Thrust and Hydrodynamic Efficiency of the Bundled Flagella

 ,
,  and
and {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Methodology
2.1. Experimental Setup and Flagella Configurations
2.2. Particle Image Velocimetry (PIV)
2.3. Volumetric PIV Reconstruction Protocol for Thrust and Dissipation
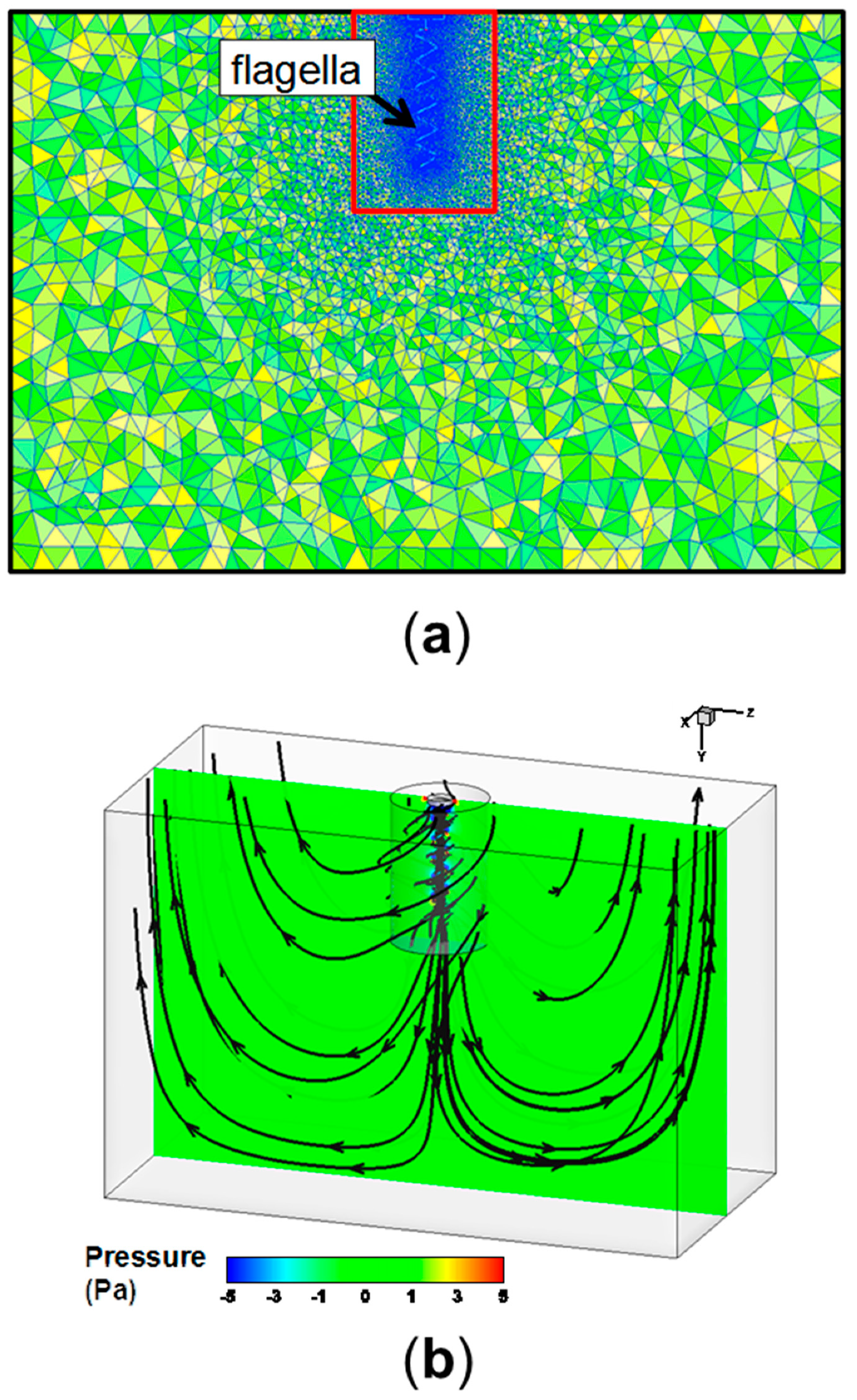
2.4. Computational Fluid Dynamics (CFD) for Experimental Planning
3. Results
3.1. Single Flagellum Flow Field
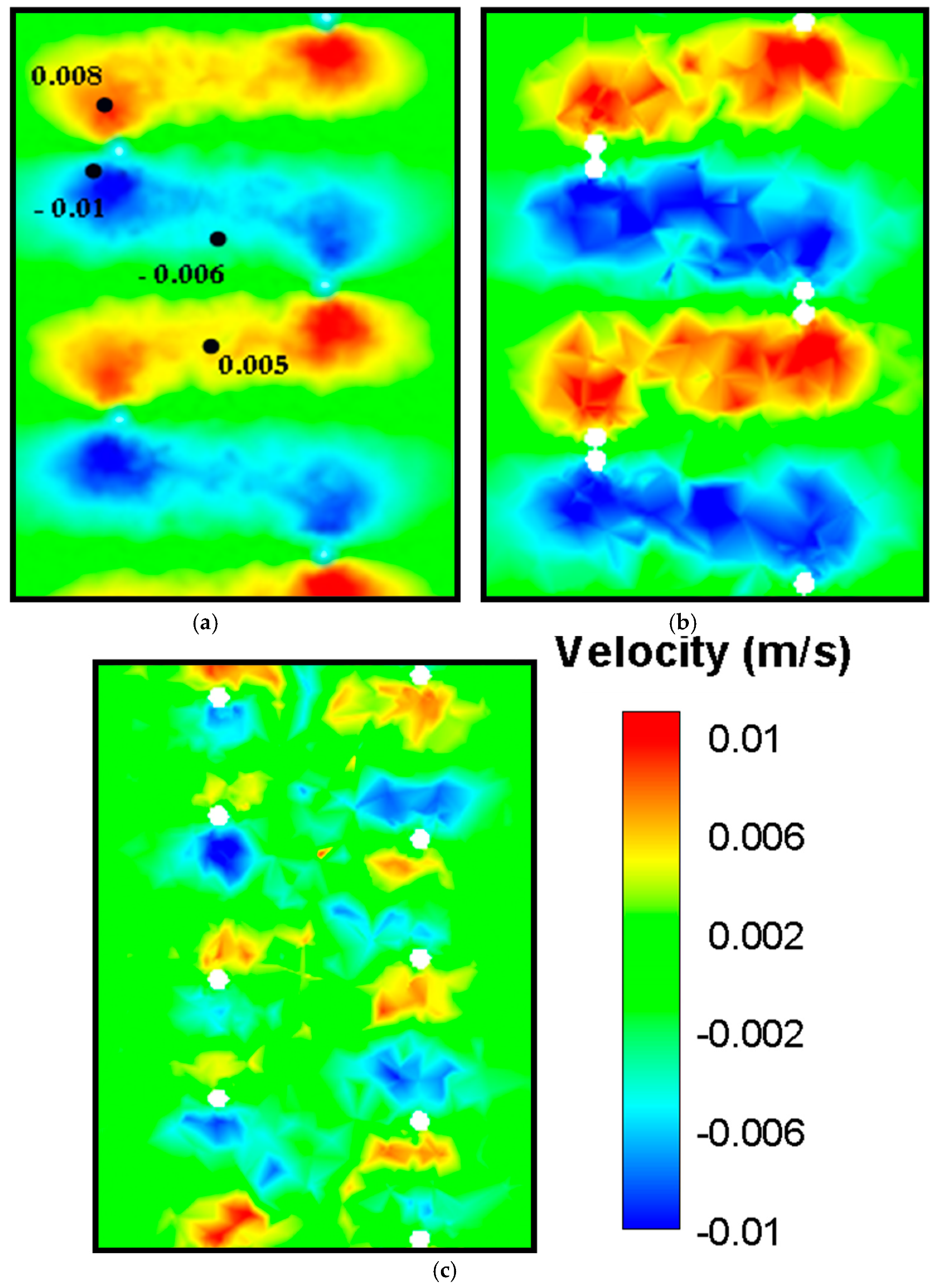
3.2. Velocity Fields Generated by Double-helix Flagella Configurations
3.3. Thrust Force Measured from Volumetric PIV vs. Load Cell
3.4. Limits of Resistive Force Theory (RFT) Predictions
3.5. Thrust Variation Along the Axis of a Rigid and Finite Flagellum
3.6. Double-Helix (Bundling States) Thrust Force Results
3.7. Hydrodynamic Efficiency
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Appendix A. CFD Methodology

References
- Taylor, G. The action of waving cylindrical tails in propelling microscopic organisms. Proc. R. Soc. Lond. Ser. A Math. Phys. Sci. 1952, 211, 225–239. [Google Scholar]
- Lauga, E.; Powers, T.R. The hydrodynamics of swimming microorganisms. Rep. Prog. Phys. 2009, 72, 096601. [Google Scholar] [CrossRef]
- Peyer, K.E.; Li, Z.; Nelson, B.J. Bio-inspired magnetic swimming microrobots for biomedical applications. Nanoscale 2013, 5, 1259–1272. [Google Scholar] [CrossRef] [PubMed]
- Martel, S. Swimming microorganisms acting as nanorobots versus artificial nanorobotic agents: A perspective view from an historical retrospective on the future of medical nanorobotics in the largest known three-dimensional biomicrofluidic networks. Biomicrofluidics 2016, 10, 021301. [Google Scholar] [CrossRef] [PubMed]
- Singh, A.V.; Hosseinidoust, Z.; Park, B.W.; Yasa, O.; Sitti, M. Microemulsion-based soft bacteria-driven microswimmers for active cargo delivery. ACS Nano 2017, 11, 9759–9769. [Google Scholar] [CrossRef] [PubMed]
- Klumpp, S.; Lefèvre, C.T.; Bennet, M.; Faivre, D. Swimming with magnets: From biological organisms to synthetic devices. Phys. Rep. 2018. [Google Scholar] [CrossRef]
- Alapan, Y.; Yasa, O.; Yigit, B.; Yasa, I.C.; Erkoc, P.; Sitti, M. Microrobotics and Microorganisms: Biohybrid Autonomous Cellular Robots. Annu. Rev. Control Robot. Auton. Syst. 2019, 2, 205–230. [Google Scholar] [CrossRef]
- Sitti, M. Miniature devices: Voyage of the microrobots. Nature 2009, 458, 1121. [Google Scholar] [CrossRef]
- Sitti, M. Mobile Microrobotics; MIT Press: Cambridge, MA, USA, 2017. [Google Scholar]
- Hu, W.; Lum, G.Z.; Mastrangeli, M.; Sitti, M. Small-scale soft-bodied robot with multimodal locomotion. Nature 2018, 554, 81. [Google Scholar] [CrossRef] [PubMed]
- Park, B.W.; Zhuang, J.; Yasa, O.; Sitti, M. Multifunctional bacteria-driven microswimmers for targeted active drug delivery. ACS Nano 2017, 11, 8910–8923. [Google Scholar] [CrossRef]
- Zhuang, J.; Park, B.W.; Sitti, M. Propulsion and Chemotaxis in Bacteria-Driven Microswimmers. Adv. Sci. 2017, 4, 1700109. [Google Scholar] [CrossRef] [PubMed]
- Stanton, M.M.; Park, B.W.; Vilela, D.; Bente, K.; Faivre, D.; Sitti, M.; Sánchez, S. Magnetotactic bacteria powered biohybrids target E. coli biofilms. ACS Nano 2017, 11, 9968–9978. [Google Scholar] [CrossRef] [PubMed]
- Niu, R.; Kreissl, P.; Brown, A.T.; Rempfer, G.; Botin, D.; Holm, C.; de Graaf, J. Microfluidic pumping by micromolar salt concentrations. Soft Matter 2017, 13, 1505–1518. [Google Scholar] [CrossRef] [PubMed]
- Niu, R.; Fischer, A.; Palberg, T.; Speck, T. Dynamics of Binary Active Clusters Driven by Ion-Exchange Particles. ACS Nano 2018, 12, 10932–10938. [Google Scholar] [CrossRef] [PubMed]
- Yang, M.; Wysocki, A.; Ripoll, M. Hydrodynamic simulations of self-phoretic microswimmers. Soft Matter 2014, 10, 6208–6218. [Google Scholar] [CrossRef] [PubMed]
- Howard, C.B.; Anderson, R.A. Bacteria swim by rotating their flagellar filaments. Nature 1973, 245, 380–382. [Google Scholar]
- Lowe, G.; Meister, M.; Berg, H.C. Rapid rotation of flagellar bundles in swimming bacteria. Nature 1987, 325, 637–640. [Google Scholar] [CrossRef]
- Purcell, E. Life at low Reynolds numbers. Am. J. Phys. 1976, 45, 3–11. [Google Scholar] [CrossRef]
- Cortez, R. The method of Regularized Stokeslets. SIAM J. Sci. Comput. 2001, 23, 204–1225. [Google Scholar] [CrossRef]
- Cortez, R.; Fauci, L.; Medovikov, A. The method of Regularized Stokeslets in three dimensions: Analysis, validation, and application to helical swimming. Phys. Fluids 2005, 17, 031504. [Google Scholar] [CrossRef]
- Lighthill, J. Helical distributions of Stokeslets. J. Eng. Math. 1996, 30, 35–78. [Google Scholar] [CrossRef]
- Rodenborn, B.; Chen, C.H.; Swinney, H.L.; Liu, B.; Zhang, H.P. Propulsion of microorganisms by a helical flagellum. Proc. Natl. Acad. Sci. USA 2013, 110, E338–E347. [Google Scholar] [CrossRef] [PubMed]
- Flores, H.; Lobaton, E.; Méndez-Diez, S.; Tlupova, S.; Cortez, R. A study of bacterial flagellar bundling. Bull. Math. Biol. 2005, 67, 137–168. [Google Scholar] [CrossRef] [PubMed]
- Turner, L.; Ryu, W.S.; Berg, H.C. Real-time imaging of fluorescent flagellar filaments. J. Bacteriol. 2000, 182, 2793–2801. [Google Scholar] [CrossRef] [PubMed]
- Shum, H.; Gaffney, E.A. Hydrodynamic analysis of flagellated bacteria swimming in corners of rectangular channels. Phys. Rev. E 2015, 92, 063016. [Google Scholar] [CrossRef] [PubMed]
- Ishimoto, K. Hydrodynamic evolution of sperm swimming: Optimal flagella by a genetic algorithm. J. Theor. Biol. 2016, 399, 166–174. [Google Scholar] [CrossRef] [PubMed]
- Ding, Y.; Qiu, F.; Casadevall i Solvas, X.; Chiu, F.; Nelson, B.; de Mello, A. Microfluidic-based droplet and cell manipulations using artificial bacterial flagella. Micromachines 2016, 7, 25. [Google Scholar] [CrossRef]
- Bet, B.; Boosten, G.; Dijkstra, M.; van Roij, R. Efficient shapes for microswimming: From three-body swimmers to helical flagella. J. Chem. Phys. 2017, 146, 084904. [Google Scholar] [CrossRef] [PubMed]
- Shum, H. Microswimmer Propulsion by Two Steadily Rotating Helical Flagella. Micromachines 2019, 10, 65. [Google Scholar] [CrossRef]
- Gray, J.; Hancock, G. The propulsion of sea urchin spermatoza. J. Exp. Biol. 1955, 32, 802–814. [Google Scholar]
- Bayly, P.V.; Lewis, B.L.; Ranz, E.C.; Okamoto, R.J.; Pless, R.B.; Dutcher, S.K. Propulsive forces on the flagellum during locomotion of Chlamydomonas reinhardtii. Biophys. J. 2011, 100, 2716–2725. [Google Scholar] [CrossRef] [PubMed]
- Blum, J.; Lubliner, J. Biophysics of flagellar motility. Annu. Rev. Biophys. Bioeng. 1973, 2, 181–219. [Google Scholar] [CrossRef] [PubMed]
- Brokaw, C. Non-sinusoidal bending waves of sperm flagella. J. Exp. Biol. 1965, 43, 155–169. [Google Scholar] [PubMed]
- Brokaw, C. Bending moments in free-swimming flagella. J. Exp. Biol. 1970, 53, 445–464. [Google Scholar] [PubMed]
- Brokaw, C. Computer simulation of flagellar movement: I. Demonstration of stable bend propagation and bend initiation by the Sliding Filament Model. Biophys. J. 1972, 12, 564–586. [Google Scholar] [CrossRef]
- Chattopadhyay, S.; Moldovan, R.; Yeung, C.; Wu, X.L. Swimming efficiency of bacterium Escherichia coli. PNAS 2006, 103, 13712–13717. [Google Scholar] [CrossRef] [PubMed]
- Darnton, N.C.; Turner, L.; Rojevsky, S.; Berg, H.C. On torque and tumbling in swimming Escherichia coli. J. Bacteriol. 2007, 189, 1756–1764. [Google Scholar] [CrossRef]
- Lauga, E.; DiLuzio, W.R.; Whitesides, G.M.; Stone, H.A. Swimming in circles: motion of bacteria near solid boundaries. Biophys. J. 2006, 90, 400–412. [Google Scholar] [CrossRef]
- Vogel, R.; Stark, H. Force-extension curves of bacterial flagella. Eur. Phys. J. E Soft Matter Biol. Phys. 2010, 33, 259–271. [Google Scholar] [CrossRef]
- Vogel, R.; Stark, H. Motor-driven bacterial flagella and buckling instabilities. Eur. Phy. J. E Soft Matter Biol. Phys. 2012, 35, 1–15. [Google Scholar] [CrossRef]
- Chattopadhyay, S.; Wu, X.L. The Effect of Long-Range Hydrodynamic Interaction on the Swimming of a Single Bacterium. Biophys. J. 2009, 96, 2023–2028. [Google Scholar] [CrossRef] [PubMed]
- Keaveny, E.; Maxey, M. Spiral swimming of an artificial micro-swimmer. J. Fluid Mech. 2008, 598, 293–319. [Google Scholar] [CrossRef]
- Espinosa-Garcia, J.; Lauga, E.; Zenit, R. Fluid elasticity increases the locomotion of flexible swimmers. Phys. Fluids 2013, 25, 031701. [Google Scholar] [CrossRef]
- Zhong, S.; Moored, K.W.; Pinedo, V.; Garcia-Gonzalez, J.; Smits, A.J. The flow field and axial thrust generated by a rotating rigid helix at low Reynolds numbers. Exp. Therm. Fluid Sci. 2013, 46, 1–7. [Google Scholar] [CrossRef]
- Hu, J.; Yang, M.; Gompper, G.; Winkler, R.G. Modelling the mechanics and hydrodynamics of swimming E. coli. Soft Matter 2015, 11, 7867–7876. [Google Scholar] [CrossRef] [PubMed]
- Mears, P.J.; Koirala, S.; Rao, C.V.; Golding, I.; Chemla, Y.R. Escherichia coli swimming is robust against variations in flagellar number. Elife 2014, 3, e01916. [Google Scholar] [CrossRef] [PubMed]
- Kanehl, P.; Ishikawa, T. Fluid mechanics of swimming bacteria with multiple flagella. Phys. Rev. E 2014, 89, 042704. [Google Scholar] [CrossRef]
- Maniyeri, R.; Kang, S. Numerical study on bacterial flagellar bundling and tumbling in a viscous fluid using an immersed boundary method. Appl. Math.Model. 2014, 38, 3567–3590. [Google Scholar] [CrossRef]
- Reigh, S.Y.; Winkler, R.G.; Gompper, G. Synchronization and bundling of anchored bacterial flagella. Soft Matter 2012, 8, 4363–4372. [Google Scholar] [CrossRef]
- Kim, M.; Bird, J.C.; Van Parys, A.J.; Breuer, K.S.; Powers, T.R. A Macroscopic Scale Model of Bacterial Flagellar Bundling. PNAS 2003, 100, 15481–15485. [Google Scholar] [CrossRef]
- Kim, M.J.; Kim, M.J.; Bird, J.C.; Park, J.; Powers, T.R.; Breuer, K.S. Particle image velocimetry experiments on a macro-scale model for bacterial flagellar bundling. Exp. Fluids 2004, 37, 782–788. [Google Scholar] [CrossRef]
- Frakes, D.H.; Pekkan, K.; Dasi, L.P.; Kitajima, H.D.; De Zelicourt, D.; Leo, H.L.; Yoganathan, A.P. Modified control grid interpolation for the volumetric reconstruction of fluid flows. Exp. Fluids 2008, 45, 987–997. [Google Scholar] [CrossRef] [PubMed]
- Behkam, B.; Sitti, M. Modeling and Testing of a Biomimetic Flagellar Propulsion Method for Microscale Biomedical Swimming Robots. In Proceedings of the IEEE/ASME International Conference on Advanced Intelligent Mechatronics, Monterey, CA, USA, 15 January 2005. [Google Scholar]
- Behkam, B.; Sitti, M. Design methodology for biomimetic propulsion of miniature swimming robots. Transactions of the ASME. J. Dyn. Syst. Meas. Control 2006, 128, 36–43. [Google Scholar] [CrossRef]
- Behkam, B.; Sitti, M. Bacterial flagella-based propulsion and on/off motion control of microscale objects. Appl. Phys. Lett. 2007, 90, 023902. [Google Scholar] [CrossRef]
- Raffel, M.; Willert, C.; Kompenhas, J. Particle Image Velocimetry; Springer: Berlin, Germany, 1998. [Google Scholar]
- Adrian, R. Dynamic range of velocity and spatial resolution of particle image velocimetry. Meas. Sci. Technol. 1997, 8, 1393–1398. [Google Scholar] [CrossRef]
- Kleinstereur, C. Engineering Fluid Dynamics; Cambridge University Press: Cambridge, UK, 1997. [Google Scholar]
- Anderson, E.J.; Grosenbaugh, M.A. Jet flow in steadily swimming adult squid. J. Exp. Biol. 2005, 208, 1125–1146. [Google Scholar] [CrossRef]
- Pierrakos, O.; Vlachos, P. The Effect of Vortex Formation on Mitral Valve Efficiency. J. Biomech. Eng. 2006, 128, 527–539. [Google Scholar] [CrossRef]
- Biewener, A. Animal Locomotion; Oxford University Press: Oxford, UK, 2003. [Google Scholar]
- Bejan, A. Entropy Generation through Heat and Fluid Flow; John Wiley & Sons: Hoboken, NJ, USA, 1982. [Google Scholar]
- Childress, S. Mechanics of Swimming and Flying; Cambridge University Press: Cambridge, UK, 1981. [Google Scholar]
- Danis, U.; Sitti, M.; Pekkan, K. Particle image velocimetry and thrust of flagellar micro propulsion systems. In Proceedings of the APS Division of Fluid Dynamics Meeting Abstracts, San Antonio, TX, USA, 23–25 November 2008. [Google Scholar]
- Aubin, J.; Fletcher, D.F.; Xuereb, C. Modeling turbulent flow in stirred tanks with CFD: the influence of the modeling approach, turbulence model and numerical scheme. Exp. Therm. Fluid Sci. 2004, 28, 431–445. [Google Scholar] [CrossRef]
- Brucato, A.; Ciofalo, M.; Grisafi, F.; Micale, G. Numerical prediction of flow fields in baffled stirred vessels: A comparison of alternative modelling approaches. Chem. Eng. Sci. 1998, 53, 3653–3684. [Google Scholar] [CrossRef]
- Bujalski, W.; Jaworski, Z.; Nienow, W. CFD Study of Homogenization with Dual Rushton Turbines—Comparison with Experimental Results Part II: The Multiple Reference Frame. Chem. Eng. Res. Des. 2002, 80, 97–104. [Google Scholar] [CrossRef]











© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Danis, U.; Rasooli, R.; Chen, C.-Y.; Dur, O.; Sitti, M.; Pekkan, K. Thrust and Hydrodynamic Efficiency of the Bundled Flagella. Micromachines 2019, 10, 449. https://doi.org/10.3390/mi10070449
Danis U, Rasooli R, Chen C-Y, Dur O, Sitti M, Pekkan K. Thrust and Hydrodynamic Efficiency of the Bundled Flagella. Micromachines. 2019; 10(7):449. https://doi.org/10.3390/mi10070449
Chicago/Turabian StyleDanis, Umit, Reza Rasooli, Chia-Yuan Chen, Onur Dur, Metin Sitti, and Kerem Pekkan. 2019. "Thrust and Hydrodynamic Efficiency of the Bundled Flagella" Micromachines 10, no. 7: 449. https://doi.org/10.3390/mi10070449
APA StyleDanis, U., Rasooli, R., Chen, C.-Y., Dur, O., Sitti, M., & Pekkan, K. (2019). Thrust and Hydrodynamic Efficiency of the Bundled Flagella. Micromachines, 10(7), 449. https://doi.org/10.3390/mi10070449