
Use of Carabids for the Post-Market Environmental Monitoring of Genetically Modified Crops

Abstract
:
1. Introduction
2. Results
2.1. Characterization and Quantitative Comparison of Carabid Communities
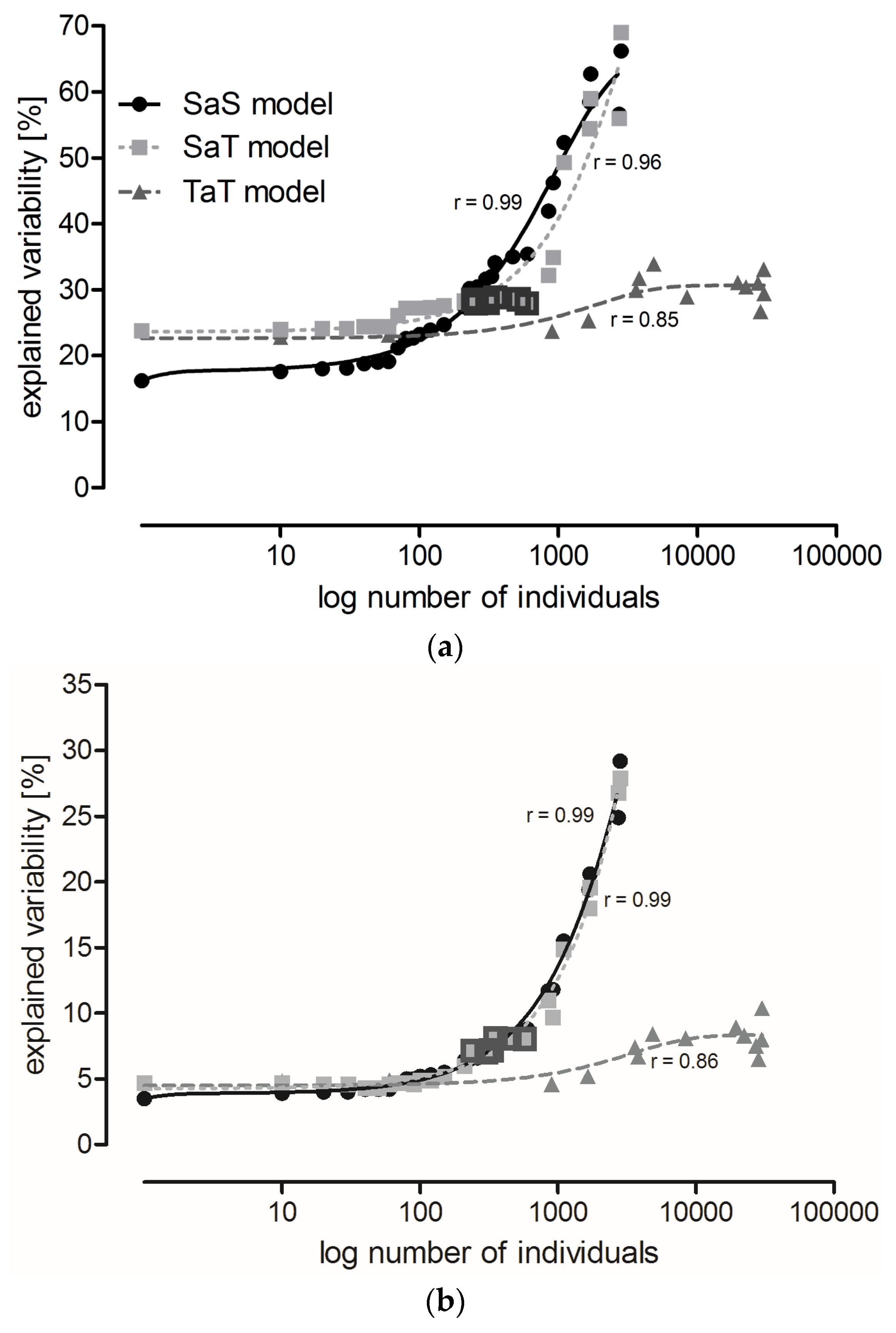
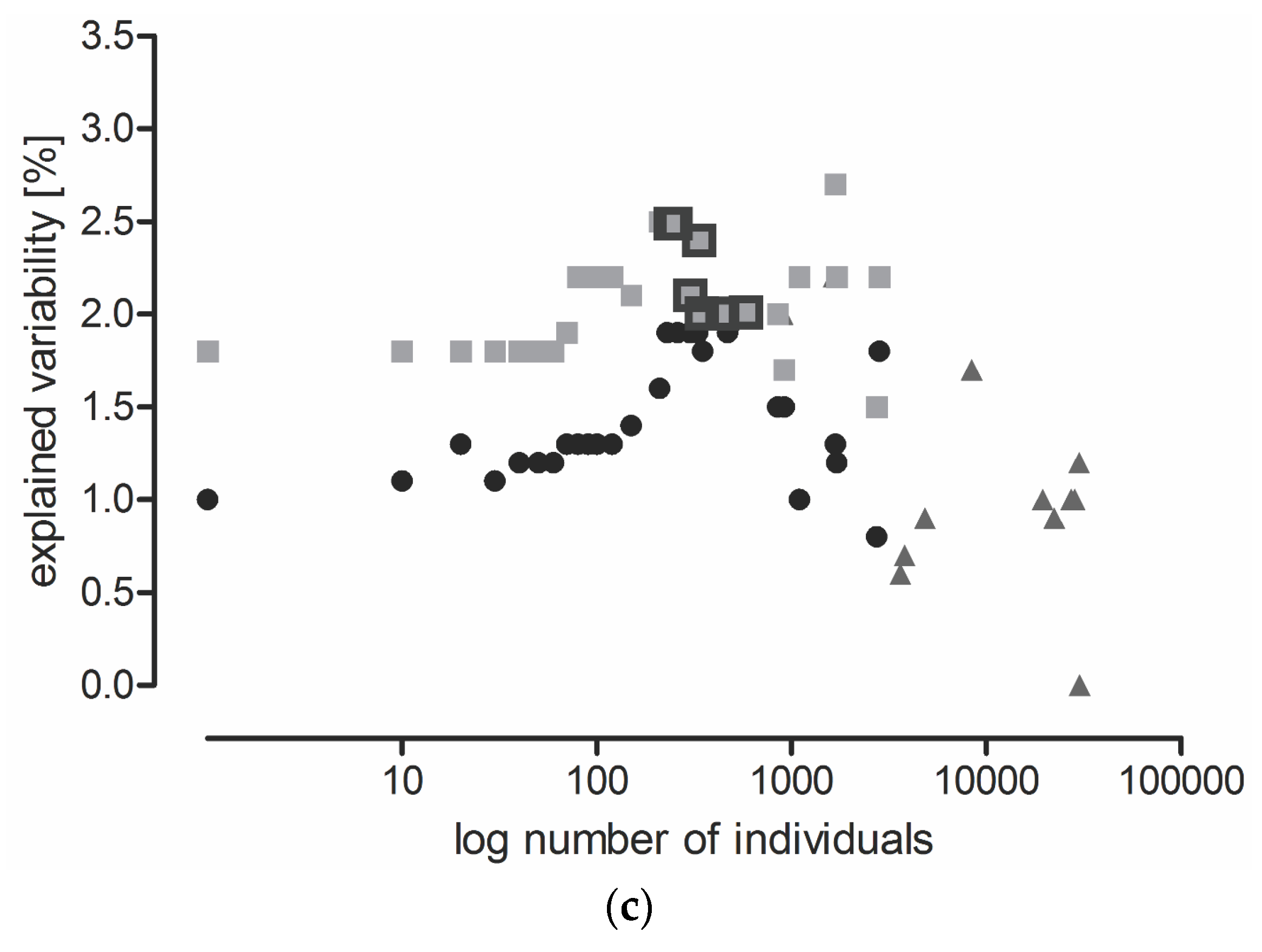
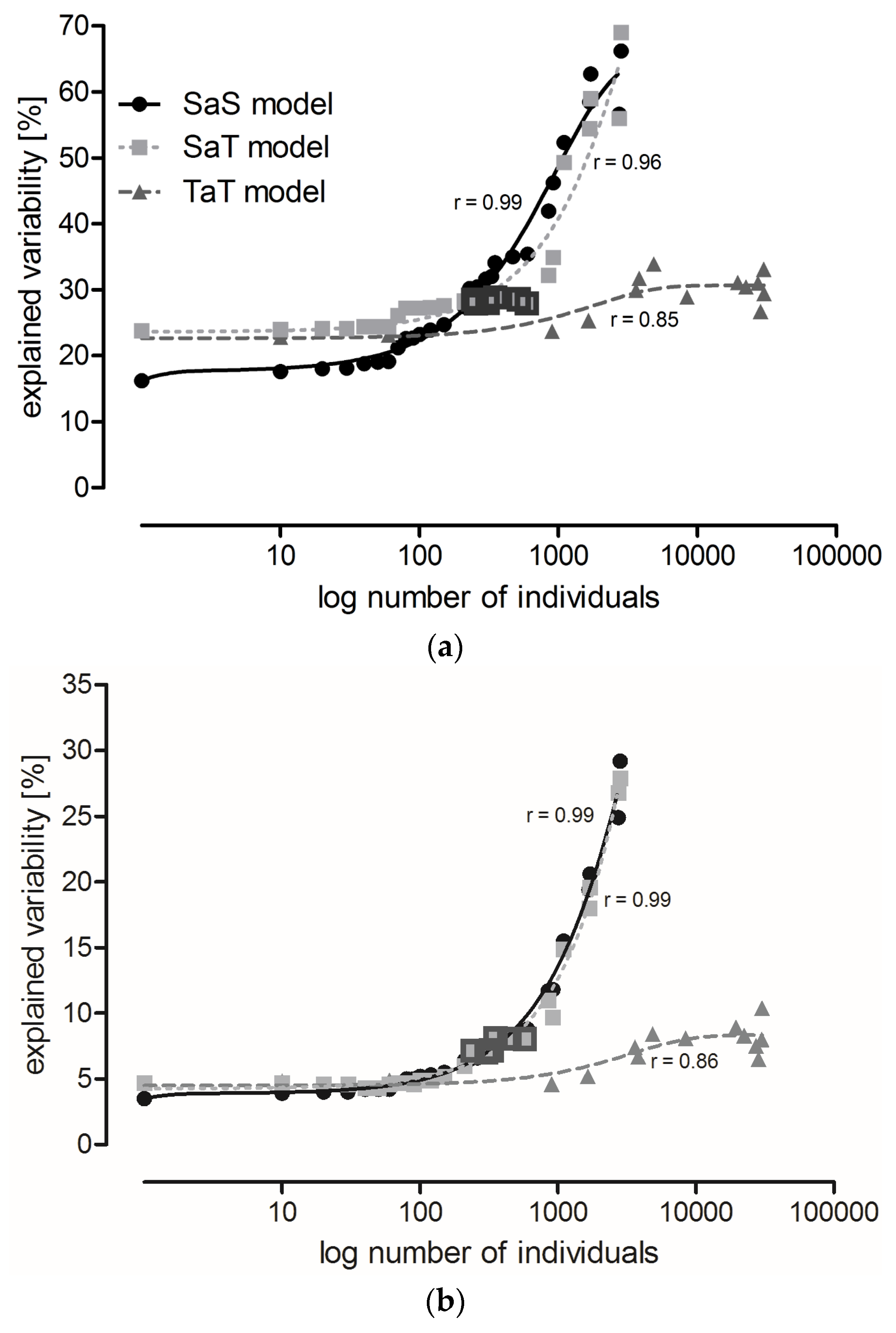
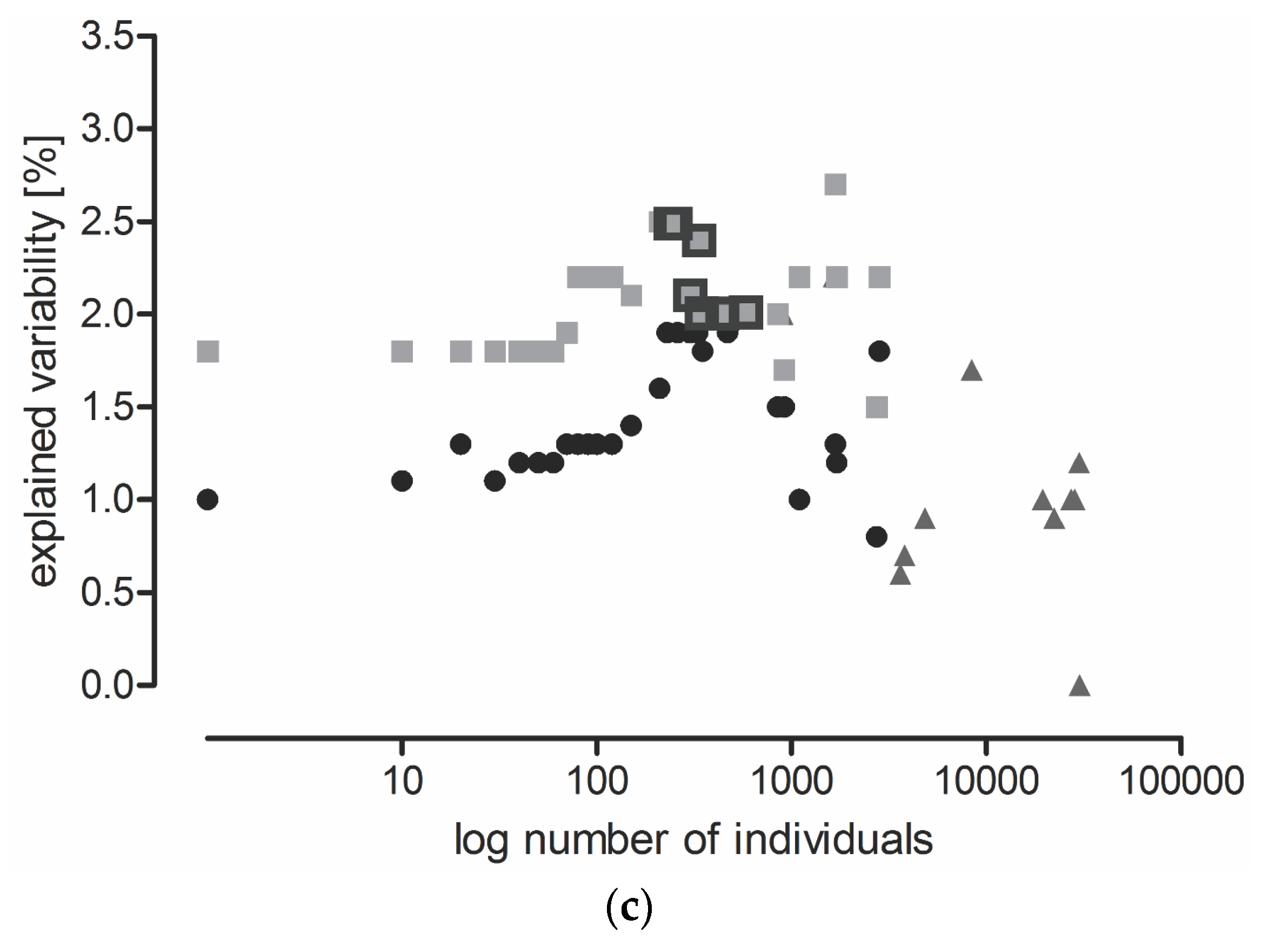
2.2. The Effect of Locality, Year, and Sampling Date on Carabid Communities (All Data Included)
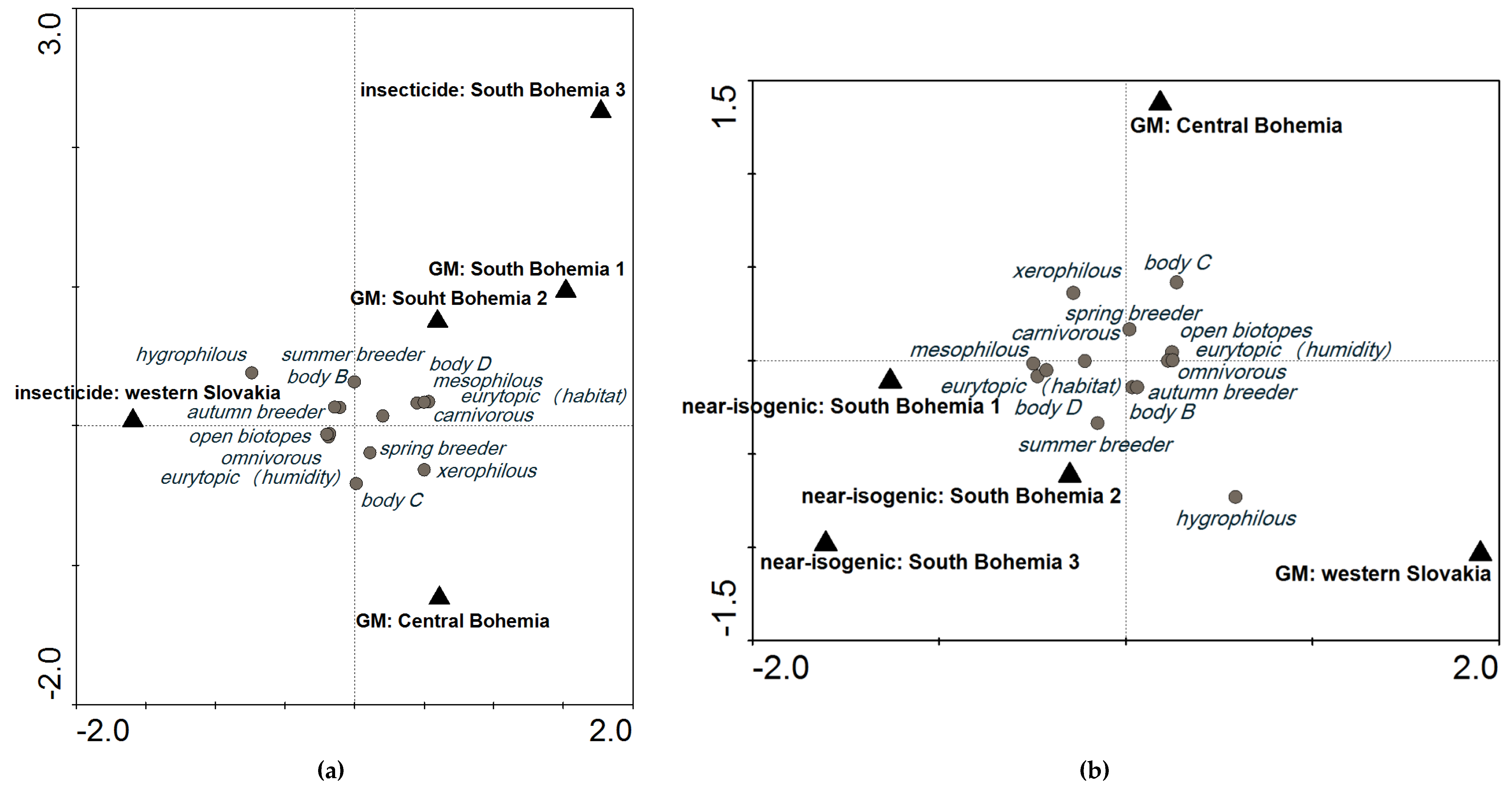
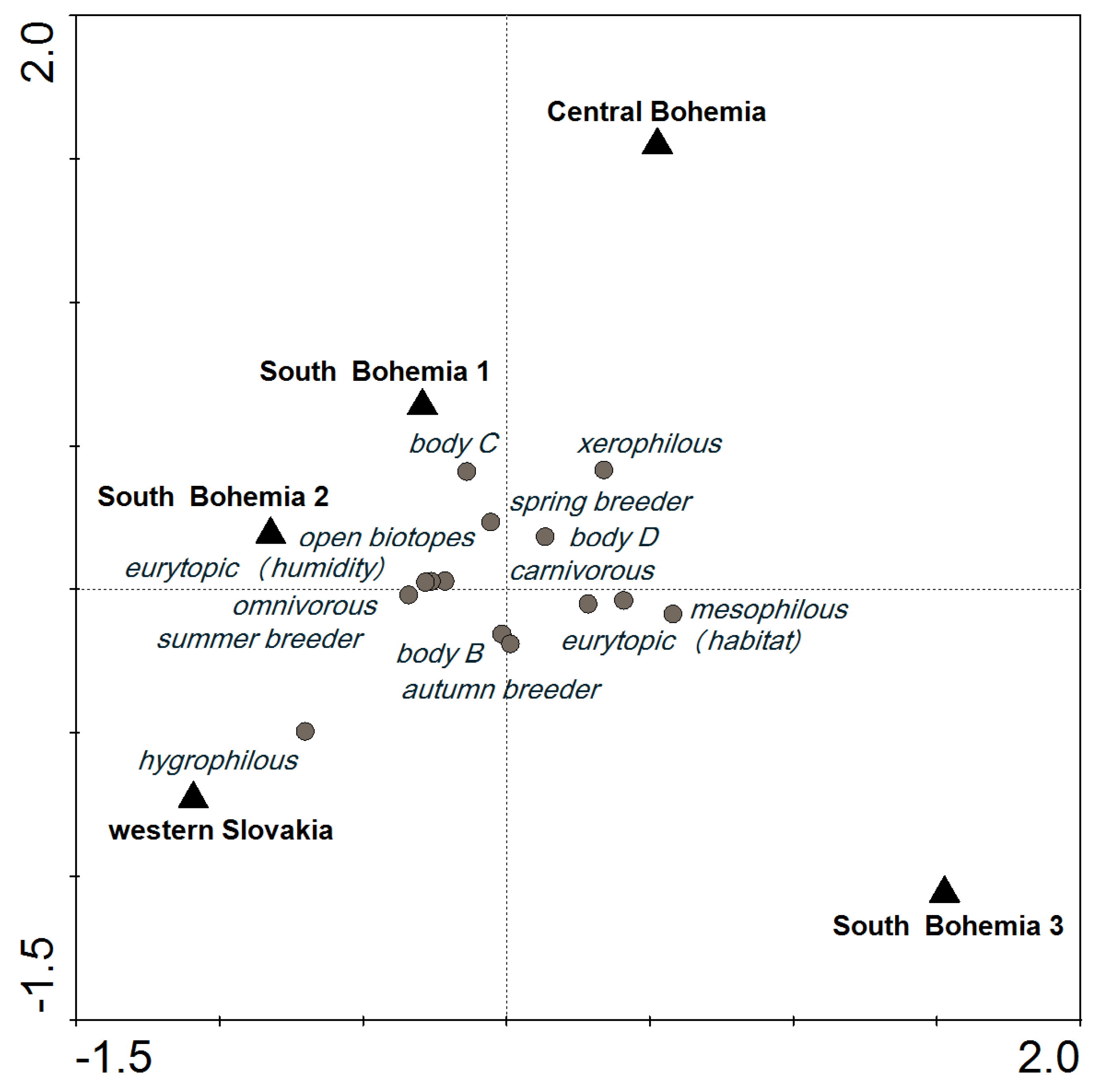
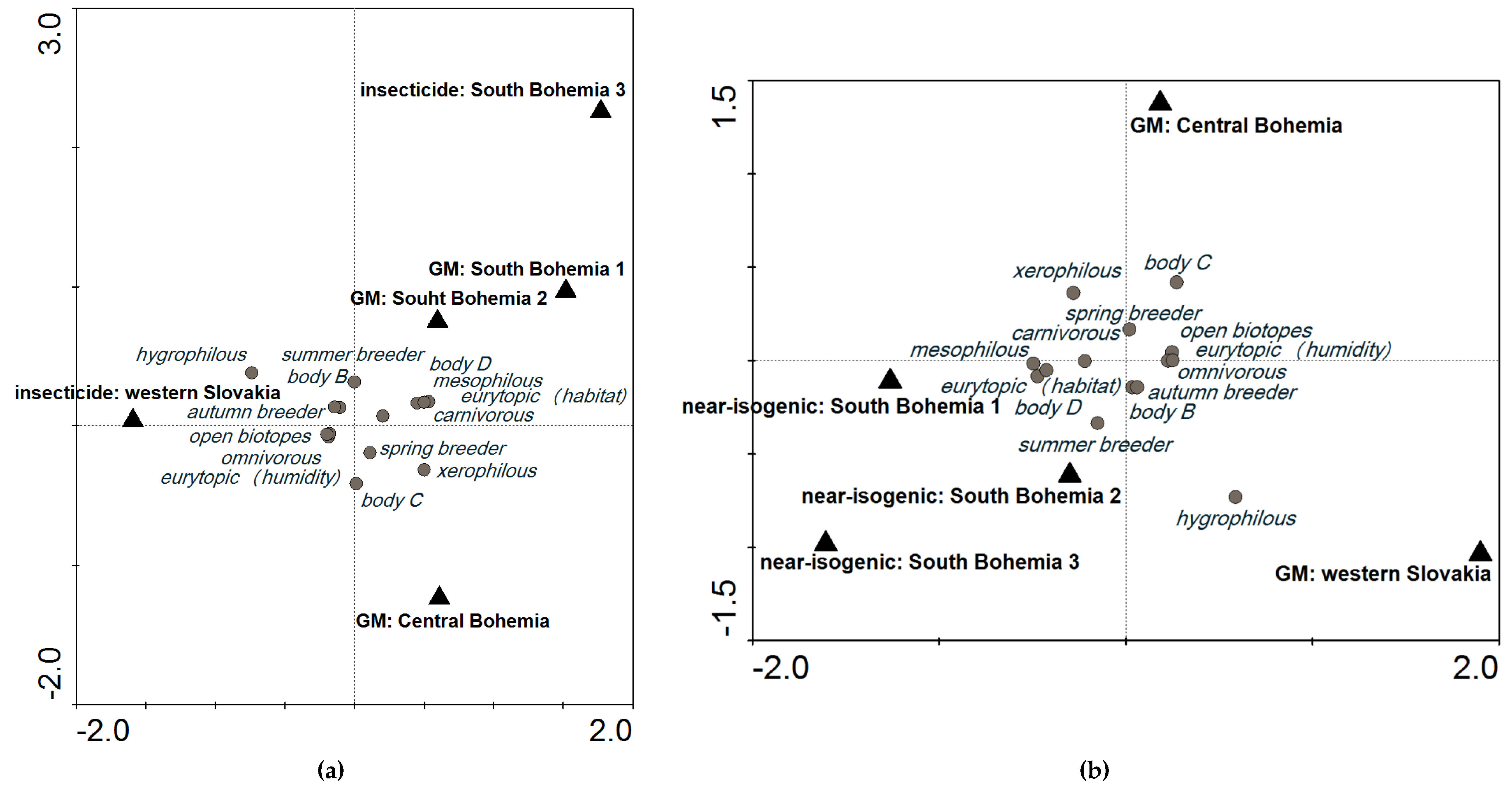
2.3. Three Possible Ways of Using Carabids in PMEM
3. Discussion
3.1. Carabid Communities in Maize Fields
3.2. Variability Explained by Locality and Environmental Variables
3.3. The Applicability of Our Findings for GS in PMEM of GM Maize
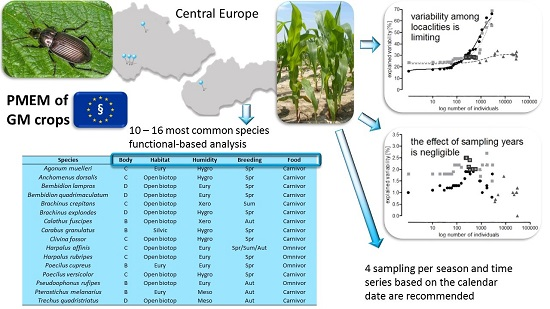
4. Conclusions
5. Materials and Methods
5.1. Experimental Localities
5.2. Capture and Identification of Carabids
5.3. Data Analysis
- SaS model: the least abundant species were eliminated and a CCA was performed at the species level (Table S4);
- SaT model: the least abundant species were eliminated and a CCA was performed at the level of functional traits (Table S5); and
- TaT model: the least frequent categories of functional traits were eliminated and a CCA was performed at the level of functional traits (Table S6).
Supplementary Materials
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Wolfenbarger, L.L.; Naranjo, S.E.; Lundgren, J.G.; Bitzer, R.J.; Watrud, L.S. Bt crop effects on functional guilds of non-target arthropods: A meta-analysis. PLoS ONE 2008, 3, e2118. [Google Scholar] [CrossRef] [PubMed]
- European Commission (EC). Directive 2001/18/EC of the European Parliament and of the Council of 12 March 2001 on the deliberate release into the environment of genetically modified organisms and repealing. Council Directive 90/220/EEC. Off. J. Eur. Communities 2001, L106, 1. [Google Scholar]
- European Commission. Council Decision of 3 October 2002 establishing guidance notes supplementing Annex VII to Directive 2001/18/EC or the European Parliament and of the Council on the deliberate release into the environment of genetically modified organisms and repealing. Council Directive 90/220/EEC. Off. J. Eur. Communities 2002, L280, 27. [Google Scholar]
- EFSA (European Food Safety Authority) Panel on GMO. Opinion of the Scientific Panel on Genetically Modified Organisms on the Post Market Environmental Monitoring (PMEM) of genetically modified plants. EFSA J. 2006, 319, 1–27. [Google Scholar]
- Sanvido, O.; De Schrijver, A.; Devos, Y.; Bartsch, D. Post market environmental monitoring of genetically modified herbicide tolerant crops (Working group report from the 4th International Workshop on PMEM of Genetically Modified Plants, Quedlinburg, Germany 2010). J. Kult. Pflanzen. 2011, 63, 211–216. [Google Scholar]
- Sanvido, O.; Widmer, F.; Winzeler, M.; Bigler, F. A conceptual framework for the design of environmental post-market monitoring of genetically modified plants. Environ. Biosaf. Res. 2005, 4, 13–27. [Google Scholar] [CrossRef]
- EFSA (European Food Safety Authority) Panel on GMO. Scientific Opinion on guidance on the Post-Market Environmental Monitoring (PMEM) of genetically modified plants. EFSA J. 2011, 9, 2316. [Google Scholar]
- Albajes, R.; Farinós, G.P.; Pérez-Hedo, M.; de la Poza, M.; Lumbierres, B.; Ortego, F.; Pons, X.; Castañera, P. Post-market environmental monitoring of Bt maize in Spain: Non-target effects of varieties derived from the event MON810 on predatory fauna. Spanish J. Agric. Res. 2012, 10, 977–985. [Google Scholar] [CrossRef]
- Albajes, R.; Lumbierres, B.; Pons, X.; Comas, J. Representative taxa in field trials for environmental risk assessment of genetically modified maize. Bull. Entomol. Res. 2013, 103, 724–733. [Google Scholar] [CrossRef] [PubMed]
- Skoková Habuštová, O.; Svobodová, Z.; Spitzer, L.; Doležal, P.; Hussein, H.M.; Sehnal, F. Communities of ground-dwelling arthropods in conventional and transgenic maize: Background data for the post-market environmental monitoring. J. Appl. Entomol. 2015, 139, 31–45. [Google Scholar]
- Lee, M.S.; Albajes, R. Monitoring carabid indicators could reveal environmental impacts of genetically modified maize. Agric. For. Entomol. 2016, 18, 238–249. [Google Scholar] [CrossRef]
- Sanvido, O.; Romeis, J.; Bigler, F. An approach for post-market monitoring of potential environmental effects of Bt-maize expressing Cry1Ab on natural enemies. J. Appl. Entomol. 2009, 133, 236–248. [Google Scholar]
- Kotze, D.J.; Brandmayr, P.; Casale, A.; Dauffy-Richard, E.; Dekoninck, W.; Koivula, M.J.; Lövei, G.L.; Mossakowski, D.; Noordijk, J.; Paarmann, W.; et al. Forty years of carabid beetle research in Europe—From taxonomy, biology, ecology and population studies to bioindication, habitat assessment and conservation. ZooKeys 2011, 100, 55–148. [Google Scholar] [CrossRef] [PubMed]
- Zahradník, J. Brouci; Aventinum: Praha, Czech Republic, 2008; p. 288. [Google Scholar]
- Hůrka, K. Střevlíkovití, Carabidae 1; Academia: Praha, Czech Republic, 1992; p. 192. [Google Scholar]
- Holland, J.M.; Luff, M.L. The effects of agricultural practices on Carabidae in temperate agroecosystems. Integr. Pest Manag. Rev. 2000, 5, 109–129. [Google Scholar] [CrossRef]
- Vician, V.; Svitok, M.; Kočík, K.; Stašiov, S. The influence of agricultural management on the structure of ground beetle (Coleoptera: Carabidae) assemblages. Biologia 2015, 70, 240–251. [Google Scholar] [CrossRef]
- Jelaska, L.Š.; Blanuš, M.; Durbešić, P.; Jelaskac, S.D. Heavy metal concentrations in ground beetles, leaf litter, and soil of a forest ecosystem. Ecotoxicol. Environ. Saf. 2007, 66, 74–81. [Google Scholar] [CrossRef] [PubMed]
- Pizzolotto, R.; Cairns, W.; Barbante, C. Pilot research on testing the reliability of studies on carabid heavy metals contamination. Balt. J. Coleopterol. 2013, 13, 1–13. [Google Scholar]
- Romeis, J.; Meissle, M.; Alvarez-Alfageme, F.; Bigler, F.; Bohan, D.A.; Devos, Y.; Malone, L.A.; Pons, X.; Rauschen, S. Potential use of an arthropod database to support the nontarget risk assessment and monitoring of transgenic plants. Transgen. Res. 2014, 23, 995–1013. [Google Scholar] [CrossRef] [PubMed]
- Comas, J.; Lumbierres, B.; Comas, C.; Pons, X.; Albajes, R. Optimizing the capacity of field trials to detect the effect of genetically modified maize on non-target organisms through longitudinal sampling. Ann. Appl. Biol. 2015, 166, 183–195. [Google Scholar] [CrossRef]
- Zetto Brandmayr, T. Spermophagous (seed-eating) ground beetles: First comparison of the diet and ecology of the harpaline genera Harpalus and Ophonus (Col., Carabidae). In The Role of Ground Beetles in Ecological and Environmental Studies; Stork, N.E., Ed.; Intercept: Andover, UK, 1990; pp. 307–316. [Google Scholar]
- Larochelle, A. The food of carabid beetles (Coleoptera: Carabidae, including Cicindelinae). Fabreries 1990, 5, 1–132. [Google Scholar]
- Talarico, F.; Giglo, A.; Pizzolotto, R.; Brandmayr, P. A synthesis of feeding habits and reproduction rhythm in Italian seed-feeding ground beetles (Coleoptera: Carabidae). Eur. J. Entomol. 2015, 113, 325–336. [Google Scholar] [CrossRef]
- Tooley, J.; Brust, G.E. Weed seed predation by carabid beetles. In The Agroecology of Carabid Beetles; Holland, J.M., Ed.; Intercept: Andover, UK, 2015; pp. 215–230. [Google Scholar]
- Honěk, A.; Martinková, Z.; Saska, P. Post-dispersal predation of Taraxacum officinale (dandelion) seed. J. Ecol. 2005, 93, 345–352. [Google Scholar] [CrossRef]
- Saska, P.; Němeček, J.; Koprdová, S.; Skuhrovec, J.; Káš, M. Weeds determine the composition of carabid assemblage in maize at a fine scale. Sci. Agric. Bohem. 2014, 45, 85–92. [Google Scholar] [CrossRef]
- Kromp, B. Carabid beetles in sustainable agriculture: A review on pest control efficacy, cultivation aspects and enhancement. Agric. Ecosyst. Environ. 1999, 74, 187–228. [Google Scholar] [CrossRef]
- Peters, R.H. The Ecological Implications of Body Size; Cambridge University Press: Cambridge, UK, 1983; p. 329. [Google Scholar]
- Sharova, I.C.H. Life Forms of Carabids; Nauka: Moscow, Russia, 1981; p. 359. [Google Scholar]
- Riedel, J.; Romeis, J.; Meissle, M. Update and expansion of the database of bio-ecological information on non-target arthropod species established to support the environmental risk assessment of genetically modified crops in the EU. EFSA Support. Publ. 2016. [Google Scholar] [CrossRef]
- Brooks, D.R.; Bohan, D.A.; Champion, G.T.; Haughton, A.J.; Hawes, C.; Heard, M.S.; Clark, J.S.; Dewar, A.M.; Firbank, L.G.; Perryl, J.N.; et al. Responses of invertebrates to contrasting herbicide regimes in genetically modified herbicide-tolerant crops. I. Soil-surface-active invertebrates. Proc. R. Soc. Lond. B Biol. Sci. 2003, 358, 1847–1862. [Google Scholar]
- Clough, Y.; Kruess, A.; Tscharntke, T. Organic versus conventional arable farming systems: Functional grouping helps understand staphylinid response. Agric. Ecosyst. Environ. 2007, 118, 285–290. [Google Scholar] [CrossRef]
- Grabowski, M.; Bereś, P.K.; Dąbrowski, Z.T. Charakterystyka wybranych gatunków biegaczowatych (Coleoptera: Carabidae) pod kątem ich przydatności dla oceny ryzyka i monitoringu uwalniania GMO do środowiska. Prog. Plant Prot. 2010, 50, 1602–1606. [Google Scholar]
- James, C. 20th Anniversary 1996 to 2015 of the Global Commercialization of Biotech Crops and Biotech Crop Highlights in 2015; ISAAA Brief No. 51; ISAAA: Ithaca, NY, USA, 2015; Available online: http://www.isaaa.org/resources/publications/briefs/51/ (accessed on 12 February 2016).
- Mast, B.; Graeff Hönninger, S.; Claupein, W. Evaluation of carabid beetle diversity in different bioenergy cropping systems. Sustain. Agric. Res. 2012, 2, 127–140. [Google Scholar] [CrossRef]
- Purchart, L.; Kula, E. Ground beetles (Coleoptera, Carabidae) agrocenoses of spring and winter wheat. Acta Univ. Agric. Silvic. Mendel. Brun. 2005, 53, 125–132. [Google Scholar] [CrossRef]
- Tancredi Caruso, T.; Pigino, G.; Bernini, F.; Bargagli, R.; Migliorini, M. The Berger–Parker index as an effective tool for monitoring the biodiversity of disturbed soils: A case study on Mediterranean oribatid (Acari: Oribatida) assemblages. Biodivers. Conserv. 2007, 16, 3277–3285. [Google Scholar]
- Irmler, U. The spatial and temporal pattern of carabid beetles on arable fields in northern Germany (Schleswig-Holstein) and their value as ecological indicators. Agric. Ecosyst. Environ. 2003, 98, 141–151. [Google Scholar] [CrossRef]
- Porhajašová, J.; Petřvalský, V.; Šustek, Z.; Urminská, J.; Ondrišík, P.; Noskovič, J. Long-termed changes in ground beetle (Coleoptera: Carabidae) assemblages in a field treated by organic fertilizers. Biologia 2008, 63, 1184–1195. [Google Scholar] [CrossRef]
- Farinós, G.P.; de la Poza, M.; Hernández-Crespo, P.; Ortego, F.; Castañera, P. Diversity and seasonal phenology of aboveground arthropods in conventional and transgenic maize crops in Central Spain. Biol. Control 2008, 44, 362–371. [Google Scholar] [CrossRef]
- Coman, D.; Roșca, I. Structure, dynamics and abundance of carabid species collected in corn fields. Sci. Pap. Ser. A Agron. 2013, 61, 477–479. [Google Scholar]
- Meissle, M.; Lang, A. Comparing methods to evaluate the effects of Bt maize and insecticide on spider assemblages. Agric. Ecosyst. Environ. 2005, 107, 359–370. [Google Scholar] [CrossRef]
- Comas, J.; Lumbierres, B.; Pons, X.; Albajes, R. Ex-ante determination of the capacity of field tests to detect effects of genetically modified corn on nontarget arthropods. J. Econom. Entomol. 2013, 106, 1659–1668. [Google Scholar] [CrossRef]
- Comas, C.; Lumbierres, B.; Pons, X.; Albajes, R. No effects of Bacillus thuringiensis maize on nontarget organisms in the field in southern Europe: A meta-analysis of 26 arthropod taxa. Transgen. Res. 2014, 23, 135–143. [Google Scholar] [CrossRef] [PubMed]
- Svobodová, Z.; Skoková Habuštová, O.; Holec, J.; Holec, M.; Boháč, J.; Jursík, M.; Soukup, J.; Sehnal, F. Weeds and epigeic arthropods communities in herbicide-tolerant maize grown under different herbicide and tillage regimes. Agric. Ecosyst. Environ. 2017. under review. [Google Scholar]
- Devos, Y.; Aguilera, J.; Diveki, Z.; Gomes, A.; Liu, Y.; Paoletti, C.; du Jardin, P.; Herman, L.; Perry, J.N.; Waigmann, E. EFSA’s scientific activities and achievements on the risk assessment of genetically modified organisms (GMOs) during its first decade of existence: Looking back and ahead. Transgen. Res. 2014, 23, 1–25. [Google Scholar] [CrossRef] [PubMed]
- Spitzer, L.; Růžička, V.; Hussein, H.; Habuštová, O.; Sehnal, F. Expression of a Bacillus thuringiensis toxin in maize does not affect epigeic communities of carabid beetles and spiders. Acta Phytotech. Zootech. 2004, 7, 110–112. [Google Scholar]
- Habuštová, O.; Doležal, P.; Spitzer, L.; Svobodová, Z.; Hussein, H.; Sehnal, F. Impact of Cry1Ab toxin expression on the non-target insects dwelling on maize plants. J. Appl. Entomol. 2014, 138, 164–172. [Google Scholar] [CrossRef]
- Svobodová, Z.; Habuštová, O.; Sehnal, F.; Holec, M.; Hussein, H.M. Epigeic spiders are not affected by the genetically modified maize MON 88017. J. App. Entomol. 2013, 137, 56–67. [Google Scholar] [CrossRef]
- Lancashire, P.D.; Bleiholder, H.; Van den Boom, T.; Langelüddecke, P.; Stauss, R.; Weber, E.; Witzen-Berger, A. An uniform decimal code for growth stages of crops and weeds. Ann. Appl. Biol. 1991, 119, 561–601. [Google Scholar] [CrossRef]
- Hůrka, K. Carabidae of the Czech and Slovak Republics; Kabourek: Zlín, Czech Republic, 1996; p. 565. [Google Scholar]
- Veselý, P.; Moravec, P.; Stanovský, J. Carabidae (střevlíkovití). In Červený Seznam Ohrožených Druhů České Republiky; Farkáč, J., Král, D., Škorpík, M., Eds.; AOPK: Praha, Czech Republic, 2005; pp. 406–411. [Google Scholar]
- Holecová, M.; Franc, V. Red (Ecosozological) list of beetles (Coleoptera) of Slovakia. In Red List of Plants and Animals of Slovakia. Ochr. Prír. 2001, 20 (Supp.), 111–128. [Google Scholar]
- Magurran, A.E. Measuring Biological Diversity; Blackwell Publishing: Oxford, Great Britain, 2004; p. 215. [Google Scholar]
- Lepš, J.; Šmilauer, P. Multivariate Analysis of Ecological Data Using CANOCO; Cambridge University Press: Cambridge, UK, 2003; p. 268. [Google Scholar]
- Krishna, K.R. Agroecosystems: Soils, Climate, Crops, Nutrient Dynamics and Productivity; CRC Press: Boca Raton, FL, USA, 2013; p. 552. [Google Scholar]
- GraphPad Software Inc. GraphPad Prism 5.0 User´s Guide; GraphPad Software Inc.: San Diego, CA, USA, 2007; Available online: http://graphpad.com/guides/prism/5/user-guide/prism5help.html (accessed on 13 August 2016).
- StatSoft Inc. Statistica Electronic Manual; StatSoft Inc.: Tulsa, OK, USA, 2015; Available online: http://documentation.statsoft.com/STATISTICAHelp.aspx?path=common/AboutSTATISTICA/ElectronicManualIndex (accessed on 13 August 2016).


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Trait | Category 1 | SB1 | SB2 | SB3 | CB | WS | Total | ||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Individuals | Species | Individuals | Species | Individuals | Species | Individuals | Species | Individuals | Species | Individuals (%) | Species (%) | ||
| Body size | |||||||||||||
| A | 36 | 1 | 14 | 3 | 2 | 1 | 0 | 0 | 3 | 2 | 55 (0.1) | 4 (5) | |
| B | 3692 | 8 | 18,349 | 10 | 14,728 | 10 | 2274 | 7 | 7214 | 13 | 46,257 (79) | 19 (22) | |
| C | 456 | 17 | 2660 | 34 | 409 | 15 | 3101 | 18 | 1765 | 12 | 8391 (14) | 44 (51) | |
| D | 1300 | 9 | 992 | 12 | 434 | 8 | 456 | 10 | 419 | 7 | 3601 (6) | 19 (22) | |
| Habitat affinity | |||||||||||||
| Silvicolous | 59 | 6 | 635 | 15 | 130 | 7 | 66 | 3 | 4 | 7 | 894 (2) | 19 (22) | |
| Open biotopes | 2673 | 16 | 4861 | 30 | 1437 | 17 | 4302 | 22 | 9009 | 23 | 22,282 (38) | 44 (51) | |
| Eurytopic | 2752 | 13 | 16,519 | 14 | 14,006 | 10 | 1463 | 10 | 388 | 7 | 35,128 (60) | 23 (27) | |
| Humidity affinity | |||||||||||||
| Hygrophilous | 329 | 15 | 2563 | 24 | 404 | 16 | 133 | 11 | 1417 | 8 | 4846 (8) | 34 (40) | |
| Mesophilous | 2038 | 6 | 3163 | 11 | 12,855 | 6 | 1161 | 4 | 275 | 6 | 19,492 (33) | 15 (17) | |
| Eurytopic | 2288 | 10 | 15,622 | 14 | 1834 | 9 | 3489 | 10 | 6935 | 8 | 30,168 (52) | 17 (20) | |
| Xerophilous | 829 | 4 | 667 | 10 | 480 | 3 | 1048 | 10 | 774 | 12 | 3798 (7) | 20 (23) | |
| Breeding period | |||||||||||||
| Spring | 2340 | 23 | 17,356 | 42 | 2080 | 24 | 3490 | 25 | 1997 | 19 | 27,263 (47) | 61 (71) | |
| Summer | 141 | 3 | 512 | 6 | 129 | 3 | 241 | 6 | 609 | 4 | 1632 (3) | 8 (9) | |
| Autumn | 3242 | 14 | 5146 | 21 | 13,610 | 11 | 2434 | 12 | 7232 | 16 | 31,664 (54) | 29 (34) | |
| Food specialization | |||||||||||||
| Carnivorous | 4258 | 24 | 7068 | 35 | 13,829 | 24 | 2322 | 20 | 2418 | 21 | 29,895 (51) | 55 (64) | |
| Omnivorous | 1226 | 11 | 14,945 | 23 | 1743 | 9 | 3508 | 14 | 6983 | 13 | 28,405 (49) | 29 (34) | |
| Granivorous | 0 | 0 | 2 | 1 | 1 | 1 | 1 | 1 | 0 | 0 | 4 (0.007) | 2 (2) | |
| Locality | No. of Tested Years | Simpson Dominance Index (D) | Berger–Parker Index (D) | Species Evenness (E) | Margalef Index (DMg) |
|---|---|---|---|---|---|
| SB1 | 1 | 0.18 | 0.32 | 0.59 | 3.95 |
| SB2 | 3 | 0.37 ± 0.08 | 0.60 | 0.44 ± 0.07 | 4.25 ± 0.55 |
| SB3 | 3 | 0.53 ± 0.15 | 0.80 | 0.38 ± 0.13 | 2.60 ± 0.23 |
| CB | 2 | 0.28 ± 0.01 | 0.47 | 0.53 ± 0.03 | 3.38 ± 0.21 |
| WS | 2 | 0.48 ± 0.08 | 0.70 | 0.39 ± 0.07 | 3.05 ± 0.30 |
| Jaccard Index (JS) | Sørensen–Dice Index (DS) | ||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| Locality | Locality | ||||||||||
| SB1 | SB2 | SB3 | CB | WS | SB1 | SB2 | SB3 | CB | WS | ||
| SB1 | SB1 | ||||||||||
| SB2 | 0.34 | SB2 | 0.51 | ||||||||
| SB3 | 0.32 | 0.32 | SB3 | 0.48 | 0.49 | ||||||
| CB | 0.26 | 0.24 | 0.23 | CB | 0.41 | 0.39 | 0.38 | ||||
| WS | 0.17 | 0.17 | 0.21 | 0.28 | WS | 0.30 | 0.29 | 0.34 | 0.43 | ||
| Environmental Variable | SaS | SaT | TaT |
|---|---|---|---|
| Locality | 5.2 (12.1) | 0.1 (6.5) | n.a. 1 (1.2) |
| Time series A (Sampling date) | 2.3 (3.6) | 0.9 (3.1) | n.a. (0.3) |
| Year | 0.1 (n. a.) | 0.5 (0) | n.a. (0.1) |
| Species | Total Abundance | Body Size | Habitat Affinity | Humidity Affinity | Breeding Period | Food Specialization |
|---|---|---|---|---|---|---|
| Agonum muelleri | 256 | C | Eurytopic | Hygrophilous | Spring | Carnivorous |
| Anchomenus dorsalis | 1099 | C | Open biotopes | Hygrophilous | Spring | Carnivorous |
| Bembidion lampros | 462 | D | Open biotopes | Eurytopic | Spring | Carnivorous |
| Bembidion quadrimaculatum | 1680 | D | Open biotopes | Eurytopic | Spring | Carnivorous |
| Brachinus crepitans | 348 | C | Open biotopes | Xerophilous | Summer | Carnivorous |
| Brachinus explondes | 294 | D | Open biotopes | Hygrophilous | Spring | Carnivorous |
| Calathus fuscipes | 2811 | B | Open biotopes | Xerophilous | Autumn | Carnivorous |
| Carabus granulatus | 596 | B | Silvicolous | Hygrophilous | Spring | Carnivorous |
| Clivina fossor | 325 | C | Open biotopes | Hygrophilous | Spring | Carnivorous |
| Harpalus affinis | 920 | C | Open biotopes | Eurytopic | Spring/summer/autumn | Omnivorous |
| Harpalus rubripes | 2734 | C | Open biotopes | Eurytopic | Spring | Omnivorous |
| Poecilus cupreus | 15,975 | B | Eurytopic | Eurytopic | Spring | Omnivorous |
| Poecilus versicolor | 1710 | C | Open biotopes | Hygrophilous | Spring | Carnivorous |
| Pseudoophonus rufipes | 7871 | B | Open biotopes | Eurytopic | Autumn | Omnivorous |
| Pterostichus melanarius | 18,297 | B | Eurytopic | Mesophilous | Autumn | Carnivorous |
| Trechus quadristriatus | 841 | D | Open biotopes | Mesophilous | Autumn | Carnivorous |
| Features | SB1 | SB2 | SB3 | CB | WS |
|---|---|---|---|---|---|
| Timing (sowing–harvest, maize stage during harvest) | 2002 (15.5–17.9. (BBCH 87)) | 2003–2005 | 2009–2011 | 2013–2014 | 2014–2015 (2014: 28.4–29.10. 2015: 5.5–30.10. (2nd trial), 4.11. (1st trial) (BBCH 89)) |
| GPS coordinates | 48°97′ N 14°44′ E | 48°58′ N 14°24′ E | 48°59′ N 14°20′ E | 50°09′ N 15°11′ E | 48°34′ N 17°43′ E |
| Altitude (m a.s.l.) | 381 | 409 | 420 | 285 | 160 |
| Climatic region | Moderately warm humid | Moderately warm humid | Moderately warm humid | Warm, slightly dry | Warm, moderate arid |
| Average annual temperature (°C) | 8.1 | 8.1 | 8.1 | 8.9 | 9.2 |
| Average annual precipitation (mm) | 623 | 623 | 623 | 596 | 593 |
| Prevalent soil type | Cambisol, sandy loam brown | Cambisol, sandy loam brown | Medium-weight, mildly humid clay-loam brown | Medium-grained black floodplain from debris | Loamy luvic chernozem |
| Trial area (ha) | 7.6 | 14 | 15 | 4.38 | 2.9 (1st trial); 0.52 (2nd trial) |
| No. of plots (plot size in ha) | 10 (0.5) | 10 (0.5) | 25 (0.5) | 54 (0.054) | 12 (0.09, 1st trial); 20 (0.01, 2nd trial); |
| No. of pitfall traps per plot/total amount | 5/50 | 5/50 | 5/125 | 2/108 | 2/24 (1st trial); 2/40 (2nd trial) |
| GM cultivar (No. of plots) | YieldGard® MON 810 1 (5) | YieldGard® MON 810 1 (5) | YieldGard VT Rootworm/RR2™ MON 88017 1 (5) | Roundup Ready™ 2 NK 603 1 (54 2) | YieldGard® MON 810 1 (4 in 1st trial; 10 in 2nd trial) |
| Near-isogenic cultivar (No. of plots) | Monumental (5) | Monumental (5) | DK 315 (5, 5 3) | None | DKC 3871 (4, 4 in 1st field trial; 10 in 2nd field trial 4) |
| Other treatments (No. of plots) | None | None | (b) Cultivar Kipous (KWS SAAT AG) (5) (c) Cv. PR38N86 (DuPont Pioneer) (5) | None | None |
| References | [48] | [10,49] | [50] | [46] | None |
© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Skoková Habuštová, O.; Svobodová, Z.; Cagáň, Ľ.; Sehnal, F. Use of Carabids for the Post-Market Environmental Monitoring of Genetically Modified Crops. Toxins 2017, 9, 121. https://doi.org/10.3390/toxins9040121
Skoková Habuštová O, Svobodová Z, Cagáň Ľ, Sehnal F. Use of Carabids for the Post-Market Environmental Monitoring of Genetically Modified Crops. Toxins. 2017; 9(4):121. https://doi.org/10.3390/toxins9040121
Chicago/Turabian StyleSkoková Habuštová, Oxana, Zdeňka Svobodová, Ľudovít Cagáň, and František Sehnal. 2017. "Use of Carabids for the Post-Market Environmental Monitoring of Genetically Modified Crops" Toxins 9, no. 4: 121. https://doi.org/10.3390/toxins9040121
APA StyleSkoková Habuštová, O., Svobodová, Z., Cagáň, Ľ., & Sehnal, F. (2017). Use of Carabids for the Post-Market Environmental Monitoring of Genetically Modified Crops. Toxins, 9(4), 121. https://doi.org/10.3390/toxins9040121