Probing Changes in Ca2+-Induced Interaction Forces between Calmodulin and Melittin by Atomic Force Microscopy

Abstract
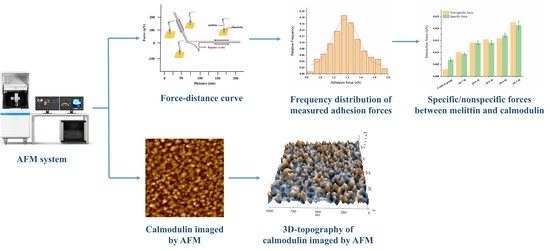
1. Introduction
2. Materials and Methods
2.1. Materials
2.2. Preparation of Gold Substrate
2.3. Self-Assembled Monolayer (SAM) of Thiol on Gold Surfaces
2.4. Protein Immobilization onto the Gold Surfaces
2.5. Functionalization of Atomic Force Microscopy (AFM) Tip
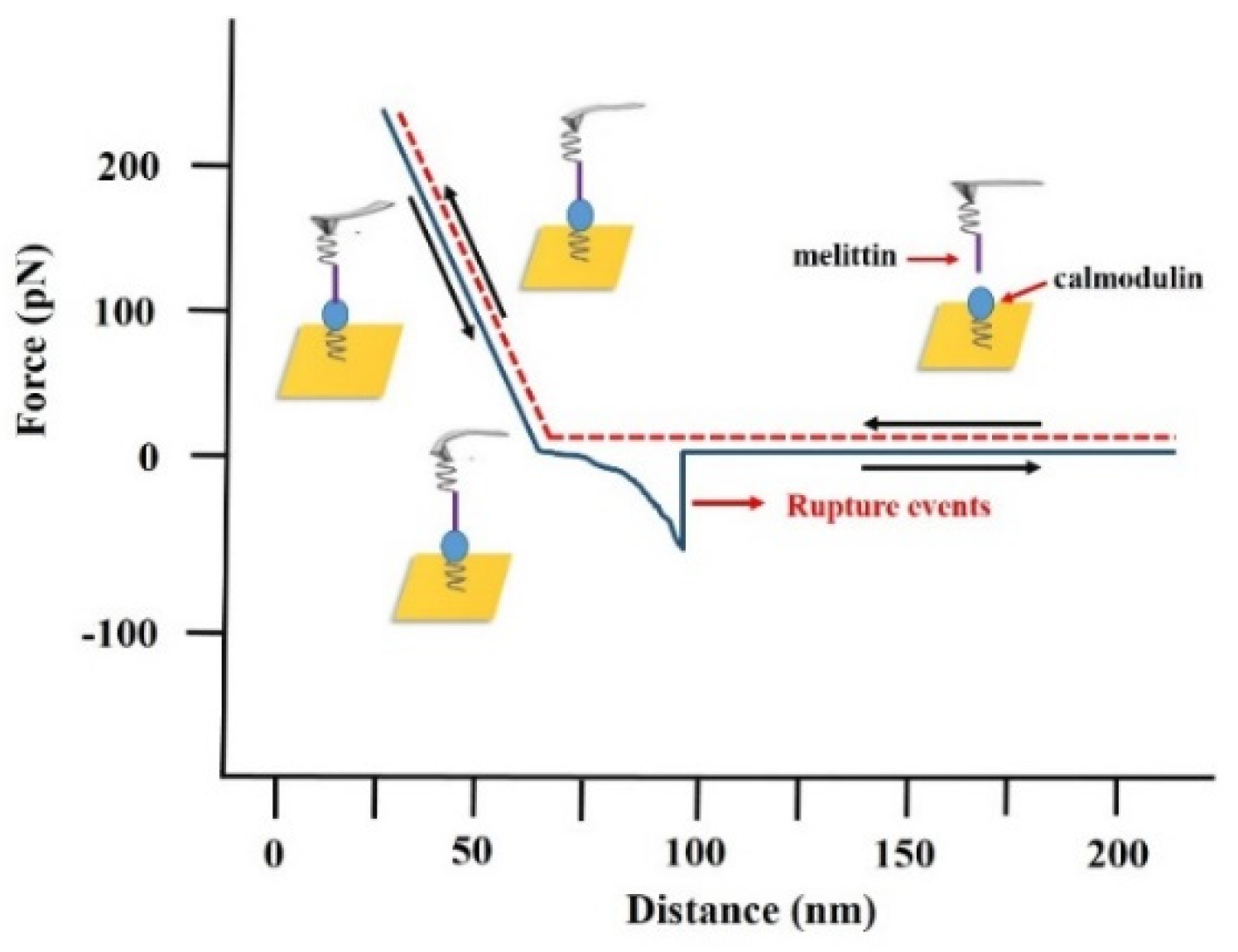
2.6. Measurement of the Melittin-Calmodulin Adhesion Force
2.7. AFM Imaging
3. Results and Discussion
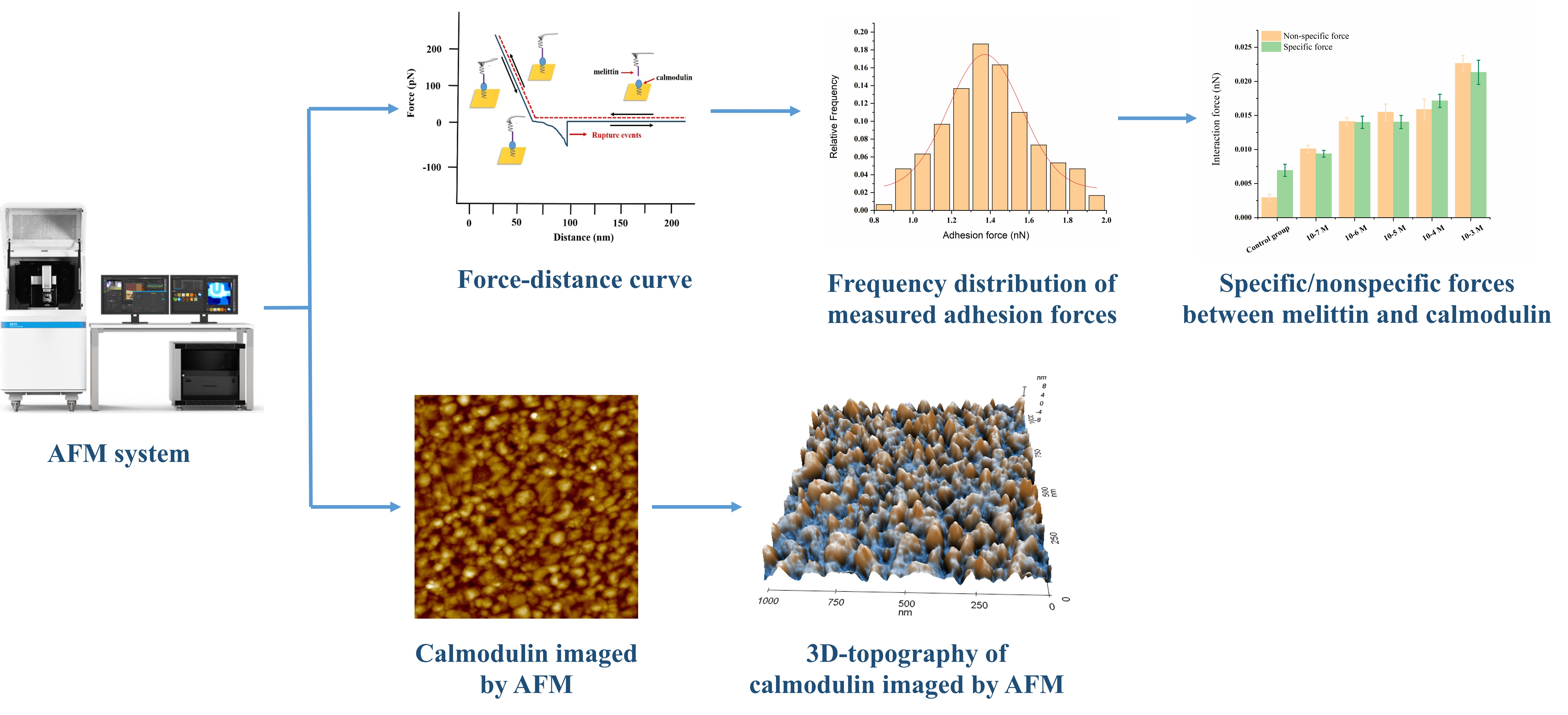
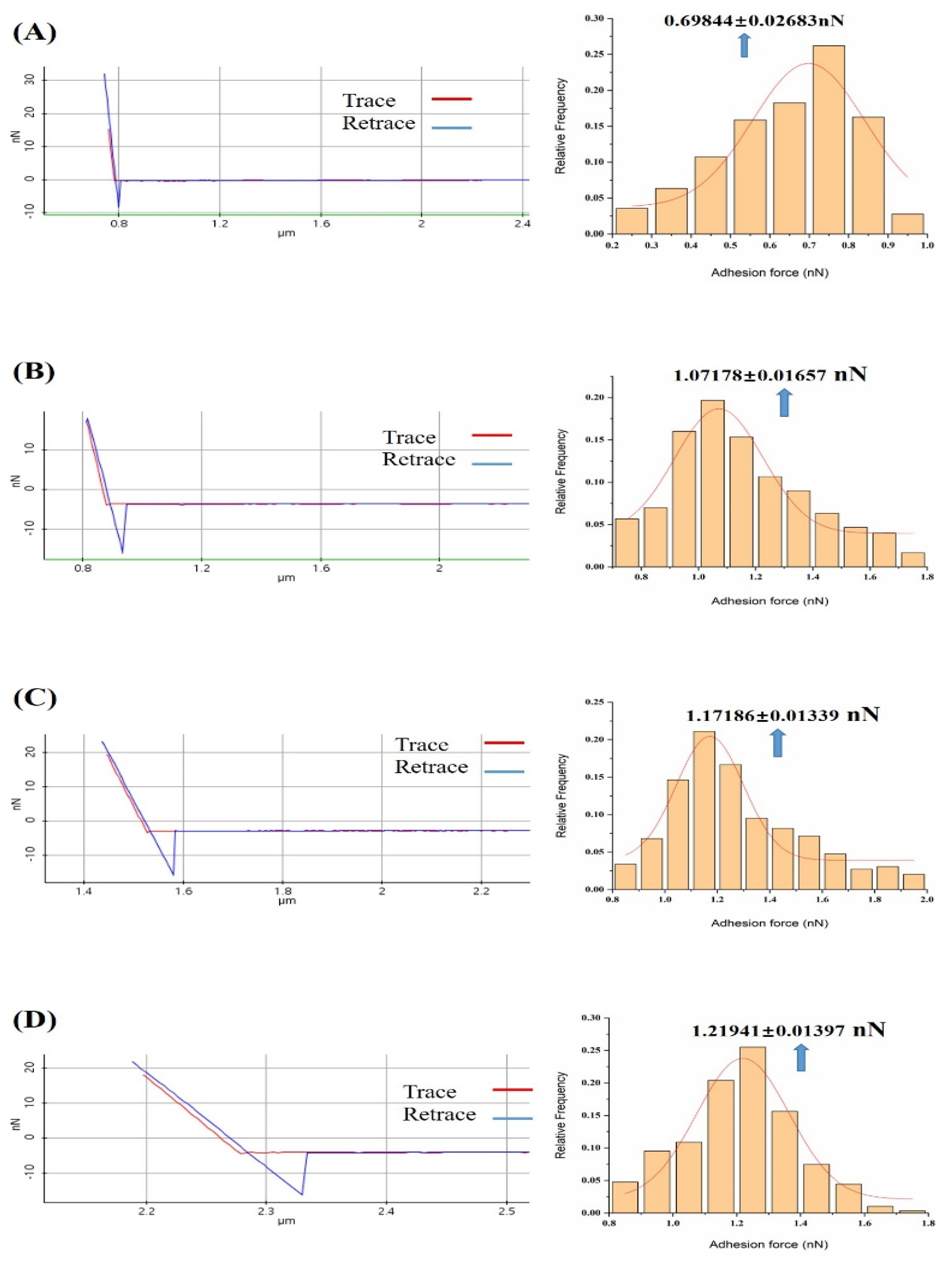
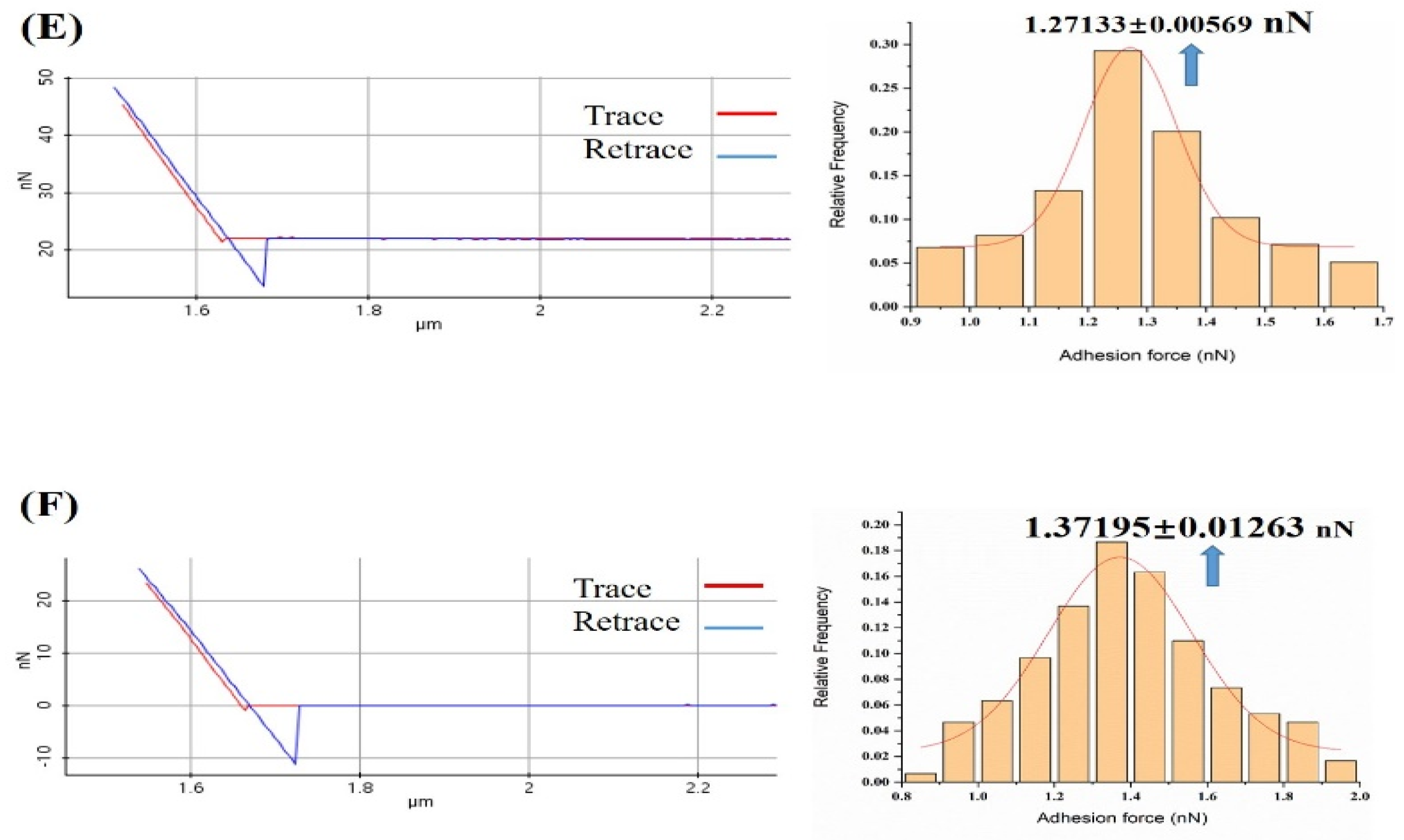
3.1. Probing the Ca2+ Effect in the Interaction Forces between Melittin and Calmodulin
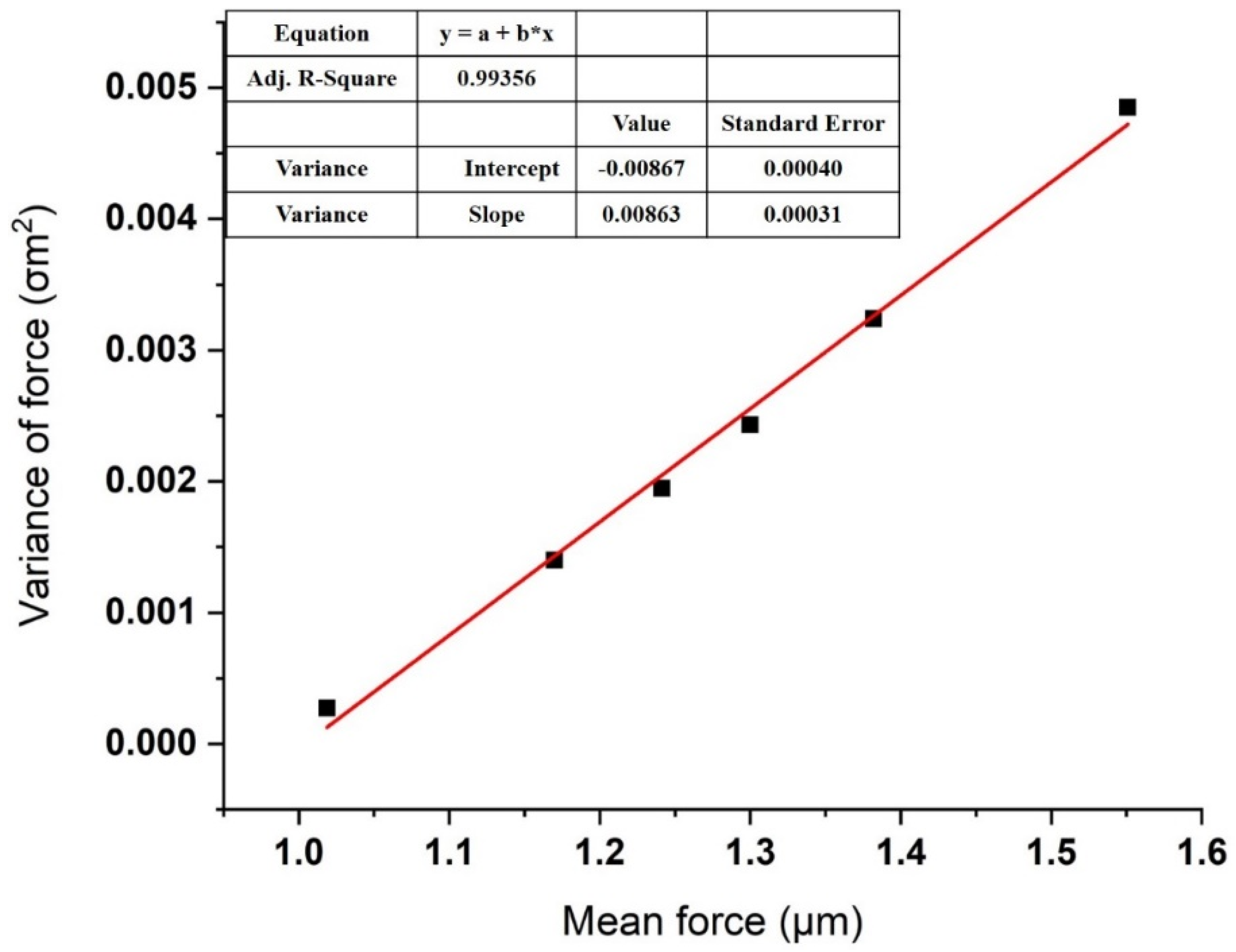
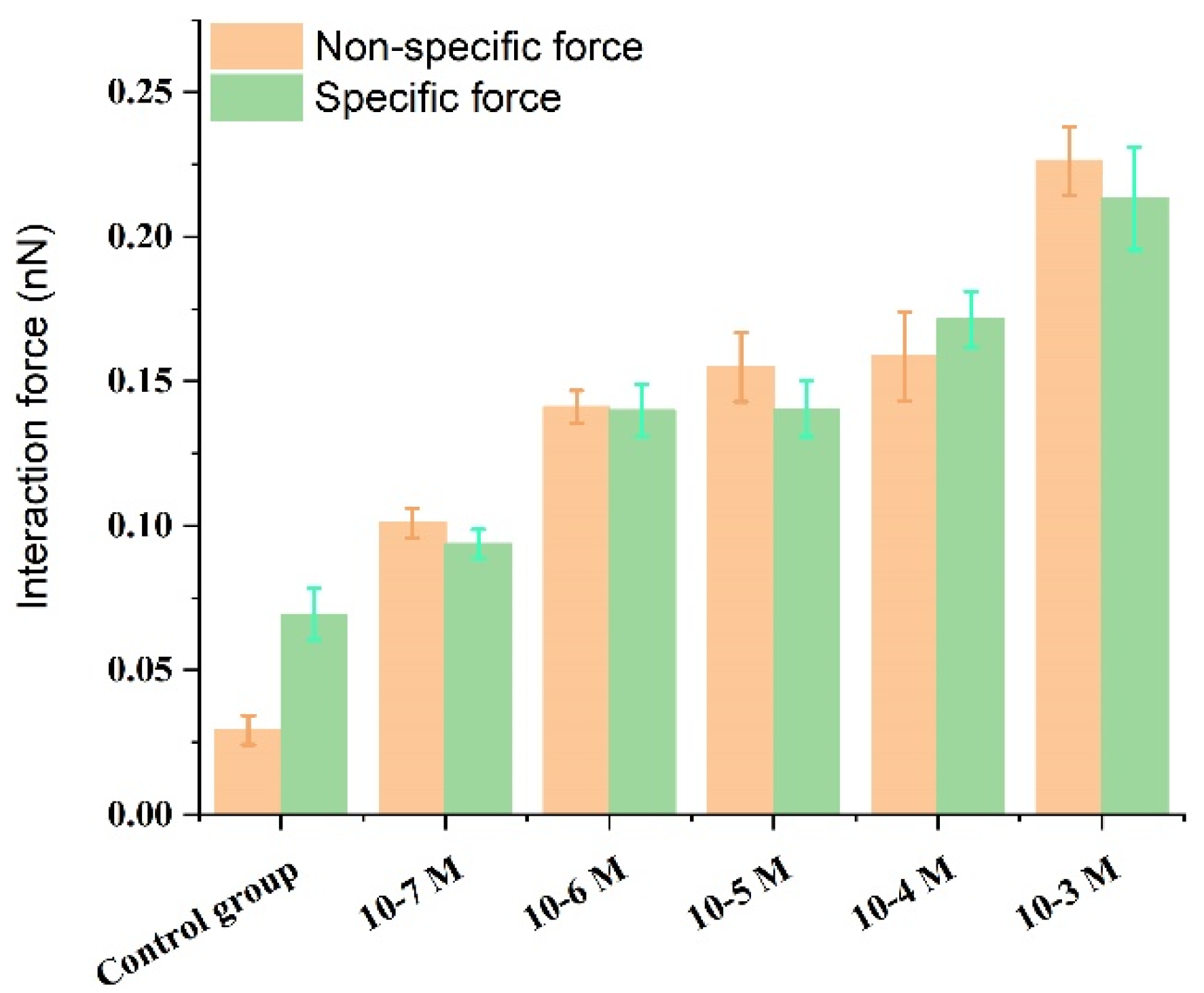
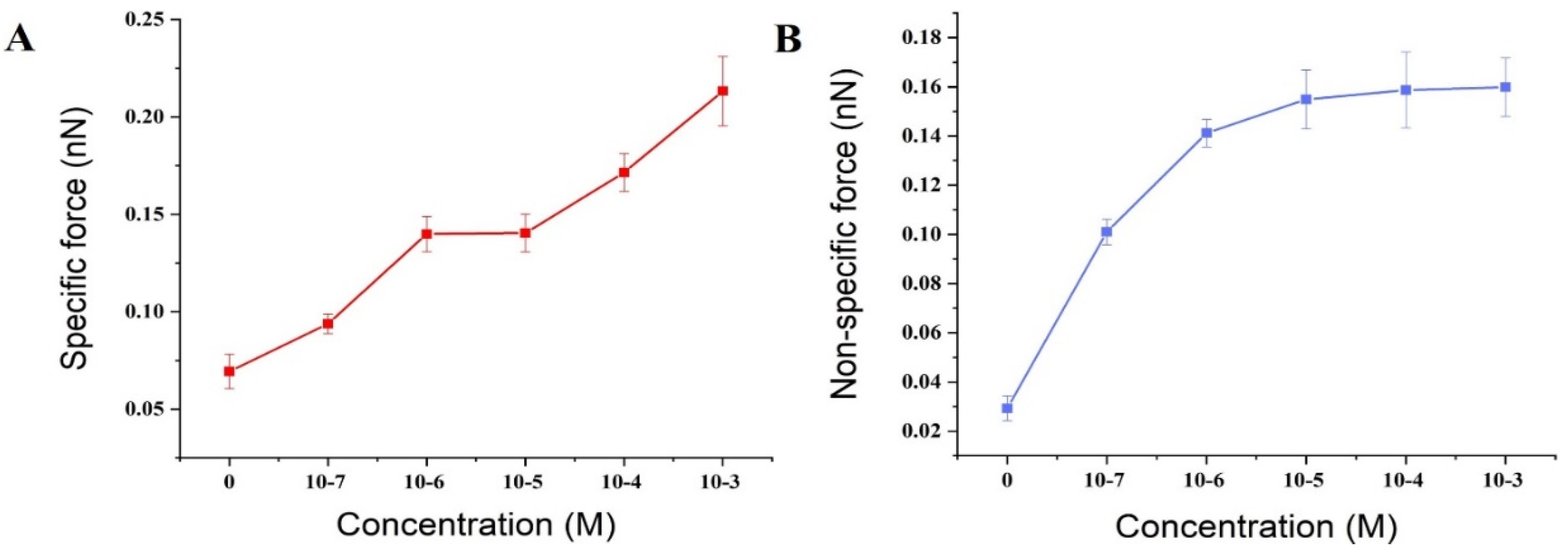
3.2. Effect of Ca2+ Concentration on Specific and Nonspecific Forces
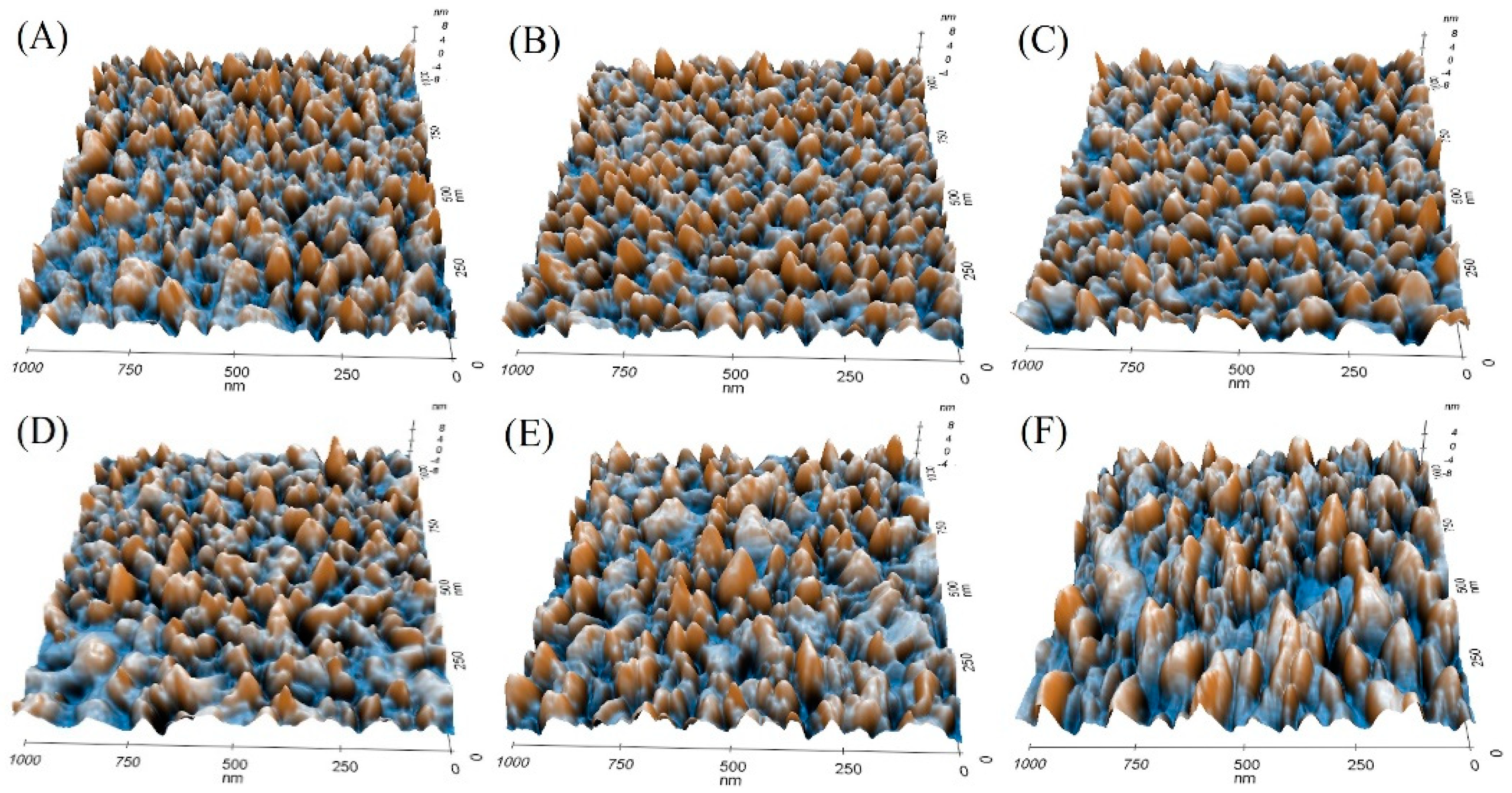
3.3. Calmodulin Imaged by AFM in Solution
4. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Wang, J.H.C.; Thampatty, B.P. An introductory review of cell mechanobiology. Biomech. Model. Mechan. 2006, 5, 1–16. [Google Scholar] [CrossRef] [PubMed]
- Jansen, K.A.; Donato, D.M.; Balcioglu, H.E.; Schmidt, T.; Danen, E.H.; Koenderink, G.H. A guide to mechanobiology: Where biology and physics meet. BBA-Mol. Cell Res. 2015, 1853, 3043–3052. [Google Scholar] [CrossRef] [PubMed]
- Gullapalli, R.R.; Tabouillot, T.; Mathura, R.A.; Dangaria, J.H.; Butler, P.J. Integrated multimodal microscopy, time-resolved fluorescence, and optical-trap rheometry: Toward single molecule mechanobiology. J. Biomed. Opt. 2007, 12, 014012. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Popa, I.; Berkovich, R. Mechanobiology: Protein refolding under force. J. Biomed. Opt. 2018, 2, 687–699. [Google Scholar]
- Meng, W.L.; Zhang, N.; Tang, H.; Tan, Z. Influence of disordered packing pattern on elastic modulus of single-stranded DNA film on substrate. Biomech. Model. Mechan. 2015, 14, 1157–1165. [Google Scholar] [CrossRef]
- Wang, C.; Jin, Y.; Desai, U.R.; Yadavalli, V.K. Investigation of the heparin–thrombin interaction by dynamic force spectroscopy. Biomech. Model. Mechan. 2015, 1850, 1099–1106. [Google Scholar] [CrossRef]
- Hinterdorfer, P.; Dufrêne, Y.F. Detection and localization of single molecular recognition events using atomic force microscopy. Nat. Methods 2006, 3, 347–355. [Google Scholar] [CrossRef]
- Bustamante, C.; Chemla, Y.R.; Forde, N.R.; Izhaky, D. Mechanical processes in biochemistry. Annu. Rev. Biochem. 2004, 73, 705–748. [Google Scholar] [CrossRef]
- Dobson, C.M. Protein folding and misfolding. Nature 2003, 426, 884–890. [Google Scholar] [CrossRef]
- Gething, M.-J.; Sambrook, J. Protein folding in the cell. Nature 1992, 355, 33–45. [Google Scholar] [CrossRef]
- Lv, Z.; Roychaudhuri, R.; Condron, M.M.; Teplow, D.B.; Lyubchenko, Y.L. Mechanism of amyloid β- protein dimerization determined using single− molecule AFM force spectroscopy. Sci. Rep. 2013, 3, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Lv, Z.; Condron, M.M.; Teplow, D.B.; Lyubchenko, Y.L. Nanoprobing of the effect of Cu 2+ cations on misfolding, interaction and aggregation of amyloid β peptide. J. Neuroimmune Pharm. 2013, 8, 262–273. [Google Scholar] [CrossRef] [PubMed]
- Xie, Y.; Wang, J.; Liu, C. Probing the role of metal cations on the aggregation behavior of amyloid β-peptide at a single molecule level by AFM. J. Nanopart. Res. 2016, 18, 287. [Google Scholar] [CrossRef]
- Ju, L.; Chen, Y.; Li, K.; Yuan, Z.; Liu, B.; Jackson, S.P.; Zhu, C. Dual Biomembrane Force Probe enables single-cell mechanical analysis of signal crosstalk between multiple molecular species. Sci. Rep. 2017, 7, 1–12. [Google Scholar] [CrossRef]
- Chen, Y.; Liu, B.; Ju, L.; Hong, J.; Ji, Q.; Chen, W.; Zhu, C. Fluorescence biomembrane force probe: Concurrent quantitation of receptor-ligand kinetics and binding-induced intracellular signaling on a single cell. JoVE 2015, 102, e52975. [Google Scholar] [CrossRef]
- Litvinov, R.I.; Shuman, H.; Bennett, J.S.; Weisel, J.W. Binding strength and activation state of single fibrinogen-integrin pairs on living cells. PNAS 2002, 99, 7426–7431. [Google Scholar] [CrossRef]
- Herbert, K.M.; La Porta, A.; Wong, B.J.; Mooney, R.A.; Neuman, K.C.; Landick, R.; Block, S.M. Sequence-resolved detection of pausing by single RNA polymerase molecules. Cell 2006, 125, 1083–1094. [Google Scholar] [CrossRef]
- Cluzel, P.; Lebrun, A.; Heller, C.; Lavery, R.; Viovy, J.L.; Chatenay, D.; Caron, F. DNA: An extensible molecule. Science 1996, 271, 792–794. [Google Scholar] [CrossRef]
- Ishijima, A.; Kojima, H.; Higuchi, H.; Harada, Y.; Funatsu, T.; Yanagida, T. Multiple-and single-molecule analysis of the actomyosin motor by nanometer-piconewton manipulation with a microneedle: Unitary steps and forces. Biophys. J. 1996, 70, 383–400. [Google Scholar] [CrossRef]
- Revyakin, A.; Ebright, R.H.; Strick, T.R. Single-molecule DNA nanomanipulation: Improved resolution through use of shorter DNA fragments. Nat. Methods 2005, 2, 127–138. [Google Scholar] [CrossRef]
- Charvin, G.; Strick, T.; Bensimon, D.; Croquette, V. Tracking topoisomerase activity at the single-molecule level. Annu. Rev. Biophys. Biomol. Struct. 2005, 34, 201–219. [Google Scholar] [CrossRef] [PubMed]
- Dietz, H.; Berkemeier, F.; Bertz, M.; Rief, M. Anisotropic deformation response of single protein molecules. PNAS 2006, 103, 12724–12728. [Google Scholar] [CrossRef] [PubMed]
- André, E.B.; Litvinov, R.I.; Discher, D.E.; Weisel, J.W. Forced unfolding of coiled-coils in fibrinogen by single-molecule AFM. Biophys. J. 2007, 92, L39–L41. [Google Scholar] [CrossRef] [PubMed]
- Greene, D.N.; Garcia, T.; Sutton, R.B.; Gernert, K.M.; Benian, G.M.; Oberhauser, A.F. Single-molecule force spectroscopy reveals a stepwise unfolding of Caenorhabditis elegans giant protein kinase domains. Biophys. J. 2008, 95, 1360–1370. [Google Scholar] [CrossRef][Green Version]
- Wang, C.; Stanciu, C.E.; Ehrhardt, C.J.; Yadavalli, V.K. The effect of growth temperature on the nanoscale biochemical surface properties of Yersinia pestis. Anal. Bioanal. Chem. 2016, 408, 5585–5591. [Google Scholar] [CrossRef]
- Osada, T.; Itoh, A.; Ikai, A. Mapping of the receptor-associated protein (RAP) binding proteins on living fibroblast cells using an atomic force microscope. Ultramicroscopy 2003, 97, 353–357. [Google Scholar] [CrossRef]
- Lv, Z.; Zhang, Y.; Krasnoslobodsev, A.; Roychaudhuri, R.; Condron, M.; Teplow, D.; Lovas, S.; Shlyakhtenko, L.; Lyubchenko, Y. Misfolding and interactions of Aß proteins: Insight from single molecule experiments and computational analyses. Mol. Neurodegener. 2013, 8, P64. [Google Scholar]
- O’Donoghue, M.B.; Shi, X.; Fang, X.; Tan, W. Single-molecule atomic force microscopy on live cells compares aptamer and antibody rupture forces. Mol. Neurodegener. 2013, 402, 3205–3209. [Google Scholar] [CrossRef]
- Alsteens, D.; Garcia, M.C.; Lipke, P.N.; Dufrêne, Y.F. Force-induced formation and propagation of adhesion nanodomains in living fungal cells. PNAS 2010, 107, 20744–20749. [Google Scholar] [CrossRef]
- Spedden, E.; White, J.D.; Naumova, E.N.; Kaplan, D.L.; Staii, C. Elasticity maps of living neurons measured by combined fluorescence and atomic force microscopy. PNAS 2010, 103, 868–877. [Google Scholar] [CrossRef]
- Taylor, J.N.; Darugar, Q.; Kourentzi, K.; Willson, R.C.; Landes, C.F. Dynamics of an anti-VEGF DNA aptamer: A single-molecule study. Biochem. Biophys. Res. Commun. 2008, 373, 213–218. [Google Scholar] [CrossRef] [PubMed]
- Lv, Z.; Wang, J.; Chen, G. Exploring the energy profile of human IgG/rat anti-human IgG interactions by dynamic force spectroscopy. Protein J. 2012, 31, 425–431. [Google Scholar] [CrossRef] [PubMed]
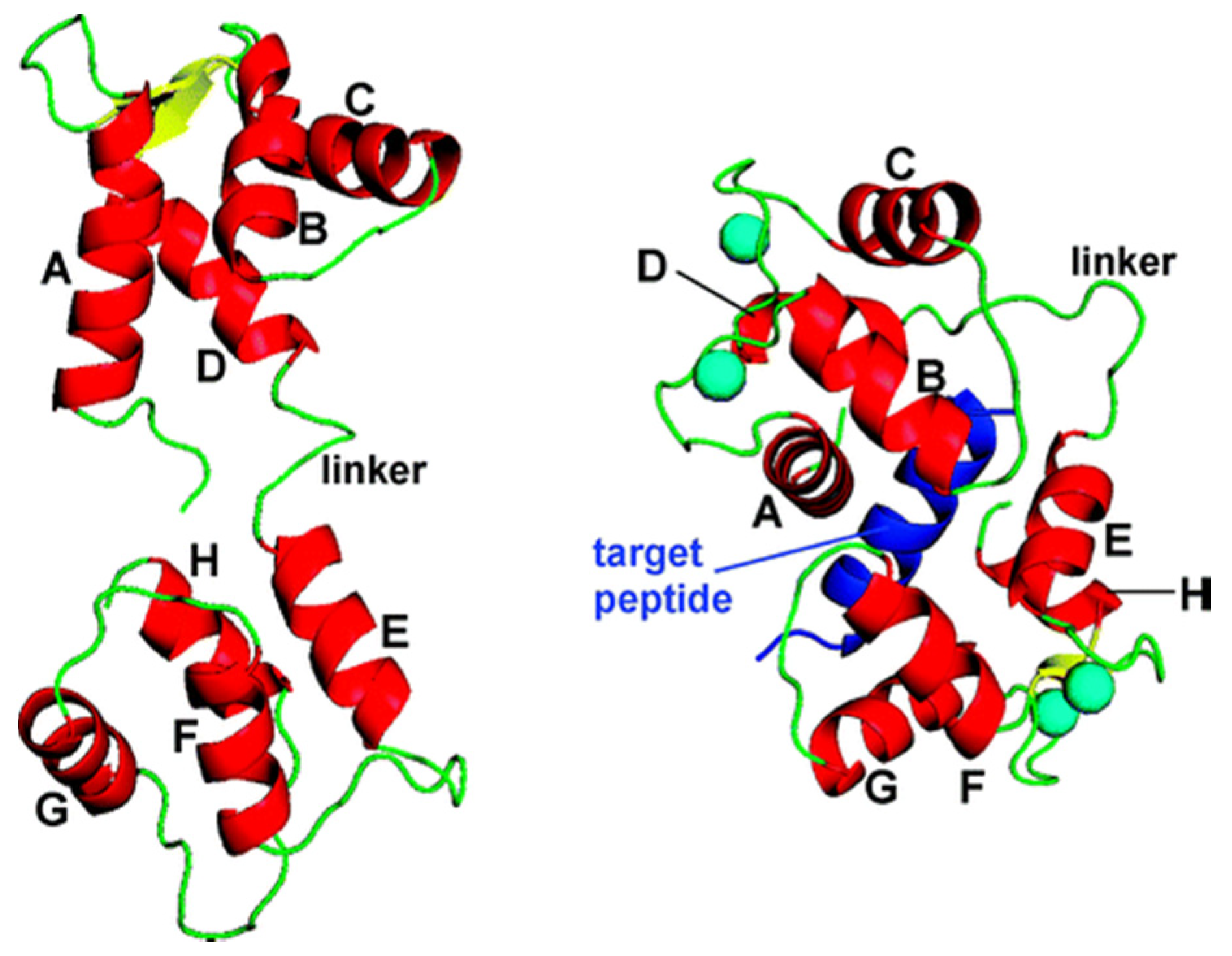
- McCormack, E.; Braam, J. Calmodulins and related potential calcium sensors of Arabidopsis. New Phytol. 2003, 159, 585–598. [Google Scholar] [CrossRef]
- Klee, C.B. Concerted regulation of protein phosphorylation and dephosphorylation by calmodulin. Neurochem. Res. 1991, 16, 1059–1065. [Google Scholar] [CrossRef]
- Li, C.J.; Heim, R.; Lu, P.; Pu, Y.; Tsien, R.Y.; Chang, D.C. Dynamic redistribution of calmodulin in HeLa cells during cell division as revealed by a GFP-calmodulin fusion protein technique. J. Cell. Sci. 1999, 112, 1567–1577. [Google Scholar]
- Jin, L.; Piao, Z.H.; Liu, C.P.; Sun, S.; Liu, B.; Kim, G.R.; Choi, S.Y.; Ryu, Y.; Kee, H.J.; Jeong, M.H. Gallic acid attenuates calcium calmodulin-dependent kinase II-induced apoptosis in spontaneously hypertensive rats. J. Cell. Mol. Med. 2018, 22, 1517–1526. [Google Scholar] [CrossRef]
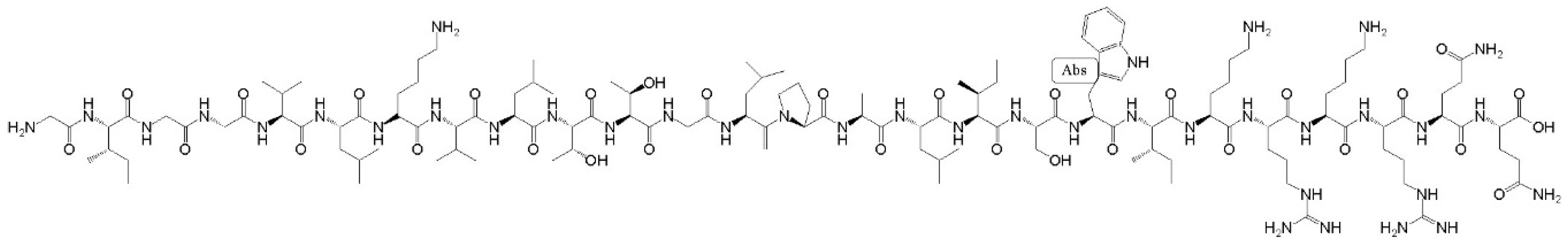
- Pan, J.; Konermann, L. Calcium-induced structural transitions of the calmodulin-melittin system studied by electrospray mass spectrometry: Conformational subpopulations and metal-unsaturated intermediates. Biochemistry 2010, 49, 3477–3486. [Google Scholar] [CrossRef]
- Gajski, G.; Garaj-Vrhovac, V. Melittin: A lytic peptide with anticancer properties. Environ. Toxicol. Phar. 2013, 36, 697–705. [Google Scholar] [CrossRef]
- Katoh, N. Inhibition by melittin of phosphorylation by protein kinase C of annexin I from cow mammary gland. J. Vet. Med. Sci. 2002, 64, 779–783. [Google Scholar] [CrossRef]
- Grandbois, M.; Dufourcq, J.; Salesse, C. Study of the synergistic action of phospholipase A2 and melittin in the hydrolysis of phospholipid monolayers. Thin Solid Films 1996, 284, 743–747. [Google Scholar] [CrossRef]
- Ghosh, A.; Greenberg, M.E. Calcium signaling in neurons: Molecular mechanisms and cellular consequences. Science 1995, 268, 239–247. [Google Scholar] [CrossRef] [PubMed]
- Yamniuk, A.P.; Vogel, H.J. Calmodulin’s flexibility allows for promiscuity in its interactions with target proteins and peptides. Mol. Biotechnol. 2004, 27, 33–57. [Google Scholar] [CrossRef]
- Lazo, J.S.; Hait, W.N.; Kennedy, K.A.; Braun, I.D.; Meandzija, B. Enhanced bleomycin-induced DNA damage and cytotoxicity with calmodulin antagonists. Mol. Pharmacol. 1985, 27, 387–393. [Google Scholar] [PubMed]
- Brokx, R.D.; Lopez, M.M.; Vogel, H.J.; Makhatadze, G.I. Energetics of target peptide binding by calmodulin reveals different modes of binding. J. Biol. Chem. 2001, 276, 14083–14091. [Google Scholar] [CrossRef]
- Schulz, D.M.; Ihling, C.; Clore, G.M.; Sinz, A. Mapping the Topology and Determination of a Low-Resolution Three-Dimensional Structure of the Calmodulin−Melittin Complex by Chemical Cross-Linking and High-Resolution FTICRMS: Direct Demonstration of Multiple Binding Modes. Biochemistry 2004, 43, 4703–4715. [Google Scholar] [CrossRef][Green Version]
- Noguchi, H.; Adachi, T.; Nakatomi, A.; Yazawa, M.; Uosaki, K. Biofunctionality of Calmodulin Immobilized on Gold Surface Studied by Surface-Enhanced Infrared Absorption Spectroscopy: Ca2+-Induced Conformational Change and Binding to a Target Peptide. J. Phys. Chem. C 2016, 120, 16035–16041. [Google Scholar] [CrossRef]
- Cinar, S.; Czeslik, C. Inhibitor and peptide binding to calmodulin characterized by high pressure Fourier transform infrared spectroscopy and Förster resonance energy transfer. BBA Proteins Proteom 2018, 1866, 617–623. [Google Scholar] [CrossRef]
- Newman, R.A.; Van Scyoc, W.S.; Sorensen, B.R.; Jaren, O.R.; Shea, M.A. Interdomain cooperativity of calmodulin bound to melittin preferentially increases calcium affinity of sites I and II. Proteins 2008, 71, 1792–1812. [Google Scholar] [CrossRef]
- Otosu, T.; Nishimoto, E.; Yamashita, S. Spectrally and Time-resolved Fluorescence Spectroscopic Study on Melittin–Calmodulin Interaction. J. Biochem. 2007, 142, 655–661. [Google Scholar] [CrossRef]
- Wang, C.; Wang, J.; Deng, L. Evaluating interaction forces between BSA and rabbit anti-BSA in sulphathiazole sodium, tylosin and levofloxacin solution by AFM. Nanoscale Res. Lett. 2011, 6, 579. [Google Scholar] [CrossRef]
- Wang, Y.; Wang, J.; Huang, S.; Liu, C.; Fu, Y. Evaluating the effect of aminoglycosides on the interaction between bovine serum albumins by atomic force microscopy. Int. J. Biol. Macromol. 2019, 134, 28–35. [Google Scholar] [CrossRef] [PubMed]
- Lv, Z.; Wang, J.; Chen, G.; Deng, L. Probing specific interaction forces between human IgG and rat anti-human IgG by self-assembled monolayer and atomic force microscopy. Nanoscale Res. Lett. 2010, 5, 1032–1038. [Google Scholar] [CrossRef] [PubMed]
- Lo, Y.S.; Huefner, N.D.; Chan, W.S.; Stevens, F.; Harris, J.M.; Beebe, T.P. Specific Interactions between Biotin and Avidin Studied by Atomic Force Microscopy Using the Poisson Statistical Analysis Method. Langmuir 1999, 15, 1373–1382. [Google Scholar] [CrossRef]
- Liu, W.; Parpura, V. Single molecule probing of SNARE proteins by atomic force microscopy. Ann. N. Y. Acad. Sci. 2009, 1152, 113–120. [Google Scholar] [CrossRef] [PubMed]
- Moorthy, A.K.; Gopal, B.; Satish, P.R.; Bhattacharya, S.; Bhattacharya, A.; Murthy, M.R.N.; Surolia, A. Thermodynamics of target peptide recognition by calmodulin and a calmodulin analogue: Implications for the role of the central linker. FEBS Lett. 1999, 461, 19–24. [Google Scholar] [CrossRef]
- Kataoka, M.; Head, J.F.; Seaton, B.A.; Engelman, D.M. Melittin binding causes a large calcium-dependent conformational change in calmodulin. PNAS 1989, 86, 6944–6948. [Google Scholar] [CrossRef]
- Seeholzer, S.H.; Cohn, M.; Wand, A.J.; Crespi, H.L.; Putkey, J.A.; Means, A.R. 1H NMR structural studies of a complex of melittin with perdeuterated calmodulin. In Calcium-Binding Proteins in Health and Disease; Academic Press: Cambridge, MA, USA, 1987; pp. 360–371. [Google Scholar]
- Itakura, M.; Iio, T. Static and Kinetic Studies of Calmodulin and Melittin Complex. J. Biochem 1992, 112, 183–191. [Google Scholar] [CrossRef]
- Bandyopadhyay, S.; Ghosh, S.K. Goat testis calmodulin: Purification and physicochemical characterization. J. Protein Chem 1990, 9, 603–611. [Google Scholar] [CrossRef]
- Wyttenbach, T.; Grabenauer, M.; Thalassinos, K.; Scrivens, J.H.; Bowers, M.T. The Effect of Calcium Ions and Peptide Ligands on the Relative Stabilities of the Calmodulin Dumbbell and Compact Structures. Phys. Chem. B 2010, 114, 437–447. [Google Scholar] [CrossRef]
- Jinesh, K.B.; Frenken, J.W.M. Capillary condensation in atomic scale friction: How water acts like a glue. Phys Rev. Lett 2006, 96, 166103. [Google Scholar] [CrossRef]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Set | Mean, μm (nN) | Number of Bonds, μn | |
|---|---|---|---|
| 1 | 1.01878 | 0.00027 | 7.4 |
| 2 | 1.1698 | 0.0014 | 8.7 |
| 3 | 1.24122 | 0.00195 | 11.6 |
| 4 | 1.3 | 0.00243 | 12.4 |
| 5 | 1.38184 | 0.00324 | 13.2 |
| 6 | 1.55082 | 0.00485 | 16.4 |
| Sample | Ra (nm) |
|---|---|
| The control | 1.616 |
| 10−7 M Ca2+ | 1.641 |
| 10−6 M Ca2+ | 1.789 |
| 10−5 M Ca2+ | 1.844 |
| 10−4 M Ca2+ | 1.849 |
| 10−3 M Ca2+ | 1.858 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Huang, S.; Wang, J.; Sun, H.; Fu, Y.; Wang, Y. Probing Changes in Ca2+-Induced Interaction Forces between Calmodulin and Melittin by Atomic Force Microscopy. Micromachines 2020, 11, 906. https://doi.org/10.3390/mi11100906
Huang S, Wang J, Sun H, Fu Y, Wang Y. Probing Changes in Ca2+-Induced Interaction Forces between Calmodulin and Melittin by Atomic Force Microscopy. Micromachines. 2020; 11(10):906. https://doi.org/10.3390/mi11100906
Chicago/Turabian StyleHuang, Sheng, Jianhua Wang, Heng Sun, Yuna Fu, and Yan Wang. 2020. "Probing Changes in Ca2+-Induced Interaction Forces between Calmodulin and Melittin by Atomic Force Microscopy" Micromachines 11, no. 10: 906. https://doi.org/10.3390/mi11100906
APA StyleHuang, S., Wang, J., Sun, H., Fu, Y., & Wang, Y. (2020). Probing Changes in Ca2+-Induced Interaction Forces between Calmodulin and Melittin by Atomic Force Microscopy. Micromachines, 11(10), 906. https://doi.org/10.3390/mi11100906