
Bee Venom Phospholipase A2: Yesterday’s Enemy Becomes Today’s Friend


Abstract
:1. Introduction
2. Bee Venom Group III sPLA2 as an Enzyme
3. Bee Venom Group III sPLA2 as a Ligand
4. Bee Venom Group III sPLA2: Yesterday’s Enemy
4.1. T Cell Responses and Anaphylaxis Induced by Bee Venom Group III sPLA2
4.2. Nociceptive Effects and Neurotoxicity of Bee Venom Group III sPLA2
5. Bee Venom Group III sPLA2: Today’s Friend
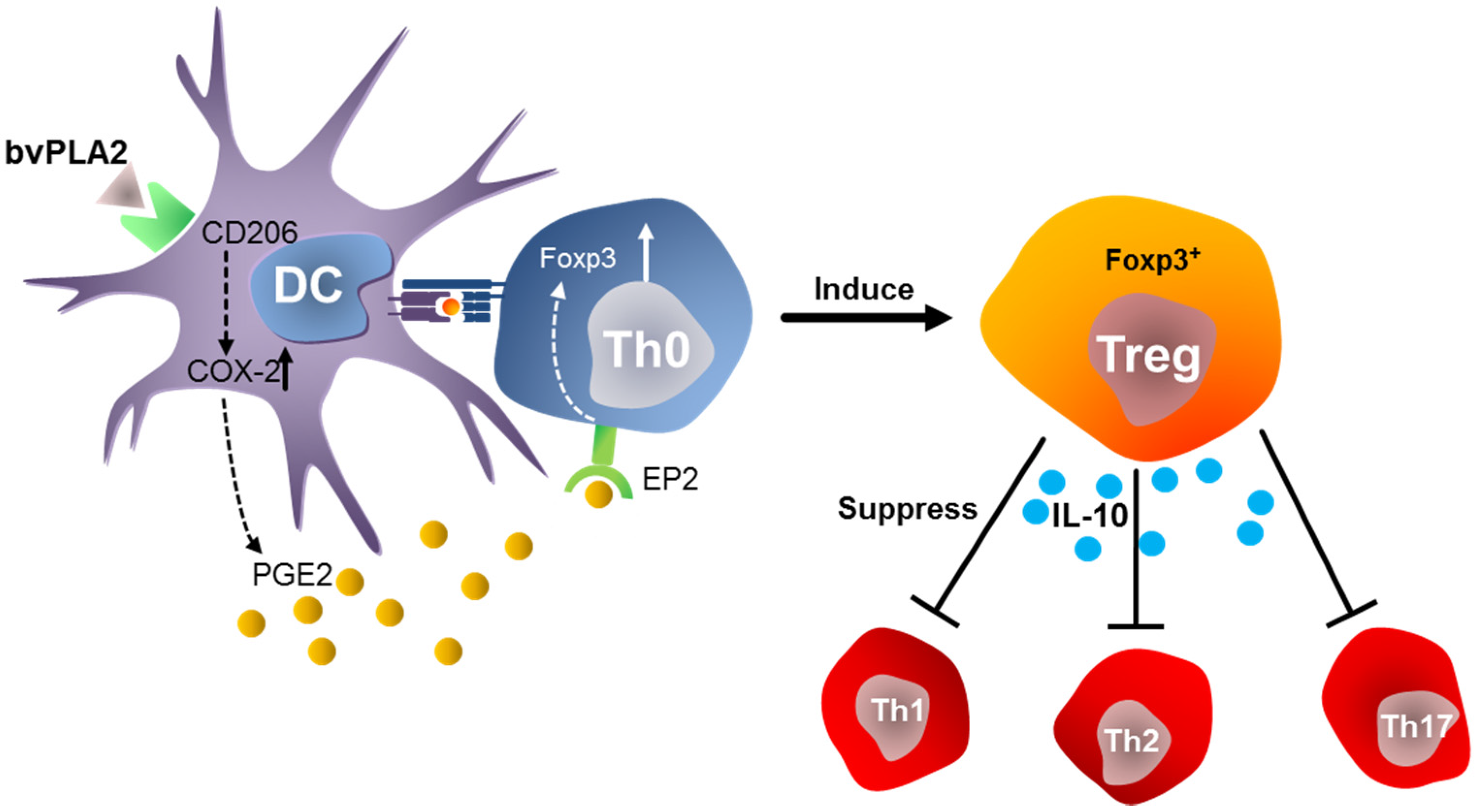
5.1. Anti-Inflammatory Effects of Bee Venom Group III sPLA2
5.2. Anti-Neuronal Injury and Anti-Nociceptive Effects of Bee Venom Group III sPLA2
5.3. Anti-Tumor Effects of Bee Venom Group III sPLA2
5.4. Vaccination Approaches
5.5. Anti-Parasite and Anti-Bacterial Effects of Bee Venom Group III sPLA2
6. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Senel, E.; Kuyucu, M.; Suslu, I. Honey and bee venom in dermatology: A novel possible alternative or complimentary therapy for psoriasis vulgaris. Anc. Sci. Life 2014, 33, 192–193. [Google Scholar] [CrossRef] [PubMed]
- Lee, S.M.; Lim, J.; Lee, J.D.; Choi, D.Y.; Lee, S. Bee venom treatment for refractory postherpetic neuralgia: A case report. J. Altern. Complement. Med. 2014, 20, 212–214. [Google Scholar] [CrossRef] [PubMed]
- Chen, J.; Lariviere, W.R. The nociceptive and anti-nociceptive effects of bee venom injection and therapy: A double-edged sword. Prog. Neurobiol. 2010, 92, 151–183. [Google Scholar] [CrossRef] [PubMed]
- Liu, X.D.; Zhang, J.L.; Zheng, H.G.; Liu, F.Y.; Chen, Y. Clinical randomized study of bee-sting therapy for rheumatoid arthritis. Zhen Ci Yan Jiu 2008, 33, 197–200. [Google Scholar] [PubMed]
- Lee, J.D.; Park, H.J.; Chae, Y.; Lim, S. An overview of bee venom acupuncture in the treatment of arthritis. Evid. Based Complement. Alternat. Med. 2005, 2, 79–84. [Google Scholar] [CrossRef] [PubMed]
- Castro, H.J.; Mendez-Lnocencio, J.I.; Omidvar, B.; Omidvar, J.; Santilli, J.; Nielsen, H.S., Jr.; Pavot, A.P.; Richert, J.R.; Bellanti, J.A. A phase I study of the safety of honeybee venom extract as a possible treatment for patients with progressive forms of multiple sclerosis. Allergy Asthma Proc. 2005, 26, 470–476. [Google Scholar] [PubMed]
- Lee, J.Y.; Kang, S.S.; Kim, J.H.; Bae, C.S.; Choi, S.H. Inhibitory effect of whole bee venom in adjuvant-induced arthritis. In Vivo 2005, 19, 801–805. [Google Scholar] [PubMed]
- Gajski, G.; Garaj-Vrhovac, V. Radioprotective effects of honeybee venom (Apis mellifera) against 915-MHZ microwave radiation-induced DNA damage in wistar rat lymphocytes: In vitro study. Int. J. Toxicol. 2009, 28, 88–98. [Google Scholar] [CrossRef] [PubMed]
- Varanda, E.A.; Monti, R.; Tavares, D.C. Inhibitory effect of propolis and bee venom on the mutagenicity of some direct- and indirect-acting mutagens. Teratog. Carcinog. Mutagen. 1999, 19, 403–413. [Google Scholar] [CrossRef]
- Park, Y.C.; Koh, P.S.; Seo, B.K.; Lee, J.W.; Cho, N.S.; Park, H.S.; Park, D.S.; Baek, Y.H. Long-term effectiveness of bee venom acupuncture and physiotherapy in the treatment of adhesive capsulitis: A one-year follow-up analysis of a previous randomized controlled trial. J. Altern. Complement. Med. 2014, 20, 919–924. [Google Scholar] [CrossRef] [PubMed]
- Kim, H.; Lee, G.; Park, S.; Chung, H.S.; Lee, H.; Kim, J.Y.; Nam, S.; Kim, S.K.; Bae, H. Bee venom mitigates cisplatin-induced nephrotoxicity by regulating CD4+CD25+Foxp3+ regulatory T cells in mice. Evid. Based Complement. Alternat. Med. 2013, 2013. [Google Scholar] [CrossRef] [PubMed]
- Choi, M.S.; Park, S.; Choi, T.; Lee, G.; Haam, K.K.; Hong, M.C.; Min, B.I.; Bae, H. Bee venom ameliorates ovalbumin induced allergic asthma via modulating CD4+CD25+ regulatory t cells in mice. Cytokine 2012, 61, 256–265. [Google Scholar] [CrossRef] [PubMed]
- Lee, K.G.; Cho, H.J.; Bae, Y.S.; Park, K.K.; Choe, J.Y.; Chung, I.K.; Kim, M.; Yeo, J.H.; Park, K.H.; Lee, Y.S.; et al. Bee venom suppresses LPS-mediated NO/iNOS induction through inhibition of PKC-α expression. J. Ethnopharmacol. 2009, 123, 15–21. [Google Scholar] [CrossRef] [PubMed]
- Park, H.J.; Lee, S.H.; Son, D.J.; Oh, K.W.; Kim, K.H.; Song, H.S.; Kim, G.J.; Oh, G.T.; Yoon, D.Y.; Hong, J.T. Antiarthritic effect of bee venom: Inhibition of inflammation mediator generation by suppression of NF-kappab through interaction with the p50 subunit. Arthritis Rheum. 2004, 50, 3504–3515. [Google Scholar] [CrossRef] [PubMed]
- Yoon, H.; Kim, M.J.; Yoon, I.; Li, D.X.; Bae, H.; Kim, S.K. Nicotinic acetylcholine receptors mediate the suppressive effect of an injection of diluted bee venom into the GV3 acupoint on oxaliplatin-induced neuropathic cold allodynia in rats. Biol. Pharm. Bull. 2015, 38, 710–714. [Google Scholar] [CrossRef] [PubMed]
- Lim, B.S.; Moon, H.J.; Li, D.X.; Gil, M.; Min, J.K.; Lee, G.; Bae, H.; Kim, S.K.; Min, B.I. Effect of bee venom acupuncture on oxaliplatin-induced cold allodynia in rats. Evid. Based Complement. Alternat. Med. 2013, 2013. [Google Scholar] [CrossRef] [PubMed]
- Baek, Y.H.; Huh, J.E.; Lee, J.D.; Choi, D.Y.; Park, D.S. Antinociceptive effect and the mechanism of bee venom acupuncture (apipuncture) on inflammatory pain in the rat model of collagen-induced arthritis: Mediation by α2-adrenoceptors. Brain Res. 2006, 1073–1074, 305–310. [Google Scholar] [CrossRef] [PubMed]
- Huh, J.E.; Baek, Y.H.; Lee, M.H.; Choi, D.Y.; Park, D.S.; Lee, J.D. Bee venom inhibits tumor angiogenesis and metastasis by inhibiting tyrosine phosphorylation of VEGFR-2 in LLC-tumor-bearing mice. Cancer Lett. 2010, 292, 98–110. [Google Scholar] [CrossRef] [PubMed]
- Moon, D.O.; Park, S.Y.; Heo, M.S.; Kim, K.C.; Park, C.; Ko, W.S.; Choi, Y.H.; Kim, G.Y. Key regulators in bee venom-induced apoptosis are Bcl-2 and caspase-3 in human leukemic U937 cells through downregulation of ERK and Akt. Int. Immunopharmacol. 2006, 6, 1796–1807. [Google Scholar] [CrossRef] [PubMed]
- Hu, H.; Chen, D.; Li, Y.; Zhang, X. Effect of polypeptides in bee venom on growth inhibition and apoptosis induction of the human hepatoma cell line SMMC-7721 in vitro and Balb/c nude mice in vivo. J. Pharm. Pharmacol. 2006, 58, 83–89. [Google Scholar] [CrossRef] [PubMed]
- Orsolic, N.; Sver, L.; Verstovsek, S.; Terzic, S.; Basic, I. Inhibition of mammary carcinoma cell proliferation in vitro and tumor growth in vivo by bee venom. Toxicon 2003, 41, 861–870. [Google Scholar] [CrossRef]
- Orsolic, N.; Knezevic, A.; Sver, L.; Terzic, S.; Hackenberger, B.K.; Basic, I. Influence of honey bee products on transplantable murine tumours. Vet. Comp. Oncol. 2003, 1, 216–226. [Google Scholar] [CrossRef] [PubMed]
- Jang, M.H.; Shin, M.C.; Lim, S.; Han, S.M.; Park, H.J.; Shin, I.; Lee, J.S.; Kim, K.A.; Kim, E.H.; Kim, C.J. Bee venom induces apoptosis and inhibits expression of cyclooxygenase-2 mRNA in human lung cancer cell line NCI-H1299. J. Pharmacol. Sci. 2003, 91, 95–104. [Google Scholar] [CrossRef] [PubMed]
- Liu, X.; Chen, D.; Xie, L.; Zhang, R. Effect of honey bee venom on proliferation of K1735M2 mouse melanoma cells in vitro and growth of murine B16 melanomas in vivo. J. Pharm. Pharmacol. 2002, 54, 1083–1089. [Google Scholar] [CrossRef] [PubMed]
- Habermann, E. Bee and wasp venoms. Science 1972, 177, 314–322. [Google Scholar] [CrossRef] [PubMed]
- Lariviere, W.R.; Melzack, R. The bee venom test: A new tonic-pain test. Pain 1996, 66, 271–277. [Google Scholar] [CrossRef]
- Son, D.J.; Lee, J.W.; Lee, Y.H.; Song, H.S.; Lee, C.K.; Hong, J.T. Therapeutic application of anti-arthritis, pain-releasing, and anti-cancer effects of bee venom and its constituent compounds. Pharmacol. Ther. 2007, 115, 246–270. [Google Scholar] [CrossRef] [PubMed]
- Sobotka, A.K.; Franklin, R.M.; Adkinson, N.F., Jr.; Valentine, M.; Baer, H.; Lichtenstein, L.M. Allergy to insect stings. II. Phospholipase A: The major allergen in honeybee venom. J. Allergy Clin. Immunol. 1976, 57, 29–40. [Google Scholar] [CrossRef]
- Von Allmen, C.E.; Schmitz, N.; Bauer, M.; Hinton, H.J.; Kurrer, M.O.; Buser, R.B.; Gwerder, M.; Muntwiler, S.; Sparwasser, T.; Beerli, R.R.; et al. Secretory phospholipase A2-IID is an effector molecule of CD4+CD25+ regulatory T cells. Proc. Natl. Acad. Sci. USA 2009, 106, 11673–11678. [Google Scholar] [CrossRef] [PubMed]
- Boilard, E.; Lai, Y.; Larabee, K.; Balestrieri, B.; Ghomashchi, F.; Fujioka, D.; Gobezie, R.; Coblyn, J.S.; Weinblatt, M.E.; Massarotti, E.M.; et al. A novel anti-inflammatory role for secretory phospholipase A2 in immune complex-mediated arthritis. EMBO Mol. Med. 2010, 2, 172–187. [Google Scholar] [CrossRef] [PubMed]
- Murakami, M.; Sato, H.; Miki, Y.; Yamamoto, K.; Taketomi, Y. A new era of secreted phospholipase A2. J. Lipid Res. 2015, 56, 1248–1261. [Google Scholar] [CrossRef] [PubMed]
- Murakami, M.; Taketomi, Y.; Miki, Y.; Sato, H.; Yamamoto, K.; Lambeau, G. Emerging roles of secreted phospholipase A2 enzymes: The 3rd edition. Biochimie 2014, 107, 105–113. [Google Scholar] [CrossRef] [PubMed]
- Sitkiewicz, I.; Stockbauer, K.E.; Musser, J.M. Secreted bacterial phospholipase A2 enzymes: Better living through phospholipolysis. Trends Microbiol. 2007, 15, 63–69. [Google Scholar] [CrossRef] [PubMed]
- Miozzi, L.; Balestrini, R.; Bolchi, A.; Novero, M.; Ottonello, S.; Bonfante, P. Phospholipase A2 up-regulation during mycorrhiza formation in tuber borchii. New Phytol. 2005, 167, 229–238. [Google Scholar] [CrossRef] [PubMed]
- Soragni, E.; Bolchi, A.; Balestrini, R.; Gambaretto, C.; Percudani, R.; Bonfante, P.; Ottonello, S. A nutrient-regulated, dual localization phospholipase A2 in the symbiotic fungus tuber borchii. EMBO J. 2001, 20, 5079–5090. [Google Scholar] [CrossRef] [PubMed]
- Lee, H.Y.; Bahn, S.C.; Shin, J.S.; Hwang, I.; Back, K.; Doelling, J.H.; Ryu, S.B. Multiple forms of secretory phospholipase A2 in plants. Prog. Lipid Res. 2005, 44, 52–67. [Google Scholar] [CrossRef] [PubMed]
- Park, Y.; Kumar, S.; Kanumuri, R.; Stanley, D.; Kim, Y. A novel calcium-independent cellular PLA2 acts in insect immunity and larval growth. Insect Biochem. Mol. Biol. 2015, 66, 13–23. [Google Scholar] [CrossRef] [PubMed]
- Kuchler, K.; Gmachl, M.; Sippl, M.J.; Kreil, G. Analysis of the cDNA for phospholipase A2 from honeybee venom glands. The deduced amino acid sequence reveals homology to the corresponding vertebrate enzymes. Eur. J. Biochem. 1989, 184, 249–254. [Google Scholar] [CrossRef] [PubMed]
- Ryu, Y.; Oh, Y.; Yoon, J.; Cho, W.; Baek, K. Molecular characterization of a gene encoding the drosophila melanogaster phospholipase A2. Biochim. Biophys. Acta 2003, 1628, 206–210. [Google Scholar] [CrossRef]
- Kini, R.M. Excitement ahead: Structure, function and mechanism of snake venom phospholipase A2 enzymes. Toxicon 2003, 42, 827–840. [Google Scholar] [CrossRef] [PubMed]
- Dennis, E.A.; Cao, J.; Hsu, Y.H.; Magrioti, V.; Kokotos, G. Phospholipase A2 enzymes: Physical structure, biological function, disease implication, chemical inhibition, and therapeutic intervention. Chem. Rev. 2011, 111, 6130–6185. [Google Scholar] [CrossRef] [PubMed]
- Murakami, M.; Taketomi, Y.; Miki, Y.; Sato, H.; Hirabayashi, T.; Yamamoto, K. Recent progress in phospholipase A2 research: From cells to animals to humans. Prog. Lipid Res. 2011, 50, 152–192. [Google Scholar] [CrossRef] [PubMed]
- Valentin, E.; Ghomashchi, F.; Gelb, M.H.; Lazdunski, M.; Lambeau, G. Novel human secreted phospholipase A2 with homology to the group III bee venom enzyme. J. Biol. Chem. 2000, 275, 7492–7496. [Google Scholar] [CrossRef] [PubMed]
- Sato, H.; Kato, R.; Isogai, Y.; Saka, G.; Ohtsuki, M.; Taketomi, Y.; Yamamoto, K.; Tsutsumi, K.; Yamada, J.; Masuda, S.; et al. Analyses of group III secreted phospholipase A2 transgenic mice reveal potential participation of this enzyme in plasma lipoprotein modification, macrophage foam cell formation, and atherosclerosis. J. Biol. Chem. 2008, 283, 33483–33497. [Google Scholar] [CrossRef] [PubMed]
- Sato, H.; Taketomi, Y.; Isogai, Y.; Miki, Y.; Yamamoto, K.; Masuda, S.; Hosono, T.; Arata, S.; Ishikawa, Y.; Ishii, T.; et al. Group III secreted phospholipase A2 regulates epididymal sperm maturation and fertility in mice. J. Clin. Investig. 2010, 120, 1400–1414. [Google Scholar] [CrossRef] [PubMed]
- Masuda, S.; Yamamoto, K.; Hirabayashi, T.; Ishikawa, Y.; Ishii, T.; Kudo, I.; Murakami, M. Human group III secreted phospholipase A2 promotes neuronal outgrowth and survival. Biochem. J. 2008, 409, 429–438. [Google Scholar] [CrossRef] [PubMed]
- Yang, X.; Sheng, W.; He, Y.; Cui, J.; Haidekker, M.A.; Sun, G.Y.; Lee, J.C. Secretory phospholipase A2 type III enhances alpha-secretase-dependent amyloid precursor protein processing through alterations in membrane fluidity. J. Lipid Res. 2009, 51, 957–966. [Google Scholar] [CrossRef] [PubMed]
- Murakami, M.; Masuda, S.; Shimbara, S.; Ishikawa, Y.; Ishii, T.; Kudo, I. Cellular distribution, post-translational modification, and tumorigenic potential of human group III secreted phospholipase A2. J. Biol. Chem. 2005, 280, 24987–24998. [Google Scholar] [CrossRef] [PubMed]
- Gijs, H.L.; Willemarck, N.; Vanderhoydonc, F.; Khan, N.A.; Dehairs, J.; Derua, R.; Waelkens, E.; Taketomi, Y.; Murakami, M.; Agostinis, P.; et al. Primary cilium suppression by SREBP1C involves distortion of vesicular trafficking by PLA2G3. Mol. Biol. Cell 2015, 26, 2321–2332. [Google Scholar] [CrossRef] [PubMed]
- Arita, H.; Hanasaki, K.; Nakano, T.; Oka, S.; Teraoka, H.; Matsumoto, K. Novel proliferative effect of phospholipase A2 in swiss 3T3 cells via specific binding site. J. Biol. Chem. 1991, 266, 19139–19141. [Google Scholar] [PubMed]
- Kanemasa, T.; Arimura, A.; Kishino, J.; Ohtani, M.; Arita, H. Contraction of guinea pig lung parenchyma by pancreatic type phospholipase A2 via its specific binding site. FEBS Lett. 1992, 303, 217–220. [Google Scholar] [PubMed]
- Lambeau, G.; Schmid-Alliana, A.; Lazdunski, M.; Barhanin, J. Identification and purification of a very high affinity binding protein for toxic phospholipases A2 in skeletal muscle. J. Biol. Chem. 1990, 265, 9526–9532. [Google Scholar] [PubMed]
- Lambeau, G.; Barhanin, J.; Schweitz, H.; Qar, J.; Lazdunski, M. Identification and properties of very high affinity brain membrane-binding sites for a neurotoxic phospholipase from the taipan venom. J. Biol. Chem. 1989, 264, 11503–11510. [Google Scholar] [PubMed]
- Hanasaki, K.; Yokota, Y.; Ishizaki, J.; Itoh, T.; Arita, H. Resistance to endotoxic shock in phospholipase A2 receptor-deficient mice. J. Biol. Chem. 1997, 272, 32792–32797. [Google Scholar] [CrossRef] [PubMed]
- Lambeau, G.; Lazdunski, M. Receptors for a growing family of secreted phospholipases A2. Trends Pharmacol. Sci. 1999, 20, 162–170. [Google Scholar] [CrossRef]
- Nicolas, J.P.; Lin, Y.; Lambeau, G.; Ghomashchi, F.; Lazdunski, M.; Gelb, M.H. Localization of structural elements of bee venom phospholipase A2 involved in n-type receptor binding and neurotoxicity. J. Biol. Chem. 1997, 272, 7173–7181. [Google Scholar] [CrossRef] [PubMed]
- Palm, N.W.; Rosenstein, R.K.; Yu, S.; Schenten, D.D.; Florsheim, E.; Medzhitov, R. Bee venom phospholipase A2 induces a primary type 2 response that is dependent on the receptor ST2 and confers protective immunity. Immunity 2013, 39, 976–985. [Google Scholar] [CrossRef] [PubMed]
- Mukhopadhyay, A.; Stahl, P. Bee venom phospholipase A2 is recognized by the macrophage mannose receptor. Arch. Biochem. Biophys. 1995, 324, 78–84. [Google Scholar] [CrossRef] [PubMed]
- Kim, H.; Lee, H.; Lee, G.; Jang, H.; Kim, S.S.; Yoon, H.; Kang, G.H.; Hwang, D.S.; Kim, S.K.; Chung, H.S.; et al. Phospholipase A2 inhibits cisplatin-induced acute kidney injury by modulating regulatory T cells by the CD206 mannose receptor. Kidney Int. 2015, 88, 550–559. [Google Scholar] [CrossRef] [PubMed]
- Chung, E.S.; Lee, G.; Lee, C.; Ye, M.; Chung, H.S.; Kim, H.; Bae, S.S.; Hwang, D.S.; Bae, H. Bee venom phospholipase A2, a novel Foxp3+ regulatory t cell inducer, protects dopaminergic neurons by modulating neuroinflammatory responses in a mouse model of parkinson’s disease. J. Immunol. 2015, 195, 4853–4860. [Google Scholar] [CrossRef] [PubMed]
- Bircher, A.J. Systemic immediate allergic reactions to arthropod stings and bites. Dermatology 2005, 210, 119–127. [Google Scholar] [CrossRef] [PubMed]
- Madero, M.F.; Gamez, C.; Madero, M.A.; Fernandez-Nieto, M.; Sastre, J.; del Pozo, V. Characterization of allergens in four south american snake species. Int. Arch. Allergy Immunol. 2009, 150, 307–310. [Google Scholar] [CrossRef] [PubMed]
- Muller, U.R. Insect venoms. Chem. Immunol. Allergy 2010, 95, 141–156. [Google Scholar] [PubMed]
- Taketomi, Y.; Ueno, N.; Kojima, T.; Sato, H.; Murase, R.; Yamamoto, K.; Tanaka, S.; Sakanaka, M.; Nakamura, M.; Nishito, Y.; et al. Mast cell maturation is driven via a group III phospholipase A2-prostaglandin D2-DP1 receptor paracrine axis. Nat. Immunol. 2013, 14, 554–563. [Google Scholar] [CrossRef] [PubMed]
- Sato, H.; Taketomi, Y.; Isogai, Y.; Masuda, S.; Kobayashi, T.; Yamamoto, K.; Murakami, M. Group III secreted phospholipase A2 transgenic mice spontaneously develop inflammation. Biochem. J. 2009, 421, 17–27. [Google Scholar] [CrossRef] [PubMed]
- Dudler, T.; Machado, D.C.; Kolbe, L.; Annand, R.R.; Rhodes, N.; Gelb, M.H.; Koelsch, K.; Suter, M.; Helm, B.A. A link between catalytic activity, IGE-independent mast cell activation, and allergenicity of bee venom phospholipase A2. J. Immunol. 1995, 155, 2605–2613. [Google Scholar] [PubMed]
- Mustafa, F.B.; Ng, F.S.; Nguyen, T.H.; Lim, L.H. Honeybee venom secretory phospholipase A2 induces leukotriene production but not histamine release from human basophils. Clin. Exp. Immunol. 2008, 151, 94–100. [Google Scholar] [CrossRef] [PubMed]
- Bourgeois, E.A.; Subramaniam, S.; Cheng, T.Y.; De Jong, A.; Layre, E.; Ly, D.; Salimi, M.; Legaspi, A.; Modlin, R.L.; Salio, M.; et al. Bee venom processes human skin lipids for presentation by CD1A. J. Exp. Med. 2015, 212, 149–163. [Google Scholar] [CrossRef] [PubMed]
- Sun, G.Y.; Xu, J.; Jensen, M.D.; Simonyi, A. Phospholipase A2 in the central nervous system: Implications for neurodegenerative diseases. J. Lipid Res. 2004, 45, 205–213. [Google Scholar] [CrossRef] [PubMed]
- Yue, H.Y.; Fujita, T.; Kumamoto, E. Phospholipase A2 activation by melittin enhances spontaneous glutamatergic excitatory transmission in rat substantia gelatinosa neurons. Neuroscience 2005, 135, 485–495. [Google Scholar] [CrossRef] [PubMed]
- Landucci, E.C.; Toyama, M.; Marangoni, S.; Oliveira, B.; Cirino, G.; Antunes, E.; de Nucci, G. Effect of crotapotin and heparin on the rat paw oedema induced by different secretory phospholipases A2. Toxicon 2000, 38, 199–208. [Google Scholar] [CrossRef]
- Clapp, L.E.; Klette, K.L.; DeCoster, M.A.; Bernton, E.; Petras, J.M.; Dave, J.R.; Laskosky, M.S.; Smallridge, R.C.; Tortella, F.C. Phospholipase A2-induced neurotoxicity in vitro and in vivo in rats. Brain Res. 1995, 693, 101–111. [Google Scholar] [CrossRef]
- Liu, N.K.; Zhang, Y.P.; Titsworth, W.L.; Jiang, X.; Han, S.; Lu, P.H.; Shields, C.B.; Xu, X.M. A novel role of phospholipase A2 in mediating spinal cord secondary injury. Ann. Neurol. 2006, 59, 606–619. [Google Scholar] [CrossRef] [PubMed]
- Titsworth, W.L.; Onifer, S.M.; Liu, N.K.; Xu, X.M. Focal phospholipases A2 group III injections induce cervical white matter injury and functional deficits with delayed recovery concomitant with schwann cell remyelination. Exp. Neurol. 2007, 207, 150–162. [Google Scholar] [CrossRef] [PubMed]
- Spence, A.; Klementowicz, J.E.; Bluestone, J.A.; Tang, Q. Targeting treg signaling for the treatment of autoimmune diseases. Curr. Opin. Immunol. 2015, 37, 11–20. [Google Scholar] [CrossRef] [PubMed]
- Josefowicz, S.Z.; Lu, L.F.; Rudensky, A.Y. Regulatory T cells: Mechanisms of differentiation and function. Ann. Rev. Immunol. 2012, 30, 531–564. [Google Scholar] [CrossRef] [PubMed]
- Park, S.; Baek, H.; Jung, K.H.; Lee, G.; Lee, H.; Kang, G.H.; Lee, G.; Bae, H. Bee venom phospholipase A2 suppresses allergic airway inflammation in an ovalbumin-induced asthma model through the induction of regulatory T cells. Immun. Inflamm. Dis. 2015, 3, 386–397. [Google Scholar] [CrossRef] [PubMed]
- Kim, H.; Keum, D.J.; Kwak, J.; Chung, H.S.; Bae, H. Bee venom phospholipase A2 protects against acetaminophen-induced acute liver injury by modulating regulatory T cells and IL-10 in mice. PLoS ONE 2014, 9. [Google Scholar] [CrossRef] [PubMed]
- Caramalho, I.; Melo, A.; Pedro, E.; Barbosa, M.M.; Victorino, R.M.; Pereira Santos, M.C.; Sousa, A.E. Bee venom enhances the differentiation of human regulatory T cells. Allergy 2015, 70, 1340–1345. [Google Scholar] [CrossRef] [PubMed]
- Pereira-Santos, M.C.; Baptista, A.P.; Melo, A.; Alves, R.R.; Soares, R.S.; Pedro, E.; Pereira-Barbosa, M.; Victorino, R.M.; Sousa, A.E. Expansion of circulating Foxp3+D25 bright CD4+ T cells during specific venom immunotherapy. Clin. Exp. Allergy 2008, 38, 291–297. [Google Scholar] [CrossRef] [PubMed]
- Marichal, T.; Starkl, P.; Reber, L.L.; Kalesnikoff, J.; Oettgen, H.C.; Tsai, M.; Metz, M.; Galli, S.J. A beneficial role for immunoglobulin e in host defense against honeybee venom. Immunity 2013, 39, 963–975. [Google Scholar] [CrossRef] [PubMed]
- Edstrom, A.; Briggman, M.; Ekstrom, P.A. Phospholipase A2 activity is required for regeneration of sensory axons in cultured adult sciatic nerves. J. Neurosci. Res. 1996, 43, 183–189. [Google Scholar] [CrossRef]
- Jeong, J.K.; Moon, M.H.; Bae, B.C.; Lee, Y.J.; Seol, J.W.; Park, S.Y. Bee venom phospholipase A2 prevents prion peptide induced-cell death in neuronal cells. Int. J. Mol. Med. 2011, 28, 867–873. [Google Scholar] [PubMed]
- Li, D.; Lee, Y.; Kim, W.; Lee, K.; Bae, H.; Kim, S.K. Analgesic effects of bee venom derived phospholipase A2 in a mouse model of oxaliplatin-induced neuropathic pain. Toxins 2015, 7, 2422–2434. [Google Scholar] [CrossRef] [PubMed]
- Putz, T.; Ramoner, R.; Gander, H.; Rahm, A.; Bartsch, G.; Bernardo, K.; Ramsay, S.; Thurnher, M. Bee venom secretory phospholipase A2 and phosphatidylinositol-homologues cooperatively disrupt membrane integrity, abrogate signal transduction and inhibit proliferation of renal cancer cells. Cancer Immunol. Immunother. 2007, 56, 627–640. [Google Scholar] [CrossRef] [PubMed]
- Putz, T.; Ramoner, R.; Gander, H.; Rahm, A.; Bartsch, G.; Thurnher, M. Antitumor action and immune activation through cooperation of bee venom secretory phospholipase A2 and phosphatidylinositol-(3,4)-bisphosphate. Cancer Immunol. Immunother. 2006, 55, 1374–1383. [Google Scholar] [CrossRef] [PubMed]
- Andreae, S.; Buisson, S.; Triebel, F. MHC class II signal transduction in human dendritic cells induced by a natural ligand, the LAG-3 protein (CD223). Blood 2003, 102, 2130–2137. [Google Scholar] [CrossRef] [PubMed]
- Appel, S.; Rupf, A.; Weck, M.M.; Schoor, O.; Brummendorf, T.H.; Weinschenk, T.; Grunebach, F.; Brossart, P. Effects of imatinib on monocyte-derived dendritic cells are mediated by inhibition of nuclear factor-kappab and akt signaling pathways. Clin. Cancer Res. 2005, 11, 1928–1940. [Google Scholar] [CrossRef] [PubMed]
- Del Prete, A.; Vermi, W.; Dander, E.; Otero, K.; Barberis, L.; Luini, W.; Bernasconi, S.; Sironi, M.; Santoro, A.; Garlanda, C.; et al. Defective dendritic cell migration and activation of adaptive immunity in PI3Kgamma-deficient mice. EMBO J. 2004, 23, 3505–3515. [Google Scholar] [CrossRef] [PubMed]
- Payrastre, B.; Missy, K.; Giuriato, S.; Bodin, S.; Plantavid, M.; Gratacap, M. Phosphoinositides: Key players in cell signalling, in time and space. Cell. Signal. 2001, 13, 377–387. [Google Scholar] [CrossRef]
- Toker, A. Phosphoinositides and signal transduction. Cell. Mol. Life Sci. 2002, 59, 761–779. [Google Scholar] [CrossRef] [PubMed]
- Gille, H.; Downward, J. Multiple ras effector pathways contribute to G(1) cell cycle progression. J. Biol. Chem. 1999, 274, 22033–22040. [Google Scholar] [CrossRef] [PubMed]
- Kennedy, S.G.; Wagner, A.J.; Conzen, S.D.; Jordan, J.; Bellacosa, A.; Tsichlis, P.N.; Hay, N. The PI 3-Kinase/Akt signaling pathway delivers an anti-apoptotic signal. Genes Dev. 1997, 11, 701–713. [Google Scholar] [CrossRef] [PubMed]
- Xu, Y.; Fang, X.J.; Casey, G.; Mills, G.B. Lysophospholipids activate ovarian and breast cancer cells. Biochem. J. 1995, 309, 933–940. [Google Scholar] [CrossRef] [PubMed]
- Sutphen, R.; Xu, Y.; Wilbanks, G.D.; Fiorica, J.; Grendys, E.C., Jr.; LaPolla, J.P.; Arango, H.; Hoffman, M.S.; Martino, M.; Wakeley, K.; et al. Lysophospholipids are potential biomarkers of ovarian cancer. Cancer Epidemiol. Biomarkers Prev. 2004, 13, 1185–1191. [Google Scholar] [PubMed]
- Babon, A.; Almunia, C.; Boccaccio, C.; Beaumelle, B.; Gelb, M.H.; Menez, A.; Maillere, B.; Abastado, J.P.; Salcedo, M.; Gillet, D. Cross-presentation of a CMV pp65 epitope by human dendritic cells using bee venom PLA2 as a membrane-binding vector. FEBS Lett. 2005, 579, 1658–1664. [Google Scholar] [CrossRef] [PubMed]
- Almunia, C.; Bretaudeau, M.; Held, G.; Babon, A.; Marchetti, C.; Castelli, F.A.; Menez, A.; Maillere, B.; Gillet, D. Bee venom phospholipase A2, a good “chauffeur” for delivering tumor antigen to the MHC I and MHC II peptide-loading compartments of the dendritic cells: The case of NY-ESO-1. PLoS ONE 2013, 8. [Google Scholar] [CrossRef] [PubMed]
- Boutrin, M.C.; Foster, H.A.; Pentreath, V.W. The effects of bee (apis mellifera) venom phospholipase A2 on trypanosoma brucei brucei and enterobacteria. Exp. Parasitol. 2008, 119, 246–251. [Google Scholar] [CrossRef] [PubMed]
- Zieler, H.; Keister, D.B.; Dvorak, J.A.; Ribeiro, J.M. A snake venom phospholipase A2 blocks malaria parasite development in the mosquito midgut by inhibiting ookinete association with the midgut surface. J. Exp. Biol. 2001, 204, 4157–4167. [Google Scholar] [PubMed]
- Moreira, L.A.; Ito, J.; Ghosh, A.; Devenport, M.; Zieler, H.; Abraham, E.G.; Crisanti, A.; Nolan, T.; Catteruccia, F.; Jacobs-Lorena, M. Bee venom phospholipase inhibits malaria parasite development in transgenic mosquitoes. J. Biol. Chem. 2002, 277, 40839–40843. [Google Scholar] [CrossRef] [PubMed]
- Annand, R.R.; Kontoyianni, M.; Penzotti, J.E.; Dudler, T.; Lybrand, T.P.; Gelb, M.H. Active site of bee venom phospholipase A2: The role of histidine-34, aspartate-64 and tyrosine-87. Biochemistry 1996, 35, 4591–4601. [Google Scholar] [CrossRef] [PubMed]


{kind=link}
| Adverse Effects | Specific Effects | Experimental System | Dose | Reference |
| Induction of Type 2 responses | Promote Th2 differentiation and ILC2 activation | mouse, in vivo, s.c. injection or mouse, in vivo, i.p. injection for 3 consecutive days | 50–100 µg/mouse | (Palm et al., 2013) [57] |
| Nociceptive effects | Induce paw oedema for less 3 h | rat, in vivo, injection into paw | 30 µg/paw | (Landucci et al., 2000) [71] |
| Neurotoxicity | Induce neuronal death | rat, in vivo, microinjection into spinal cord | 0.05–0.5 µg/rat | (Liu et al., 2006) [73] |
| Create demyelination and remyelination | rat, in vivo, microinjection into spinal cord | 1.5–6 ng/rat | (Titsworth et al., 2007) [74] | |
| Beneficial Effects | Specific Effects | Experimental System | Dose | Reference |
| Anti-inflammatory effects | Promote Treg differentiation | mouse, in vivo, i.p. injection | 0.1–1 mg/kg | (Chung et al., 2015) [60] |
| Supress airway inflammation | mouse, in vivo, i.p. injection | 0.2 mg/kg | (Park et al., 2015) [77] | |
| Protect cisplatin-induced renal inflammation | mouse, in vivo, i.p. injection | 0.2 mg/kg | (Kim et al., 2015) [59] | |
| Protect acetaminophen-induced liver inflammation | mouse, in vivo, i.p. injection | 0.2 mg/kg | (Kim et al., 2014) [78] | |
| Anti-nociceptive effects | Reduce oxaliplatin-induced neuropathic pain | mouse, in vivo, i.p. injection | 0.2 mg/kg | (Li et al., 2015) [84] |
| Anti-neuronal injury | Prevent MPTP-induced neurotoxicity | mouse, in vivo, i.p. injection | 0.2 mg/kg | (Chung et al., 2015) [60] |
| Inhibit PrP(106–126)-induced neuronal cell death | human neuroblastoma cell lines (SH-SY5Y), in vitro | 50 nM | (Jeong et al., 2011) [83] | |
| Anti-tumor effects | Inhibit growth of various cancer cell lines synergistically with PtdIns(3,4)P2 | A498, DU145, BEAS-2B, T-47D cell lines, in vitro | 10 µg/mL | (Putz et al., 2006) [86] |
| Inhibit A498 cell line growth synergistically with PtdIns(3,4)P2 | human kidney carcinoma cell line (A498), in vitro | 10 µg/mL | (Putz et al., 2006) [85] | |
| Anti-parasite effects | Inhibit ookinete binding on mosquito midgut | mosquito, ex vivo | 3.2 µM | (Zieler et al., 2001) [99] |
© 2016 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons by Attribution (CC-BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lee, G.; Bae, H. Bee Venom Phospholipase A2: Yesterday’s Enemy Becomes Today’s Friend. Toxins 2016, 8, 48. https://doi.org/10.3390/toxins8020048
Lee G, Bae H. Bee Venom Phospholipase A2: Yesterday’s Enemy Becomes Today’s Friend. Toxins. 2016; 8(2):48. https://doi.org/10.3390/toxins8020048
Chicago/Turabian StyleLee, Gihyun, and Hyunsu Bae. 2016. "Bee Venom Phospholipase A2: Yesterday’s Enemy Becomes Today’s Friend" Toxins 8, no. 2: 48. https://doi.org/10.3390/toxins8020048
APA StyleLee, G., & Bae, H. (2016). Bee Venom Phospholipase A2: Yesterday’s Enemy Becomes Today’s Friend. Toxins, 8(2), 48. https://doi.org/10.3390/toxins8020048