
Detection of Abrin Holotoxin Using Novel Monoclonal Antibodies



 and
and
Abstract
:1. Introduction
2. Results
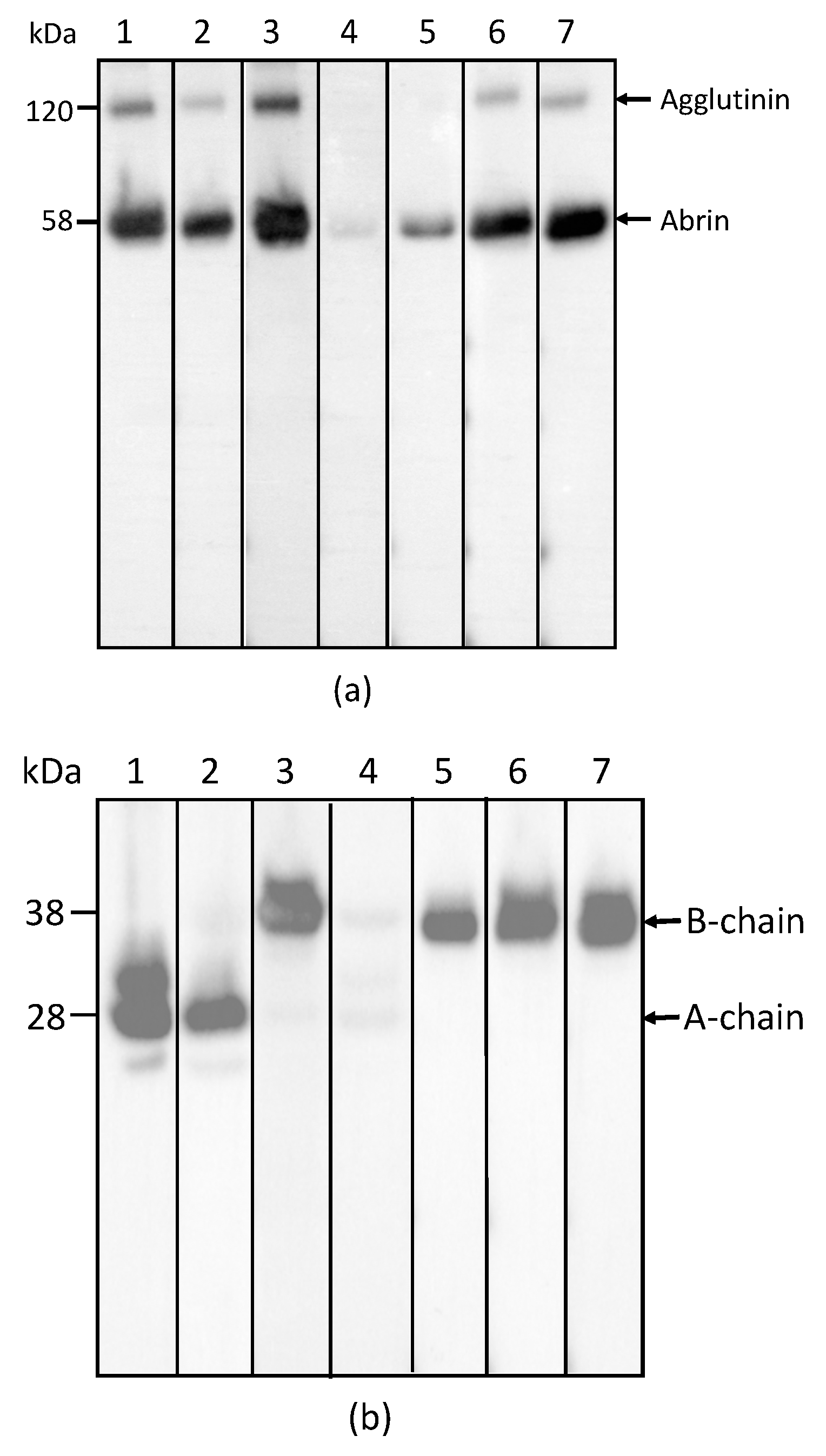
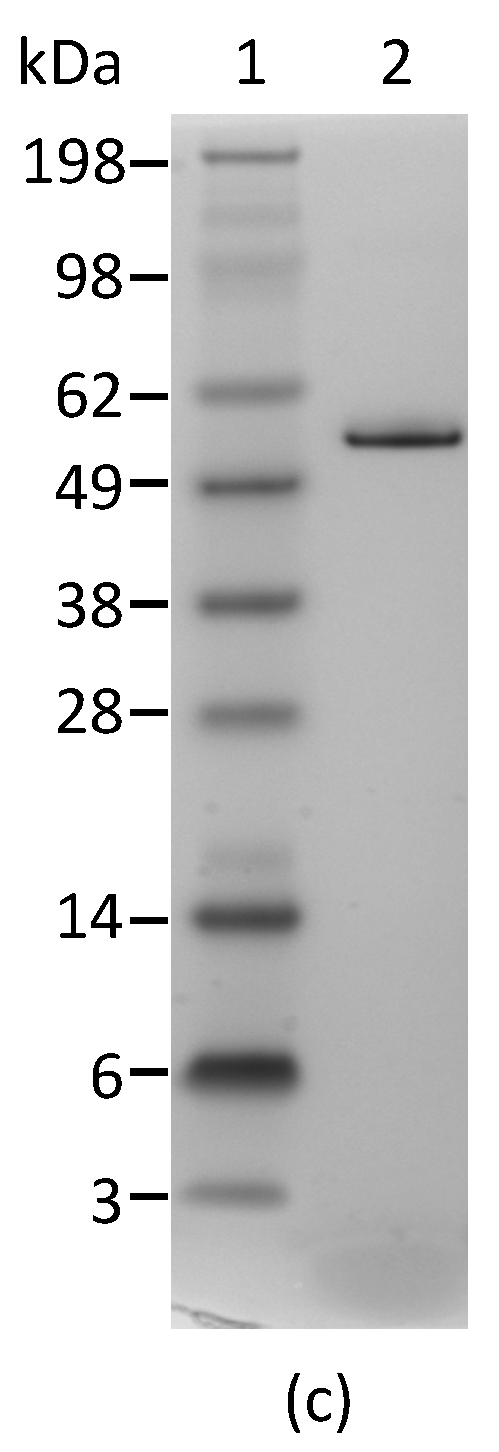
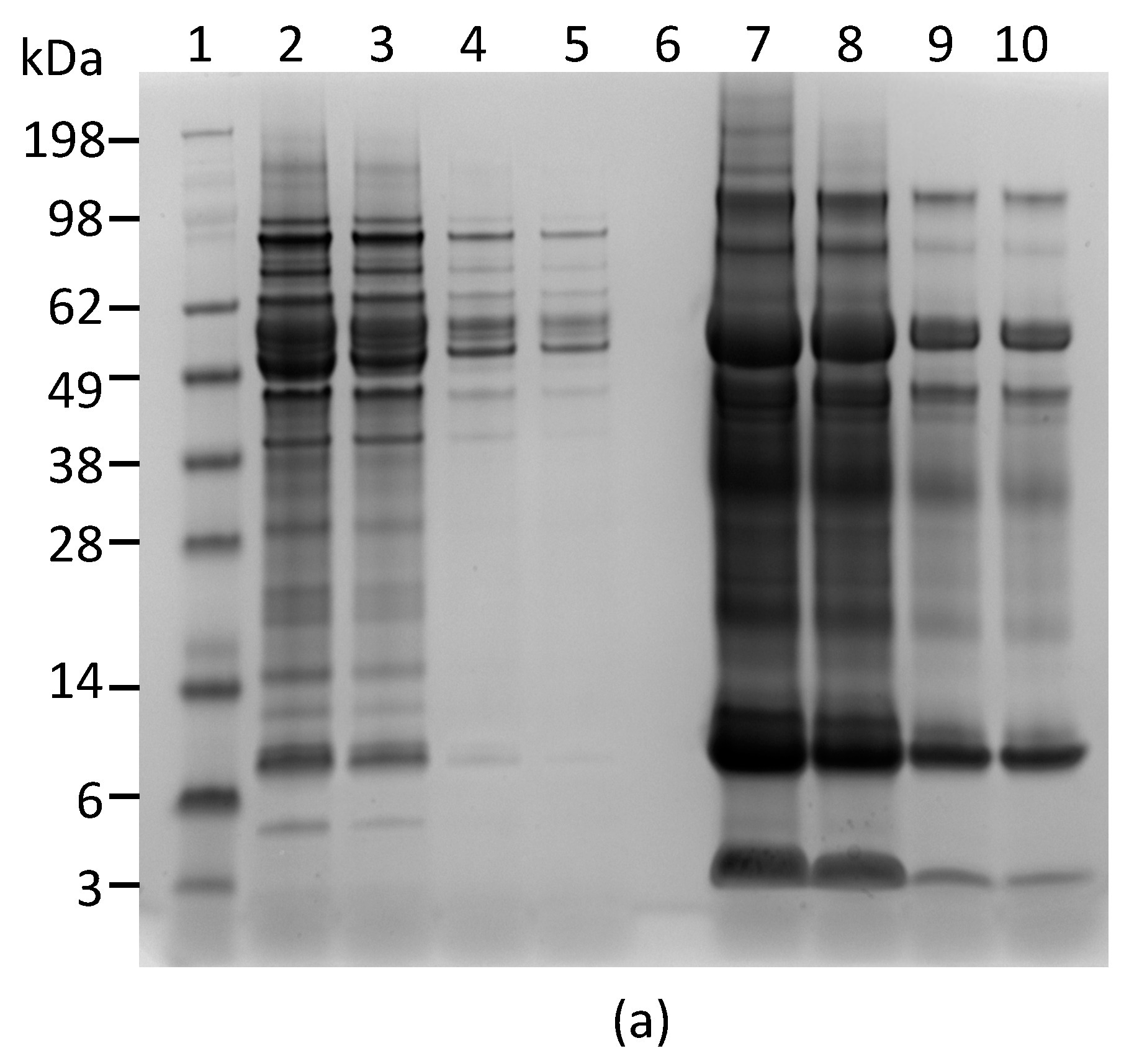
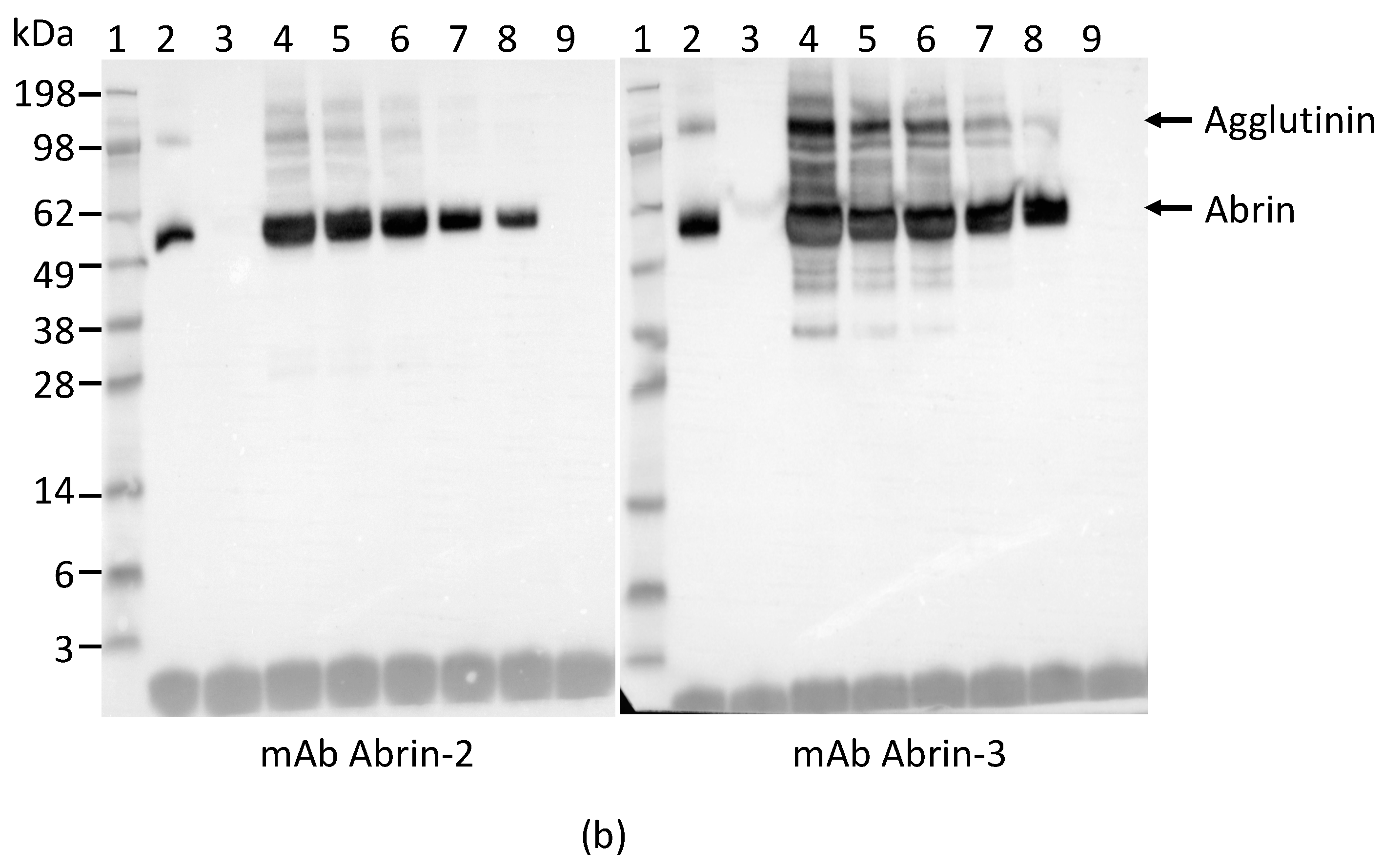
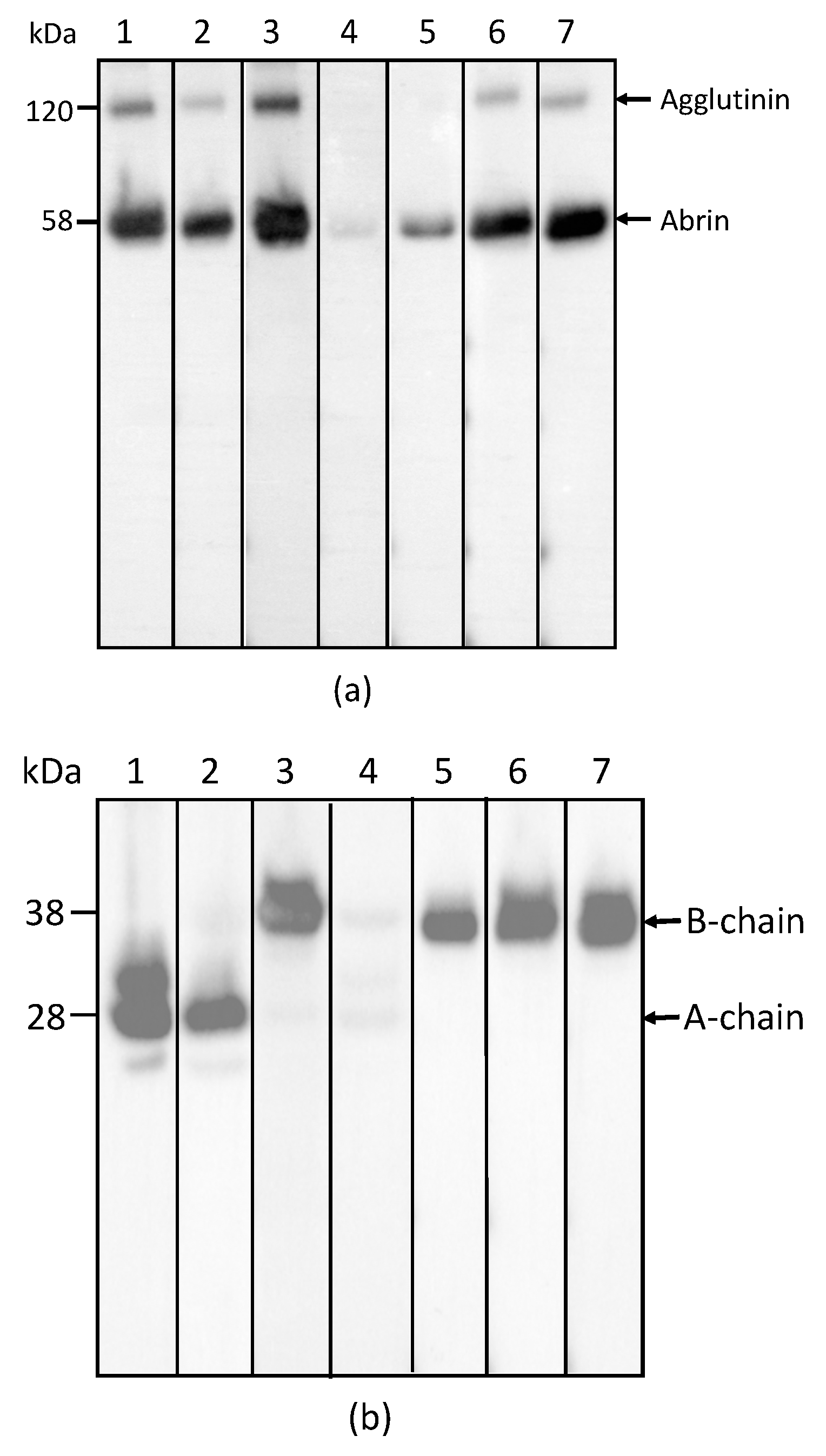
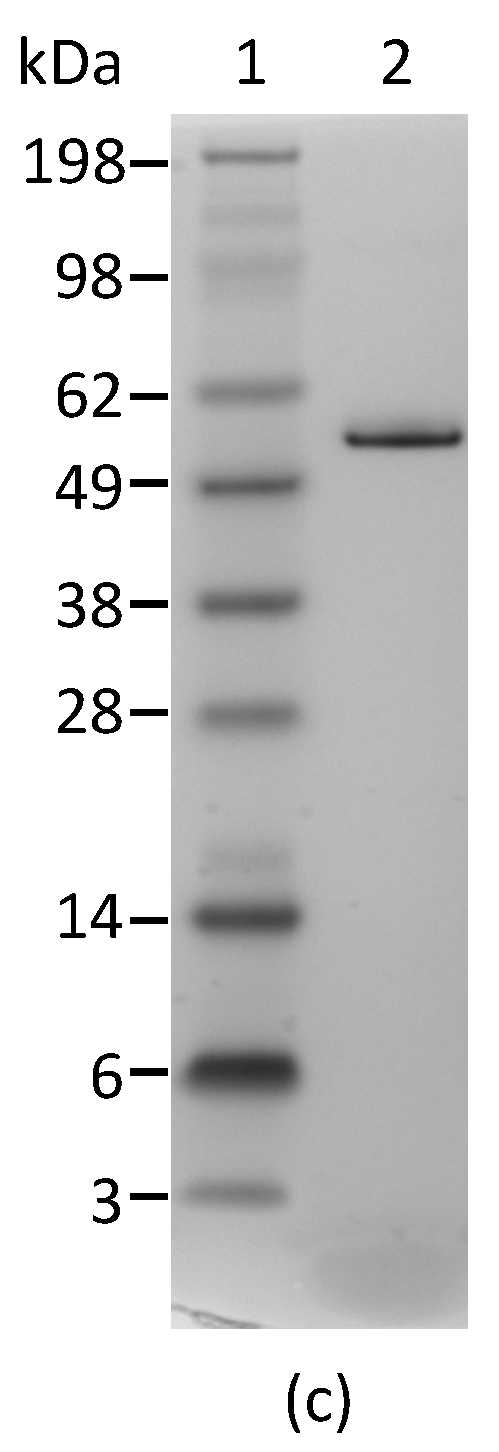
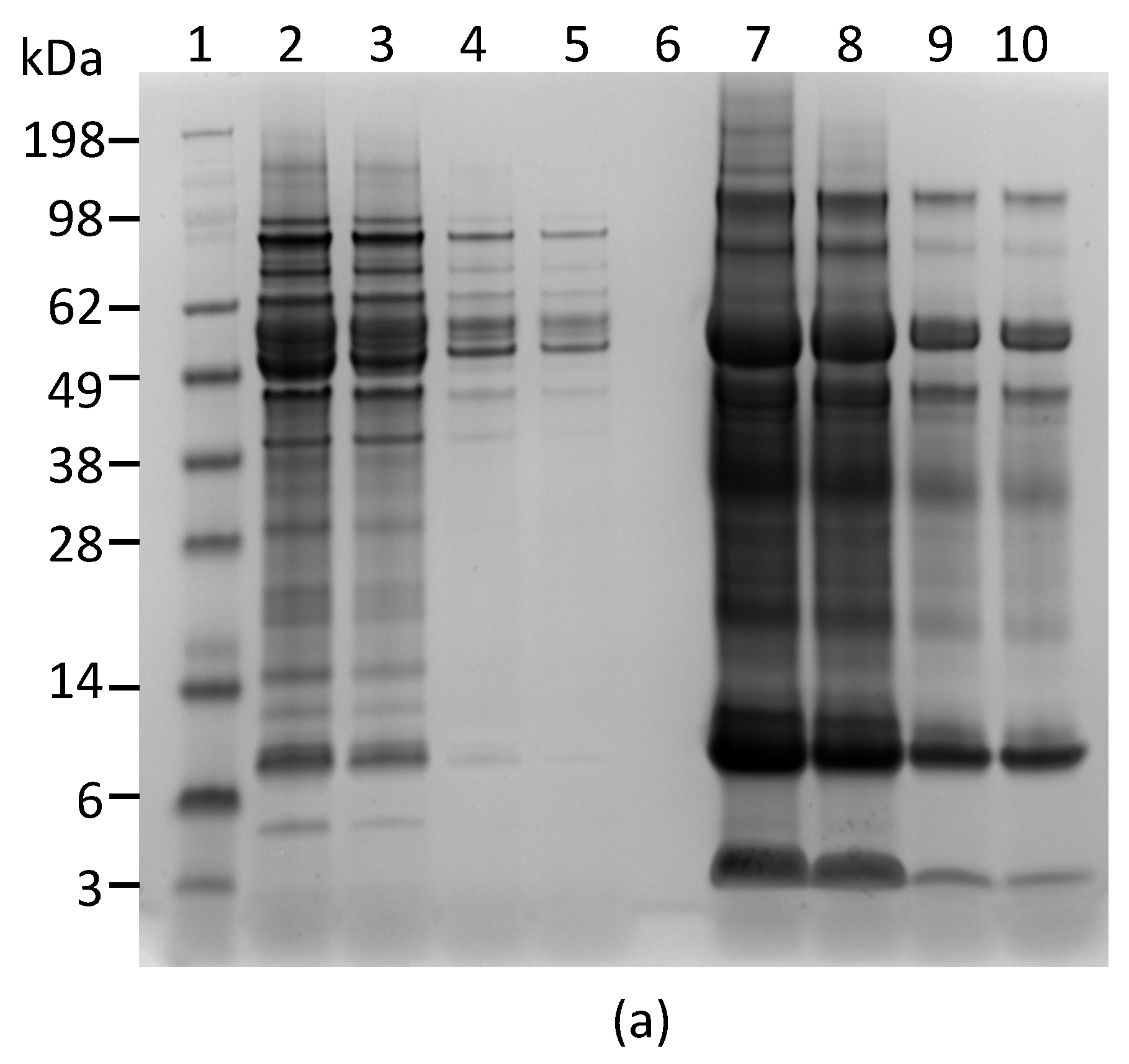
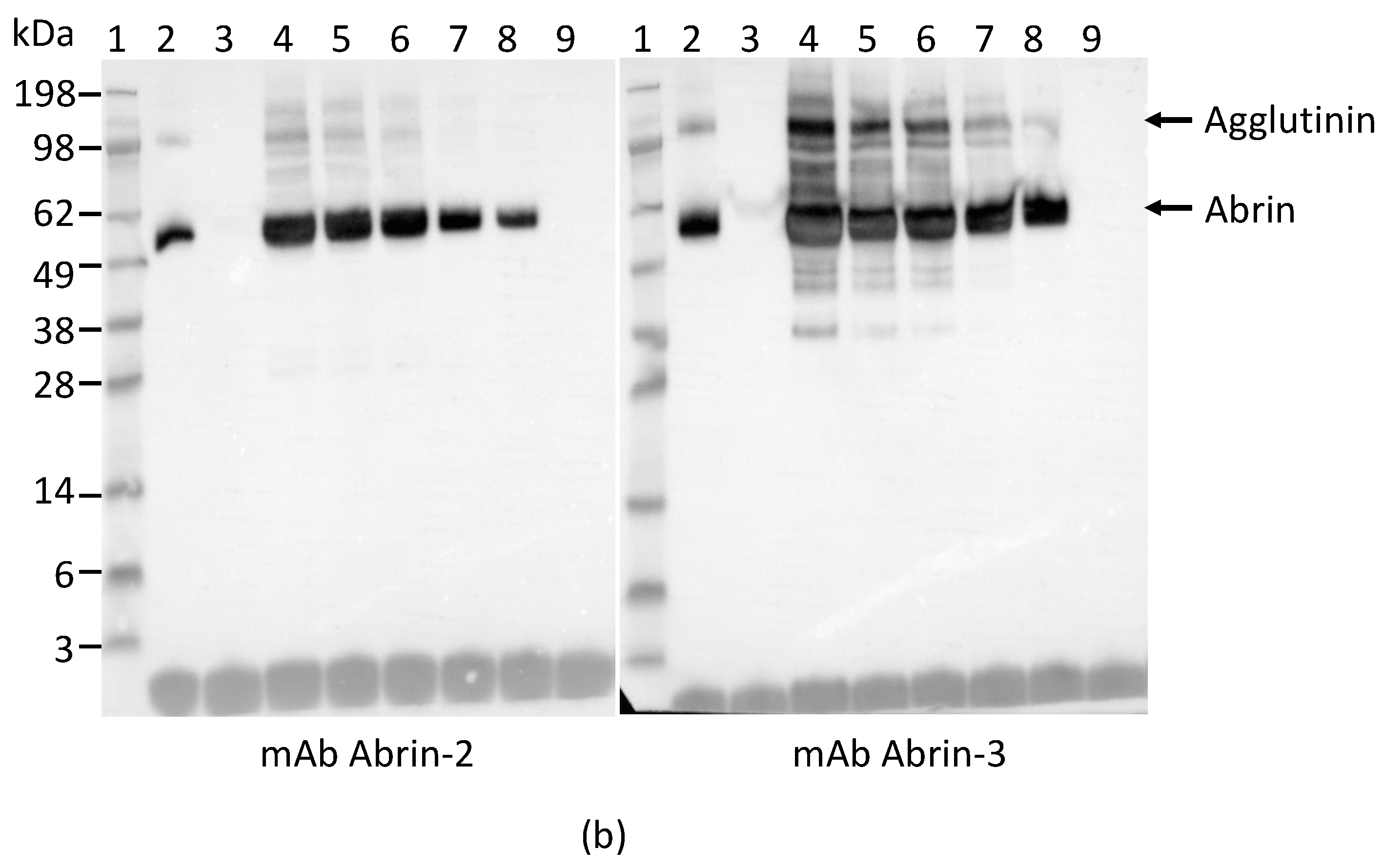
2.1. Development and Characterization of Monoclonal Antibodies against Abrin
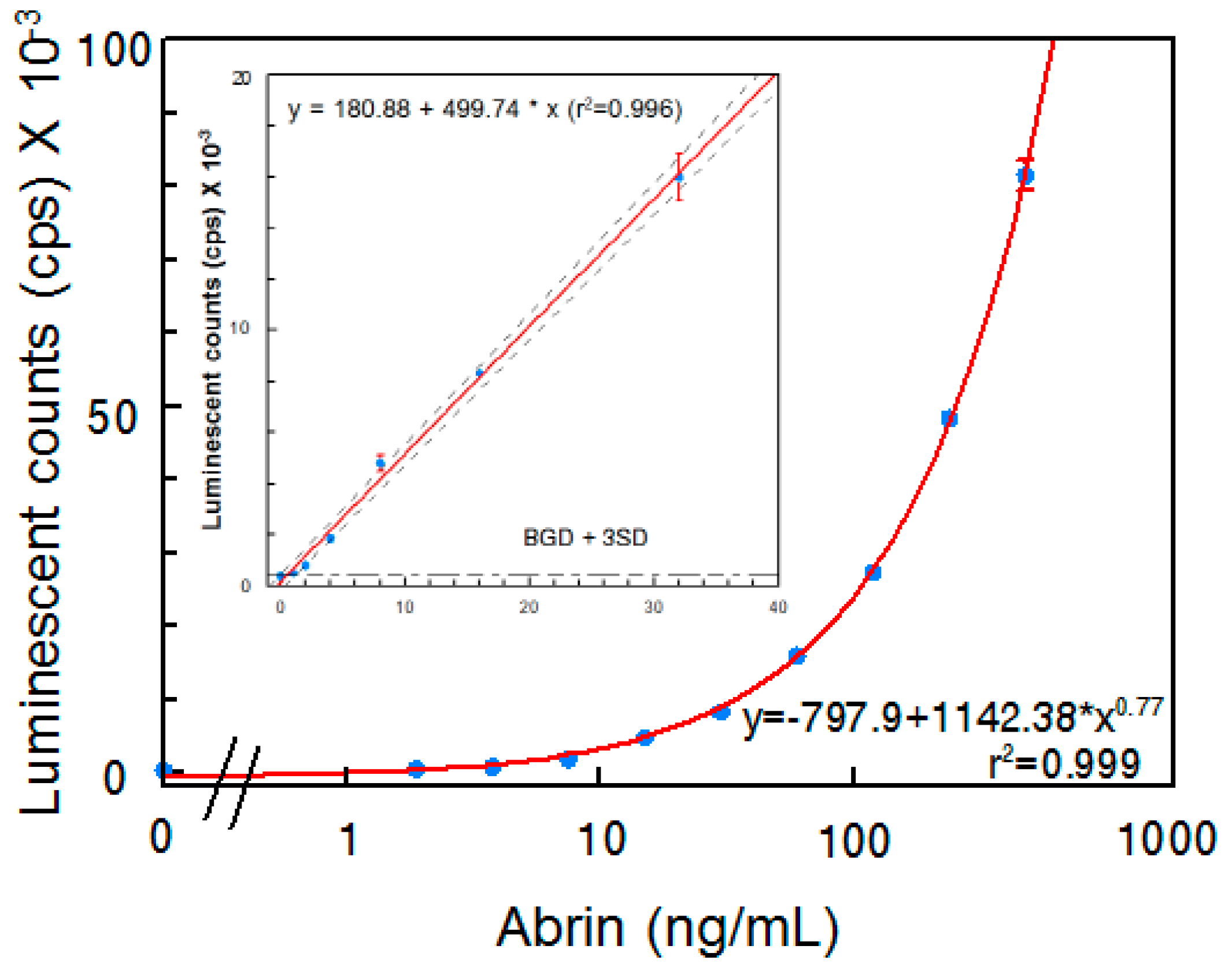
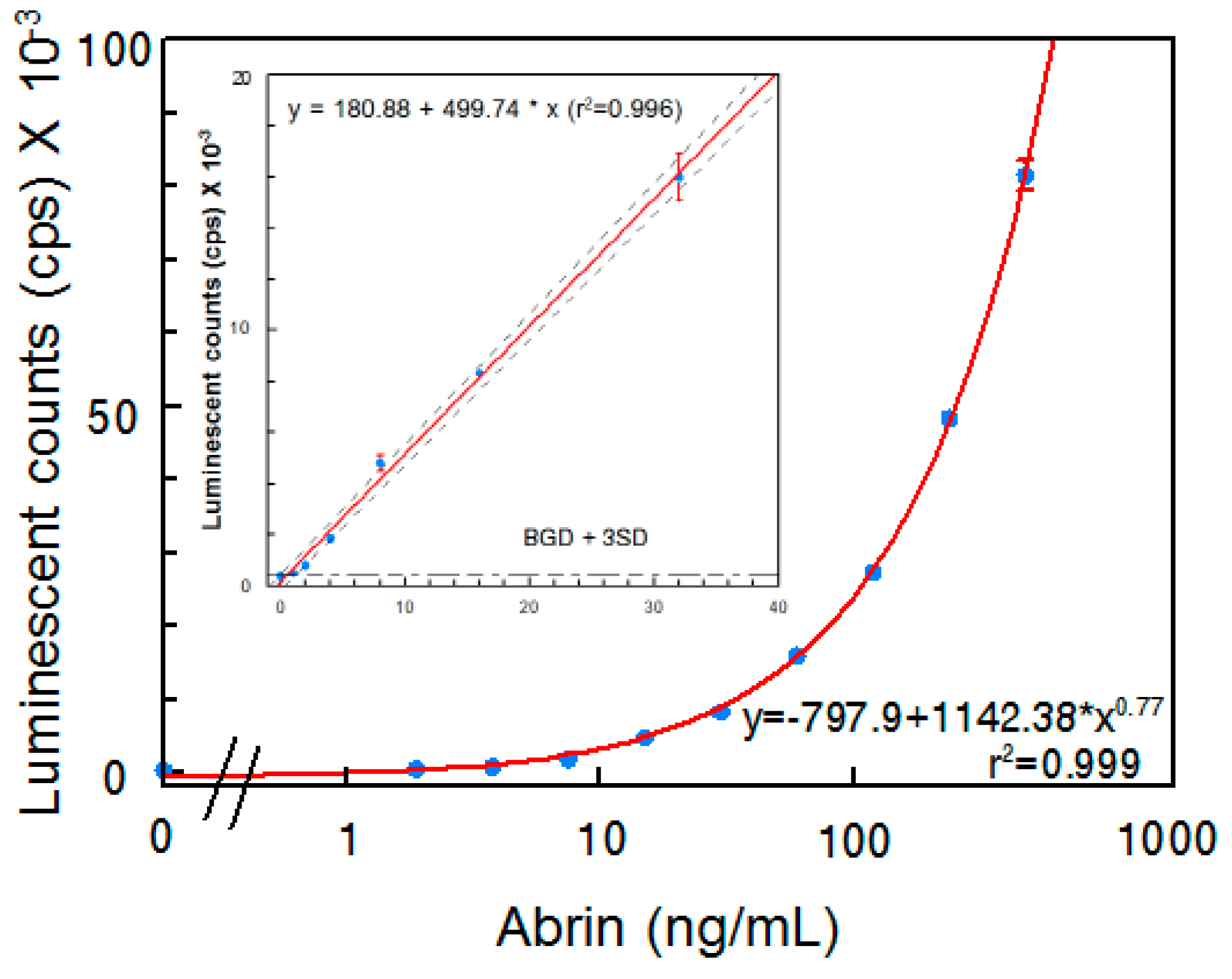
2.2. Establishment of an ELISA that Detects Abrin Holotoxin
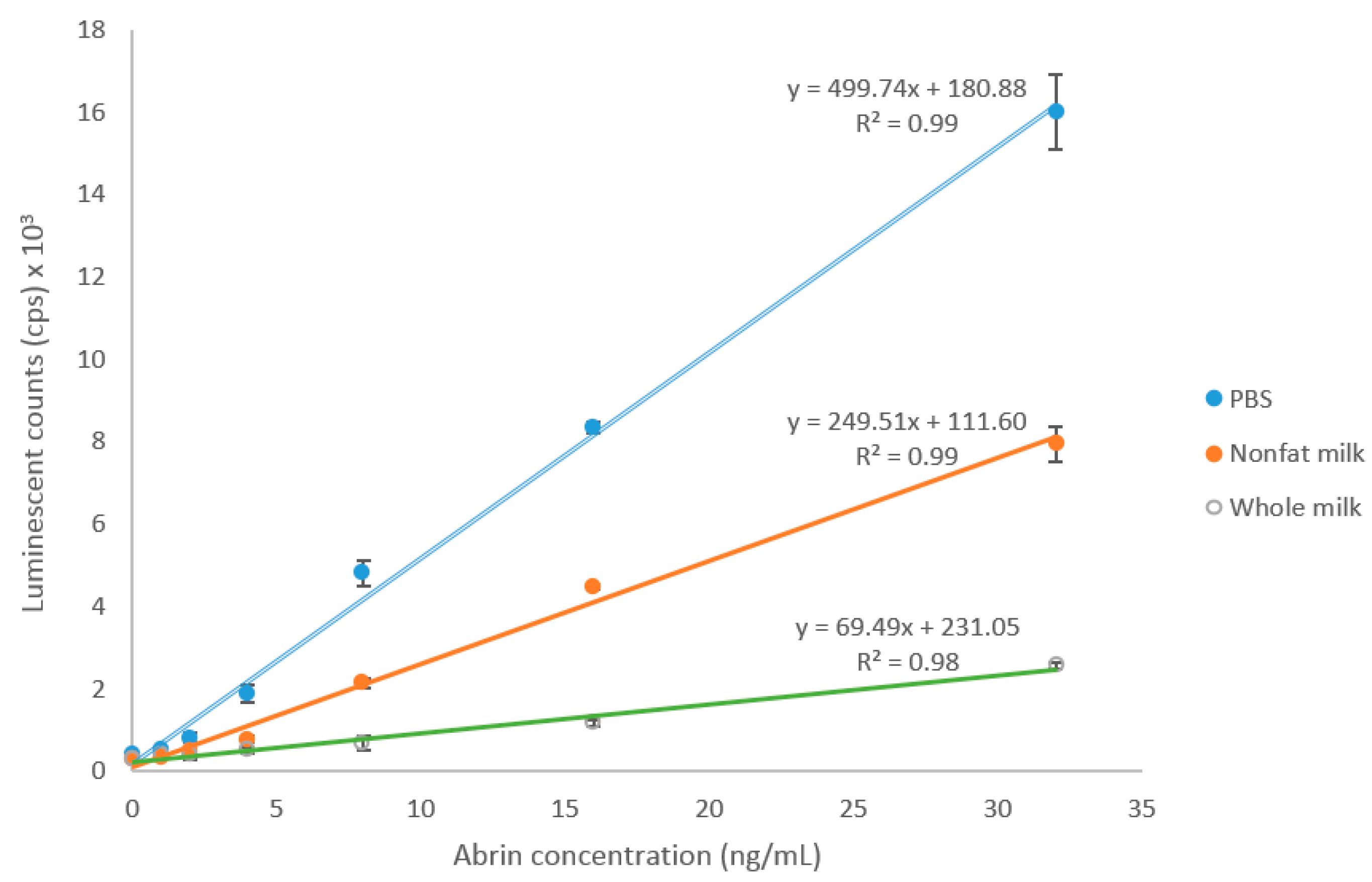
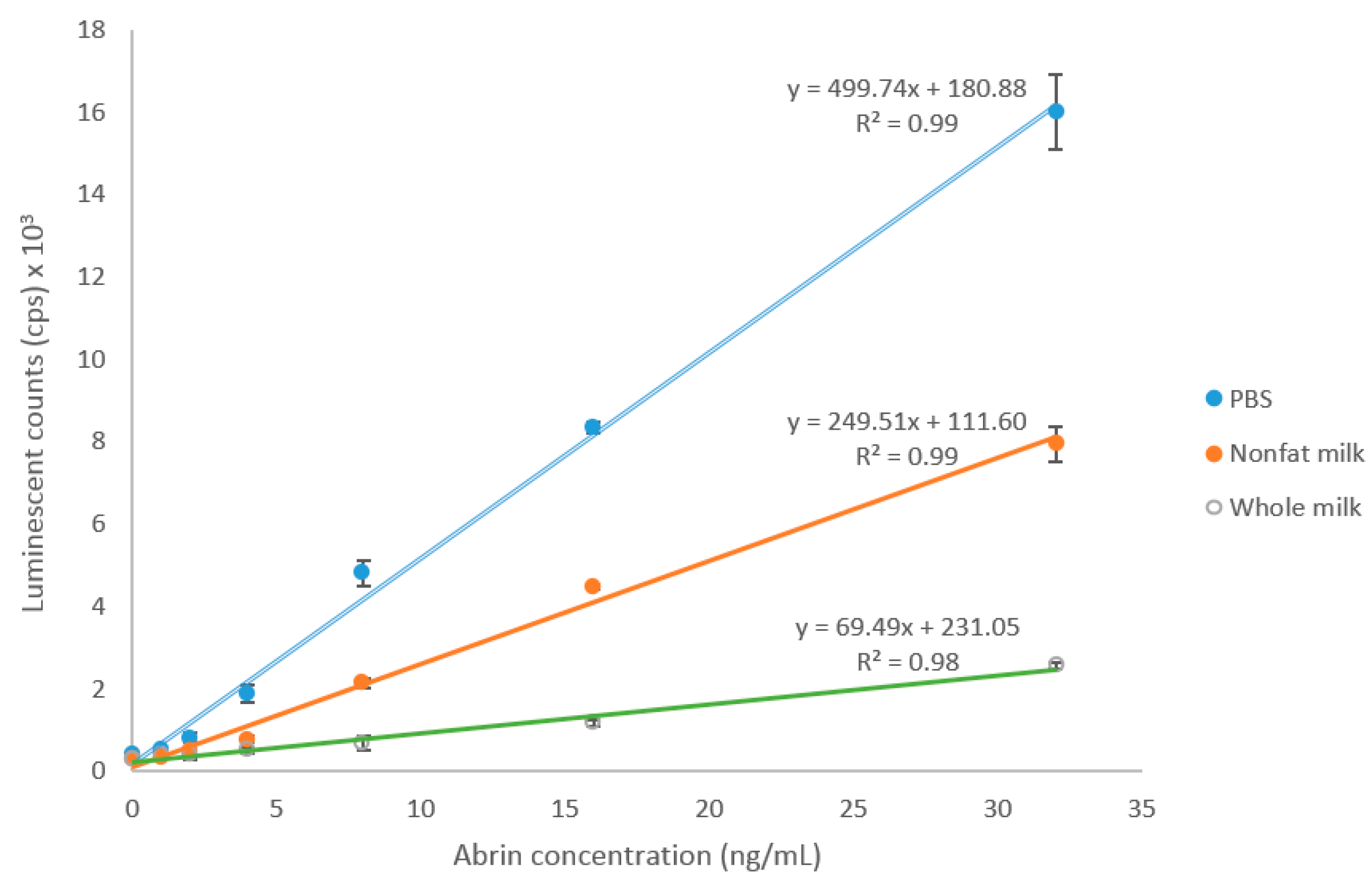
2.3. Detection of Abrin Produced by Plants and Spiked in Milk
3. Discussion
4. Materials and Methods
4.1. Ethics Statement
4.2. Monoclonal Antibody Production and Purification
4.3. Enzyme-Linked Immunosorbent Assay (ELISA)
4.4. Polyacrylamide Gel Electrophoresis (PAGE) and Western Blot Analyses
4.5. Preparation of Plant Seed Extracts
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Wood, K.A.; Lord, J.M.; Wawrzynczak, E.J.; Piatak, M. Preproabrin: genomic cloning, characterisation and the expression of the A-chain in Escherichia coli. Eur. J. Biochem. 1991, 198, 723–732. [Google Scholar] [CrossRef] [PubMed]
- Tahirov, T.H.; Lu, T.H.; Liaw, Y.C.; Chen, Y.L.; Lin, J.Y. Crystal structure of abrin-a at 2.14 a. J. Mol. Biol. 1995, 250, 354–367. [Google Scholar] [CrossRef] [PubMed]
- Endo, Y.; Mitsui, K.; Motizuki, M.; Tsurugi, K. The mechanism of action of ricin and related toxic lectins on eukaryotic ribosomes. The site and the characteristics of the modification in 28 s ribosomal RNA caused by the toxins. J. Biol. Chem. 1987, 262, 5908–5912. [Google Scholar] [PubMed]
- Endo, Y.; Tsurugi, K. RNA N-glycosidase activity of ricin a-chain. Mechanism of action of the toxic lectin ricin on eukaryotic ribosomes. J. Biol. Chem. 1987, 262, 8128–8130. [Google Scholar] [PubMed]
- Sandvig, K.; van Deurs, B. Entry of ricin and Shiga toxin into cells: Molecular mechanisms and medical perspectives. EMBO J. 2000, 19, 5943–5950. [Google Scholar] [CrossRef] [PubMed]
- Bagaria, A.; Surendranath, K.; Ramagopal, U.A.; Ramakumar, S.; Karande, A.A. Structure-function analysis and insights into the reduced toxicity of Abrus precatorius agglutinin I in relation to abrin. J. Biol. Chem. 2006, 281, 34465–34474. [Google Scholar] [CrossRef] [PubMed]
- Lin, J.Y.; Lee, T.C.; Hu, S.T.; Tung, T.C. Isolation of four isotoxic proteins and one agglutinin from jequiriti bean (Abrus precatorius). Toxicon 1981, 19, 41–51. [Google Scholar] [PubMed]
- Kimura, M.; Sumizawa, T.; Funatsu, G. The complete amino acid sequences of the b-chains of abrin-a and abrin-b, toxic proteins from the seeds of Abrus precatorius. Biosci. Biotechnol. Biochem. 1993, 57, 166–169. [Google Scholar] [CrossRef] [PubMed]
- Olsnes, S. The history of ricin, abrin and related toxins. Toxicon 2004, 44, 361–370. [Google Scholar] [CrossRef] [PubMed]
- Dickers, K.J.; Bradberry, S.M.; Rice, P.; Griffiths, G.D.; Vale, J.A. Abrin poisoning. Toxicol. Rev. 2003, 22, 137–142. [Google Scholar] [CrossRef] [PubMed]
- Gadadhar, S.; Karande, A.A. Abrin immunotoxin: Targeted cytotoxicity and intracellular trafficking pathway. PLoS ONE 2013, 8, e58304. [Google Scholar] [CrossRef] [PubMed]
- Jang, D.H.; Hoffman, R.S.; Nelson, L.S. Attempted suicide, by mail order: Abrus precatorius. J. Med. Toxicol. 2010, 6, 427–430. [Google Scholar] [CrossRef] [PubMed]
- Alhamdani, M.; Brown, B.; Narula, P. Abrin poisoning in an 18-month-old child. Am. J. Case Rep. 2015, 16, 146–148. [Google Scholar] [PubMed]
- Robb, C.S. The analysis of abrin in food and beverages. TrAC Trends Anal. Chem. 2015, 67, 100–106. [Google Scholar] [CrossRef]
- Garber, E.A.; Walker, J.L.; O’Brien, T.W. Detection of abrin in food using enzyme-linked immunosorbent assay and electrochemiluminescence technologies. J. Food Prot. 2008, 71, 1868–1874. [Google Scholar] [CrossRef]
- Liu, X.; Zhao, Y.; Sun, C.; Wang, X.; Wang, X.; Zhang, P.; Qiu, J.; Yang, R.; Zhou, L. Rapid detection of abrin in foods with an up-converting phosphor technology-based lateral flow assay. Sci. Rep. 2016, 6, 34926. [Google Scholar] [CrossRef] [PubMed]
- Wein, L.M.; Liu, Y.F. Analyzing a bioterror attack on the food supply: The case of botulinum toxin in milk. Proc. Natl. Acad. Sci. USA 2005, 102, 9984–9989. [Google Scholar] [CrossRef] [PubMed]
- Goldman, E.R.; Anderson, G.P.; Zabetakis, D.; Walper, S.; Liu, J.L.; Bernstein, R.; Calm, A.; Carney, J.P.; O’Brien, T.W.; Walker, J.L.; et al. Llama-derived single domain antibodies specific for Abrus agglutinin. Toxins 2011, 3, 1405–1419. [Google Scholar] [CrossRef] [PubMed]
- Zhou, Y.; Tian, X.L.; Li, Y.S.; Pan, F.G.; Zhang, Y.Y.; Zhang, J.H.; Wang, X.R.; Ren, H.L.; Lu, S.Y.; Li, Z.H.; et al. Development of a monoclonal antibody-based sandwich-type enzyme-linked immunosorbent assay (ELISA) for detection of abrin in food samples. Food Chem. 2012, 135, 2661–2665. [Google Scholar] [CrossRef] [PubMed]
- Bradberry, S.M.; Dickers, K.J.; Rice, P.; Griffiths, G.D.; Vale, J.A. Ricin poisoning. Toxicol. Rev. 2003, 22, 65–70. [Google Scholar] [CrossRef] [PubMed]
- Federal Select Agent Program. Select Agents and Toxins List. Available online: https://www.selectagents.gov/selectagentsandtoxinslist.html (accessed on 24 July 2017).
- Li, X.B.; Yang, W.; Zhang, Y.; Zhang, Z.G.; Kong, T.; Li, D.N.; Tang, J.J.; Liu, L.; Liu, G.W.; Wang, Z. Preparation and identification of monoclonal antibody against abrin-a. J. Agric. Food Chem. 2011, 59, 9796–9799. [Google Scholar] [CrossRef] [PubMed]
- Olsnes, S.; Pihl, A. Treatment of abrin and ricin with mercaptoethanol opposite effects on their toxicity in mice and their ability to inhibit protein synthesis in a cell-free system. FEBS Lett. 1972, 28, 48–50. [Google Scholar] [CrossRef]
- United States Department of Agriculture. Milk Production, Disposition, and Income Annual Summary. Available online: http://usda.mannlib.cornell.edu/usda/current/milkproddi/milkproddi-04-27-2017.pdf (accessed on 22 August 2017).
- He, X.; McMahon, S.; Skinner, C.; Merrill, P.; Scotcher, M.C.; Stanker, L.H. Development and characterization of monoclonal antibodies against Shiga toxin 2 and their application for toxin detection in milk. J. Immunol. Methods 2013, 389, 18–28. [Google Scholar] [CrossRef] [PubMed]
- Bigbee, W.L.; Vanderlaan, M.; Fong, S.S.; Jensen, R.H. Monoclonal antibodies specific for the M- and N-forms of human glycophorinA. Mol. Immunol. 1983, 20, 1353–1362. [Google Scholar] [CrossRef]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Antibody | Isotype | Subunit Specificity |
|---|---|---|
| Abrin-1 | IgG2a, kappa | A |
| Abrin-2 | IgG2a, kappa | A |
| Abrin-3 | IgG2b, kappa | B |
| Abrin-4 | IgG1, kappa | Nd * |
| Abrin-5 | IgG2b, kappa | B |
| Abrin-6 | IgG1, kappa | B |
| Abrin-7 | IgG2a, kappa | B |
| Plant | Extract (wt/vol) | Average ELISA Reading (cps) | SD |
|---|---|---|---|
| Rosary pea | 0.02% | 43,285 | 3155 |
| 0.20% | 74,465 | 2335 | |
| 2.00% | 98,690 | 930 | |
| Castor bean | 0.02% | 160 | 10 |
| 0.20% | 210 | 30 | |
| 2.00% | 150 | 30 | |
| PBS | 0 | 227 | 30 |
© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
He, X.; Patfield, S.; Cheng, L.W.; Stanker, L.H.; Rasooly, R.; McKeon, T.A.; Zhang, Y.; Brandon, D.L. Detection of Abrin Holotoxin Using Novel Monoclonal Antibodies. Toxins 2017, 9, 386. https://doi.org/10.3390/toxins9120386
He X, Patfield S, Cheng LW, Stanker LH, Rasooly R, McKeon TA, Zhang Y, Brandon DL. Detection of Abrin Holotoxin Using Novel Monoclonal Antibodies. Toxins. 2017; 9(12):386. https://doi.org/10.3390/toxins9120386
Chicago/Turabian StyleHe, Xiaohua, Stephanie Patfield, Luisa W. Cheng, Larry H. Stanker, Reuven Rasooly, Thomas A. McKeon, Yuzhu Zhang, and David L. Brandon. 2017. "Detection of Abrin Holotoxin Using Novel Monoclonal Antibodies" Toxins 9, no. 12: 386. https://doi.org/10.3390/toxins9120386
APA StyleHe, X., Patfield, S., Cheng, L. W., Stanker, L. H., Rasooly, R., McKeon, T. A., Zhang, Y., & Brandon, D. L. (2017). Detection of Abrin Holotoxin Using Novel Monoclonal Antibodies. Toxins, 9(12), 386. https://doi.org/10.3390/toxins9120386