- Kansara, M.; Teng, M.W.; Smyth, M.J.; Thomas, D.M. Translational biology of osteosarcoma. Nat. Rev. Cancer 2014, 14, 722–735. [Google Scholar] [CrossRef] [PubMed]
- Kenkre, J.S.; Bassett, J. The bone remodelling cycle. Ann. Clin. Biochem. 2018, 4563218759371. [Google Scholar] [CrossRef] [PubMed]
- Walsh, M.C.; Choi, Y. Biology of the RANKL-RANK-OPG System in Immunity, Bone, and Beyond. Front. Immunol. 2014, 5, 511. [Google Scholar] [CrossRef] [PubMed]
- Xiong, J.; Onal, M.; Jilka, R.L.; Weinstein, R.S.; Manolagas, S.C.; O’Brien, C.A. Matrix-embedded cells control osteoclast formation. Nat. Med. 2011, 17, 1235–1241. [Google Scholar] [CrossRef] [PubMed]
- Rao, S.; Cronin, S.J.F.; Sigl, V.; Penninger, J.M. RANKL and RANK: From Mammalian Physiology to Cancer Treatment. Trends Cell Biol. 2018, 28, 213–223. [Google Scholar] [CrossRef] [PubMed]
- Theoleyre, S.; Wittrant, Y.; Tat, S.K.; Fortun, Y.; Redini, F.; Heymann, D. The molecular triad OPG/RANK/RANKL: Involvement in the orchestration of pathophysiological bone remodeling. Cytokine Growth Factor Rev. 2004, 15, 457–475. [Google Scholar] [CrossRef] [PubMed]
- Santini, D.; Perrone, G.; Roato, I.; Godio, L.; Pantano, F.; Grasso, D.; Russo, A.; Vincenzi, B.; Fratto, M.E.; Sabbatini, R.; et al. Expression pattern of receptor activator of NFκB (RANK) in a series of primary solid tumors and related bone metastases. J. Cell. Physiol. 2011, 226, 780–784. [Google Scholar] [CrossRef] [PubMed]
- Mori, K.; Le Goff, B.; Charrier, C.; Battaglia, S.; Heymann, D.; Rédini, F. DU145 human prostate cancer cells express functional receptor activator of NFκB: New insights in the prostate cancer bone metastasis process. Bone 2007, 40, 981–990. [Google Scholar] [CrossRef] [PubMed]
- Sisay, M.; Mengistu, G.; Edessa, D. The RANK/RANKL/OPG system in tumorigenesis and metastasis of cancer stem cell: Potential targets for anticancer therapy. OncoTargets Ther. 2017, 10, 3801–3810. [Google Scholar] [CrossRef] [PubMed]
- Sigl, V.; Jones, L.P.; Penninger, J.M. RANKL/RANK: From bone loss to the prevention of breast cancer. Open Biol. 2016, 6, 160230. [Google Scholar] [CrossRef] [PubMed]
- Lamoureux, F.; Richard, P.; Wittrant, Y.; Battaglia, S.; Pilet, P.; Trichet, V.; Blanchard, F.; Gouin, F.; Pitard, B.; Heymann, D.; et al. Therapeutic relevance of osteoprotegerin gene therapy in osteosarcoma: Blockade of the vicious cycle between tumor cell proliferation and bone resorption. Cancer Res. 2007, 67, 7308–7318. [Google Scholar] [CrossRef] [PubMed]
- Grimaud, E.; Soubigou, L.; Couillaud, S.; Coipeau, P.; Moreau, A.; Passuti, N.; Gouin, F.; Redini, F.; Heymann, D. Receptor activator of nuclear factor kappaB ligand (RANKL)/osteoprotegerin (OPG) ratio is increased in severe osteolysis. Am. J. Pathol. 2003, 163, 2021–2031. [Google Scholar] [CrossRef]
- Guise, T.A. The vicious cycle of bone metastases. J. Musculoskelet. Neuronal Interact. 2002, 2, 570–572. [Google Scholar] [PubMed]
- Beristain, A.G.; Narala, S.R.; Di Grappa, M.A.; Khokha, R. Homotypic RANK signaling differentially regulates proliferation, motility and cell survival in osteosarcoma and mammary epithelial cells. J. Cell Sci. 2012, 125, 943–955. [Google Scholar] [CrossRef] [PubMed]
- Castaneda, B.; Simon, Y.; Ferbus, D.; Robert, B.; Chesneau, J.; Mueller, C.; Berdal, A.; Lézot, F. Role of RANKL (TNFSF11)-dependent osteopetrosis in the dental phenotype of Msx2 null mutant mice. PLoS ONE 2013, 8, e80054. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.; Di Grappa, M.A.; Molyneux, S.D.; McKee, T.D.; Waterhouse, P.; Penninger, J.M.; Khokha, R. RANKL blockade prevents and treats aggressive osteosarcomas. Sci. Transl. Med. 2015, 7, 317ra197. [Google Scholar] [CrossRef] [PubMed]
- Renema, N.; Navet, B.; Heymann, M.-F.; Lezot, F.; Heymann, D. RANK-RANKL signalling in cancer. Biosci. Rep. 2016, 36. [Google Scholar] [CrossRef] [PubMed]
- Bago-Horvath, Z.; Schmid, K.; Rössler, F.; Nagy-Bojarszky, K.; Funovics, P.; Sulzbacher, I. Impact of RANK signalling on survival and chemotherapy response in osteosarcoma. Pathology 2014, 46, 411–415. [Google Scholar] [CrossRef] [PubMed]
- Mori, K.; Le Goff, B.; Berreur, M.; Riet, A.; Moreau, A.; Blanchard, F.; Chevalier, C.; Guisle-Marsollier, I.; Léger, J.; Guicheux, J.; et al. Human osteosarcoma cells express functional receptor activator of nuclear factor-kappa B. J. Pathol. 2007, 211, 555–562. [Google Scholar] [CrossRef] [PubMed]
- Wittrant, Y.; Lamoureux, F.; Mori, K.; Riet, A.; Kamijo, A.; Heymann, D.; Redini, F. RANKL directly induces bone morphogenetic protein-2 expression in RANK-expressing POS-1 osteosarcoma cells. Int. J. Oncol. 2006, 28, 261–269. [Google Scholar] [CrossRef] [PubMed]
- Trieb, K.; Windhager, R. Receptor activator of nuclear factor κB expression is a prognostic factor in human osteosarcoma. Oncol. Lett. 2015, 10, 1813–1815. [Google Scholar] [CrossRef] [PubMed]
- Safa, A.R. c-FLIP, a master anti-apoptotic regulator. Exp. Oncol. 2012, 34, 176–184. [Google Scholar] [PubMed]
- Stine, Z.E.; Walton, Z.E.; Altman, B.J.; Hsieh, A.L.; Dang, C.V. MYC, Metabolism, and Cancer. Cancer Discov. 2015, 5, 1024–1039. [Google Scholar] [CrossRef] [PubMed]
- Gonzalez-Suarez, E.; Jacob, A.P.; Jones, J.; Miller, R.; Roudier-Meyer, M.P.; Erwert, R.; Pinkas, J.; Branstetter, D.; Dougall, W.C. RANK ligand mediates progestin-induced mammary epithelial proliferation and carcinogenesis. Nature 2010, 468, 103–107. [Google Scholar] [CrossRef] [PubMed]
- Lamoureux, F.; Baud’huin, M.; Rodriguez Calleja, L.; Jacques, C.; Berreur, M.; Rédini, F.; Lecanda, F.; Bradner, J.E.; Heymann, D.; Ory, B. Selective inhibition of BET bromodomain epigenetic signalling interferes with the bone-associated tumour vicious cycle. Nat. Commun. 2014, 5, 3511. [Google Scholar] [CrossRef] [PubMed]
- Hanada, R.; Hanada, T.; Penninger, J.M. Physiology and pathophysiology of the RANKL/RANK system. Biol. Chem. 2010, 391, 1365–1370. [Google Scholar] [CrossRef] [PubMed]
- Duheron, V.; Hess, E.; Duval, M.; Decossas, M.; Castaneda, B.; Klöpper, J.E.; Amoasii, L.; Barbaroux, J.-B.; Williams, I.R.; Yagita, H.; et al. Receptor activator of NF-kappaB (RANK) stimulates the proliferation of epithelial cells of the epidermo-pilosebaceous unit. Proc. Natl. Acad. Sci. USA 2011, 108, 5342–5347. [Google Scholar] [CrossRef] [PubMed]
- Sigl, V.; Penninger, J.M. RANKL/RANK-from bone physiology to breast cancer. Cytokine Growth Factor Rev. 2014, 25, 205–214. [Google Scholar] [CrossRef] [PubMed]
- Thomas, D.M.; Skubitz, K.M. Giant cell tumour of bone. Curr. Opin. Oncol. 2009, 21, 338–344. [Google Scholar] [CrossRef] [PubMed]
- Ren, T.; Piperdi, S.; Koirala, P.; Park, A.; Zhang, W.; Ivenitsky, D.; Zhang, Y.; Villanueva-Siles, E.; Hawkins, D.S.; Roth, M.; et al. CD49b inhibits osteogenic differentiation and plays an important role in osteosarcoma progression. Oncotarget 2017, 8, 87848–87859. [Google Scholar] [CrossRef] [PubMed]
- Canon, J.; Bryant, R.; Roudier, M.; Branstetter, D.G.; Dougall, W.C. RANKL inhibition combined with tamoxifen treatment increases anti-tumor efficacy and prevents tumor-induced bone destruction in an estrogen receptor-positive breast cancer bone metastasis model. Breast Cancer Res. Treat. 2012, 135, 771–780. [Google Scholar] [CrossRef] [PubMed]
- Christoph, F.; König, F.; Lebentrau, S.; Jandrig, B.; Krause, H.; Strenziok, R.; Schostak, M. RANKL/RANK/OPG cytokine receptor system: mRNA expression pattern in BPH, primary and metastatic prostate cancer disease. World J. Urol. 2018, 36, 187–192. [Google Scholar] [CrossRef] [PubMed]
- Casimiro, S.; Mohammad, K.S.; Pires, R.; Tato-Costa, J.; Alho, I.; Teixeira, R.; Carvalho, A.; Ribeiro, S.; Lipton, A.; Guise, T.A.; et al. RANKL/RANK/MMP-1 molecular triad contributes to the metastatic phenotype of breast and prostate cancer cells in vitro. PLoS ONE 2013, 8, e63153. [Google Scholar] [CrossRef] [PubMed]
- Thirumala, R.; Ramaswamy, M.; Chawla, S. Diagnosis and management of infectious complications in critically ill patients with cancer. Crit. Care Clin. 2010, 26, 59–91. [Google Scholar] [CrossRef] [PubMed]
- Avnet, S.; Longhi, A.; Salerno, M.; Halleen, J.M.; Perut, F.; Granchi, D.; Ferrari, S.; Bertoni, F.; Giunti, A.; Baldini, N. Increased osteoclast activity is associated with aggressiveness of osteosarcoma. Int. J. Oncol. 2008, 33, 1231–1238. [Google Scholar] [CrossRef] [PubMed]
- Luo, J.; Yang, Z.; Ma, Y.; Yue, Z.; Lin, H.; Qu, G.; Huang, J.; Dai, W.; Li, C.; Zheng, C.; et al. LGR4 is a receptor for RANKL and negatively regulates osteoclast differentiation and bone resorption. Nat. Med. 2016, 22, 539–546. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.; Wei, W.; Guo, C.-A.; Han, N.; Pan, J.; Fei, T.; Yan, Z. Stat3 Upregulates Leucine-Rich Repeat-Containing G Protein-Coupled Receptor 4 Expression in Osteosarcoma Cells. BioMed Res. Int. 2013, 2013, 310691. [Google Scholar] [CrossRef] [PubMed]
- Luo, W.; Tan, P.; Rodriguez, M.; He, L.; Tan, K.; Zeng, L.; Siwko, S.; Liu, M. Leucine-rich repeat-containing G protein-coupled receptor 4 (Lgr4) is necessary for prostate cancer metastasis via epithelial-mesenchymal transition. J. Biol. Chem. 2017, 292, 15525–15537. [Google Scholar] [CrossRef] [PubMed]
- Joliat, M.J.; Umeda, S.; Lyons, B.L.; Lynes, M.A.; Shultz, L.D. Establishment and characterization of a new osteogenic cell line (MOS-J) from a spontaneous C57BL/6J mouse osteosarcoma. In Vivo Athens Greece 2002, 16, 223–228. [Google Scholar]
- Dull, T.; Zufferey, R.; Kelly, M.; Mandel, R.J.; Nguyen, M.; Trono, D.; Naldini, L. A third-generation lentivirus vector with a conditional packaging system. J. Virol. 1998, 72, 8463–8471. [Google Scholar] [PubMed]
- Lézot, F.; Chesneau, J.; Navet, B.; Gobin, B.; Amiaud, J.; Choi, Y.; Yagita, H.; Castaneda, B.; Berdal, A.; Mueller, C.G.; et al. Skeletal consequences of RANKL-blocking antibody (IK22-5) injections during growth: Mouse strain disparities and synergic effect with zoledronic acid. Bone 2015, 73, 51–59. [Google Scholar] [CrossRef] [PubMed]
- Kamijo, S.; Nakajima, A.; Ikeda, K.; Aoki, K.; Ohya, K.; Akiba, H.; Yagita, H.; Okumura, K. Amelioration of bone loss in collagen-induced arthritis by neutralizing anti-RANKL monoclonal antibody. Biochem. Biophys. Res. Commun. 2006, 347, 124–132. [Google Scholar] [CrossRef] [PubMed]
- Dumars, C.; Ngyuen, J.-M.; Gaultier, A.; Lanel, R.; Corradini, N.; Gouin, F.; Heymann, D.; Heymann, M.-F. Dysregulation of macrophage polarization is associated with the metastatic process in osteosarcoma. Oncotarget 2016, 7, 78343–78354. [Google Scholar] [CrossRef] [PubMed]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Genes | Fold Change HOS + RKL vs. HOS | Fold Change HOSRK + RKL vs. HOSRK |
|---|---|---|
| RANK | 1.10 ± 0.18 | 1.58 ± 0.14 |
| RANKL | 4.44 ± 1.99 | 77.94 ± 28.37 |
| OPG | 1.00 ± 0.10 | 1.09 ± 0.12 |
| Apoptosis and proliferation | ||
| Bax | 1.18 ± 0.12 | 1.21 ± 0.11 |
| Bcl2 | −1.04 ± 0.23 | 1.28 ± 0.15 |
| p21 | 1.18 ± 0.21 | 4.22 ± 0.57 |
| p53 | 1.05 ± 0.08 | −1.23 ± 0.12 |
| Differentiation | ||
| RUNX2 | 1.01 ± 0.07 | −1.28 ± 0.54 |
| SOX9 | −1.17 ± 0.07 | −2.14 ± 0.05 |
| COLL1a1 | −1.09 ± 0.07 | −6.00 ± 0.05 |
| OCN | 1.70 ± 0.14 | 1.41 ± 0.15 |
| OPN | 1.58 ± 0.12 | 2.81 ± 0.42 |
| BSP | −1.06 ± 0.32 | 50.25 ± 20.44 |
| Migration | ||
| MMP2 | −1.31 ± 0.16 | −1.43 ± 0.06 |
| MMP9 | −1.03 ± 0.14 | 7.98 ± 0.97 |
| MMP13 | 1.24 ± 0.30 | 11.73 ± 1.41 |
| TIMP1 | 1.07 ± 0.08 | 1.07 ± 0.12 |
| TIMP2 | −1.05 ± 0.13 | −1.38 ± 0.11 |
| Oncogenes | ||
| c-FLIP | −1.23 ± 0.07 | 2.59 ± 0.44 |
| c-MET | −1.13 ± 0.10 | −1.11 ± 0.25 |
| c-MYC | −1.17 ± 0.07 | 3.58 ± 0.31 |
| Others | ||
| NFATC1 | 1.20 ± 0.14 | 7.13 ± 3.48 |
| NFkB | 1.05 ± 0.22 | 2.07 ± 0.53 |
| BMP2 | 1.21 ± 0.35 | 3.24 ± 0.51 |
| TGFb1 | 1.01 ± 0.07 | 1.13 ± 0.20 |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Navet, B.; Ando, K.; Vargas-Franco, J.W.; Brion, R.; Amiaud, J.; Mori, K.; Yagita, H.; Mueller, C.G.; Verrecchia, F.; Dumars, C.; et al. The Intrinsic and Extrinsic Implications of RANKL/RANK Signaling in Osteosarcoma: From Tumor Initiation to Lung Metastases. Cancers 2018, 10, 398. https://doi.org/10.3390/cancers10110398
Navet B, Ando K, Vargas-Franco JW, Brion R, Amiaud J, Mori K, Yagita H, Mueller CG, Verrecchia F, Dumars C, et al. The Intrinsic and Extrinsic Implications of RANKL/RANK Signaling in Osteosarcoma: From Tumor Initiation to Lung Metastases. Cancers. 2018; 10(11):398. https://doi.org/10.3390/cancers10110398
Chicago/Turabian StyleNavet, Benjamin, Kosei Ando, Jorge William Vargas-Franco, Régis Brion, Jérome Amiaud, Kanji Mori, Hideo Yagita, Christopher G. Mueller, Franck Verrecchia, Clotilde Dumars, and et al. 2018. "The Intrinsic and Extrinsic Implications of RANKL/RANK Signaling in Osteosarcoma: From Tumor Initiation to Lung Metastases" Cancers 10, no. 11: 398. https://doi.org/10.3390/cancers10110398
APA StyleNavet, B., Ando, K., Vargas-Franco, J. W., Brion, R., Amiaud, J., Mori, K., Yagita, H., Mueller, C. G., Verrecchia, F., Dumars, C., Heymann, M.-F., Heymann, D., & Lézot, F. (2018). The Intrinsic and Extrinsic Implications of RANKL/RANK Signaling in Osteosarcoma: From Tumor Initiation to Lung Metastases. Cancers, 10(11), 398. https://doi.org/10.3390/cancers10110398