
Combined Effects of Lipophilic Phycotoxins (Okadaic Acid, Azapsiracid-1 and Yessotoxin) on Human Intestinal Cells Models

 , , and
, , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Cell Cultures
2.2. Toxins
2.3. Neutral Red Uptake Assay
2.4. Combinations of Toxins
2.5. Data Analysis
3. Results
3.1. Individual Cytotoxicity of OA, AZA-1 and YTX
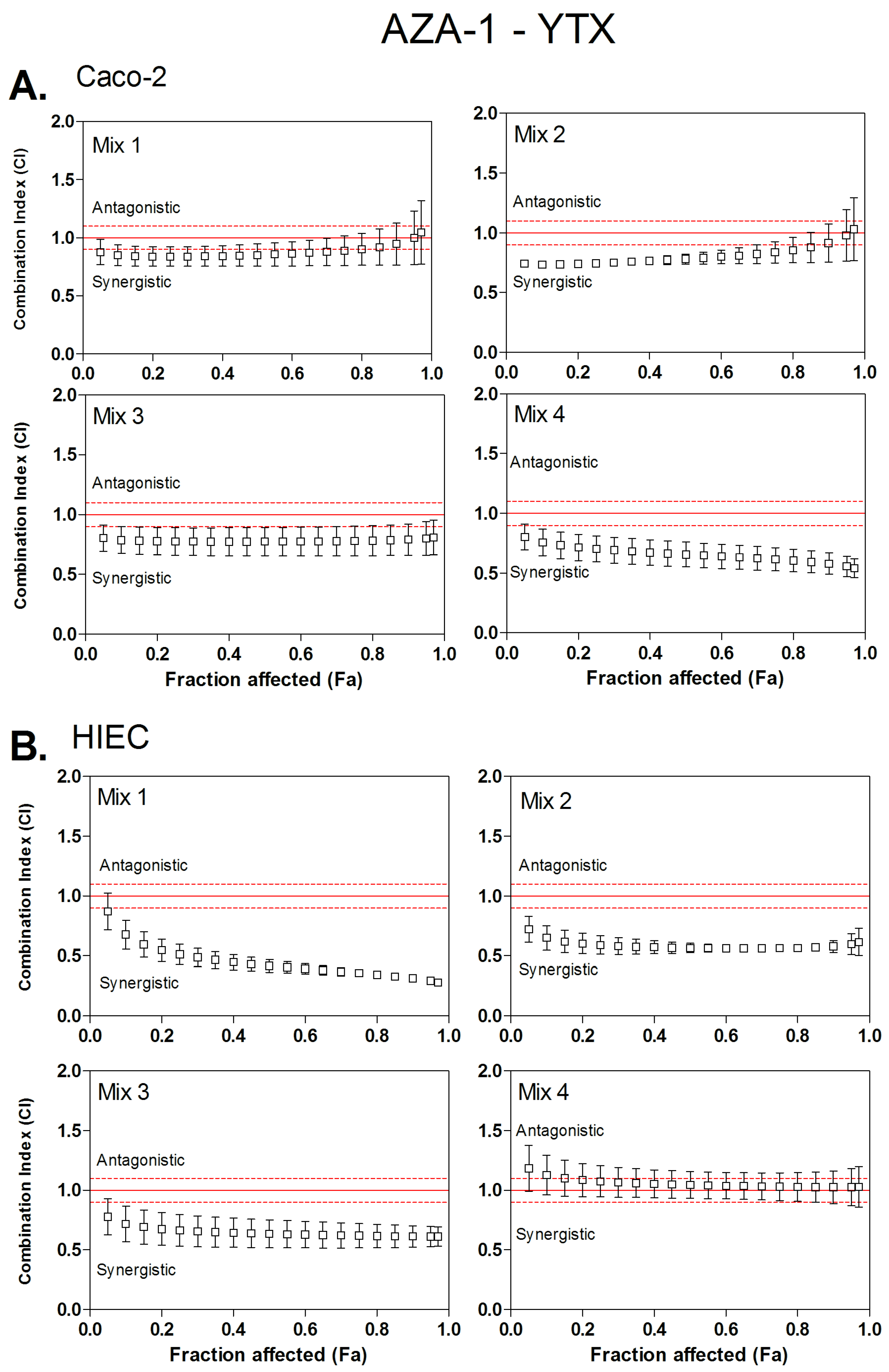
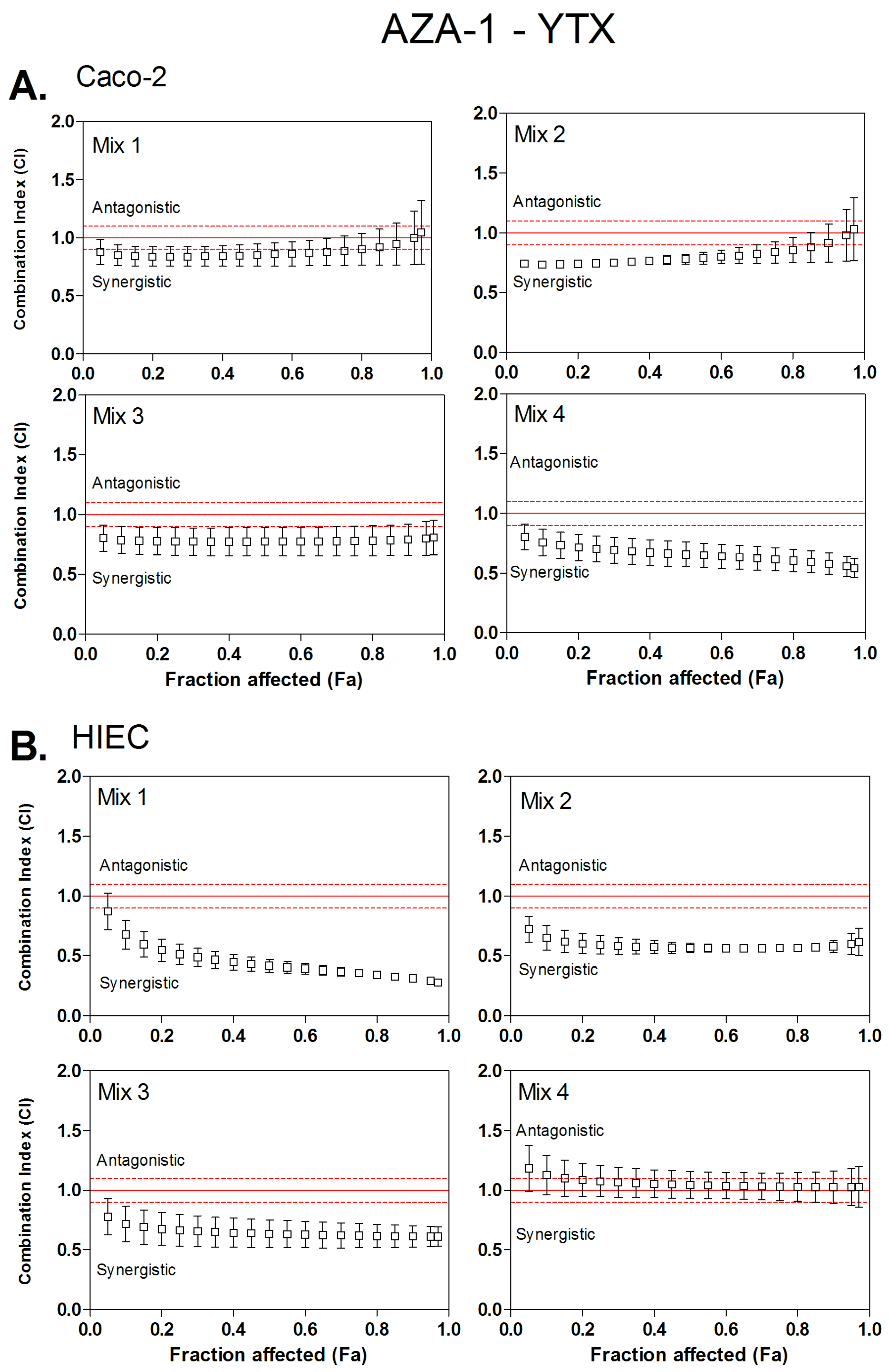
3.2. Combined Cytotoxicity of AZA-1 and YTX
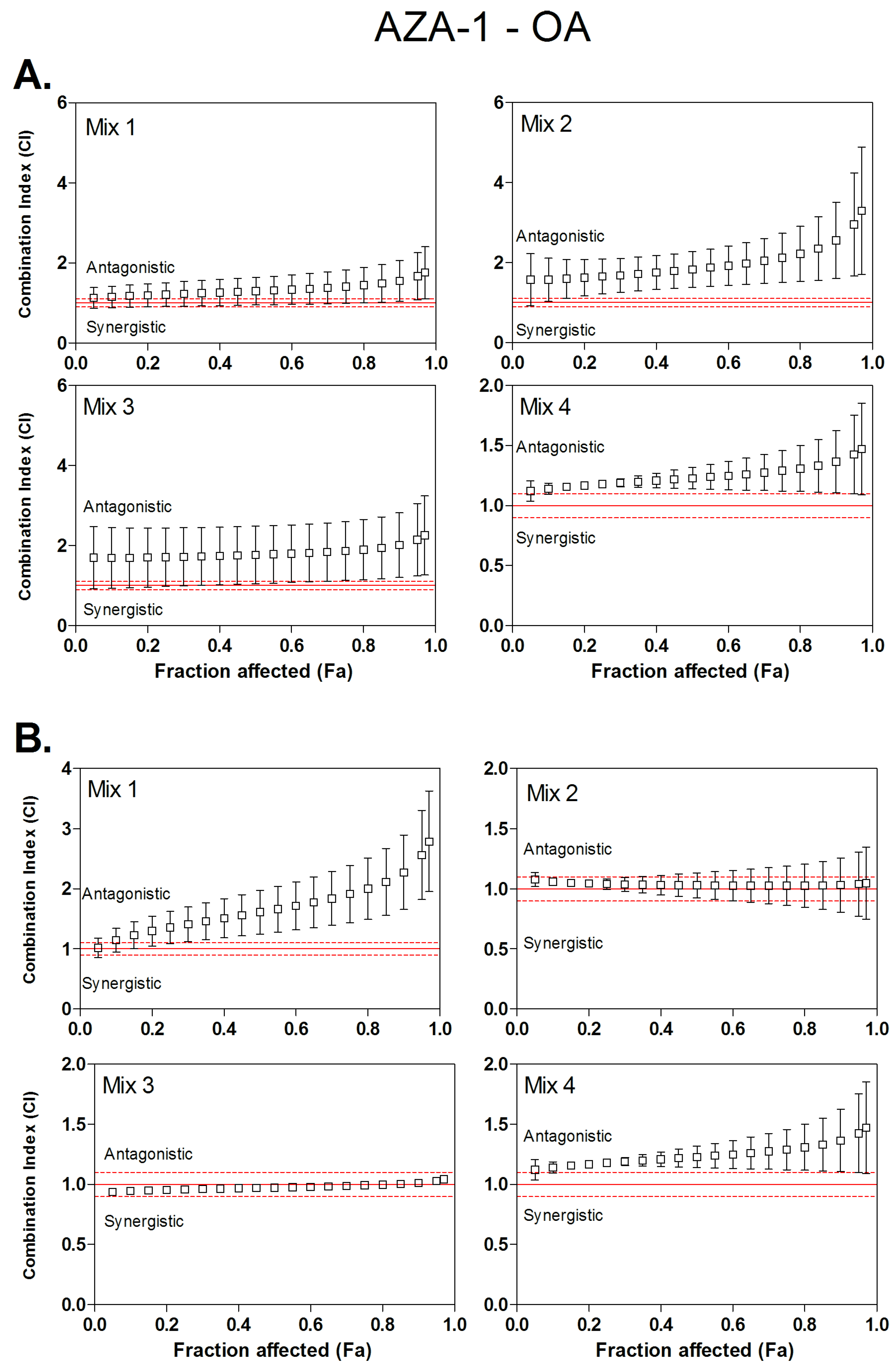
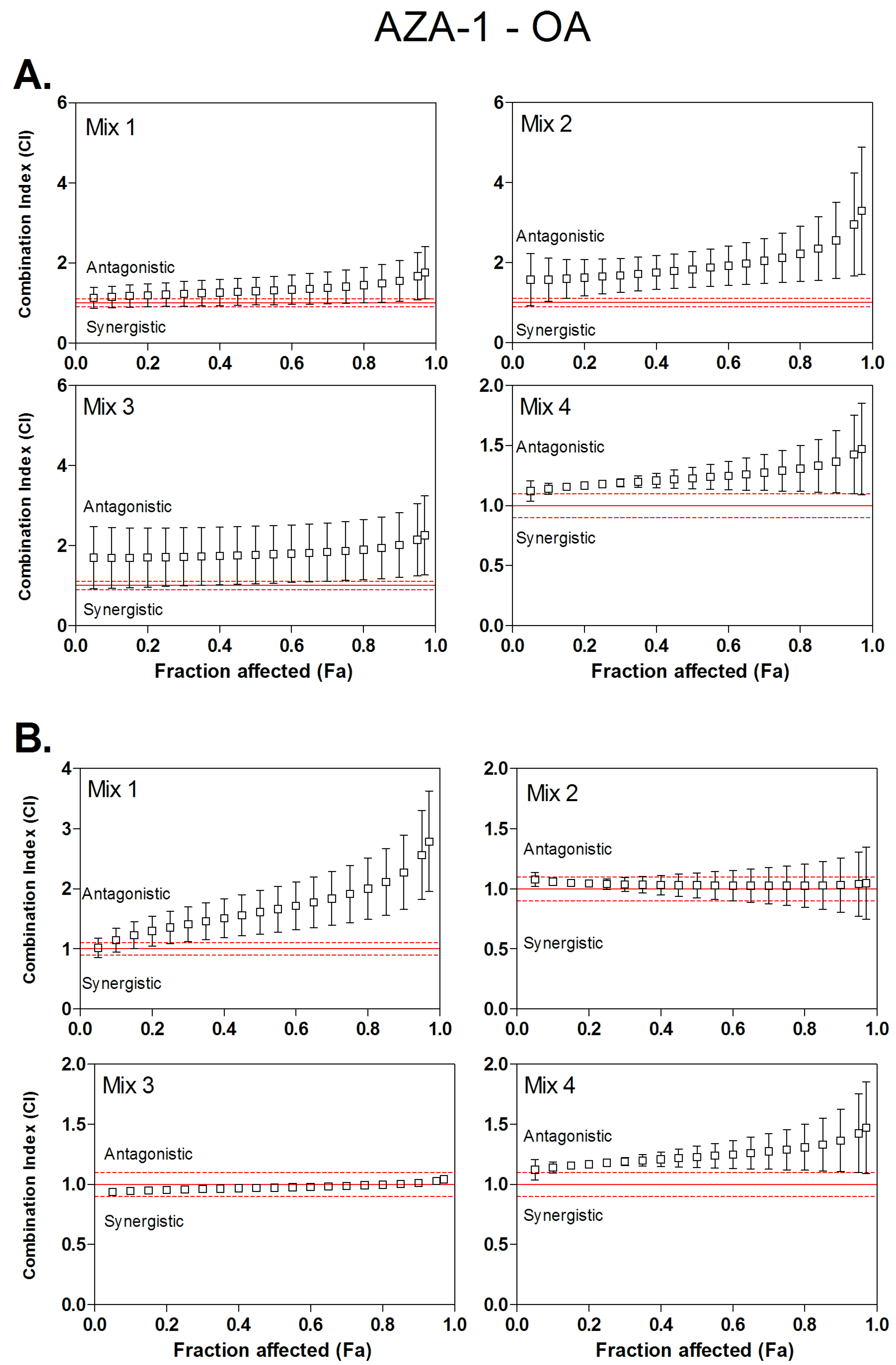
3.3. Combined Cytotoxicity of AZA-1 and OA
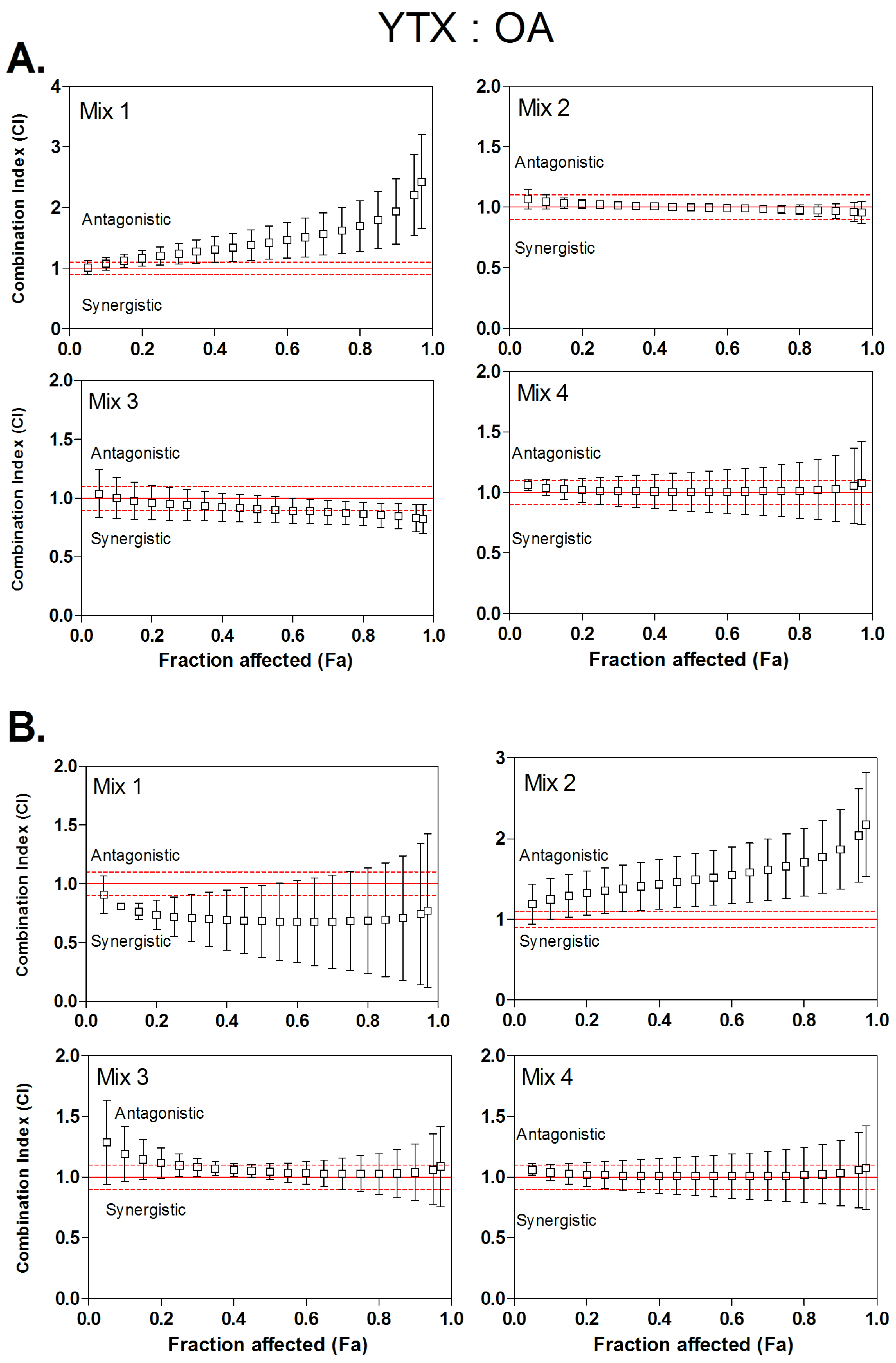
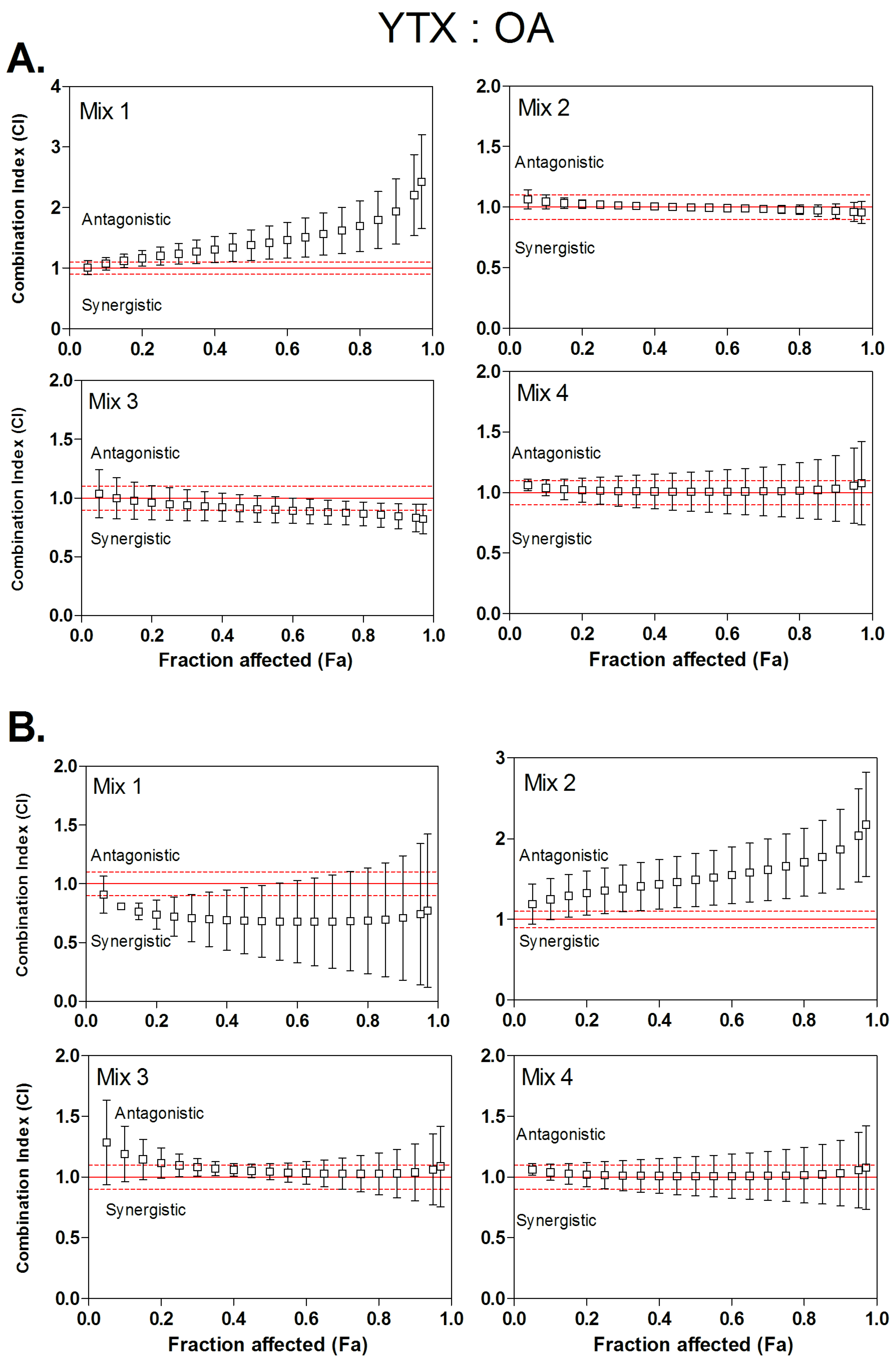
3.4. Combined Cytotoxicity of YTX and OA
4. Discussion
5. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Rossini, G.P.; Hess, P. Phycotoxins: Chemistry, mechanisms of action and shellfish poisoning. In Molecular, Clinical and Environmental Toxicology; Luch, A., Ed.; Birkhäuser: Basel, Switzerland, 2010; pp. 65–122. [Google Scholar]
- Marine Biotoxins in Shellfish—Summary on Regulated Marine Biotoxins. Available online: http://www.elika.net/datos/articulos/archivo_en448/marinebiotoxins_en.pdf (accessed on 19 February 2016).
- Dominguez, H.J.; Paz, B.; Daranas, A.H.; Norte, M.; Franco, J.M.; Fernández, J.J. Dinoflagellate polyether within the yessotoxin, pectenotoxin and okadaic acid toxin groups: Characterization, analysis and human health implications. Toxicon 2010, 56, 191–217. [Google Scholar] [CrossRef] [PubMed]
- Yasumoto, T.; Murata, M.; Oshima, Y.; Sano, M.; Matsumoto, G.K.; Clardy, J. Diarrhetic shellfish toxins. Tetrahedron 1985, 41, 1019–1025. [Google Scholar] [CrossRef]
- Dounay, A.B.; Forsyth, C.J. Okadaic acid: The archetypal serine/threonine protein phosphatase inhibitor. Curr. Med. Chem. 2002, 9, 1939–1980. [Google Scholar] [CrossRef] [PubMed]
- Furey, A.; O’Doherty, S.; O’Callaghan, K.; Lehane, M.; James, K.J. Azaspiracid poisoning (AZP) toxins in shellfish: Toxicological and health considerations. Toxicon Off. J. Int. Soc. Toxinol. 2010, 56, 173–190. [Google Scholar] [CrossRef] [PubMed]
- Ito, E.; Satake, M.; Ofuji, K.; Kurita, N.; McMahon, T.; James, K.; Yasumoto, T. Multiple organ damage caused by a new toxin azaspiracid, isolated from mussels produced in Ireland. Toxicon 2000, 38, 917–930. [Google Scholar] [CrossRef]
- Twiner, M.J.; Rehmann, N.; Hess, P.; Doucette, G.J. Azaspiracid shellfish poisoning: A review on the chemistry, ecology, and toxicology with an emphasis on human health impacts. Mar. Drugs 2008, 6, 39–72. [Google Scholar] [CrossRef] [PubMed]
- Gu, H.; Luo, Z.; Krock, B.; Witt, M.; Tillmann, U. Morphology, phylogeny and azaspiracid profile of Azadinium poporum (Dinophyceae) from the China Sea. Harmful Algae 2013, 64–75. [Google Scholar] [CrossRef]
- Taleb, H.; Vale, P.; Amanhir, R.; Benhadouch, A.; Sagou, R.; Chafik, A. First detection of azaspiracids in mussels in North West Africa. J. Shellfish Res. 2006, 25, 1067–1070. [Google Scholar]
- Tillmann, U.; Elbrächter, M.; Krock, B.; John, U.; Cembella, A. Azadinium spinosum gen. et sp. nov. (Dinophyceae) identified as a primary producer of azaspiracid toxins. Eur. J. Phycol. 2009, 44, 63–79. [Google Scholar] [CrossRef]
- Potvin, É.; Jeong, H.J.; Kang, N.S.; Tillmann, U.; Krock, B. First report of the photosynthetic dinoflagellate genus Azadinium in the Pacific Ocean: Morphology and molecular characterization of Azadinium cf. poporum. J. Eukaryot. Microbiol. 2012, 59, 145–156. [Google Scholar] [CrossRef] [PubMed]
- Krock, B.; Tillmann, U.; Witt, M.; Gu, H. Azaspiracid variability of Azadinium poporum (Dinophyceae) from the China Sea. Harmful Algae 2014, 36, 22–28. [Google Scholar] [CrossRef]
- Sala, G.L.; Bellocci, M.; Callegari, F.; Rossini, G.P. Azaspiracid-1 inhibits the maturation of cathepsin D in mammalian cells. Chem. Res. Toxicol. 2013, 26, 444–455. [Google Scholar] [CrossRef] [PubMed]
- Vilariño, N. Marine toxins and the cytoskeleton: Azaspiracids. FEBS J. 2008, 275, 6075–6081. [Google Scholar] [CrossRef] [PubMed]
- Paz, B.; Daranas, A.H.; Norte, M.; Riobó, P.; José, M.F.; José, J.F. Yessotoxins, a group of marine polyether toxins: An overview. Mar. Drugs 2008, 6, 73–102. [Google Scholar] [CrossRef] [PubMed]
- Rubiolo, J.A.; López-Alonso, H.; Martínez, P.; Millán, A.; Cagide, E.; Vieytes, M.R.; Vega, F.V.; Botana, L.M. Yessotoxin induces ER-stress followed by autophagic cell death in glioma cells mediated by mTOR and BNIP3. Cell. Signal. 2014, 26, 419–432. [Google Scholar] [CrossRef] [PubMed]
- Korsnes, M.S. Yessotoxin as a tool to study induction of multiple cell death pathways. Toxins 2012, 4, 568–579. [Google Scholar] [CrossRef] [PubMed]
- Callegari, F.; Rossini, G.P. Yessotoxin inhibits the complete degradation of E-cadherin. Toxicology 2008, 244, 133–144. [Google Scholar] [CrossRef] [PubMed]
- Amzil, Z.; Sibat, M.; Royer, F.; Savar, V. First report on azaspiracid and yessotoxin groups detection in French shellfish. Toxicon 2008, 52, 39–48. [Google Scholar] [CrossRef] [PubMed]
- Trainer, V.; Moore, L.; Bill, B.; Adams, N.; Harrington, N.; Borchert, J.; da Silva, D.; Eberhart, B.-T. Diarrhetic shellfish toxins and other lipophilic toxins of human health concern in Washington State. Mar. Drugs 2013, 11, 1815–1835. [Google Scholar] [CrossRef] [PubMed]
- Zamorano, R.; Marín, M.; Cabrera, F.; Figueroa, D.; Contreras, C.; Barriga, A.; Lagos, N.; García, C. Determination of the variability of both hydrophilic and lipophilic toxins in endemic wild bivalves and carnivorous gastropods from the Southern part of Chile. Food Addit. Contam. Part A 2013, 30, 1660–1677. [Google Scholar] [CrossRef] [PubMed]
- Sosa, S.; Ardizzone, M.; Beltramo, D.; Vita, F.; Dell’Ovo, V.; Barreras, A.; Yasumoto, T.; Tubaro, A. Repeated oral co-exposure to yessotoxin and okadaic acid: A short term toxicity study in mice. Toxicon 2013, 76, 94–102. [Google Scholar] [CrossRef] [PubMed]
- Aune, T.; Espenes, A.; Aasen, J.A.B.; Quilliam, M.A.; Hess, P.; Larsen, S. Study of possible combined toxic effects of azaspiracid-1 and okadaic acid in mice via the oral route. Toxicon 2012, 60, 895–906. [Google Scholar] [CrossRef] [PubMed]
- Aasen, J.A.B.; Espenes, A.; Miles, C.O.; Samdal, I.A.; Hess, P.; Aune, T. Combined oral toxicity of azaspiracid-1 and yessotoxin in female NMRI mice. Toxicon 2011, 57, 909–917. [Google Scholar] [CrossRef] [PubMed]
- Franchini, A.; Marchesini, E.; Poletti, R.; Ottaviani, E. Lethal and sub-lethal yessotoxin dose-induced morpho-functional alterations in intraperitoneal injected Swiss CD1 mice. Toxicon 2004, 44, 83–90. [Google Scholar] [CrossRef] [PubMed]
- Tubaro, A.; Sosa, S.; Carbonatto, M.; Altinier, G.; Vita, F.; Melato, M.; Satake, M.; Yasumoto, T. Oral and intraperitoneal acute toxicity studies of yessotoxin and homoyessotoxins in mice. Toxicon 2003, 41, 783–792. [Google Scholar] [CrossRef]
- Ehlers, A.; These, A.; Hessel, S.; Preiss-Weigert, A.; Lampen, A. Active elimination of the marine biotoxin okadaic acid by P-glycoprotein through an in vitro gastrointestinal barrier. Toxicol. Lett. 2014, 225, 311–317. [Google Scholar] [CrossRef] [PubMed]
- Ferron, P.-J.; Hogeveen, K.; Fessard, V.; Hégarat, L. Comparative analysis of the cytotoxic effects of okadaic acid-group toxins on human intestinal cell lines. Mar. Drugs 2014, 12, 4616–4634. [Google Scholar] [CrossRef] [PubMed]
- Leira, F.; Alvarez, C.; Cabado, A.G.; Vieites, J.M.; Vieytes, M.R.; Botana, L.M. Development of a F actin-based live-cell fluorimetric microplate assay for diarrhetic shellfish toxins. Anal. Biochem. 2003, 317, 129–135. [Google Scholar] [CrossRef]
- Ronzitti, G.; Hess, P.; Rehmann, N.; Rossini, G.P. Azaspiracid-1 alters the E-cadherin pool in epithelial cells. Toxicol. Sci. Off. J. Soc. Toxicol. 2007, 95, 427–435. [Google Scholar] [CrossRef] [PubMed]
- Beaulieu, J.-F.; Ménard, D. Isolation, characterization, and culture of normal human intestinal crypt and villus cells. Methods Mol. Biol. Clifton NJ 2012, 806, 157–173. [Google Scholar]
- Perreault, N.; Beaulieu, J.F. Use of the dissociating enzyme thermolysin to generate viable human normal intestinal epithelial cell cultures. Exp. Cell Res. 1996, 224, 354–364. [Google Scholar] [CrossRef] [PubMed]
- Guezguez, A.; Paré, F.; Benoit, Y.D.; Basora, N.; Beaulieu, J.-F. Modulation of stemness in a human normal intestinal epithelial crypt cell line by activation of the WNT signaling pathway. Exp. Cell Res. 2014, 355–364. [Google Scholar] [CrossRef] [PubMed]
- Pageot, L.-P.; Perreault, N.; Basora, N.; Francoeur, C.; Magny, P.; Beaulieu, J.-F. Human cell models to study small intestinal functions: Recapitulation of the crypt-villus axis. Microsc. Res. Tech. 2000, 49, 394–406. [Google Scholar] [CrossRef]
- Chou, T.C.; Talalay, P. Quantitative analysis of dose-effect relationships: the combined effects of multiple drugs or enzyme inhibitors. Adv. Enzyme Regul. 1984, 22, 27–55. [Google Scholar] [CrossRef]
- Wang, D.-Z. Neurotoxins from marine dinoflagellates: A brief review. Mar. Drugs 2008, 6, 349–371. [Google Scholar] [CrossRef] [PubMed]
- Takenaka, T.; Harada, N.; Kuze, J.; Chiba, M.; Iwao, T.; Matsunaga, T. Human small intestinal epithelial cells differentiated from adult intestinal stem cells as a novel system for predicting oral drug absorption in humans. Drug Metab. Dispos. Biol. Fate Chem. 2014, 42, 1947–1954. [Google Scholar] [CrossRef] [PubMed]
- Sérandour, A.-L.; Ledreux, A.; Morin, B.; Derick, S.; Augier, E.; Lanceleur, R.; Hamlaoui, S.; Moukha, S.; Furger, C.; Biré, R.; et al. Collaborative study for the detection of toxic compounds in shellfish extracts using cell-based assays. Part I: Screening strategy and pre-validation study with lipophilic marine toxins. Anal. Bioanal. Chem. 2012, 403, 1983–1993. [Google Scholar] [CrossRef] [PubMed]
- Azaspiracids—Toxicological Evaluation, Test Methods and Identification of the Source Organism (ASTOX II). Available online: http://oar.marine.ie/handle/10793/970 (accessed on 19 February 2016).
- Le Hégarat, L.; Jacquin, A.-G.; Bazin, E.; Fessard, V. Genotoxicity of the marine toxin okadaic acid, in human Caco-2 cells and in mice gut cells. Environ. Toxicol. 2006, 21, 55–64. [Google Scholar] [CrossRef] [PubMed]
- Aune, T.; Sørby, R.; Yasumoto, T.; Ramstad, H.; Landsverk, T. Comparison of oral and intraperitoneal toxicity of yessotoxin towards mice. Toxicon 2002, 40, 77–82. [Google Scholar] [CrossRef]
- Gauthier, R.; Harnois, C.; Drolet, J.F.; Reed, J.C.; Vézina, A.; Vachon, P.H. Human intestinal epithelial cell survival: Differentiation state-specific control mechanisms. Am. J. Physiol. Cell Physiol. 2001, 280, C1540–C1554. [Google Scholar] [PubMed]
- Korsnes, M.S.; Espenes, A.; Hetland, D.L.; Hermansen, L.C. Paraptosis-like cell death induced by yessotoxin. Toxicol. In Vitro 2011, 25, 1764–1770. [Google Scholar] [CrossRef] [PubMed]
- Chou, T.-C. Theoretical basis, experimental design, and computerized simulation of synergism and antagonism in drug combination studies. Pharmacol. Rev. 2006, 58, 621–681. [Google Scholar] [CrossRef] [PubMed]
- Rodríguez, L.; González, V.; Martínez, A.; Paz, B.; Lago, J.; Cordeiro, V.; Blanco, L.; Vieites, J.; Cabado, A. Occurrence of lipophilic marine toxins in shellfish from galicia (NW of Spain) and Synergies among them. Mar. Drugs 2015, 13, 1666–1687. [Google Scholar] [CrossRef] [PubMed]
- Chou, T.-C. Frequently asked questions in drug combinations and the mass-action law-based answers. Synergy 2014, 1, 3–21. [Google Scholar] [CrossRef]
- Boobis, A.; Budinsky, R.; Collie, S.; Crofton, K.; Embry, M.; Felter, S.; Hertzberg, R.; Kopp, D.; Mihlan, G.; Mumtaz, M.; et al. Critical analysis of literature on low-dose synergy for use in screening chemical mixtures for risk assessment. Crit. Rev. Toxicol. 2011, 41, 369–383. [Google Scholar] [CrossRef] [PubMed]
- Alassane-Kpembi, I.; Kolf-Clauw, M.; Gauthier, T.; Abrami, R.; Abiola, F.A.; Oswald, I.P.; Puel, O. New insights into mycotoxin mixtures: The toxicity of low doses of Type B trichothecenes on intestinal epithelial cells is synergistic. Toxicol. Appl. Pharmacol. 2013, 272, 191–198. [Google Scholar] [CrossRef] [PubMed]

{kind=link}
{kind=link}
{kind=link}
| Cytotoxicity | Caco-2 | HIEC | ||||
|---|---|---|---|---|---|---|
| IC50 (nM) | 95% CI | n | IC50 (nM) | 95% CI | n | |
| OA | 78.52 | 46.48–111.60 | 6 | 65.29 | 49.14–81.45 | 6 |
| AZA-1 | 4.03 * | 2.05–6.01 | 6 | 12.52 * | 8.64–16.40 | 6 |
| YTX | 4.08 | 1.18–7.83 | 6 | 3.36 | 1.17–5.93 | 6 |
| Toxins Mixture | Molar Combination Ratio | |||
|---|---|---|---|---|
| Mix 1 | Mix 2 | Mix 3 | Mix 4 | |
| AZA:YTX | 1:0.8 | 1:1.3 | 1:2.4 | 1:3.6 |
| AZA:OA | 1:51 | 1:27.2 | 1:15.3 | 1:8.2 |
| YTX:OA | 1:26.5 | 1:14.1 | 1:7.9 | 1:4.2 |
| Phycotoxins | Ratio | Caco-2 | HIEC | ||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| IC25 | IC50 | IC75 | IC25 | IC50 | IC75 | ||||||||
| CI | DRI | CI | DRI | CI | DRI | CI | DRI | CI | DRI | CI | DRI | ||
| AZA:YTX | |||||||||||||
| Mix 1 | 1:0.8 | 0.84 | / | 0.85 | / | 0.88 | / | 0.51 | 3.1:2.8 | 0.42 | 3.4:4.3 | 0.36 | 3.8:6.8 |
| Mix 2 | 1:1.3 | 0.74 | 3.9:2.1 | 0.78 | 2.0:4.0 | 0.83 | / | 0.59 | 2.3:3.9 | 0.57 | 2.3:5.2 | 0.56 | 2.3:7.2 |
| Mix 3 | 1:2.4 | 0.77 | 1.5:5.0 | 0.77 | 1.4:4.9 | 0.78 | 1.3:4.8 | 0.66 | 1.7:5.2 | 0.63 | 1.7:7.5 | 0.62 | 1.8:10.9 |
| Mix 4 | 1:3.6 | 0.7 | 1.4:8.6 | 0.66 | 1.5:9.4 | 0.61 | 1.4:10.4 | 1.07 | / | 1.04 | / | 1.03 | / |
| AZA:OA | |||||||||||||
| Mix 1 | 1:8.2 | 1.20 | / | 1.29 | / | 1.40 | / | 1.35 | / | 1.60 | / | 1.91 | / |
| Mix 2 | 1:15.3 | 1.64 | / | 1.82 | / | 2.11 | / | 1.04 | / | 1.00 | / | 1.02 | / |
| Mix 3 | 1:27.2 | 1.70 | / | 1.76 | / | 1.86 | / | 0.95 | / | 0.97 | / | 1.00 | / |
| Mix 4 | 1:51 | 1.66 | / | 1.85 | / | 2.13 | / | 1.17 | / | 1.22 | / | 1.28 | / |
| YTX:OA | |||||||||||||
| Mix 1 | 1:4.2 | 1.2 | / | 1.38 | / | 1.63 | / | 0.72 | 3.4:1.8 | 0.68 | 3.5:1.4 | 0.68 | 3.4:1.2 |
| Mix 2 | 1:7.9 | 1.02 | / | 1.00 | / | 0.98 | / | 1.35 | / | 1.49 | / | 1.66 | / |
| Mix 3 | 1:14.1 | 0.95 | / | 0.91 | 2.1:2.0 | 0.87 | / | 1.10 | / | 1.04 | / | 1.02 | / |
| Mix 4 | 1:26.5 | 1.14 | / | 1.14 | / | 1.16 | / | 1.01 | / | 1.01 | / | 1.01 | / |
© 2016 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons by Attribution (CC-BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ferron, P.-J.; Dumazeau, K.; Beaulieu, J.-F.; Le Hégarat, L.; Fessard, V. Combined Effects of Lipophilic Phycotoxins (Okadaic Acid, Azapsiracid-1 and Yessotoxin) on Human Intestinal Cells Models. Toxins 2016, 8, 50. https://doi.org/10.3390/toxins8020050
Ferron P-J, Dumazeau K, Beaulieu J-F, Le Hégarat L, Fessard V. Combined Effects of Lipophilic Phycotoxins (Okadaic Acid, Azapsiracid-1 and Yessotoxin) on Human Intestinal Cells Models. Toxins. 2016; 8(2):50. https://doi.org/10.3390/toxins8020050
Chicago/Turabian StyleFerron, Pierre-Jean, Kevin Dumazeau, Jean-François Beaulieu, Ludovic Le Hégarat, and Valérie Fessard. 2016. "Combined Effects of Lipophilic Phycotoxins (Okadaic Acid, Azapsiracid-1 and Yessotoxin) on Human Intestinal Cells Models" Toxins 8, no. 2: 50. https://doi.org/10.3390/toxins8020050
APA StyleFerron, P.-J., Dumazeau, K., Beaulieu, J.-F., Le Hégarat, L., & Fessard, V. (2016). Combined Effects of Lipophilic Phycotoxins (Okadaic Acid, Azapsiracid-1 and Yessotoxin) on Human Intestinal Cells Models. Toxins, 8(2), 50. https://doi.org/10.3390/toxins8020050