
Analysis of the Masked Metabolite of Deoxynivalenol and Fusarium Resistance in CIMMYT Wheat Germplasm


Abstract
1. Introduction
2. Results
2.1. FHB Progression
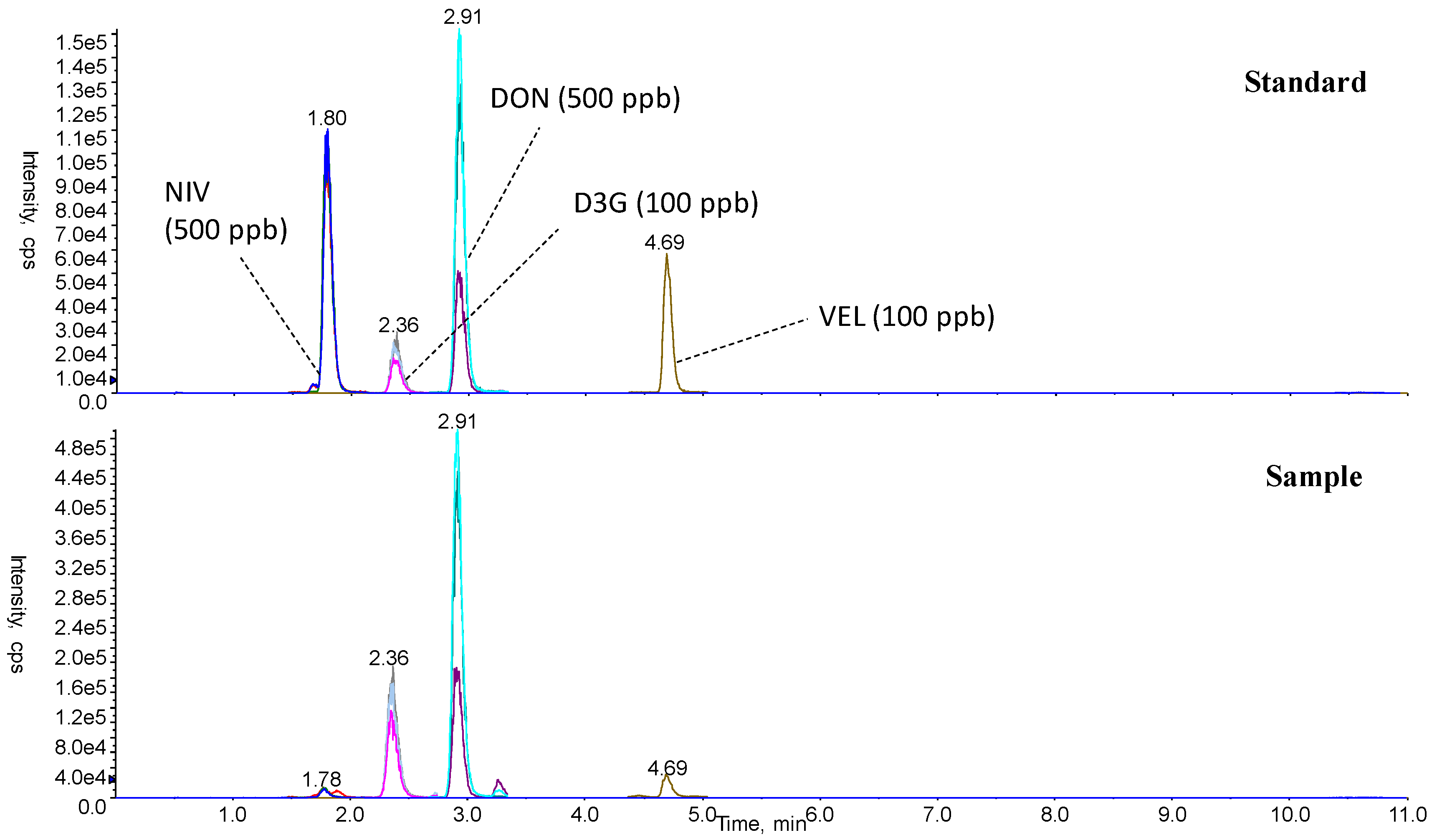
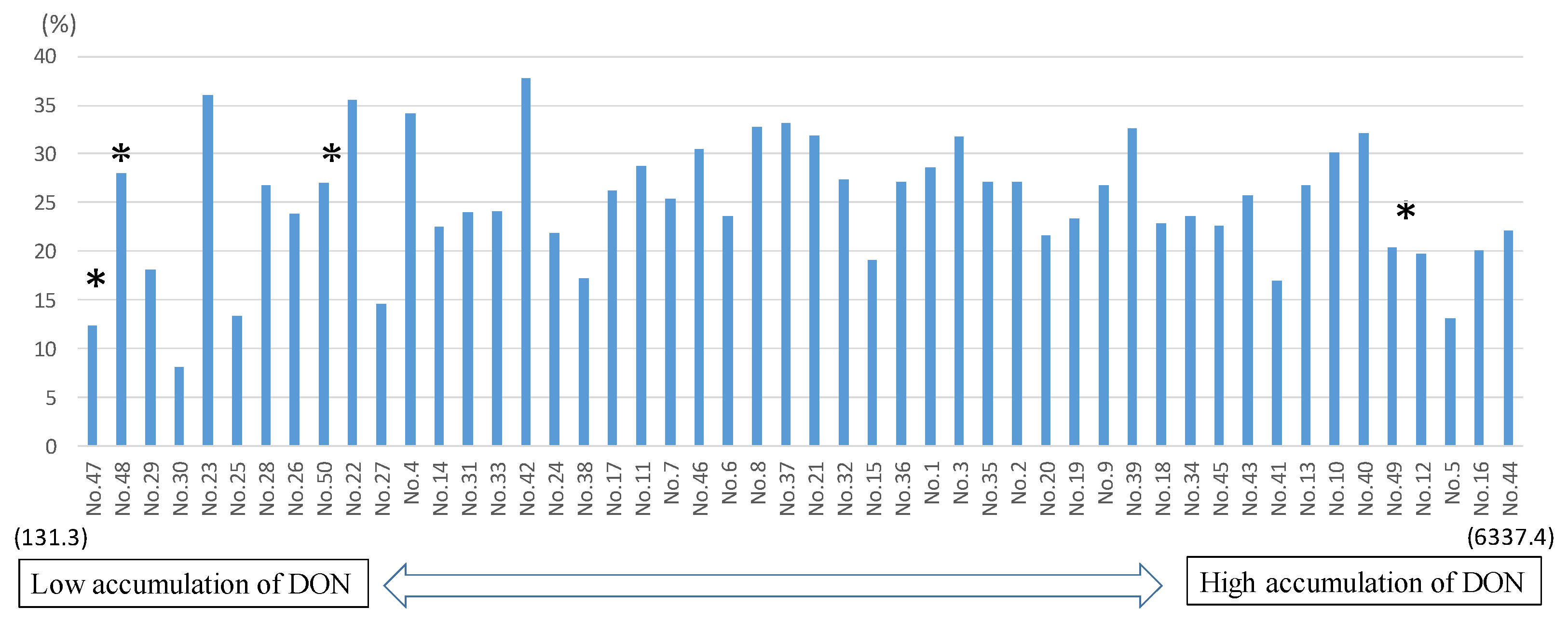
2.2. Determination of Mycotoxins and ERG
2.3. Correlations among Toxin-Related Traits
3. Discussion
4. Conclusions
5. Materials and Methods
5.1. Plant Materials
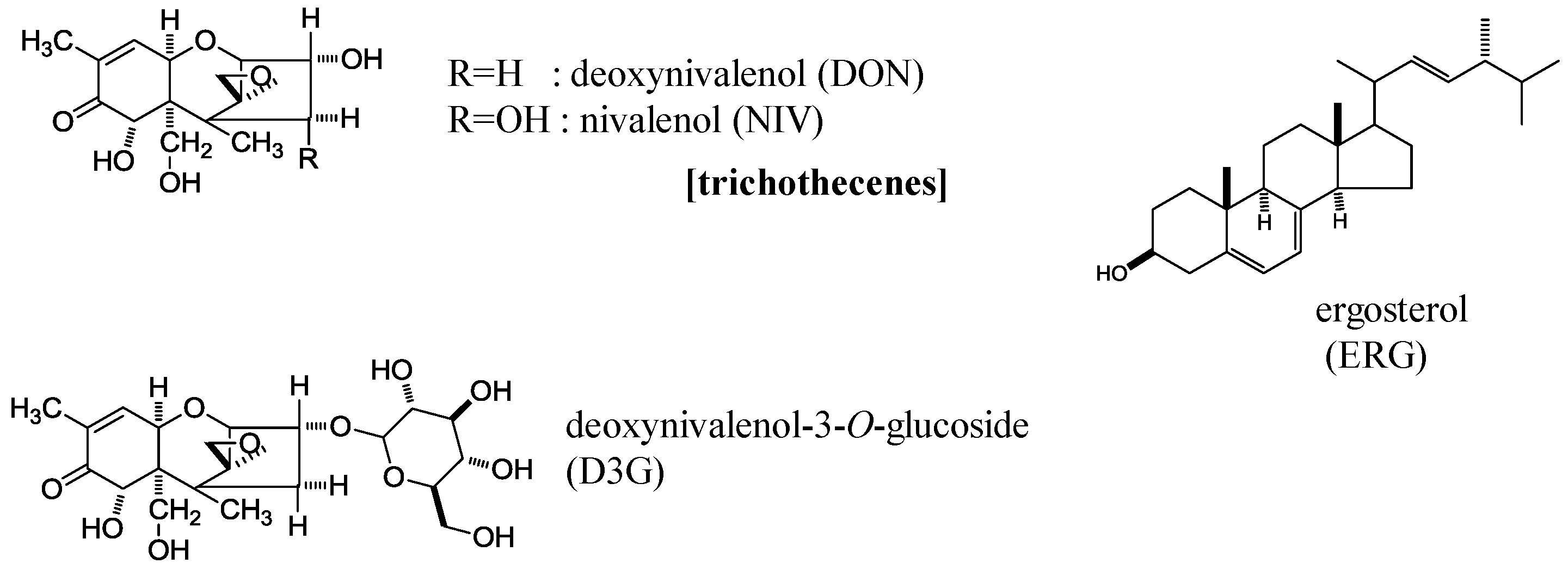
5.2. Chemicals and Standards
5.3. Fungal Strains for Inoculation
5.4. Field Trials and FHB Evaluation
5.5. Preparation of Stock and Working Solutions for Chemical Analysis
5.6. Extraction and Purification of Mycotoxins and ERG
5.7. Analysis of Mycotoxins and ERG
Supplementary Materials
Acknowledgments
Author Contributions
Conflicts of Interest
Abbreviations
References
- Duveiller, E.; Singh, P.K.; Mezzalama, M.; Singh, R.P.; Dababat, A.A. Wheat Diseases and Pests: A Guide for Field Identification; CIMMYT: Mexico DF, Mexico, 2012; 138p, Available online: http://repository.cimmyt.org/xmlui/handle/10883/1115 (accessed on 27 July 2017).
- Gilbert, J.; Haber, S. Overview of some recent research developments in fusarium head blight of wheat. Can. J. Plant Pathol. 2013, 35, 149–174. [Google Scholar] [CrossRef]
- Zhang, X.; Halder, J.; White, R.P.; Hughes, D.J.; Ye, Z.; Wang, C.; Xu, R.; Gan, B.; Fitt, B.D. Climate change increases risk of fusarium ear blight on wheat in central china. Ann. Appl. Biol. 2014, 164, 384–395. [Google Scholar] [CrossRef]
- Yoshizawa, T.; Morooka, N. Deoxynivalenol and its monoacetate: New trichothecene mycotoxin from Fusarium roseum and moldy barley. Agric. Biol. Chem. 1973, 37, 2933–2934. [Google Scholar] [CrossRef]
- Kushiro, M. Effects of milling and cooking processes on the deoxynivalenol content in wheat. Int. J. Mol. Sci. 2008, 9, 2127–2145. [Google Scholar] [CrossRef] [PubMed]
- Joint FAO/WHO Expert Committee on Food Additives (JECFA). WHO Food Additives Series 47: Safety Evaluation of Certain Mycotoxins in Food; World Health Organization: Geneva, Switzerland, 2001; pp. 419–555. Available online: https://www.efsa.europa.eu/en/efsajournal/pub/4655 (accessed on 27 July 2017).
- Food Safety Commission of Japan (FSCJ). Risk Assessment Report: Deoxynivalenol and Nivalenol (Mycotoxin). Risk Assessment Report—Veterinary Medicines FS/872/2010. Available online: https://www.fsc.go.jp/fsciis/evaluationDocument/show/kya20101118001 (accessed on 27 July 2017).
- Paul, P.A.; Lipps, P.E.; Madden, L.V. Relationship between visual estimates of fusarium head blight intensity and deoxynivalenol accumulation in harvested wheat grain: A meta-analysis. Phytopathology 2005, 95, 1225–1236. [Google Scholar] [CrossRef] [PubMed]
- Nakagawa, H. Mycotoxin glucosides (Masked mycotoxins). In Glucosides: Sources, Applications, and New Research; Boyd, I., Ed.; Nova Science Publishers Inc.: Hauppauge, NY, USA, 2016. [Google Scholar]
- EFSA Panel on Contaminants in the Food Chain (CONTAM). Scientific opinion on the appropriateness to set a group health based guidance value for T2 and HT2 toxin and its modified forms. EFSA J. 2017. [Google Scholar] [CrossRef]
- Mesterhazy, A.; Bartok, T.; Kaszonyi, G.; Varga, M.; Toth, B.; Varga, J. Common resistance to different fusarium spp. Causing fusarium head blight in wheat. Eur. J. Plant Pathol. 2005, 112, 267–281. [Google Scholar] [CrossRef]
- He, X.; Bonnett, D.; Singh, P.K.; Hyles, J.; Spielmeyer, W.; Dreisigacker, S. Advanced wheat breeding lines combining fhb1 and sr2 in different genetic backgrounds. In Proceedings of the 9th International Wheat Conference, Sydney, Australia, 20–25 September 2015; p. 140. [Google Scholar]
- Nakagawa, H.; Naito, S.; Kitani, Y.; Ito, Y.; Aoyama, Y.; Koyama, M.; Hiejima, Y.; Nakamura, K.; Miyazaki, H.; Morita, H.-I.; et al. Harmonized Collaborative Validation of a Simultaneous and Multiple Determination Method for Nivalenol, Deoxynivalenol, T-2 Toxin, HT-2 Toxin, and Zearalenone in Wheat and Barley by Liquid Chromatography Coupled to Tandem Mass Spectrometry (LC-MS/MS). J. Anal. Bioanal. Tech. 2014. [Google Scholar] [CrossRef]
- Thammawong, M.; Okadome, H.; Shiina, T.; Nakagawa, H.; Nagashima, H.; Nakajima, T.; Kushiro, M. Distinct distribution of deoxynivalenol, nivalenol, and ergosterol in Fusarium-infected Japanese soft red winter wheat milling fractions. Mycopathologia 2011, 172, 323–330. [Google Scholar] [CrossRef] [PubMed]
- Ban, T.; Suenaga, K. Genetic analysis of resistance to Fusarium head blight caused by Fusarium graminearum in Chinese wheat cultivar Sumai 3 and the Japanese cultivar Saikai 165. Euphytica 2000, 113, 87–99. [Google Scholar] [CrossRef]
- Lemmens, M.; Scholz, U.; Berthiller, F.; Dall’Asta, C.; Koutnik, A.; Schuhmacher, R.; Adam, G.; Buerstmayr, H.; Mesterházy, A.; Krska, R.; et al. The ability to detoxify the mycotoxin deoxynivalenol colocalizes with a major quantitative trait locus for Fusarium head blight resistance in wheat. Mol. Plant Microbe Interact. 2005, 18, 1318–1324. [Google Scholar] [CrossRef] [PubMed]
- Poppenberger, B.; Berthiller, F.; Lucyshyn, D.; Sieberer, T.; Schuhmacher, R.; Krska, R.; Kuchler, K.; Glössl, J.; Luschnig, C.; Adam, G. Detoxification of the Fusarium Mycotoxin Deoxynivalenol by a UDP-glucosyltransferase from Arabidopsis thaliana. J. Biol. Chem. 2003, 278, 47905–47914. [Google Scholar] [CrossRef] [PubMed]
- Schweiger, W.; Boddu, J.; Shin, S.; Poppenberger, B.; Berthiller, F.; Lemmens, M.; Muehlbauer, G.J.; Adam, G. Validation of a Candidate Deoxynivalenol-Inactivating UDP-Glucosyltransferase from Barley by Heterologous Expression in Yeast. Mol. Plant Microbe Interact. 2010, 23, 977–986. [Google Scholar] [CrossRef] [PubMed]
- Rawat, N.; Pumphrey, M.O.; Liu, S.; Zhang, X.; Tiwari, V.K.; Ando, K.; Trick, H.N.; Bockus, W.W.; Akhunov, E.; Anderson, J.A.; et al. Wheat Fhb1 encodes a chimeric lectin with agglutinin domains and a pore-forming toxin-like domain conferring resistance to Fusarium head blight. Nat. Genet. 2016, 48, 1576–1580. [Google Scholar] [CrossRef] [PubMed]
- Steiner, B.; Zimmerl, S.; Polzer, R.; Mühl, S.; Lemmens, M.; Adam, G.; Till, B.; Schweiger, W.; Buerstmayr, H. Functional identification of the wheat gene enhancing mycotoxin detoxification of the major Fusarium resistance QTL Fhb1. In Proceedings of the 13th International Wheat Genetics Symposium, Tulln, Austria, 23–28 April 2017; Buerstmayr, H., Lang-Mladek, C., Steiner, B., Michel, S., Buerstmayr, M., Lemmens, M., Vollmann, J., Grausgruber, H., Eds.; p. 70. [Google Scholar]
- Berthiller, F.; Crews, C.; Dall’Asta, C.; Saeger, S.D.; Haesaert, G.; Karlovsky, P.; Oswald, I.P.; Seefelder, W.; Speijers, G.; Stroka, J. Masked mycotoxins: A review. Mol. Nutr. Food Res. 2013, 57, 165–186. [Google Scholar] [CrossRef] [PubMed]
- Lemmens, M.; Steiner, B.; Sulyok, M.; Nicholson, P.; Mesterhazy, A.; Buerstmayr, H. Masked mycotoxins: Does breeding for enhanced Fusarium head blight resistance result in more deoxynivalenol-3-glucoside in new wheat varieties? World Mycotoxin J. 2016, 9, 741–754. [Google Scholar] [CrossRef]
- Audenaert, K.; De Boevre, M.; Vanheule, A.; Callewaert, J.; Bekaert, B.; Höfte, M.; De Saeger, S.; Haesaert, G. Mycotoxin glucosylation in commercial wheat varieties: Impact on resistance to fusarium graminearum under laboratory and field conditions. Food Control 2013, 34, 756–762. [Google Scholar] [CrossRef]
- He, X.; Singh, P.K.; Duveiller, E.; Schlang, N.; Dreisigacker, S.; Singh, R.P. Identification and characterization of international fusarium head blight screening nurseries of wheat at cimmyt, mexico. Eur. J. Plant Pathol. 2013, 136, 123–134. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| ppb | Number of Countries |
|---|---|
| 1100–2000 | 4 |
| 1000 | 9 |
| 750 | 19 |
| 300–700 | 5 |
| Analyte | Precursor Ion | Product Ion | Retention Time (min) |
|---|---|---|---|
| NIV (C15H20O7) | 371.1 [C15H20O7+CH3COO]− | 280.8 | 1.74–1.81 |
| 310.7 | 1.71–1.87 | ||
| 59.0 | 1.74–1.87 | ||
| D3G (C21H30O11) | 517.1 [C21H30O11+CH3COO]− | 456.9 | 2.28–2.48 |
| 426.9 | 2.29–2.49 | ||
| 58.9 | 2.28–2.48 | ||
| DON (C15H20O6) | 355.1 [C15H20O6+CH3COO]− | 294.8 | 2.86–3.00 |
| 264.8 | 2.86–3.00 | ||
| 58.9 | 2.86–3.00 |
| FHB Index | ERG (ppm) | NIV (ppb) | D3G (ppb) | DON (ppb) | D3G/DON Ratio (%) | |
|---|---|---|---|---|---|---|
| Range | 1.48–87.22 | 1.49–29.57 | 3.7–496.5 | 23.5–2682.6 | 131.3–6337.4 | 8.1–37.7 |
| Average | 19.03 | 8.99 | 96.8 | 1145 | 3161 | 24.9 |
| FHB | ERG | D3G | DON | D3G/DON | NIV | |
|---|---|---|---|---|---|---|
| FHB | 1 | |||||
| ERG | 0.37 ** | 1 | ||||
| D3G | 0.32 * | 0.55 *** | 1 | |||
| DON | 0.36 ** | 0.55 *** | 0.91 *** | 1 | ||
| D3G/DON | 0.03 | 0.09 | 0.32 * | −0.01 | 1 | |
| NIV | 0.35 * | 0.25 | 0.45 *** | 0.42 ** | 0.17 | 1 |
© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Nakagawa, H.; He, X.; Matsuo, Y.; Singh, P.K.; Kushiro, M. Analysis of the Masked Metabolite of Deoxynivalenol and Fusarium Resistance in CIMMYT Wheat Germplasm. Toxins 2017, 9, 238. https://doi.org/10.3390/toxins9080238
Nakagawa H, He X, Matsuo Y, Singh PK, Kushiro M. Analysis of the Masked Metabolite of Deoxynivalenol and Fusarium Resistance in CIMMYT Wheat Germplasm. Toxins. 2017; 9(8):238. https://doi.org/10.3390/toxins9080238
Chicago/Turabian StyleNakagawa, Hiroyuki, Xinyao He, Yosuke Matsuo, Pawan K. Singh, and Masayo Kushiro. 2017. "Analysis of the Masked Metabolite of Deoxynivalenol and Fusarium Resistance in CIMMYT Wheat Germplasm" Toxins 9, no. 8: 238. https://doi.org/10.3390/toxins9080238
APA StyleNakagawa, H., He, X., Matsuo, Y., Singh, P. K., & Kushiro, M. (2017). Analysis of the Masked Metabolite of Deoxynivalenol and Fusarium Resistance in CIMMYT Wheat Germplasm. Toxins, 9(8), 238. https://doi.org/10.3390/toxins9080238