
A Novel Microfluidic Device for the Neutrophil Functional Phenotype Analysis: Effects of Glucose and Its Derivatives AGEs


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Materials and Methods
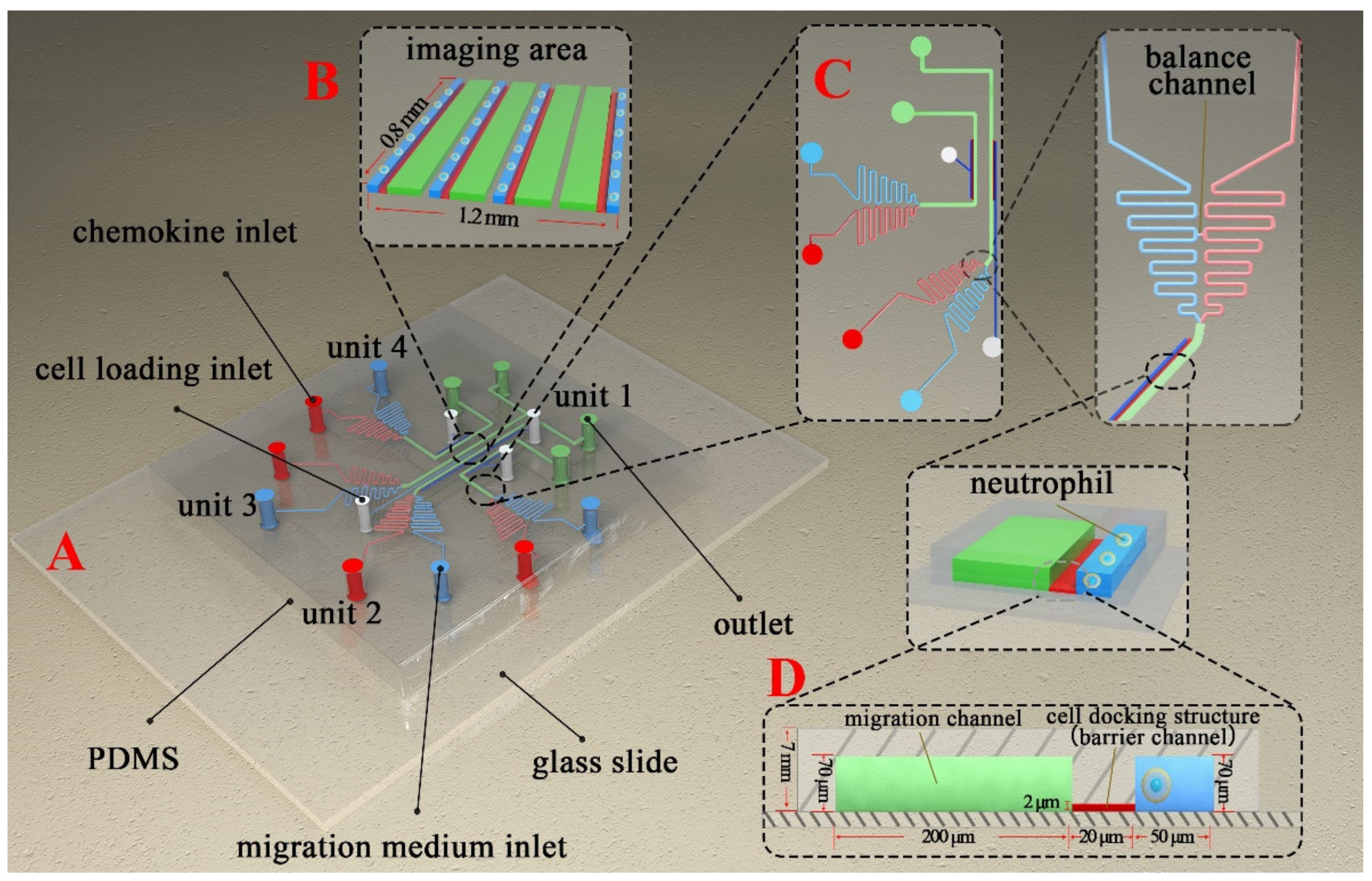
2.1. F4-Chip Design and Fabrication
2.2. Neutrophil Isolation
2.3. Neutrophil Pretreatment
2.4. Cell Migration Experiments
2.4.1. Cell Loading
2.4.2. Gradient Generation
2.4.3. Image Capture
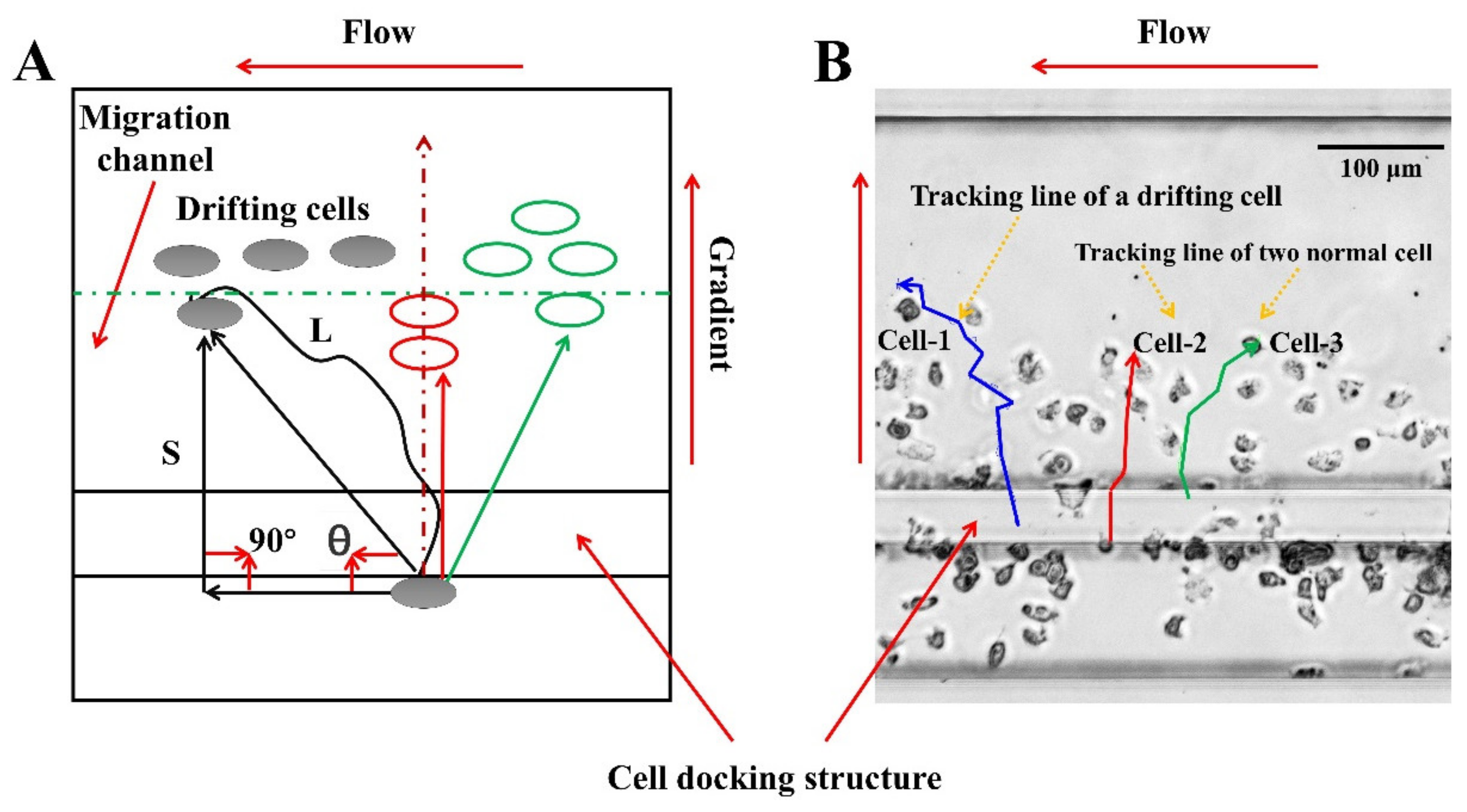
2.5. Data Analysis
3. Results
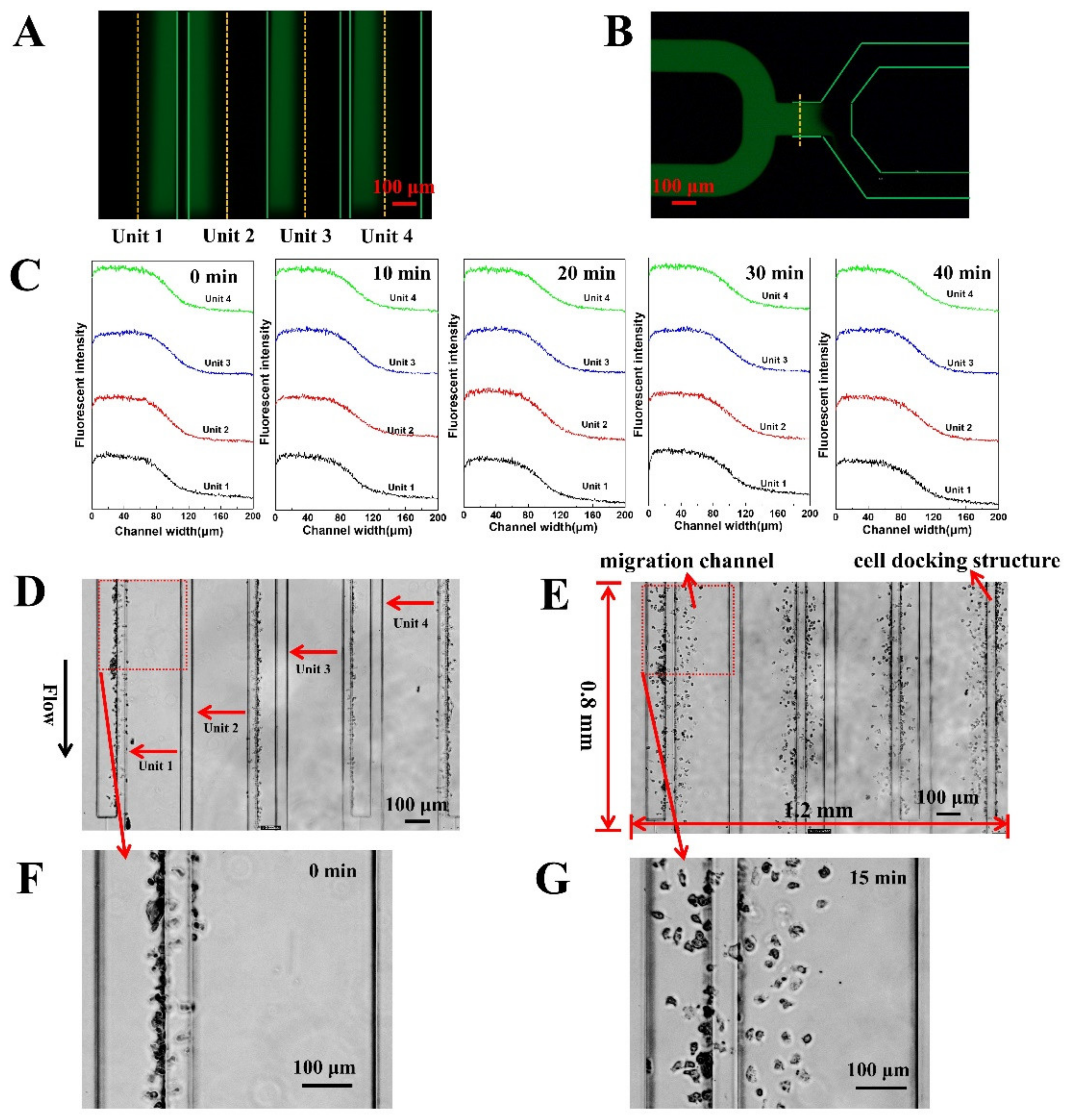
3.1. The F4-Chip and Gradient Characterization
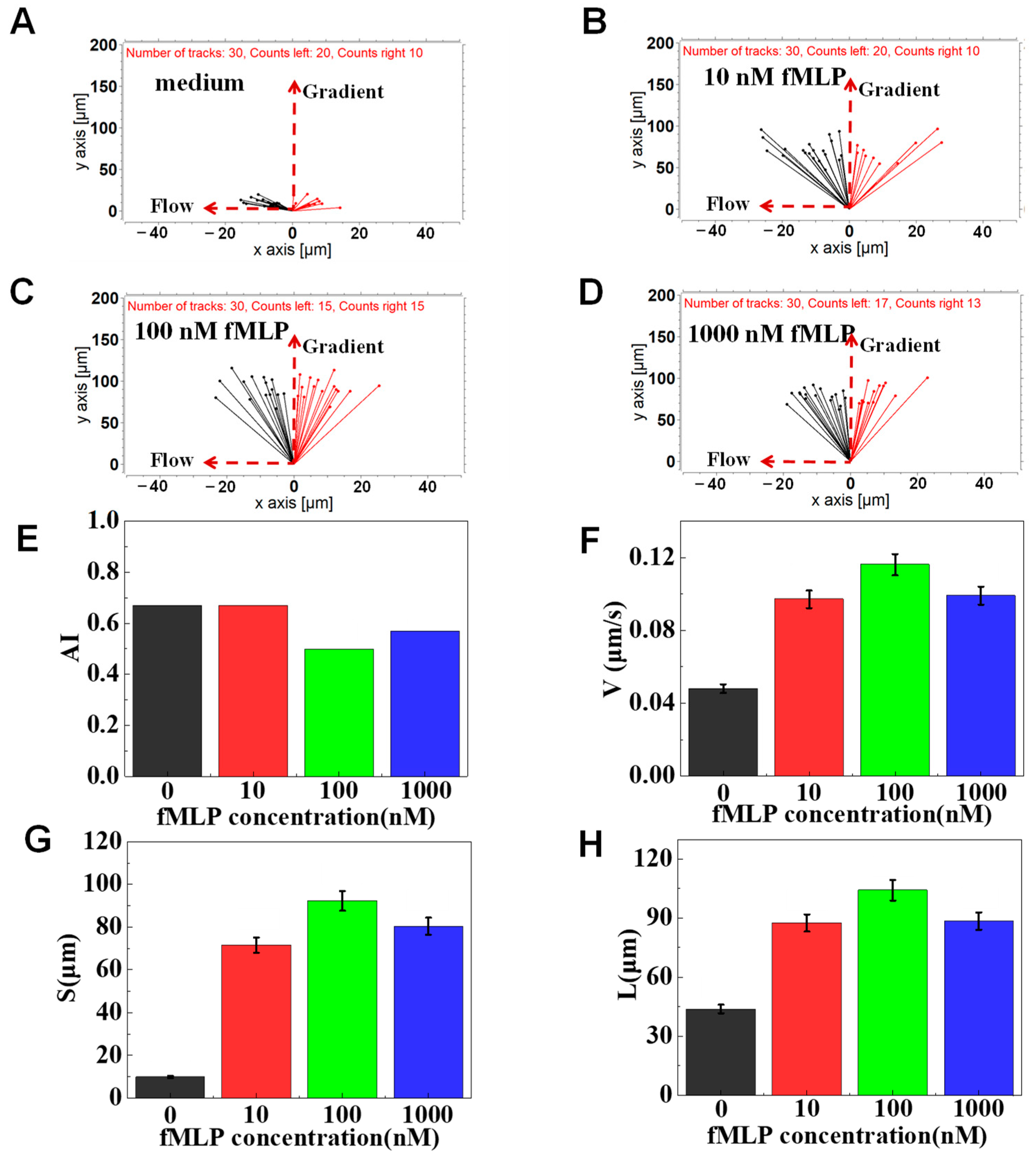
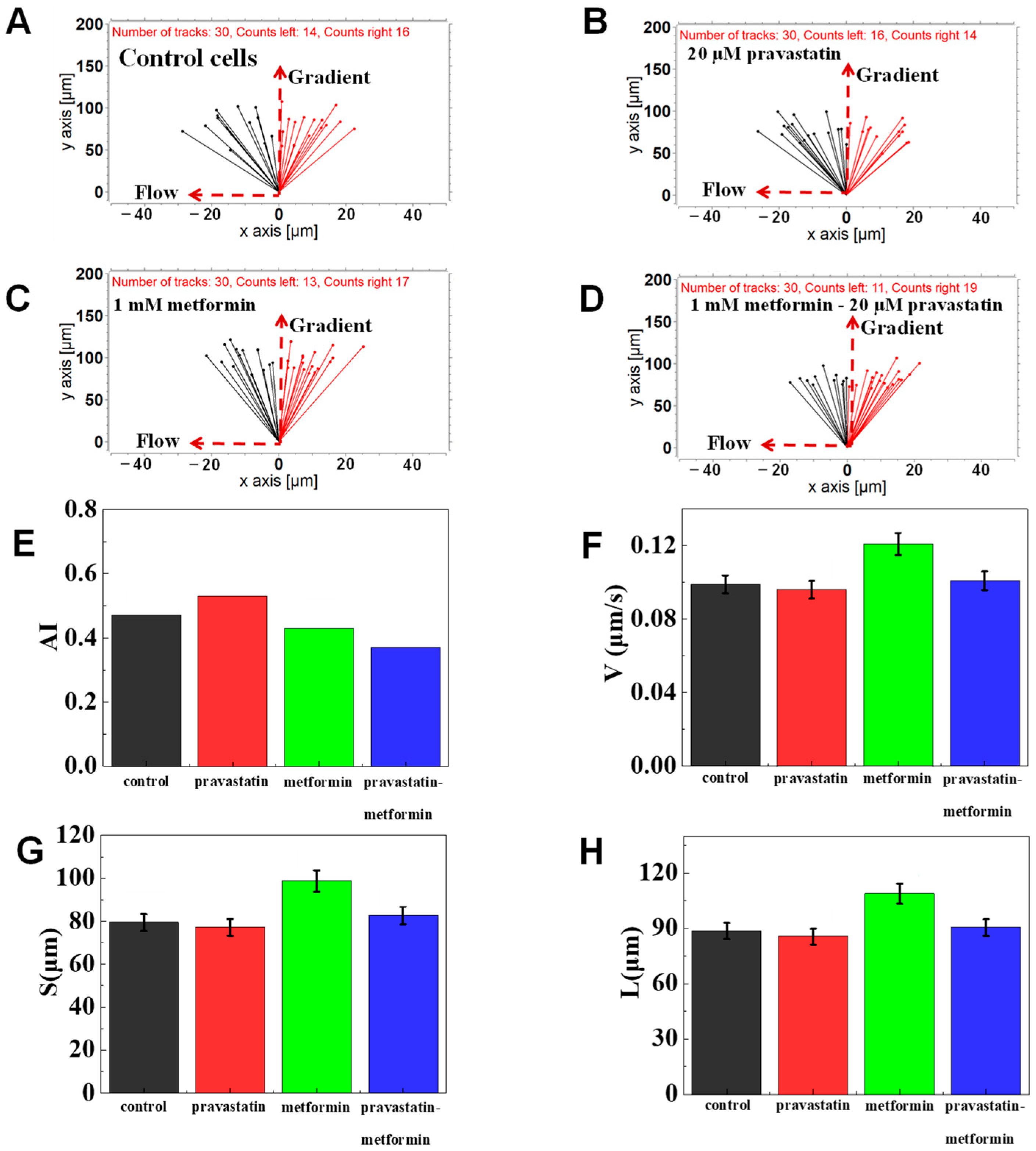
3.2. On-Chip Neutrophil Chemotaxis Assay
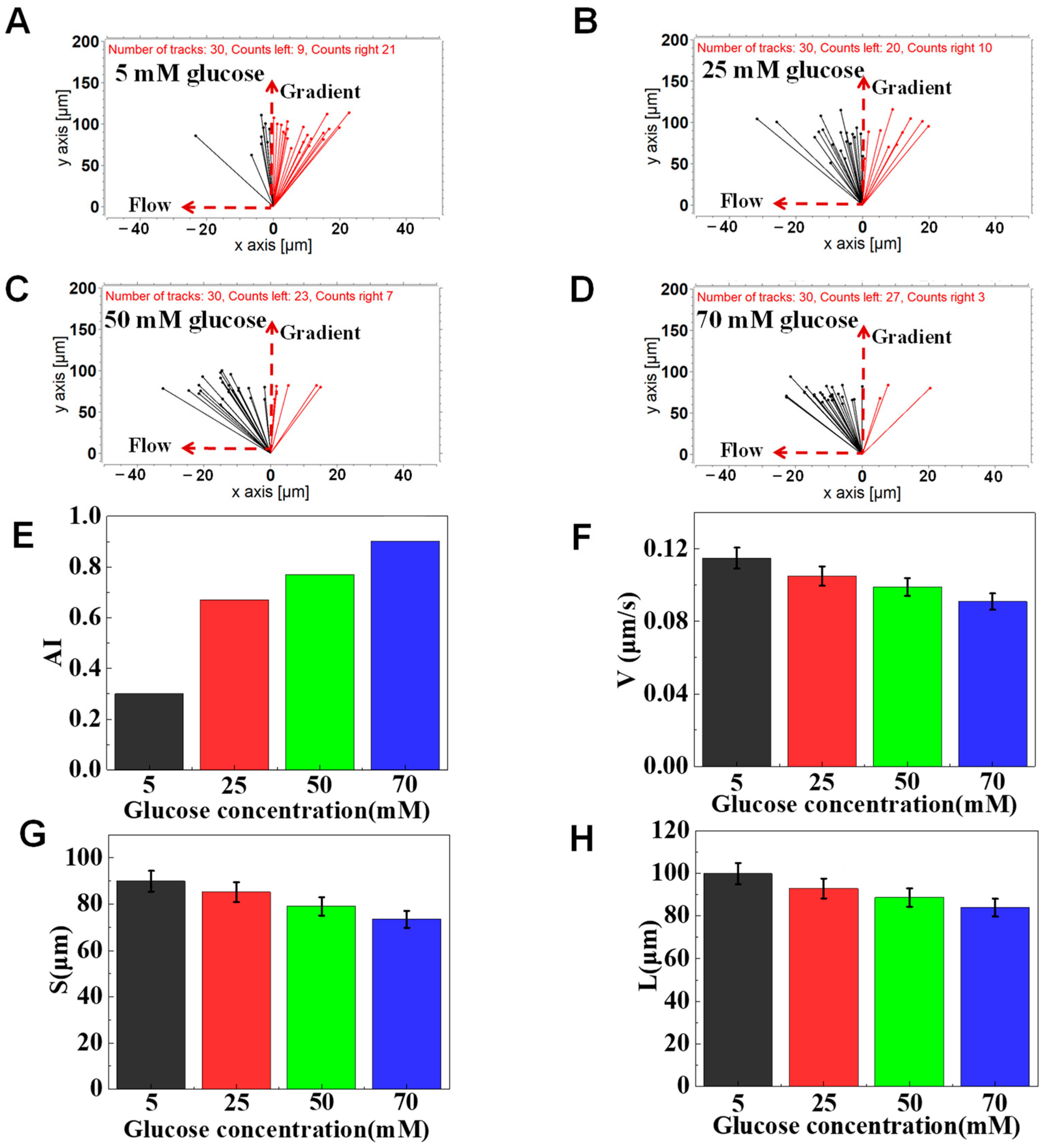
3.3. The Function of Glucose on Neutrophil Chemotaxis
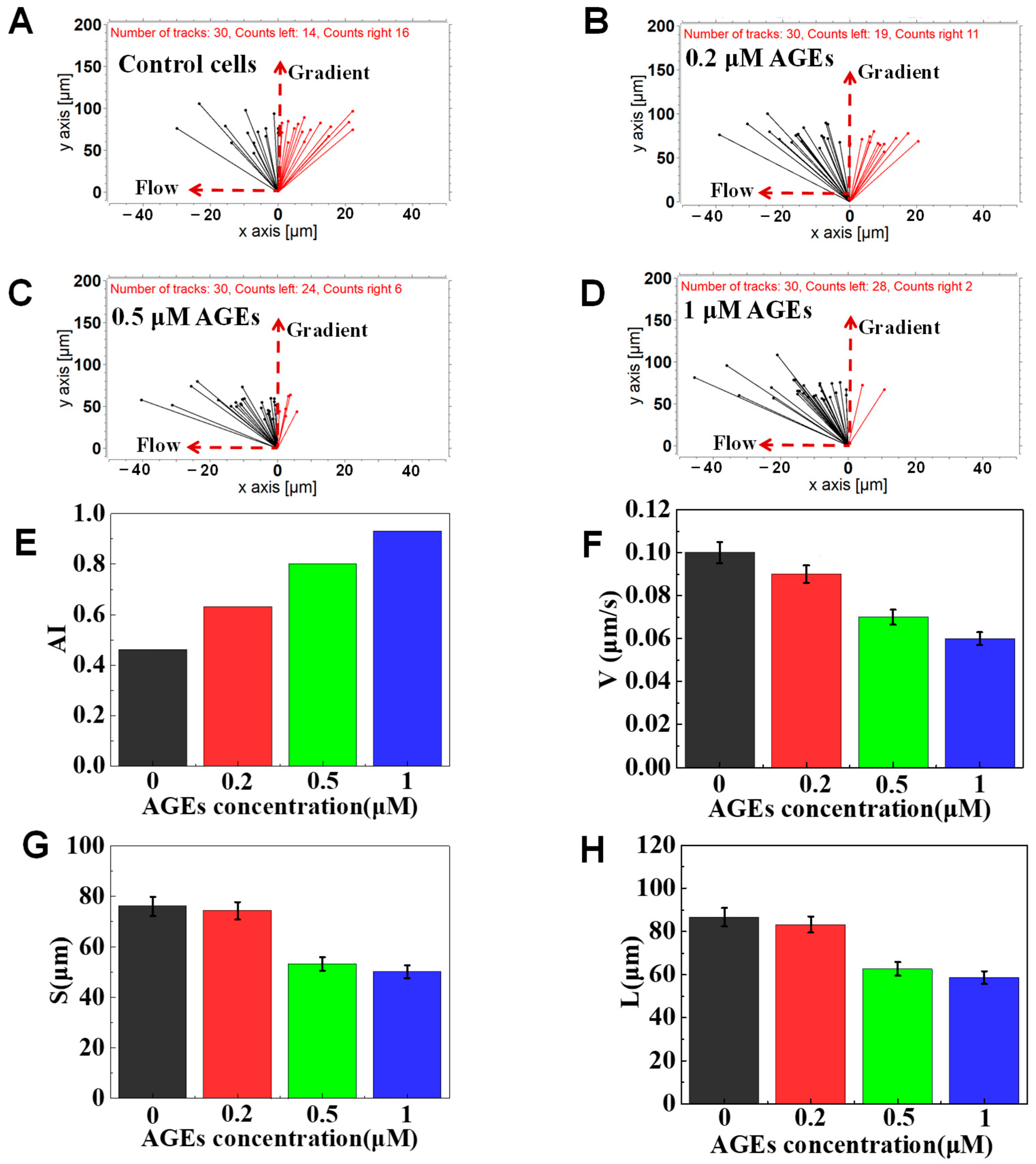
3.4. The Function of AGEs on Neutrophil Chemotaxis
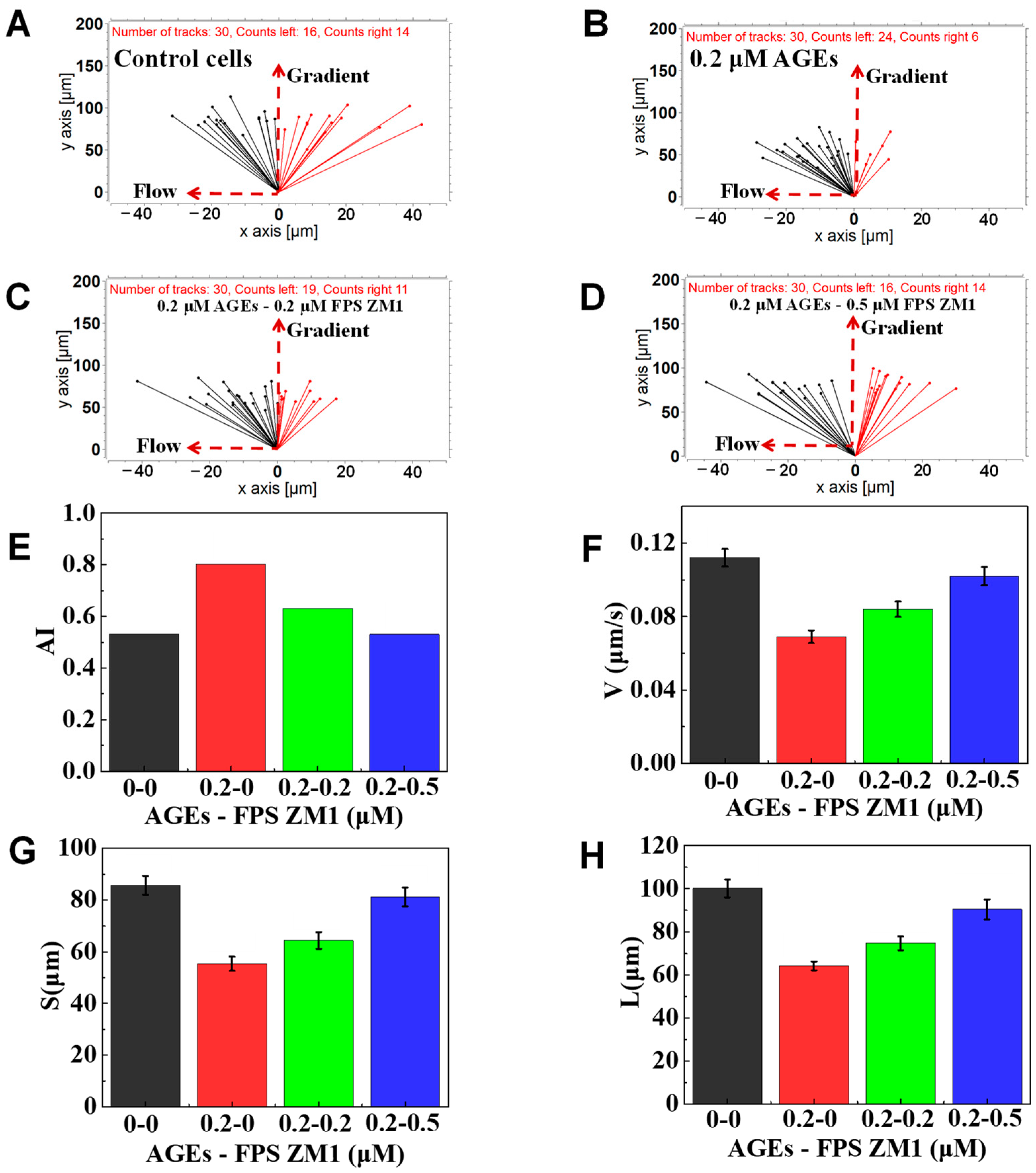
3.5. The Function of FPS-ZM1 on Neutrophil Chemotaxis
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Golden, T.N.; Simmons, R.A. Immune dysfunction in developmental programming of type 2 diabetes mellitus. Nat. Rev. Endocrinol. 2021, 17, 235–245. [Google Scholar] [CrossRef] [PubMed]
- Cole, J.B.; Florez, J.C. Genetics of diabetes mellitus and diabetes complications. Nat. Rev. Nephrol. 2020, 16, 377–390. [Google Scholar] [CrossRef]
- Lin, Q.; Zhou, W.; Wang, Y.; Huang, J.; Hui, X.; Zhou, Z.; Xiao, Y. Abnormal Peripheral Neutrophil Transcriptome in Newly Diagnosed Type 2 Diabetes Patients. J. Diabetes Res. 2020, 2020, 9519072. [Google Scholar] [CrossRef]
- Klopf, J.; Brostjan, C.; Eilenberg, W.; Neumayer, C. Neutrophil Extracellular Traps and Their Implications in Cardiovascular and Inflammatory Disease. Int. J. Mol. Sci. 2021, 22, 559. [Google Scholar] [CrossRef] [PubMed]
- Hughes, M.J.; McGettrick, H.M.; Sapey, E. Shared mechanisms of multimorbidity in COPD, atherosclerosis and type-2 diabetes: The neutrophil as a potential inflammatory target. Eur. Respir. Rev. 2020, 29, 190102. [Google Scholar] [CrossRef] [PubMed]
- Hirota, T.; Levy, J.H.; Iba, T. The influence of hyperglycemia on neutrophil extracellular trap formation and endothelial glycocalyx damage in a mouse model of type 2 diabetes. Microcirculation 2020, 27, e12617. [Google Scholar] [CrossRef] [PubMed]
- Yu, Y.; Lin, Q.; Ye, D.; Wang, Y.; He, B.; Li, Y.; Huang, G.; Zhou, Z.; Xiao, Y. Neutrophil count as a reliable marker for diabetic kidney disease in autoimmune diabetes. BMC Endocr. Disord. 2020, 20, 158. [Google Scholar] [CrossRef]
- Raj, P.N.; Shaji, B.V.; Haritha, V.H.; Anie, Y. Neutrophil secretion modulates neutrophil and monocyte functions during hyperglucose and/or hyperinsulin conditions in vitro. J. Cell. Immunother. 2018, 4, 65–70. [Google Scholar] [CrossRef]
- Schuetz, P.; Castro, P.; Shapiro, N.I. Diabetes and Sepsis: Preclinical Findings and Clinical Relevance. Diabetes Care 2011, 34, 771. [Google Scholar] [CrossRef]
- Insuela, D.; Coutinho, D.; Martins, M.; Ferrero, M.; Carvalho, V. Neutrophil Function Impairment Is a Host Susceptibility Factor to Bacterial Infection in Diabetes. Cells Immune Syst. 2019, 1–22. [Google Scholar] [CrossRef]
- Wierusz-Wysocka, B.; Wysocki, H.; Wykrętowicz, A.; Klimas, R. The influence of increasing glucose concentrations on selected functions of polymorphonuclear neutrophils. Acta Diabetol. Lat. 1988, 25, 283–288. [Google Scholar] [CrossRef]
- Milicevic, Z.; Raz, I.; Beattie, S.D.; Campaigne, B.N.; Hanefeld, M.J.D.C. Natural History of Cardiovascular Disease in Patients With Diabetes. Diabetes Care 2008, 31 Suppl. S2, S155–S160. [Google Scholar] [CrossRef]
- Yubero-Serrano, E.M.; Pérez-Martínez, P. Advanced Glycation End Products and Their Involvement in Cardiovascular Disease. Angiology 2020, 71, 698–700. [Google Scholar] [CrossRef] [PubMed]
- Nin, J.W.; Jorsal, A.; Ferreira, I.; Schalkwijk, C.G.; Prins, M.H.; Parving, H.-H.; Tarnow, L.; Rossing, P.; Stehouwer, C.D. Higher Plasma Levels of Advanced Glycation End Products Are Associated With Incident Cardiovascular Disease and All-Cause Mortality in Type 1 Diabetes. Diabetes Care 2011, 34, 442–447. [Google Scholar] [CrossRef] [PubMed]
- Yan, S.F.; Ramasamy, R.; Schmidt, A.M. Mechanisms of Disease: Advanced glycation end-products and their receptor in inflammation and diabetes complications. Nat. Clin. Pract. Endocrinol. Metab. 2008, 4, 285–293. [Google Scholar] [CrossRef] [PubMed]
- Spadaccio, C.; De Marco, F.; Di Domenico, F.; Coccia, R.; Lusini, M.; Barbato, R.; Covino, E.; Chello, M. Simvastatin attenuates the endothelial pro-thrombotic shift in saphenous vein grafts induced by Advanced glycation endproducts. Thromb. Res. 2014, 133, 418–425. [Google Scholar] [CrossRef] [PubMed]
- Touré, F.; Zahm, J.-M.; Garnotel, R.; Lambert, E.; Bonnet, N.; Schmidt, A.M.; Vitry, F.; Chanard, J.; Gillery, P.; Rieu, P. Receptor for advanced glycation end-products (RAGE) modulates neutrophil adhesion and migration on glycoxidated extracellular matrix. Biochem. J. 2008, 416, 255–261. [Google Scholar] [CrossRef] [PubMed]
- Wong, R.K.M.; Pettit, A.I.; Davies, J.E.; Ng, L.L. Augmentation of the Neutrophil Respiratory Burst Through the Action of Advanced Glycation End Products. Diabetes 2002, 51, 2846–2853. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Collison, K.S.; Parhar, R.S.; Saleh, S.S.; Meyer, B.F.; Kwaasi, A.A.; Hammami, M.M.; Schmidt, A.M.; Stern, D.M.; Al-Mohanna, F.A. RAGE-mediated neutrophil dysfunction is evoked by advanced glycation end products (AGEs). J. Leukoc. Biol. 2002, 71, 433. [Google Scholar]
- Satti, S.; Deng, P.; Matthews, K.; Duffy, S.P.; Ma, H. Multiplexed end-point microfluidic chemotaxis assay using centrifugal alignment. Lab Chip 2020, 20, 3096–3103. [Google Scholar] [CrossRef]
- Garcia-Seyda, N.; Aoun, L.; Tishkova, V.; Seveau, V.; Biarnes-Pelicot, M.; Bajénoff, M.; Valignat, M.-P.; Theodoly, O. Microfluidic device to study flow-free chemotaxis of swimming cells. Lab Chip 2020, 20, 1639–1647. [Google Scholar] [CrossRef]
- Grigolato, F.; Egholm, C.; Impellizzieri, D.; Arosio, P.; Boyman, O. Establishment of a scalable microfluidic assay for characterization of population-based neutrophil chemotaxis. Allergy 2020, 75, 1382–1393. [Google Scholar] [CrossRef]
- Zhao, W.; Zhao, H.; Li, M.; Huang, C. Microfluidic devices for neutrophil chemotaxis studies. J. Transl. Med. 2020, 18, 168. [Google Scholar] [CrossRef]
- Ren, X.; Wu, J.; Levin, D.; Santos, S.; de Faria, R.L.; Zhang, M.; Lin, F. Sputum from chronic obstructive pulmonary disease patients inhibits T cell migration in a microfluidic device. Ann. New York Acad. Sci. 2019, 1445, 52–61. [Google Scholar] [CrossRef] [PubMed]
- Srisawat, N.; Kulvichit, W.; Tungsanga, S.; Peerapornratana, S.; Vorasitchai, S.; Tangkanakul, C.; Lumlertgul, N.; Komaenthammasophon, C.; Praditpornsilpa, K.; Tungsanga, K.; et al. The role of neutrophil chemotaxis activity as an immunologic biomarker to predict mortality in critically-ill patients with severe sepsis. J. Crit. Care 2020, 56, 215–221. [Google Scholar] [CrossRef] [PubMed]
- Otawara, M.; Roushan, M.; Wang, X.; Ellett, F.; Yu, Y.-M.; Irimia, D. Microfluidic Assay Measures Increased Neutrophil Extracellular Traps Circulating in Blood after Burn Injuries. Sci. Rep. 2018, 8, 16983. [Google Scholar] [CrossRef]
- Wang, X.; Irimia, D. Neutrophil Chemotaxis in One Droplet of Blood Using Microfluidic Assays. Methods Mol. Biol. 2018, 1749, 351. [Google Scholar]
- Muldur, S.; Ellett, F.; Marand, A.L.; Marvil, C.; Branda, J.A.; LeMieux, J.E.; Raff, A.B.; Strle, K.; Irimia, D. Microfluidic Assays for Probing Neutrophil-Borrelia Interactions in Blood During Lyme Disease. Cells Tissues Organs 2021. [Google Scholar] [CrossRef] [PubMed]
- Virumbrales-Muñoz, M.; Ayuso, J.M.; Gong, M.M.; Humayun, M.; Livingston, M.K.; Lugo-Cintrón, K.M.; McMinn, P.; Álvarez-García, Y.R.; Beebe, D.J. Microfluidic lumen-based systems for advancing tubular organ modeling. Chem. Soc. Rev. 2020, 49, 6402–6442. [Google Scholar] [CrossRef]
- McMinn, P.H.; Hind, L.E.; Huttenlocher, A.; Beebe, D.J. Neutrophil trafficking on-a-chip: An in vitro, organotypic model for investigating neutrophil priming, extravasation, and migration with spatiotemporal control. Lab Chip 2019, 19, 3697–3705. [Google Scholar] [CrossRef]
- Ren, X.; Getschman, A.E.; Hwang, S.; Volkman, B.F.; Klonisch, T.; Levin, D.; Zhao, M.; Santos, S.; Liu, S.; Cheng, J.; et al. Investigations on T cell transmigration in a human skin-on-chip (SoC) model. Lab Chip 2021, 21, 1527–1539. [Google Scholar] [CrossRef] [PubMed]
- Wu, J.; Kumar-Kanojia, A.; Hombach-Klonisch, S.; Klonisch, T.; Lin, F. A radial microfluidic platform for higher throughput chemotaxis studies with individual gradient control. Lab Chip 2018, 18, 3855–3864. [Google Scholar] [CrossRef]
- Hou, H.W.; Petchakup, C.; Tay, H.M.; Tam, Z.Y.; Dalan, R.; Chew, D.E.K.; Li, K.H.H.; Boehm, B.O. Rapid and label-free microfluidic neutrophil purification and phenotyping in diabetes mellitus. Sci. Rep. 2016, 6, 29410. [Google Scholar] [CrossRef]
- Yang, K.; Peretz-Soroka, H.; Wu, J.; Zhu, L.; Cui, X.; Zhang, M.; Rigatto, C.; Liu, Y.; Lin, F. Fibroblast growth factor 23 weakens chemotaxis of human blood neutrophils in microfluidic devices. Sci. Rep. 2017, 7, 3100. [Google Scholar] [CrossRef]
- Yang, K.; Wu, J.; Peretz-Soroka, H.; Zhu, L.; Li, Z.; Sang, Y.; Hipolito, J.; Zhang, M.; Santos, S.; Hillier, C.; et al. Mkit: A cell migration assay based on microfluidic device and smartphone. Biosens. Bioelectron. 2018, 99, 259–267. [Google Scholar] [CrossRef]
- Salek, M.M.; Carrara, F.; Fernandez, V.; Guasto, J.S.; Stocker, R. Bacterial chemotaxis in a microfluidic T-maze reveals strong phenotypic heterogeneity in chemotactic sensitivity. Nat. Commun. 2019, 10, 1877. [Google Scholar] [CrossRef] [PubMed]
- Coluccio, M.L.; D’Attimo, M.A.; Cristiani, C.M.; Candeloro, P.; Parrotta, E.; Dattola, E.; Guzzi, F.; Cuda, G.; Lamanna, E.; Carbone, E.; et al. A Passive Microfluidic Device for Chemotaxis Studies. Micromachines 2019, 10, 551. [Google Scholar] [CrossRef]
- Qin, D.; Xia, Y.; Whitesides, G.M.J.N.P. Soft lithography for micro-and nanoscale patterning. Nature Protoc. 2010, 5, 491. [Google Scholar] [CrossRef]
- Hipolito, J.; Peretz-Soroka, H.; Torres, A.M.; Booy, E.; Yang, K.; Gupta, M.; Meier, M.; McKenna, S.; Koch, M.; Santos, S.; et al. Microfluidic Devices for Studying the Effect of Netrin-1 on Neutrophil and Breast Cancer Cell Migration. Adv. Biosyst. 2018, 2, 1700178. [Google Scholar] [CrossRef]
- Irimia, D.; Charras, G.; Agrawal, N.; Mitchison, T.; Toner, M. Polar stimulation and constrained cell migration in microfluidic channels. Lab Chip 2007, 7, 1783–1790. [Google Scholar] [CrossRef] [PubMed]
- Daughaday, C.C.; Mehta, J.; Spilberg, I.; Atkinson, J.P. Deactivation of guinea pig pulmonary alveolar macrophage responses to N-formyl-methionyl-leucyl-phenylalanine: Chemotaxis, superoxide generation, and binding. J. Immunol. 1985, 134, 1823–1826. [Google Scholar]
- Nast, C.C.; LeDuc, L.E. Chemotactic peptides. Dig. Dis. Sci. 1988, 33, 50S–57S. [Google Scholar] [CrossRef]
- Park, D.W.; Jiang, S.; Tadie, J.-M.; Stigler, W.S.; Gao, Y.; Deshane, J.; Abraham, E.; Zmijewski, J.W. Activation of AMPK Enhances Neutrophil Chemotaxis and Bacterial Killing. Mol. Med. 2013, 19, 387–398. [Google Scholar] [CrossRef] [PubMed]
- Dunzendorfer, S.; Rothbucher, D.; Schratzberger, P.; Reinisch, N.; Kähler, C.M.; Wiedermann, C.J. Mevalonate-Dependent Inhibition of Transendothelial Migration and Chemotaxis of Human Peripheral Blood Neutrophils by Pravastatin. Circ. Res. 1997, 81, 963–969. [Google Scholar] [CrossRef] [PubMed]
- Makita, Z.; Vlassara, H.; Cerami, A.; Bucala, R.J.J.O.B.C. Immunochemical detection of advanced glycosylation end products. J. Biol. Chem. 1992, 267, 5133–5138. [Google Scholar] [CrossRef]
- Advani, A.; Marshall, S.M.; Thomas, T.H. Impaired neutrophil store-mediated calcium entry in Type 2 diabetes. Eur. J. Clin. Investig. 2004, 34, 43–49. [Google Scholar] [CrossRef] [PubMed]
- Na, M.; Mohammad, M.; Fei, Y.; Wang, W.; Holdfeldt, A.; Forsman, H.; Ali, A.; Pullerits, R.; Jin, T. Lack of Receptor for Advanced Glycation End Products Leads to Less Severe Staphylococcal Skin Infection but More Skin Abscesses and Prolonged Wound Healing. J. Infect. Dis. 2018, 218, 791–800. [Google Scholar] [CrossRef]
- Mihaylova, M.M.; Shaw, R.J. The AMPK signalling pathway coordinates cell growth, autophagy and metabolism. Nat Cell Biol 2011, 13, 1016–1023. [Google Scholar] [CrossRef] [PubMed]
- Diaz-Morales, N.; Rovira-Llopis, S.; Bañuls, C.; Escribano-Lopez, I.; de Marañon, A.M.; Lopez-Domenech, S.; Orden, S.; Roldan-Torres, I.; Alvarez, A.; Veses, S.; et al. Are Mitochondrial Fusion and Fission Impaired in Leukocytes of Type 2 Diabetic Patients? Antioxid. Redox Signal. 2016, 25, 108–115. [Google Scholar] [CrossRef] [PubMed]
- Al-Awar, A.; Kupai, K.; Veszelka, M.; Szűcs, G.; Attieh, Z.; Murlasits, Z.; Török, S.; Pósa, A.; Varga, C. Experimental Diabetes Mellitus in Different Animal Models. J. Diabetes Res. 2016, 2016, 9051426. [Google Scholar] [CrossRef]
- Rezende, L.P.; Galheigo, M.R.U.; Landim, B.C.; Cruz, A.R.; Botelho, F.V.; Zanon, R.G.; Góes, R.M.; Ribeiro, D.L. Effect of glucose and palmitate environment on proliferation and migration of PC3-prostate cancer cells. Cell Biol. Int. 2019, 43, 373–383. [Google Scholar] [CrossRef]
- Yang, L.; Liu, J.; Shan, Q.; Geng, G.; Shao, P. High glucose inhibits proliferation and differentiation of osteoblast in alveolar bone by inducing pyroptosis. Biochem. Biophys. Res. Commun. 2020, 522, 471–478. [Google Scholar] [CrossRef] [PubMed]
- Willershausen-Zönnchen, B.; Lemmen, C.; Hamn, G. Influence of high glucose concentrations on glycosaminoglycan and collagen synthesis in cultured human gingival fibroblasts. J. Clin. Periodontol. 1991, 18, 190–195. [Google Scholar] [CrossRef]
- Deveci, M.; Gilmont, R.R.; Dunham, W.R.; Mudge, B.P.; Smith, D.J.; Marcelo, C.L. Glutathione enhances fibroblast collagen contraction and protects keratinocytes from apoptosis in hyperglycaemic culture. Br. J. Dermatol. 2005, 152, 217–224. [Google Scholar] [CrossRef]
- Takeuchi, M.; Yamagishi, S.-I. Involvement of Toxic AGEs (TAGE) in the Pathogenesis of Diabetic Vascular Complications and Alzheimer’s Disease. J. Alzheimer’s Dis. 2009, 16, 845–858. [Google Scholar] [CrossRef] [PubMed]
- Sionov, R.V.; Fainsod-Levi, T.; Zelter, T.; Polyansky, L.; Pham, C.T.; Granot, Z. Neutrophil Cathepsin G and Tumor Cell RAGE Facilitate Neutrophil Anti-Tumor Cytotoxicity. OncoImmunology 2019, 8, e1624129. [Google Scholar] [CrossRef] [PubMed]
- Ribeiro, D.; Freitas, M.; Rocha, S.; Lima, J.L.F.C.; Carvalho, F.; Fernandes, E. Calcium Pathways in Human Neutrophils—The Extended Effects of Thapsigargin and ML-9. Cells 2018, 7, 204. [Google Scholar] [CrossRef]








Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yang, K.; Yang, X.; Gao, C.; Hua, C.; Hong, C.; Zhu, L. A Novel Microfluidic Device for the Neutrophil Functional Phenotype Analysis: Effects of Glucose and Its Derivatives AGEs. Micromachines 2021, 12, 944. https://doi.org/10.3390/mi12080944
Yang K, Yang X, Gao C, Hua C, Hong C, Zhu L. A Novel Microfluidic Device for the Neutrophil Functional Phenotype Analysis: Effects of Glucose and Its Derivatives AGEs. Micromachines. 2021; 12(8):944. https://doi.org/10.3390/mi12080944
Chicago/Turabian StyleYang, Ke, Xiao Yang, Chaoru Gao, Changyi Hua, Chenggang Hong, and Ling Zhu. 2021. "A Novel Microfluidic Device for the Neutrophil Functional Phenotype Analysis: Effects of Glucose and Its Derivatives AGEs" Micromachines 12, no. 8: 944. https://doi.org/10.3390/mi12080944
APA StyleYang, K., Yang, X., Gao, C., Hua, C., Hong, C., & Zhu, L. (2021). A Novel Microfluidic Device for the Neutrophil Functional Phenotype Analysis: Effects of Glucose and Its Derivatives AGEs. Micromachines, 12(8), 944. https://doi.org/10.3390/mi12080944