
Differential Properties of Venom Peptides and Proteins in Solitary vs. Social Hunting Wasps

Abstract
:1. Introduction

2. Venom of Hunting Wasps
2.1. General Properties and Origin
2.2. Solitary vs. Social Wasp Venom
3. Comparative Studies of Solitary vs. Social Wasp Venoms

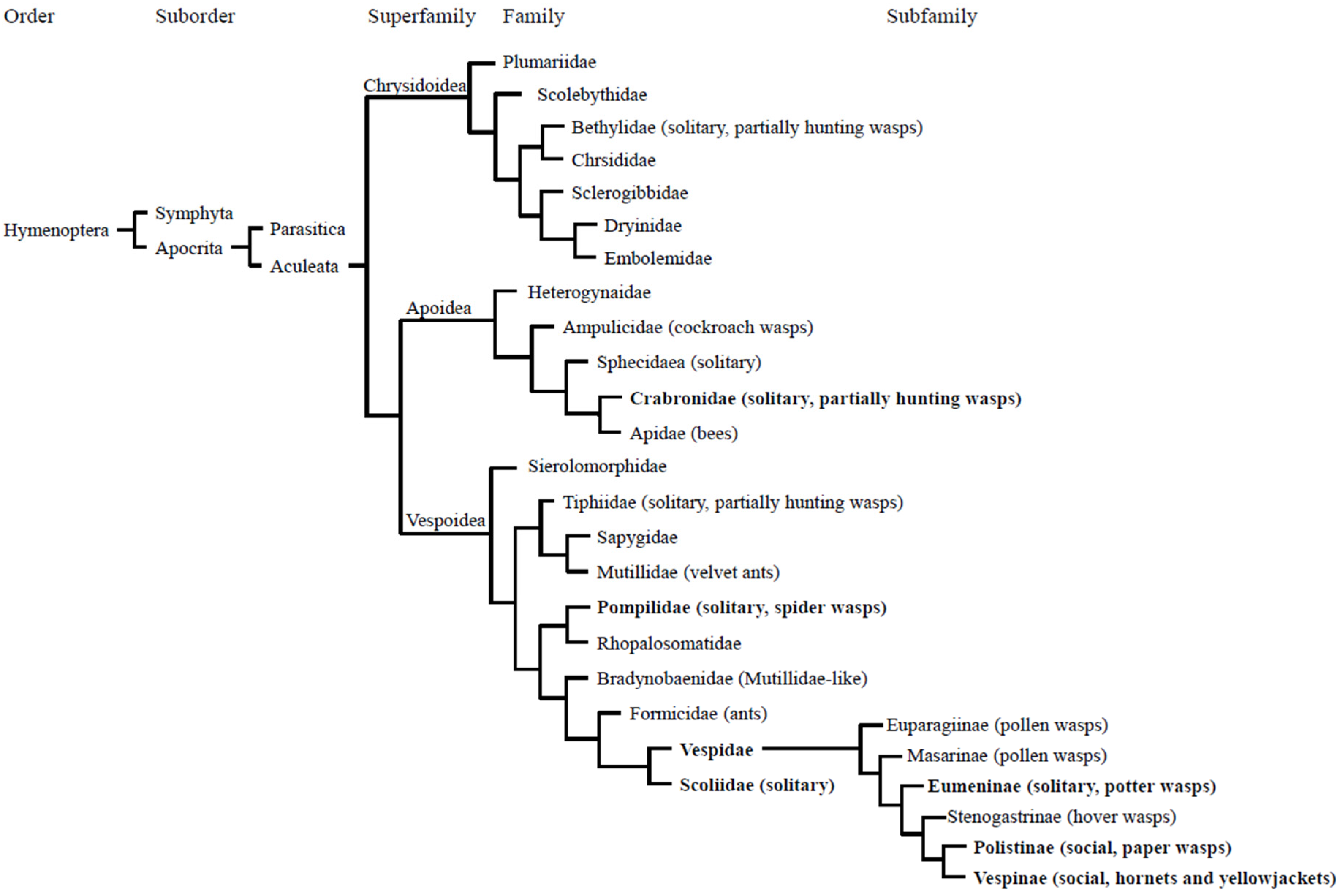{kind=link}
| Protein/Peptide | Putative Function a | Species b | References |
|---|---|---|---|
| Neurotoxins | |||
| α-pompilidiotoxin | Paralysis (Na+ channel blocking) | As | [23] |
| β-pompilidiotoxin | Paralysis (Na+ channel blocking) | Bm | [23] |
| Dendrotoxin-like | Paralysis (K+ channel blocking) | Ep | [16,18] |
| Kinins | |||
| Wasp kinin | Pain production | Ca, Cd, Ci, Cm, Mf, Mp | [24,25,26] |
| Mast cell degranulating peptides | |||
| Mastoparan-like | Allergic inflammation (Mast cell degranulation), Antimicrobial activity | Af, As, Ep, Er, Od1, Od2 | [16,18,27,28,29,30,31,32] |
| Chemotactic peptides | |||
| Wasp chemotactic peptide | Inflammatory activity Antimicrobial activity | Cf, Ep, Od1, Rb | [16,18,30,33] |
| Enzymes | |||
| Acetyl-CoA synthase | Involvement in metabolism of acetate | Rb | [19] |
| Alcohol dehydrogenase | Oxidation of ethanol to acetaldehyde | Ep | [18] |
| Amidophosphoribosyltransferase | Regulation of cell growth | Rb | [19] |
| Arginine kinase | Paralysis | Cd, Od1, Ep, Rb | [16,19,34] |
| ATP synthase | ATP synthesis | Od1, Ep, Rb | [17,18] |
| Carboxylesterase | Lipid metabolism | Rb | [19] |
| Citrate synthase | Catalyzing the citric acid cycle | Rb | [19] |
| Cytochrome P450 monooxygenase | Metabolism of toxic compounds | Od1, Rb | [17,19] |
| DNA-directed RNA polymerase | Synthesis of mRNA precursor | Rb | [19] |
| Farnesoic acid O-methyltransferase | Regulation of biosynthetic pathway of juvenile hormone | Rb | [19] |
| Glutamate decarboxylase | Involvement in beta-cell-specific autoimmunity | Ep | [18] |
| Glyceraldehyde-3-phosphate dehydrogenase | Direct hemolytic factor | Rb | [19] |
| Glycogenin | Synthesis of glycogen | Rb | [19] |
| HECT E3 ubiquitin ligase | Regulation of cell trafficking | Ep | [18] |
| Hyaluronidase | Venom dissemination | Ep, Rb | [18,19] |
| Myo inositol monophosphatase | Regulation of inositol homeostasis | Rb | [19] |
| Phospholipase A2 | Hydolysis of lecithins | Ep, Rb | [18,19] |
| Protein tyrosin phosphatase | Regulation of cellular processes | Rb | [19] |
| Serine/threonine protein phosphatase | Regulation of biochemical pathways | Rb | [19] |
| Tyrosine 3-monooxygenase | Regulation of dopamine synthesis | Od1, Ep | [16,18] |
| Hemostasis affecting proteins | |||
| Metalloendopeptidase | Inhibition of platelet aggregation | Od1, Ep, Rb | [18,19] |
| Neprilysin | Inhibition of platelet aggregation | Rb | [19] |
| Serine protease/Chymotrypsin/Thrombin-like | Fibrinolytic activity Kinin releasing activity Melanization | Od1, Ep | [16] |
| Muscle-related proteins | |||
| Actin | Regulation of hemocyte cytoskeleton gene expression | Od1, Ep, Rb | [16,19] |
| Ankyrin | Attachment of membrane proteins to membrane cytoskeleton | Rb | [19] |
| Bmkettin | Development of flight muscles | Od1, Rb | [17,19] |
| Calponin | Regulation of myogenesis | Od1, Ep, Rb | [17,18,19] |
| Muscle LIM protein | Regulation of myogenesis | Od1, Ep, Rb | [17,19] |
| Muscle protein 20 Myomesin | Regulation of muscle cotraciton | Od1, Ep | [17,18] |
| Anchoring the thick filaments | Rb | [19] | |
| Myosin heavy chain | Regulation of muscle functions | Od1, Ep, Rb | [17,18,19] |
| Myosin light chain | Modulation of the affinity of myosin for actin | Od1, Ep, Rb | [17,18,19] |
| Paramyosin | Regulation of thick filament in muscles | Od1, Ep, Rb | [17,18,19] |
| Titin | Assembly of contractile machinery in muscle cells | Od1, Rb | [17,19] |
| Tropomyosin | Muscle contraction | Od1, Rb | [17,19] |
| Troponin | Muscle contraction | Od1, Ep, Rb | [17,18,19] |
| Tubulin | Regulation of hemocyte skeleton genes expression | Od1, Ep, Rb | [17,19] |
| Other proteins/peptides | |||
| Chemosensory protein | Transferring metabolism-related small molecules | Rb | [19] |
| Cytochrom C | Protein wire | Od1, Rb | [17,19] |
| Heat shock proteins | Prevention of protein misfolding | Od1, Ep, Rb | [16,19] |
| Insulin-like peptide binding protein | Developmental arrest (Inhibition of insulin signaling) | Ep | [18] |
| Sialin | Nitrate transporter | Rb | [19] |
| Sugar transporter | Maintenance of glucose homeostasis | Rb | [19] |
| Protein/Peptide | Putative Function a | Species b | References |
|---|---|---|---|
| Neurotoxins | |||
| AvTx-7,8 | Paralysis (K+ channel blocking) | Av | [35,36] |
| Agatoxin-like | Paralysis (Ca2+ channel blocking) | Vv1 | [21] |
| Analgesic polypeptide | Paralysis (Na+ channel blocking) | Vv1 | [21] |
| Calsyntenin | Paralysis (Ca2+ channel blocking) | Vc | [20] |
| Conophysin-R | Paralysis (Ca2+ channel blocking) | Vv1 | [21] |
| Latrotoxin-like | Channel formation | Vv1 | [21] |
| Leucine rich repeat domain-containing protein | Paralysis (Involvement in synaptic vescle trafficking) | Va2, Vc | [20] |
| Orientotoxin-like | Paralysis (Presynaptic effect, lysophospholipase activity) | Vo, Vv1 | [21,37] |
| Kinins | |||
| Wasp kinin | Pain production | Pa, Pc, Pe1, Pe2, Pf, Pi, Pj, Pm1, Pm2, Pp, Pr, Va2, Vc, Vm2, Vt, Vx | [20,25,38,39,40,41] |
| Mast cell degranulating peptides | |||
| Mastoparan | Allergic inflamation (Mast cell degranulation) | Ap, Pe2, Pi, Pj, Pm2, Pp, Ps, Rs, Va1, Va2, Vb1, Vb2, Vc, Vd, Vl, Vm1, Vm2, Vo, Vt, Vv1, Vx | [19,20,38,42,43,44,45,46,47,48,49,50,51,52,53] |
| Chemotactic peptides | |||
| Wasp chemotactic peptide | Inflammatory activity Antimicrobial activity | Ap, Pl, Pm2, Pp, Ps, Va2, Vb2, Vc, Vm1, Vm2, Vo, Vt, Vx | [20,38,40,41,45,49,50,52,54] |
| Enzymes | |||
| Acetylcholinesterase | Pain processing (Hydrolysis of neurotransmitter) | Va2, Vc, Vv1 | [20,21] |
| Acetyltransferase | Synthesis of acetylcholine | Vt | [19] |
| Acid phosphatase | Female reproduction | Va2, Vc | [20] |
| Acyl-CoA delta-9 desaturase | Insertion of double bond in fatty acids | Vt | [19] |
| AMP dependent coa ligase | Production of fatty acyl-CoA esters | Vt | [19] |
| Arginine kinase | Paralysis | Va2, Vc | [20] |
| Argininosuccinate synthase | Arginine synthesis | Vt | [19] |
| ATP-dependent protease | Mediation of protein quality | Vt | [19] |
| Carboxylesterase | Lipid metabolism | Va2, Vc | [20] |
| Chitinase | Chitinolysis | Va2, Vt | [19,20] |
| Core alpha 1,3-fructosyltransferase A | Glycoprotein production | Vt | [19] |
| Cytochrome P450 monooxygenase | Metabolism of toxic compounds | Vt | [20] |
| Dipeptidyl peptidase IV | Liberation of bioactive peptides | Va2, Vb1 | [20,44] |
| Esterase FE4 | Sequestration | Vc | [20] |
| Fatty acid synthase | Biosynthesis of hormones | Vt | [19] |
| Fibrinogenase brevinase | Fibrinolysis | Vv1 | [21] |
| Glyceraldehyde-3-phosphate dehydrogenase Glycerol-3-phosphate acyltransferase Glycogenin GTP cyclohydrolase I isoform A | Direct hemolytic factor | Va2, Vc | [20] |
| Synthesis of triacylglycerol | Vt | [19] | |
| Synthesis of glycogen | Va2, Vc | [20] | |
| Production of neurotransmitter | Vt | [19] | |
| Hyaluronidase | Venom dissemination | Dm, Pa, Pp, Va2, Vc, Vm1, Vt, Vv3 | [19,20,55,56,57] |
| Laccase | Oxidation, cuticle sclerotization | Vc | [20] |
| Myosin light chain kinase | Muscle contraction | Va2, Vc | [20] |
| O-linked n-acetylglucosamine transferase | Insulin signaling reduction | Vt | [19] |
| Peptidyl-prolyl cis-trans isomerase | Immune mediator | Vt | [19] |
| Phospholipase A1 | Production of lipid mediator | Dm, Pa, Va1, Va2, Vc, Vv3 | [20,57,58] |
| Phospholipase A2 | Hydrolysis of lecithins | Va2, Vc, Vv1 | [20,21] |
| Phospholipase B1 | Hydrolysis of lysolecithins | Va2 | [20] |
| Phospholipase D | Induction of inflammatory responses | Va2, Vc | [20] |
| Phospholipase DDHD | Synaptic organization | Va2, Vc | [20] |
| Purine nucleoside phosphorylase | Apoptosis of lymphocytes | Vt | [19] |
| Reverse transcriptase | Production of high venom yield | Vt | [19] |
| Thrombin-like enzyme | Coagulation factor | Va2, Vv1 | [20,21] |
| γ-glutamyl transpeptidase | Apoptosis of ovariole cells | Va2, Vc | [20] |
| CAP superfamily | |||
| Defensin | Antimicrobial activity | Va2, Vc | [20] |
| Venom allergen 5 | Allergenic activity | Dm, Pa, Pe1, Pf, Va2, Vc, Vf, Vg, Vm1, Vm3, Vp, Vs, Vt, Vv2, Vv3 | [20,56,57,59,60,61,62] |
| Hemostasis affecting proteins | |||
| Blarina toxin | Production of kinins | Vv1 | [21] |
| Coagulation factor Disintegrin | Platelet aggregation Platelet aggregation | Vv1 Va2 | [21] [20] |
| Factor V activator | Coagulation factor | Vv1 | [21] |
| Lectoxin-Enh4 Metalloendopeptidase Nematocyte expressed protein-6 Neprilysin | Anticoagulant factor Inhibition of platelet aggregation Inhibition of platelet aggregation Inhibition of platelet aggregation | Vv1 Va2, Vc, Vv1 Vv1 Va2, Vc | [21] [20,21] [21] [20] |
| Oscutarin-C | Fibrinolysis | Vv1 | [21] |
| Ryncolin-3/4 | Platelet aggregation | Vv1 | [21] |
| Serine protease/Chymotrypsin/Thrombin-like | Fibrinolytic activity Kinin releasing activity Melanization | Va2, Vc, Vm1, Vt, Vv1 | [19,20,21,63] |
| Snaclec | Platelet aggregation | Vv1 | [21] |
| Vescular endothelial growth factor | Coagulation factor | Vv1 | [21] |
| Veficolin | Platelet aggregation | Vv1 | [21] |
| Venom plasminogen activator | Fibrinolysis | Vv1 | [21] |
| Venom prothrombin activator | Fibrinolysis | Vv1 | [21] |
| Muscle-related proteins | |||
| Actin | Expression of hemocyte cytoskeleton | Va2, Vc | [20] |
| Calponin | Binding with actin | Va2, Vc | [20] |
| Muscle LIM protein | Regulation of myogenesis | Va2, Vc | [20] |
| Myosin heavy chain | Regulation of muscle functions | Va2, Vc | [20] |
| Paramyosin | Regulation of thick filament in muscle | Va2, Vc | [20] |
| Tropomyosin | Muscle contraction | Va2, Vc | [20] |
| Troponin | Muscle contraction | Va2, Vc | [20] |
| Vespin | Smooth muscle contraction | Vm1 | [64] |
| Protease inhibitor | |||
| Leukocyte elastase inhibitor isoform | Reduction of tissue damage | Vc | [20] |
| Serpin | Immune suppression (Inhibition of melanization) | Va2, Vc | [20] |
| Other proteins/peptides | |||
| Anaphase-promoting complex subunit 13 | Protein degradation | Vt | [19] |
| Apolipophorin-III | Lipid transport | Vt | [19] |
| Bhlh factor math 6 | Regulation of developmental process | Vt | [19] |
| Bombolitin | Antimicrobial activity | Vc | [20] |
| CRAL/TRIO domain-containing protein | Regulation of cell growth | Vt | [19] |
| Cytochrome b | Transferring electrons | Vt | [19] |
| Doublesex isoform 1 | Sex determination factor | Vt | [19] |
| Ejaculatory bulb-specific protein 3 | Odorant binding protein | Va2, Vc | [20] |
| Elongation factor 2 | Protein synthesis | Va2, Vc | [20] |
| Endopeptidase inhibitor | Inhibition of atrial natriuretic peptides | Vt | [21] |
| Endoplasmin | Protein folding | Va2, Vc | [20] |
| ETR-3 like factor 2 | Pre-mRNA alternative splicing | Vt | [19] |
| Gigantoxin-1 | Hemolytic activity | Vv1 | [21] |
| Growth hormone inducible transmembrane protein | Apoptosis | Vt | [19] |
| GTPase-activating protein | Regulation of G protein signaling | Va2 | [20] |
| Heat shock proteins | Prevention of protein misfolding | Vt | [19] |
| Insulin binding protein | Inhibition of insulin signaling | Va2, Vc | [20] |
| NADH-ubiquinone oxidoreductase chain 4 | Involvement in respiratory chain | Vt | [19] |
| Natterin-4 | Kininogenase activity | Vv1 | [21] |
| Peptidoglycan-recognition protein 1 | Antimicrobial activity | Vt | [19] |
| Phd finger protein | Protein-protein interaction | Vt | [19] |
| Plancitoxin | DNase activity | Vv1 | [21] |
| Polyubiquitin | Proteolysis | Vt | [19] |
| SE-cephalotoxin | Paralysis | Vv1 | [21] |
3.1. High Throughput Identification of Wasp Venom Components
3.2. Comparative Aspects of Venom Components between Solitary vs. Social Wasp
3.2.1. Solitary Wasp Venom-Specific Features
3.2.2. Social Wasp Venom-Specific Features
4. Venom Peptides
4.1. Neurotoxic Peptides
4.2. Kinins
4.3. Mastoparans
4.4. Chemotactic Peptides
4.5. Other Venom Peptides
5. Useful Wasp Venom Components for Pharmacological, Medical, and Agricultural Applications
5.1. Antimicrobial Agents
5.2. Antitumor Agents
5.3. Venom Allergy Diagnosis and Immunotherapy
5.4. Biopesticides
6. Concluding Remarks
Acknowledgments
Author Contributions
Conflicts of Interest
Appendix A
| Sociality | Name | Sequence | Length (a.a) | Species | References |
|---|---|---|---|---|---|
| Neurotoxins | |||||
| Solitary | α-PMTX | RIKIGLFQDLSKL | 13 | Anoplius samariensis | [23] |
| β-PMTX | RIKIGLFQDLSRL | 13 | Batozonellus maculifrons | [23] | |
| Social | AvTx-7 | 1210 Da (α-PMTX 1530 Da) | - | Agelaia vicina | [36] |
| AvTx-8 | 1567 Da | - | Agelaia vicina | [35] | |
| Kinins | |||||
| Solitary | Bradykinin (BK) | RPPGFSPFR | 9 | mammal | - |
| Megascoliakinin | RPPGFTPFRKA | 11 | Megascolia flavifrons | [145] | |
| Bradykinin | RPPGFSPFR | 9 | Megacampsomeris prismatica | [24] | |
| Thr6-BK | RPPGFTPFR | 9 | Megacampsomeris prismatica, Campsomeriella annulata annulata, Carinoscolia melanosoma fascinate, Cyphononyx dorsalis, Megascolia flavifrons, Colpa interrupta | [24,105,145] | |
| Social | RA-Thr6-Bradykinin | RARPPGFTPFR | 11 | Polybia paulista | [40] |
| RA-Thr6-Bradykinin-DT | RARPPGFTPFRDT | 13 | Polybia paulista | [40] | |
| Vespakinin-M | GRPHypGFSPFRID | 14 | Vespa mandarinia | [146] | |
| Vespakinin-X | ARPPGFSPFRIV | 12 | Vespa xanthoptera | [147] | |
| Vespakinin-A | GRPPGFSPFRVI | 12 | Vespa analis | [148] | |
| Vespakinin-AP ** | ELPPGFTPFRII | 12 | Vespa analis parallela | [20] | |
| Vespakinin-T | GRPHypGFSPFRVI | 12 | Vespa tropica | [101] | |
| Vespakinin-C | KLPPGFTPFRII | 12 | Vespa crabro flavogasciata | [20] | |
| Vespulakinin | TAT(carbhy)T(carbhy)RRRGRPPGFSPFR | 17 | (Para)Vespula maculifrons | [149] | |
| Vespulakinin-L | TAR(NAcGal-Gal)TKRRGRPPGFSPFR | 17 | Vespula lewisii | [101] | |
| Polisteskinin 3 | PyrTNKKKLRGRPPGFSPFR | 18 | Polistes exclamans, Polistes annularis, Polistes fuscramatus | [25] | |
| Polisteskinin-R | ARRPPGFTPFR | 11 | Polistes rothneyi | [25] | |
| Polisteskinin-J | RRRPPGFT(S)PFR | 11 | Polistes jadwigae | [101] | |
| Polisteskinin-C | SKRPPGFSPFR | 11 | Polistes chnensis | [101] | |
| PMM1 | KRRPPGFTPFR | 11 | Polistes major major | [38] | |
| Protopolybiakinin-I | DKNKKPIRVGGRRPPGFTR | 19 | Protopolybia exigua | [39] | |
| Protopolybiakinin-II | DKNKKPIWMAGFPGFTPIR | 19 | Protopolybia exigua | [39] | |
| Mastoparan-like Peptides | |||||
| Solitary | EMP-AF | INLLKIAKGIIKSL-NH2 | 14 | Anterhynchium flavormarginatum micado | [150] |
| Eumenitin | LNLKGIFKKVASLLT | 15 | Eumenes rubronotatus | [29] | |
| EMP-OD (OdVP1) | GRILSFIKGLAEHL-NH2 | 14 | Orancistrocerus drewseni | [30,66,114] | |
| OdVP3 a | KDLHTVVSAILQAL-NH2 | 14 | Orancistrocerus drewseni | [66,114] | |
| EpVP1 a | INLKGLIKKVASLLT | 15 | Eumenes pomiformis | [18,114] | |
| EpVP2a a | FDLLGLVKKVASAL-NH2 | 14 | Eumenes pomiformis | [18,114] | |
| EpVP2b a | FDLLGLVKSVVSAL-NH2 | 14 | Eumenes pomiformis | [18,114] | |
| Social | Mastoparan (MP) | INLKALAALAKKIL-NH2 | 14 | Vespula lewisii | [110] |
| Mastoparan-X | INWKGIAAMAKKLL-NH2 | 14 | Vespa xanthoptera | [46] | |
| Mastoparan-A | IKWKAILDAVKKVL(I)-NH2 | 14 | Vespa analis | [20,30] | |
| Mastoparan-B | LKLKSIVSWAKKVL-NH2 | 14 | Vespa basalis | [30] | |
| Mastoparan-C | INW(L)KALLAVAKKIL-NH2 | 14 | Vespa crabro | [20,30] | |
| Mastoparan-II | INLKALAALVKKVL-NH2 | 14 | Vespa orientalis | [30] | |
| HR1 | INLKAIAALVKKVL-NH2 | 14 | Vespa orientalis | [37] | |
| Mastoparan-T1 * | INLKVFAALVKKFL-NH2 | 14 | Vespa tropica | [19] | |
| Mastoparan-T2 * | INLKVFAALVKKLL-NH2 | 14 | Vespa tropica | [19] | |
| Mastoparan-T3 * | INLRGFAALVKKFL-NH2 | 14 | Vespa tropica | [19] | |
| Mastoparan-T4 * | INLFGFAALVKKFL-NH2 | 14 | Vespa tropica | [19] | |
| protopolybia-MP I | INWLKLGKKVSAIL-NH2 | 14 | Protopolybia exigua | [46] | |
| protopolybia-MP II | INWKAIIEAAKQAL-NH2 | 14 | Protopolybia exigua | [46] | |
| protopolybia-MP III | INWLKLGKAVIDAL-NH2 | 14 | Protopolybia exigua | [46] | |
| P-8 | INWKALLDAAKKVL-NH2 | 14 | Protonectarina sylveirae | [151] | |
| polybia-MP I | IDWKKLLDAAKQIL-NH2 | 14 | Polybia paulista | [43] | |
| polybia-MP II | INWLKLGKMVIDAL-NH2 | 14 | Polybia paulista | [42] | |
| polybia-MP III | IDWLKLGKMVMDVL-NH2 | 14 | Polybia paulista | [42] | |
| polybia-MP IV | IDWLKLRVISVIDL-NH2 | 14 | Polybia paulista | [40] | |
| polybia-MP V | INWHDIAIKNIDAL-NH2 | 14 | Polybia paulista | [40] | |
| polybia-MP VI | IDWLKLGKMVM | 11 | Polybia paulista | [40] | |
| parapolybia-MP | INWKKMAATALKMI-NH2 | 14 | Parapolybia indica | [46] | |
| parapolybia-MP | INWAKLGKLALEVI-NH2 | 14 | Parapolybia indica | [30] | |
| Ropalidia-MP | INWAKLGKLALQAL-NH2 | 14 | Ropalidia | [46] | |
| polistes-MP | VDWKKIGQHILSVL-NH2 | 14 | Polistes jadwigae | [46] | |
| PMM2 | INTKKIASIGKEVLKAL-NH2 | 17 | Polistes major major | [38] | |
| Agelaia MP-I | INWLKLGKAIIDAL-NH2 | 14 | Agelaia pallipes pallipes | [52] | |
| Chemotactic Peptides | |||||
| Solitary | OdVP2 (Orancis-protonectin) | ILGIITSLLKSL-NH2 | 12 | Orancistrocerus drewseni | [30,66,114] |
| EpVP6 b | FGPVIGLLSGILKSLL | 16 | Eumenes pomiformis | [18,114] | |
| RbVP1 *,b | FLGGLIKGLVKAL-NH2 | 13 | Rhynchium brunneum | [19] | |
| Social | Protonectin | ILGTILGLLKGL-NH2 | 12 | Protonectarina sylveirae, Agelaia pallipes pallipes | [52] |
| Protonectin(1-6) | ILGTIL-NH2 | 6 | Agelaia pallipes pallipes | [152] | |
| Paulista-CP (polybia-CP) | ILGTILGLLKSL-NH2 | 12 | Polybia paulista | [43] | |
| Polybia-CP 2 | ILGTILGKIL | 10 | Polybia paulista | [40] | |
| Polybia-CP 3 | ILGTILGTFKSL-NH2 | 12 | Polybia paulista | [40] | |
| Crabrolin | FLPLILRKIVTAL-NH2 | 13 | Vespa crabro | [153] | |
| Ves-CP-T | FLPILGKILGGLL-NH2 | 13 | Vespa tropica | P17231, [19] | |
| Ves-CP-T2 * | FLPIIGKLLSGLL-NH2 | 13 | Vespa tropica | [19] | |
| Ves-CP-M | FLPIIGKLLSGLL-NH2 | 13 | Vespa mandarina | P17232 | |
| Ves-CP-A | FLPMIAKLLGGLL-NH2 | 13 | Vespa analis, Vespa analis parallela | P17233, [20] | |
| Ves-CP-X | FLPIIAKLLGGLL-NH2 | 13 | Vespa xanthoptera | P17234 | |
| Ves-CP-L | FLPIIAKLVSGLL-NH2 | 13 | Vespula lewisi | P17235 | |
| VCP-5e | FLPIIAKLLGGLL-NH2 | 13 | Vespa magnifica | [45] | |
| VCP-5f | FLPIPRPILLGLL-NH2 | 13 | Vespa magnifica | [45] | |
| VCP-5g | FLIIRRPIVLGLL-NH2 | 13 | Vespa magnifica | [45] | |
| VCP-5h | FLPIIGKLLSGLL-NH2 | 13 | Vespa magnifica | [45] | |
| HP-1 | LFRLIAKTLGSLM | 13 | Vespa basalis | [154] | |
| HP-2 | LFRLLANTLGKIL | 13 | Vespa basalis | [154] | |
| HP-3 | IFGLLAKTLGNLF | 13 | Vespa basalis | [154] | |
| HR2 | FLPLILGKLVKGLL-NH2 | 14 | Vespa orientalis | [37] | |
| PMM3 | FLSALLGMLKNL-NH2 | 12 | Polistes major major | [38] | |
| Uncategorized Peptides | |||||
| Solitary | Anoplin | GLLKRIKTLL-NH2 | 10 | Anoplius samariensis | [27] |
| Decoralin | SLLSLIRKLIT-NH2 | 11 | Oreumenes decoratus | [28] | |
| OdVP4 | LDPKVVQSLL-NH2 | 10 | Orancistrocerus drewseni | [66,114] | |
| EpVP3 | AINPKSVQSLL-NH2 | 11 | Eumenes pomiformis | [18,114] | |
| EpVP3S | INPKSVQSLL-NH2 | 10 | Eumenes pomiformis | [18,114] | |
| EpVP4a | LSPAVMASLA-NH2 | 10 | Eumenes pomiformis | [18,114] | |
| EpVP4b | LSPAAMASLA-NH2 | 10 | Eumenes pomiformis | [18,114] | |
| EpVP5 | VHVPPICSHRECRK | 14 | Eumenes pomiformis | [18,114] | |
| As-peptide126 | QDPPVVKMK-NH2 | 9 | Anoplius samariensis | BAF65255 | |
| Cd-125 | DTARLKWH | 8 | Cyphononyx dorsalis | [107] | |
| Cd-146 | SETGNTVTVKGFSPLR | 16 | Cyphononyx dorsalis | [107] | |
| EpDTX | IATICNLPIVSGNGQEEHIRWAYSIITHVCVSFRYTGKGGNRNNFFTERECRSYCYF | 57 | Eumenes pomiformis | [18,114] | |
| As-fr-19 | VSFCLLPIVPGPCTQYVIRYAFQPSISACRRFTFGGCEGNDNNFMTRRDCEHYCEELL | 58 | Anoplius samariensis | [79] | |
| Social | Vespin | CYQRRVAITAGGLKHRLMSSLIIIIIIRINYLRDNSVIILESSY | 44 | Vespa magnifica | [64] |
References
- Piek, T. Venoms of the Hymenoptera: Biochemical, Pharmacological, and Behavioural Aspects; Academic Press: London, UK, 1986; p. 570. [Google Scholar]
- Brothers, D.J. Phylogeny and evolution of wasps, ants and bees (Hymenoptera, Chrysidoidea, Vespoidea and Apoidea). Zool. Scr. 1999, 28, 233–249. [Google Scholar] [CrossRef]
- Dowton, M.; Austin, A.D. Simultaneous analysis of 16S, 28S, COI and morphology in the Hymenoptera: Apocrita—Evolutionary transitions among parasitic wasps. Biol. J. Linn. Soc. 2001, 74, 87–111. [Google Scholar]
- O’Neill, K.M. Solitary Wasps: Behavior and Natural History; Comstock Pub. Associates: Ithaca, NY, USA, 2001; p. 406. [Google Scholar]
- Moreau, S.J.M.; Asgari, S. Venom proteins from parasitoid wasps and their biological functions. Toxins 2015, 7, 2385–2412. [Google Scholar] [CrossRef] [PubMed]
- Evans, D.L.; Schmidt, J.O. Insect Defenses: Adaptive Mechanisms and Strategies of Prey And Predators; State University of New York Press: Albany, NY, USA, 1990; p. 482. [Google Scholar]
- Spradbery, J.P. Wasps: An Account of the Biology and Natural History of Solitary and Social Wasps; University of Washington Press: Seattle, WA, USA, 1973; p. 408. [Google Scholar]
- Hunting Wasp. Available online: http://en.wikipedia.org/wiki/Hunting_wasp (accessed on 16 December 2015).
- Tree of Life Web Project. 1995. Aculeata. Version 01 January 1995 (Temporary). Available online: http://tolweb.Org/aculeata/11184/1995.01.01 in the Tree of Life Web Project; http://tolweb.Org/.; http://tolweb.org/Aculeata/11184/1995 (accessed on 17 December 2015).
- Clemetson, K.; Kini, R.M. Introduction. In Toxins and Hemostasis: From Bench to Bedside; Kini, R.M., Clemetson, K.J., Markland, F.S., McLane, M.A., Morita, T., Eds.; Springer: Dordrecht, The Netherlands, 2011; pp. 1–9. [Google Scholar]
- Fry, B.G.; Roelants, K.; Champagne, D.E.; Scheib, H.; Tyndall, J.D.A.; King, G.F.; Nevalainen, T.J.; Norman, J.A.; Lewis, R.J.; Norton, R.S.; et al. The toxicogenomic multiverse: Convergent recruitment of proteins into animal venoms. Annu. Rev. Genomics Hum. Genet. 2009, 10, 483–511. [Google Scholar] [CrossRef] [PubMed]
- Fry, B.G.; Roelants, K.; Norman, J.A. Tentacles of venom: Toxic protein convergence in the Kingdom Animalia. J. Mol. Evol. 2009, 68, 311–321. [Google Scholar] [CrossRef] [PubMed]
- Lai, R.; Liu, C. Bioactive peptides and proteins from wasp venoms. In Toxins and Hemostasis: From Bench to Bedside; Kini, R.M., Clemetson, K.J., Markland, F.S., McLane, M.A., Morita, T., Eds.; Springer: Dordrecht, The Netherlands, 2011; pp. 83–96. [Google Scholar]
- Palma, M.S. Chapter 56—Insect venom peptides. In Handbook of Biologically Active Peptides; Kastin, A.J., Ed.; Academic Press: Burlington, NJ, USA, 2006; pp. 389–396. [Google Scholar]
- National center for biotechnology information. Available online: http://www.ncbi.nlm.nih.gov/ (accessed on 16 December 2015).
- Baek, J.H.; Lee, S.H. Identification and characterization of venom proteins of two solitary wasps. Toxicon 2010, 56, 554–562. [Google Scholar] [CrossRef] [PubMed]
- Baek, J.H.; Woo, T.H.; Kim, C.B.; Park, J.H.; Kim, H.; Lee, S.; Lee, S.H. Differential gene expression profiles in the venom gland/sac of Orancistrocerus drewseni (Hymenoptera: Eumenidae). Arch. Insect Biochem. Physiol. 2009, 71, 205–222. [Google Scholar] [CrossRef] [PubMed]
- Baek, J.H.; Lee, S.H. Differential gene expression profiles in the venom gland/sac of Eumenes pomiformis (Hymenoptera: Eumenidae). Toxicon 2010, 55, 1147–1156. [Google Scholar] [CrossRef] [PubMed]
- Baek, J.H.; Oh, J.H.; Kim, Y.H.; Lee, S.H. Comparative transcriptome analysis of the venom sac and gland of social wasp Vespa tropica and solitary wasp Rhynchium brunneum. J. Asia Pac. Entomol. 2013, 16, 497–502. [Google Scholar] [CrossRef]
- Yoon, K.A.; Kim, K.; Nguyen, P.; Seo, J.B.; Park, Y.H.; Kim, K.-G.; Seo, H.-Y.; Koh, Y.H.; Lee, S.H. Comparative functional venomics of social hornets Vespa crabro and Vespa analis. J. Asia Pac. Entomol. 2015, 18, 815–823. [Google Scholar] [CrossRef]
- Liu, Z.R.; Chen, S.G.; Zhou, Y.; Xie, C.H.; Zhu, B.F.; Zhu, H.M.; Liu, S.P.; Wang, W.; Chen, H.Z.; Ji, Y.H. Deciphering the venomic transcriptome of killer-wasp Vespa velutina. Sci Rep. 2015, 5. [Google Scholar] [CrossRef] [PubMed]
- Dos Santos, L.D.; Santos, K.S.; Pinto, J.R.A.; Dias, N.B.; Souza, B.M.D.; dos Santos, M.F.; Perales, J.; Domont, G.B.; Castro, F.M.; Kalil, J.E.; et al. Profiling the proteome of the venom from the social wasp Polybia paulista: A clue to understand the envenoming mechanism. J. Proteome Res. 2010, 9, 3867–3877. [Google Scholar] [CrossRef] [PubMed]
- Konno, K.; Hisada, M.; Itagaki, Y.; Naoki, H.; Kawai, N.; Miwa, A.; Yasuhara, T.; Takayama, H. Isolation and structure of pompilidotoxins, novel peptide neurotoxins in solitary wasp venoms. Biochem. Biophys. Res. Commun. 1998, 250, 612–616. [Google Scholar] [CrossRef] [PubMed]
- Konno, K.; Palma, M.S.; Hitara, I.Y.; Juliano, M.A.; Juliano, L.; Yasuhara, T. Identification of bradykinins in solitary wasp venoms. Toxicon 2002, 40, 309–312. [Google Scholar] [CrossRef]
- Piek, T. Wasp kinins and kinin analogues. In Animal Toxins: Facts and Protocols; Rochat, H., Martin-Eauclaire, M.-F., Eds.; Birkhauser Verlag: Boston, MA, USA, 2000; pp. 99–115. [Google Scholar]
- Piek, T.; Mantel, P.; van Ginkel, C.J.W. Megascoliakinin, a bradykinin-like compound in the venom of Megascolia flavifrons fab. (Hymenoptera: Scoliidae). Comp. Biochem. Physiol. C 1984, 78, 473–474. [Google Scholar] [CrossRef]
- Konno, K.; Hisada, M.; Fontana, R.; Lorenzi, C.C.; Naoki, H.; Itagaki, Y.; Miwa, A.; Kawai, N.; Nakata, Y.; Yasuhara, T.; et al. Anoplin, a novel antimicrobial peptide from the venom of the solitary wasp Anoplius samariensis. Biochim. Biophys. Acta 2001, 1550, 70–80. [Google Scholar] [CrossRef]
- Konno, K.; Rangel, M.; Oliveira, J.S.; dos Santos Cabrera, M.P.; Fontana, R.; Hirata, I.Y.; Hide, I.; Nakata, Y.; Mori, K.; Kawano, M.; et al. Decoralin, a novel linear cationic α-helical peptide from the venom of the solitary Eumenine wasp Oreumenes decoratus. Peptides 2007, 28, 2320–2327. [Google Scholar] [CrossRef] [PubMed]
- Konno, K.; Hisada, M.; Naoki, H.; Itagaki, Y.; Fontana, R.; Rangel, M.; Oliveira, J.S.; Cabreraf, M.P.D.; Neto, J.R.; Hide, I.; et al. Eumenitin, a novel antimicrobial peptide from the venom of the solitary Eumenine wasp Eumenes rubronotatus. Peptides 2006, 27, 2624–2631. [Google Scholar] [CrossRef] [PubMed]
- Murata, K.; Shinada, T.; Ohfune, Y.; Hisada, M.; Yasuda, A.; Naoki, H.; Nakajima, T. Novel mastoparan and protonectin analogs isolated from a solitary wasp, Orancistrocerus drewseni drewseni. Amino Acids 2009, 37, 389–394. [Google Scholar] [CrossRef] [PubMed]
- Cabrera, M.P.D.; de Souza, B.M.; Fontana, R.; Konno, K.; Palma, M.S.; de Azevedo, W.F.; Neto, J.R. Conformation and lytic activity of Eumenine mastoparan: A new antimicrobial peptide from wasp venom. J. Pept. Res. 2004, 64, 95–103. [Google Scholar] [CrossRef] [PubMed]
- Sforca, M.L.; Oyama, S.; Canduri, F.; Lorenzi, C.C.B.; Pertinhez, T.A.; Konno, K.; Souza, B.M.; Palma, N.S.; Neto, J.R.; Azevedo, W.F.; et al. How C-terminal carboxyamidation alters the biological activity of peptides from the venom of the Eumenine solitary wasp. Biochemistry 2004, 43, 5608–5617. [Google Scholar] [CrossRef] [PubMed]
- Picolo, G.; Hisada, M.; Moura, A.B.; Machado, M.F.M.; Sciani, J.M.; Conceicao, I.M.; Melo, R.L.; Oliveira, V.; Lima-Landman, M.T.R.; Cury, Y.; et al. Bradykinin-related peptides in the venom of the solitary wasp Cyphononyx fulvognathus. Biochem. Pharmacol. 2010, 79, 478–486. [Google Scholar] [CrossRef] [PubMed]
- Yamamoto, T.; Arimoto, H.; Kinumi, T.; Oba, Y.; Uemura, D. Identification of proteins from venom of the paralytic spider wasp, Cyphononyx dorsalis. Insect Biochem. Mol. 2007, 37, 278–286. [Google Scholar] [CrossRef] [PubMed]
- De Oliveira, L.; Cunha, A.O.; Mortari, M.R.; Pizzo, A.B.; Miranda, A.; Coimbra, N.C.; dos Santos, W.F. Effects of microinjections of neurotoxin AvTx8, isolated from the social wasp Agelaia vicina (Hymenoptera, Vespidae) venom, on GABAergic nigrotectal pathways. Brain Res. 2005, 1031, 74–81. [Google Scholar] [CrossRef] [PubMed]
- Pizzo, A.B.; Beleboni, R.O.; Fontana, A.C.; Ribeiro, A.M.; Miranda, A.; Coutinho-Netto, J.; dos Santos, W.F. Characterization of the actions of AvTx 7 isolated from Agelaia vicina (Hymenoptera: Vespidae) wasp venom on synaptosomal glutamate uptake and release. J. Biochem. Mol. Toxicol. 2004, 18, 61–68. [Google Scholar] [CrossRef] [PubMed]
- Tuichibaev, M.U.; Akhmedova, N.U.; Kazakov, I.; Korneev, A.S.; Gagel’gans, A.I. Low molecular weight peptides from the venom of the giant hornet Vespa orientalis. Structure and function. Biokhimiia 1988, 53, 219–226. [Google Scholar] [PubMed]
- Cerovsky, V.; Pohl, J.; Yang, Z.; Alam, N.; Attygalle, A.B. Identification of three novel peptides isolated from the venom of the neotropical social wasp Polistes major major. J. Pept. Sci. 2007, 13, 445–450. [Google Scholar] [CrossRef] [PubMed]
- Mendes, M.A.; Palma, M.S. Two new bradykinin-related peptides from the venom of the social wasp Protopolybia exigua (saussure). Peptides 2006, 27, 2632–2639. [Google Scholar] [CrossRef] [PubMed]
- Dias, N.B.; de Souza, B.M.; Gomes, P.C.; Brigatte, P.; Palma, M.S. Peptidome profiling of venom from the social wasp Polybia paulista. Toxicon 2015, 107, 290–303. [Google Scholar] [CrossRef] [PubMed]
- Nakajima, T.; Yasuhara, T.; Horikawa, R.; Pisano, J.; Erspamer, V. A new structural class of biologically active peptide in non-mammals. In Advances in Experimental Medicine and Biology; Abe, K., Moriya, H., Fujii, S., Eds.; Springer: New York, NY, USA, 1989; pp. 215–220. [Google Scholar]
- De Souza, B.M.; da Silva, A.V.R.; Resende, V.M.F.; Arcuri, H.A.; dos Santos Cabrera, M.P.; Ruggiero Neto, J.; Palma, M.S. Characterization of two novel polyfunctional mastoparan peptides from the venom of the social wasp Polybia paulista. Peptides 2009, 30, 1387–1395. [Google Scholar] [CrossRef] [PubMed]
- Souza, B.M.; Mendes, M.A.; Santos, L.D.; Marques, M.R.; César, L.M.M.; Almeida, R.N.A.; Pagnocca, F.C.; Konno, K.; Palma, M.S. Structural and functional characterization of two novel peptide toxins isolated from the venom of the social wasp Polybia paulista. Peptides 2005, 26, 2157–2164. [Google Scholar] [CrossRef] [PubMed]
- Lee, V.S.; Tu, W.C.; Jinn, T.R.; Peng, C.C.; Lin, L.J.; Tzen, J.T. Molecular cloning of the precursor polypeptide of mastoparan B and its putative processing enzyme, dipeptidyl peptidase IV, from the black-bellied hornet, Vespa basalis. Insect Mol. Biol. 2007, 16, 231–237. [Google Scholar] [CrossRef] [PubMed]
- Xu, X.; Li, J.; Lu, Q.; Yang, H.; Zhang, Y.; Lai, R. Two families of antimicrobial peptides from wasp (Vespa magnifica) venom. Toxicon 2006, 47, 249–253. [Google Scholar] [CrossRef] [PubMed]
- Mendes, M.A.; de Souza, B.M.; Palma, M.S. Structural and biological characterization of three novel mastoparan peptides from the venom of the neotropical social wasp Protopolybia exigua (saussure). Toxicon 2005, 45, 101–106. [Google Scholar] [CrossRef] [PubMed]
- Lin, C.H.; Tzen, J.T.C.; Shyu, C.L.; Yang, M.J.; Tu, W.C. Structural and biological characterization of mastoparans in the venom of Vespa species in taiwan. Peptides 2011, 32, 2027–2036. [Google Scholar] [CrossRef] [PubMed]
- Ho, C.L.; Hwang, L.L. Structure and biological activities of a new mastoparan isolated from the venom of the hornet Vespa basalis. Biochem. J. 1991, 274, 453–456. [Google Scholar] [CrossRef] [PubMed]
- Chen, W.; Yang, X.; Yang, X.; Zhai, L.; Lu, Z.; Liu, J.; Yu, H. Antimicrobial peptides from the venoms of Vespa bicolor fabricius. Peptides 2008, 29, 1887–1892. [Google Scholar] [CrossRef] [PubMed]
- Yang, X.W.; Wang, Y.; Lee, W.H.; Zhang, Y. Antimicrobial peptides from the venom gland of the social wasp Vespa tropica. Toxicon 2013, 74, 151–157. [Google Scholar] [CrossRef] [PubMed]
- Baptista-Saidemberg, N.B.; Saidemberg, D.M.; Ribeiro, R.A.; Arcuri, H.A.; Palma, M.S.; Carneiro, E.M. Agelaia MP-I: A peptide isolated from the venom of the social wasp, Agelaia pallipes pallipes, enhances insulin secretion in mice pancreatic islets. Toxicon 2012, 60, 596–602. [Google Scholar] [CrossRef] [PubMed]
- Mendes, M.A.; de Souza, B.M.; Marques, M.R.; Palma, M.S. Structural and biological characterization of two novel peptides from the venom of the neotropical social wasp Agelaia pallipes pallipes. Toxicon 2004, 44, 67–74. [Google Scholar] [CrossRef] [PubMed]
- Piek, T.O.M.; Spanjer, W. 5—Chemistry and pharmacology of solitary wasp venoms. In Venoms of the Hymenoptera: Biochemical, Pharmacological and Behavioural Aspects; Piek, T., Ed.; Academic Press: London, UK, 1986; pp. 161–307. [Google Scholar]
- Yu, H.N.; Yang, H.L.; Ma, D.Y.; Lv, Y.; Liu, T.G.; Zhang, K.Y.; Lai, R.; Liu, J.Z. Vespid chemotactic peptide precursor from the wasp, Vespa magnifica (smith). Toxicon 2007, 50, 377–382. [Google Scholar] [CrossRef] [PubMed]
- Justo Jacomini, D.L.; Campos Pereira, F.D.; Aparecido dos Santos Pinto, J.R.; dos Santos, L.D.; da Silva Neto, A.J.; Giratto, D.T.; Palma, M.S.; de Lima Zollner, R.; Brochetto Braga, M.R. Hyaluronidase from the venom of the social wasp Polybia paulista (Hymenoptera, Vespidae): Cloning, structural modeling, purification, and immunological analysis. Toxicon 2013, 64, 70–80. [Google Scholar] [CrossRef] [PubMed]
- An, S.; Chen, L.; Wei, J.F.; Yang, X.; Ma, D.; Xu, X.; Xu, X.; He, S.; Lu, J.; Lai, R. Purification and characterization of two new allergens from the venom of Vespa magnifica. PLoS ONE 2012, 7. [Google Scholar] [CrossRef] [PubMed]
- King, T.P.; Spangfort, M.D. Structure and biology of stinging insect venom allergens. Int. Arch. Allergy Immunol. 2000, 123, 99–106. [Google Scholar] [CrossRef] [PubMed]
- Sukprasert, S.; Rungsa, P.; Uawonggul, N.; Incamnoi, P.; Thammasirirak, S.; Daduang, J.; Daduang, S. Purification and structural characterisation of phospholipase A1 (Vespapase, Ves a 1) from thai banded tiger wasp (Vespa affinis) venom. Toxicon 2013, 61, 151–164. [Google Scholar] [CrossRef] [PubMed]
- Hoffman, D.R. Allergens in hymenoptera venom 25: The amino-acid-sequences of antigen 5 molecules and the structural basis of antigenic cross-reactivity. J. Allergy Clin. Immunol. 1993, 92, 707–716. [Google Scholar] [CrossRef]
- Monsalve, R.I.; Lu, G.; King, T.P. Expressions of recombinant venom allergen, antigen 5 of yellowjacket (Vespula vulgaris) and paper wasp (Polistes annularis), in bacteria or yeast. Protein Expr. Purif. 1999, 16, 410–416. [Google Scholar] [CrossRef] [PubMed]
- Henriksen, A.; King, T.P.; Mirza, O.; Monsalve, R.I.; Meno, K.; Ipsen, H.; Larsen, J.N.; Gajhede, M.; Spangfort, M.D. Major venom allergen of yellow jackets, Ves v 5: Structural characterization of a pathogenesis-related protein superfamily. Proteins 2001, 45, 438–448. [Google Scholar] [CrossRef] [PubMed]
- King, T.P.; Moran, D.; Wang, D.F.; Kochoumian, L.; Chait, B.T. Structural studies of a hornet venom allergen antigen 5, Dol m V and its sequence similarity with other proteins. Protein Seq. Data Anal. 1990, 3, 263–266. [Google Scholar] [PubMed]
- Han, J.Y.; You, D.W.; Xu, X.Q.; Han, W.; Lu, Y.; Lai, R.; Meng, Q.X. An anticoagulant serine protease from the wasp venom of Vespa magnifica. Toxicon 2008, 51, 914–922. [Google Scholar] [CrossRef] [PubMed]
- Chen, L.; Chen, W.; Yang, H.; Lai, R. A novel bioactive peptide from wasp venom. J. Venom Res. 2010, 1, 43–47. [Google Scholar] [PubMed]
- Yoon, K.A.; Kim, K.; Nguyen, P.; Seo, J.B.; Park, Y.H.; Kim, K.-G.; Seo, H.-Y.; Koh, Y.H.; Lee, S.H. Comparative bioactivities of mastoparans from social hornets Vespa crabro and Vespa analis. J. Asia Pac. Entomol. 2015, 18, 825–829. [Google Scholar] [CrossRef]
- Baek, J.H.; Lee, S.H. Isolation and molecular cloning of venom peptides from Orancistrocerus drewseni (Hymenoptera: Eumenidae). Toxicon 2010, 55, 711–718. [Google Scholar] [CrossRef] [PubMed]
- Hsu, C.-C.; Huang, T.-F. Biological activities of snake venom metalloproteinases on platelets, neutrophils, endothelial cells, and extracellular matrices. In Toxins and Hemostasis: From Bench to Bedside; Kini, R.M., Clemetson, K.J., Markland, F.S., McLane, M.A., Morita, T., Eds.; Springer: Dordrecht, The Netherlands, 2011; pp. 723–732. [Google Scholar]
- Fernandez-Patron, C.; Leung, D. Emergence of a metalloproteinase/phospholipase A2 axis of systemic inflammation. Metalloproteinases Med. 2015, 2, 29–38. [Google Scholar] [CrossRef] [PubMed]
- Itino, T. Comparison of life tables between the solitary Eumenid wasp Antherhynchium flavomarginatum and the subsocial Eumenid wasp Orancistrocerus drewseni to evaluate the adaptive significance of maternal care. Res. Popul. Ecol. 1986, 28, 185–199. [Google Scholar] [CrossRef]
- Munoz-Torres, M.C.; Reese, J.T.; Childers, C.P.; Bennett, A.K.; Sundaram, J.P.; Childs, K.L.; Anzola, J.M.; Milshina, N.; Elsik, C.G. Hymenoptera genome database: Integrated community resources for insect species of the order Hymenoptera. Nucleic Acids Res. 2011, 39, D658–D662. [Google Scholar] [CrossRef] [PubMed]
- Heavner, M.E.; Gueguen, G.; Rajwani, R.; Pagan, P.E.; Small, C.; Govind, S. Partial venom gland transcriptome of a Drosophila parasitoid wasp, Leptopilina heterotoma, reveals novel and shared bioactive profiles with stinging Hymenoptera. Gene 2013, 526, 195–204. [Google Scholar] [CrossRef] [PubMed]
- Zhu, J.Y.; Fang, Q.; Wang, L.; Hu, C.; Ye, G.Y. Proteomic analysis of the venom from the endoparasitoid wasp Pteromalus puparum (Hymenoptera: Pteromalidae). Arch. Insect Biochem. 2010, 75, 28–44. [Google Scholar] [CrossRef] [PubMed]
- Kreil, G. Hyaluronidases—A group of neglected enzymes. Protein Sci. 1995, 4, 1666–1669. [Google Scholar] [CrossRef] [PubMed]
- Kemparaju, K.; Girish, K.S. Snake venom hyaluronidase: A therapeutic target. Cell Biochem. Funct. 2006, 24, 7–12. [Google Scholar] [CrossRef] [PubMed]
- Nagaraju, S.; Devaraja, S.; Kemparaju, K. Purification and properties of hyaluronidase from Hippasa partita (funnel web spider) venom gland extract. Toxicon 2007, 50, 383–393. [Google Scholar] [CrossRef] [PubMed]
- Girish, K.S.; Kemparaju, K. The magic glue hyaluronan and its eraser hyaluronidase: A biological overview. Life Sci. 2007, 80, 1921–1943. [Google Scholar] [CrossRef] [PubMed]
- Konno, K.; Miwa, A.; Takayama, H.; Hisada, M.; Itagaki, Y.; Naoki, H.; Yasuhara, T.; Kawai, N. Alpha-pompilidotoxin (α-pmtx), a novel neurotoxin from the venom of a solitary wasp, facilitates transmission in the crustacean neuromuscular synapse. Neurosci. Lett. 1997, 238, 99–102. [Google Scholar] [CrossRef]
- Sahara, Y.; Gotoh, M.; Konno, K.; Miwa, A.; Tsubokawa, H.; Robinson, H.P.; Kawai, N. A new class of neurotoxin from wasp venom slows inactivation of sodium current. Eur. J. Neurosci. 2000, 12, 1961–1970. [Google Scholar] [CrossRef] [PubMed]
- Hisada, M.; Satake, H.; Masuda, K.; Aoyama, M.; Murata, K.; Shinada, T.; Iwashita, T.; Ohfune, Y.; Nakajima, T. Molecular components and toxicity of the venom of the solitary wasp, Anoplius samariensis. Biochem. Biophys. Res. Commun. 2005, 330, 1048–1054. [Google Scholar] [CrossRef] [PubMed]
- Piek, T. Delta-philanthotoxin, a semi-irreversible blocker of ion-channels. Comp. Biochem. Phys. C 1982, 72, 311–315. [Google Scholar] [CrossRef]
- Vanmarle, J.; Piek, T.; Lind, A.; Vanweerenkramer, J. Inhibition of the glutamate uptake in the excitatory neuromuscular synapse of the locust by delta-philanthotoxin—A component of the venom of the solitary wasp Philanthus-triangulum f a high-resolution autoradiographic study. Comp. Biochem Phys. C 1984, 79, 213–215. [Google Scholar] [CrossRef]
- Piek, T.; Hue, B.; Pelhate, M.; David, J.A.; Spanjer, W.; Veldsemacurrie, R.D. Effects of the venom of Philanthus triangulum F. (Hym-Sphecidae) and beta-philanthotoxin and delta-philanthotoxin on axonal excitability and synaptic transmission in the cockroach CNS. Arch. Insect Biochem. 1984, 1, 297–306. [Google Scholar] [CrossRef]
- Moore, E.L.; Haspel, G.; Libersat, F.; Adams, M.E. Parasitoid wasp sting: A cocktail of GABA, taurine, and beta-alanine opens chloride channels for central synaptic block and transient paralysis of a cockroach host. J. Neurobiol. 2006, 66, 811–820. [Google Scholar] [CrossRef] [PubMed]
- Baek, J.H.; Lee, S.H.; Kim, W.-Y.; Kim, M.G. An insulin-binding protein from the venom of a solitary wasp Eumenes pomiformis binds to apolipophorin III in Lepidopteran hemolymph. Toxicon 2016, 111, 62–64. [Google Scholar] [CrossRef] [PubMed]
- Danneels, E.L.; Rivers, D.B.; de Graaf, D.C. Venom proteins of the parasitoid wasp Nasonia vitripennis: Recent discovery of an untapped pharmacopee. Toxins 2010, 2, 494–516. [Google Scholar] [CrossRef] [PubMed]
- Lee, K.M.; Lee, K.Y.; Choi, H.W.; Cho, M.Y.; Kwon, T.H.; Kawabata, S.; Lee, B.L. Activated phenoloxidase from tenebrio molitor larvae enhances the synthesis of melanin by using a vitellogenin-like protein in the presence of dopamine. Eur. J. Biochem. 2000, 267, 3695–3703. [Google Scholar] [CrossRef] [PubMed]
- Fang, K.S.Y.; Vitale, M.; Fehlner, P.; King, T.P. CDNA cloning and primary structure of a white-face hornet venom allergen, antigen-5. Proc. Natl. Acad. Sci. USA 1988, 85, 895–899. [Google Scholar] [CrossRef] [PubMed]
- Vincent, B.; Kaeslin, M.; Roth, T.; Heller, M.; Poulain, J.; Cousserans, F.; Schaller, J.; Poirie, M.; Lanzrein, B.; Drezen, J.M.; et al. The venom composition of the parasitic wasp Chelonus inanitus resolved by combined expressed sequence tags analysis and proteomic approach. BMC Genomics 2010, 11. [Google Scholar] [CrossRef] [PubMed]
- Hoffman, D.R. Hymenoptera venom allergens. Clin. Rev. Allergy Immunol. 2006, 30, 109–128. [Google Scholar] [CrossRef]
- Barboni, E.; Kemeny, D.M.; Campos, S.; Vernon, C.A. The purification of acid-phosphatase from honey-bee venom (Apis-mellifica). Toxicon 1987, 25, 1097–1103. [Google Scholar] [CrossRef]
- Kim, B.Y.; Jin, B.R. Molecular characterization of a venom acid phosphatase Acph-1-like protein from the Asiatic honeybee Apis cerana. J. Asia Pac. Entomol. 2014, 17, 695–700. [Google Scholar] [CrossRef]
- Hoffman, D.R. Allergens in bee venom: III. Identification of allergen-B of bee venom as an acid-phosphatase. J. Allergy Clin. Immunol. 1977, 59, 364–366. [Google Scholar] [CrossRef]
- De Graaf, D.C.; Aerts, M.; Brunain, M.; Desjardins, C.A.; Jacobs, F.J.; Werren, J.H.; Devreese, B. Insights into the venom composition of the ectoparasitoid wasp Nasonia vitripennis from bioinformatic and proteomic studies. Insect Mol. Biol. 2010, 19, 11–26. [Google Scholar] [CrossRef] [PubMed]
- Grunwald, T.; Bockisch, B.; Spillner, E.; Ring, J.; Bredehorst, R.; Ollert, M.W. Molecular cloning and expression in insect cells of honeybee venom allergen acid phosphatase (Api m 3). J. Allergy Clin. Immunol. 2006, 117, 848–854. [Google Scholar] [CrossRef] [PubMed]
- The Chemical Compositions of Insect Venoms. Available online: http://www.compoundchem.com/2014/08/28/insectvenoms/ (accessed on 18 December 2015).
- Yang, J.; Zhao, Y.; Pan, Y.; Lu, G.; Lu, L.; Wang, D.; Wang, J. Acetylcholine participates in pain modulation by influencing endogenous opiate peptides in rat spinal cord. World J. Neurosci. 2012, 2, 15–22. [Google Scholar] [CrossRef]
- Gorrell, M.D. Dipeptidyl peptidase iv and related enzymes in cell biology and liver disorders. Clin. Sci. (Lond.) 2005, 108, 277–292. [Google Scholar] [CrossRef] [PubMed]
- Eipper, B.A.; Milgram, S.L.; Husten, E.J.; Yun, H.Y.; Mains, R.E. Peptidylglycine alpha-amidating monooxygenase: A multifunctional protein with catalytic, processing, and routing domains. Protein Sci. 1993, 2, 489–497. [Google Scholar] [CrossRef] [PubMed]
- Kreil, G.; Kreil-Kiss, G. The isolation of N-formylglycine from a polypeptide present in bee venom. Biochem. Biophys. Res. Commun. 1967, 27, 275–280. [Google Scholar] [CrossRef]
- Schachter, M.; Thain, E.M. Chemical and pharmacological properties of the potent, slow contracting substance (kinin) in wasp venom. Br. J. Pharmacol. Chemother. 1954, 9, 352–359. [Google Scholar] [CrossRef] [PubMed]
- Nakajima, T. Pharmacological biochemistry of vespid venoms. In Venoms of the Hymenoptera: Biochemical, Pharmacological and Behavioural Aspects; Piek, T., Ed.; Academic: London, UK, 1986; pp. 309–327. [Google Scholar]
- Rocha, E.S.M.; Beraldo, W.T.; Rosenfeld, G. Bradykinin, a hypotensive and smooth muscle stimulating factor released from plasma globulin by snake venoms and by trypsin. Am. J. Physiol. 1949, 156, 261–273. [Google Scholar]
- Podvin, S.; Bundey, R.; Toneff, T.; Ziegler, M.; Hook, V. Profiles of secreted neuropeptides and catecholamines illustrate similarities and differences in response to stimulation by distinct secretagogues. Mol. Cell. Neurosci. 2015, 68, 177–185. [Google Scholar] [CrossRef] [PubMed]
- Piek, T.; Hue, B.; Pelhate, M.; Mony, L. The venom of the wasp Campsomeris sexmaculata (F.) blocks synaptic transmission in insect CNS. Comp. Biochem. Physiol. C 1987, 87, 283–286. [Google Scholar] [CrossRef]
- Piek, T.; Hue, B.; Mantel, P.; Nakajima, T.; Pelhate, M.; Yasuhara, T. Threonine6-bradykinin in the venom of the wasp Colpa interrupta (F.) presynaptically blocks nicotinic synaptic transmission in the insect CNS. Comp. Biochem. Physiol. C 1990, 96, 157–162. [Google Scholar] [CrossRef]
- Mortari, M.R.; Cunha, A.O.; Carolino, R.O.; Coutinho-Netto, J.; Tomaz, J.C.; Lopes, N.P.; Coimbra, N.C.; dos Santos, W.F. Inhibition of acute nociceptive responses in rats after i.c.v. Injection of Thr6-bradykinin, isolated from the venom of the social wasp, Polybia occidentalis. Br. J. Pharmacol. 2007, 151, 860–869. [Google Scholar] [CrossRef] [PubMed]
- Konno, K.; Hisada, M.; Naoki, H.; Itagaki, Y.; Yasuhara, T.; Juliano, M.A.; Juliano, L.; Palma, M.S.; Yamane, T.; Nakajima, T. Isolation and sequence determination of peptides in the venom of the spider wasp (Cyphononyx dorsalis) guided by matrix-assisted laser desorption/ionization time of flight (MALDI-TOF) mass spectrometry. Toxicon 2001, 39, 1257–1260. [Google Scholar] [CrossRef]
- Alsop, D.W.; Bettini, S. Arthropod Venoms; Springer-Verlag: Berlin, Germany; New York, NY, USA, 1978; p. 977. [Google Scholar]
- Piek, T.; Hue, B.; Lind, A.; Mantel, P.; van Marle, J.; Visser, J.H. The venom of ampulex compressa—Effects on behaviour and synaptic transmission of cockroaches. Comp. Biochem. Physiol. C 1989, 92, 175–183. [Google Scholar] [CrossRef]
- Hirai, Y.; Yasuhara, T.; Yoshida, H.; Nakajima, T.; Fujino, M.; Kitada, C. A new mast cell degranulating peptide “mastoparan” in the venom of Vespula lewisii. Chem. Pharm. Bull. 1979, 27, 1942–1944. [Google Scholar] [CrossRef] [PubMed]
- Brogden, K.A. Antimicrobial peptides: Pore formers or metabolic inhibitors in bacteria? Nat. Rev. Microbiol. 2005, 3, 238–250. [Google Scholar] [CrossRef] [PubMed]
- Higashijima, T.; Uzu, S.; Nakajima, T.; Ross, E.M. Mastoparan, a peptide toxin from wasp venom, mimics receptors by activating GTP-binding regulatory proteins (G proteins). J. Biol. Chem. 1988, 263, 6491–6494. [Google Scholar] [PubMed]
- Kurihara, H.; Kitajima, K.; Senda, T.; Fujita, H.; Nakajima, T. Multigranular exocytosis induced by phospholipase A2-activators, melittin and mastoparan, in rat anterior pituitary cells. Cell Tissue Res. 1986, 243, 311–316. [Google Scholar] [CrossRef] [PubMed]
- Baek, J.H.; Ji, Y.; Shin, J.S.; Lee, S.; Lee, S.H. Venom peptides from solitary hunting wasps induce feeding disorder in Lepidopteran larvae. Peptides 2011, 32, 568–572. [Google Scholar] [CrossRef] [PubMed]
- Moreno, M.; Giralt, E. Three valuable peptides from bee and wasp venoms for therapeutic and biotechnological use: Melittin, apamin and mastoparan. Toxins 2015, 7, 1126–1150. [Google Scholar] [CrossRef] [PubMed]
- Rocha, T.; de Souza, B.M.; Palma, M.S.; da Cruz-Hofling, M.A. Myotoxic effects of mastoparan from Polybia paulista (Hymenoptera, Epiponini) wasp venom in mice skeletal muscle. Toxicon 2007, 50, 589–599. [Google Scholar] [CrossRef] [PubMed]
- Silva, J.; Monge-Fuentes, V.; Gomes, F.; Lopes, K.; dos Anjos, L.; Campos, G.; Arenas, C.; Biolchi, A.; Goncalves, J.; Galante, P.; et al. Pharmacological alternatives for the treatment of neurodegenerative disorders: Wasp and bee venoms and their components as new neuroactive tools. Toxins 2015, 7, 3179–3209. [Google Scholar] [CrossRef] [PubMed]
- Moreno, M.; Zurita, E.; Giralt, E. Delivering wasp venom for cancer therapy. J. Control. Release 2014, 182, 13–21. [Google Scholar] [CrossRef] [PubMed]
- Kastin, A.J. Handbook of Biologically Active Peptides; Academic Press: Amsterdam, The Netherlands; Boston, MA, USA, 2006; p. 1595. [Google Scholar]
- Jindrichova, B.; Burketova, L.; Novotna, Z. Novel properties of antimicrobial peptide anoplin. Biochem. Biophys. Res. Commun. 2014, 444, 520–524. [Google Scholar] [CrossRef] [PubMed]
- Ifrah, D.; Doisy, X.; Ryge, T.S.; Hansen, P.R. Structure-activity relationship study of anoplin. J. Pept. Sci. 2005, 11, 113–121. [Google Scholar] [CrossRef] [PubMed]
- Spillner, E.; Blank, S.; Jakob, T. Hymenoptera allergens: From venom to “venome”. Front. Immunol. 2014, 5. [Google Scholar] [CrossRef] [PubMed]
- Boyle, R.J.; Elremeli, M.; Hockenhull, J.; Cherry, M.G.; Bulsara, M.K.; Daniels, M.; Elberink, J.N.G.O. Venom immunotherapy for preventing allergic reactions to insect stings. Cochrane Database Syst. Rev. 2012, 10. [Google Scholar] [CrossRef]
- Beckage, N.E.; Gelman, D.B. Wasp parasitoid disruption of host development: Implications for new biologically based strategies for insect control. Annu. Rev. Entomol. 2004, 49, 299–330. [Google Scholar] [CrossRef] [PubMed]
- Rydlo, T.; Miltz, J.; Mor, A. Eukaryotic antimicrobial peptides: Promises and premises in food safety. J. Food Sci. 2006, 71, R125–R135. [Google Scholar] [CrossRef]
- Bulet, P.; Stocklin, R.; Menin, L. Anti-microbial peptides: From invertebrates to vertebrates. Immunol. Rev. 2004, 198, 169–184. [Google Scholar] [CrossRef] [PubMed]
- Reddy, K.V.R.; Yedery, R.D.; Aranha, C. Antimicrobial peptides: Premises and promises. Int. J. Antimicrob. Agents 2004, 24, 536–547. [Google Scholar] [CrossRef] [PubMed]
- Vasilev, K.; Cook, J.; Griesser, H.J. Antibacterial surfaces for biomedical devices. Expert Rev. Med. Devices 2009, 6, 553–567. [Google Scholar] [CrossRef] [PubMed]
- Basu, A.; Mishra, B.; Leong, S.S.J. Immobilization of polybia-mpi by allyl glycidyl ether based brush chemistry to generate a novel antimicrobial surface. J. Mater. Chem. B 2013, 1, 4746–4755. [Google Scholar] [CrossRef]
- Jones, S.; Howl, J. Charge delocalisation and the design of novel mastoparan analogues: Enhanced cytotoxicity and secretory efficacy of [Lys5, Lys8, Aib10]MP. Regul. Pept. 2004, 121, 121–128. [Google Scholar] [CrossRef] [PubMed]
- Jones, S.; Martel, C.; Belzacq-Casagrande, A.S.; Brenner, C.; Howl, J. Mitoparan and target-selective chimeric analogues: Membrane translocation and intracellular redistribution induces mitochondrial apoptosis. Biochim. Biophys. Acta 2008, 1783, 849–863. [Google Scholar] [CrossRef] [PubMed]
- Wang, K.R.; Zhang, B.Z.; Zhang, W.; Yan, J.X.; Li, J.; Wang, R. Antitumor effects, cell selectivity and structure-activity relationship of a novel antimicrobial peptide Polybia-MPI. Peptides 2008, 29, 963–968. [Google Scholar] [CrossRef] [PubMed]
- Wang, K.R.; Yan, J.X.; Zhang, B.Z.; Song, J.J.; Jia, P.F.; Wang, R. Novel mode of action of Polybia-MPI, a novel antimicrobial peptide, in multi-drug resistant leukemic cells. Cancer Lett. 2009, 278, 65–72. [Google Scholar] [CrossRef] [PubMed]
- Leite, N.B.; Aufderhorst-Roberts, A.; Palma, M.S.; Connell, S.D.; Neto, J.R.; Beales, P.A. PE and PS lipids synergistically enhance membrane poration by a peptide with anticancer properties. Biophys. J. 2015, 109, 936–947. [Google Scholar] [CrossRef] [PubMed]
- Aalberse, R.C.; Akkerdaas, J.H.; van Ree, R. Cross-reactivity of ige antibodies to allergens. Allergy 2001, 56, 478–490. [Google Scholar] [CrossRef] [PubMed]
- Muller, U.R. Recombinant Hymenoptera venom allergens. Allergy 2002, 57, 570–576. [Google Scholar] [CrossRef] [PubMed]
- Muller, U.; Fricker, M.; Wymann, D.; Blaser, K.; Crameri, R. Increased specificity of diagnostic tests with recombinant major bee venom allergen phospholipase A2. Clin. Exp. Allergy 1997, 27, 915–920. [Google Scholar] [CrossRef] [PubMed]
- Magloire, V.; Czarnecki, A.; Anwander, H.; Streit, J. Beta-pompilidotoxin modulates spontaneous activity and persistent sodium currents in spinal networks. Neuroscience 2011, 172, 129–138. [Google Scholar] [CrossRef] [PubMed]
- Quistad, G.B.; Nguyen, Q.; Bernasconi, P.; Leisy, D.J. Purification and characterization of insecticidal toxins from venom glands of the parasitic wasp, Bracon hebetor. Insect Biochem. Mol. 1994, 24, 955–961. [Google Scholar] [CrossRef]
- Smith, J.J.; Herzig, V.; King, G.F.; Alewood, P.F. The insecticidal potential of venom peptides. Cell. Mol. Life Sci. 2013, 70, 3665–3693. [Google Scholar] [CrossRef] [PubMed]
- Down, R.E.; Fitches, E.C.; Wiles, D.P.; Corti, P.; Bell, H.A.; Gatehouse, J.A.; Edwards, J.P. Insecticidal spider venom toxin fused to snowdrop lectin is toxic to the peach-potato aphid, Myzus persicae (Hemiptera: Aphididae) and the rice brown planthopper, Nilaparvata lugens (Hemiptera : Delphacidae). Pest. Manag. Sci. 2006, 62, 77–85. [Google Scholar] [CrossRef] [PubMed]
- Fitches, E.C.; Pyati, P.; King, G.F.; Gatehouse, J.A. Fusion to snowdrop lectin magnifies the oral activity of insecticidal omega-hexatoxin-Hv1a peptide by enabling its delivery to the central nervous system. PLoS ONE 2012, 7. [Google Scholar] [CrossRef]
- Nakasu, E.Y.T.; Williamson, S.M.; Edwards, M.G.; Fitches, E.C.; Gatehouse, J.A.; Wright, G.A.; Gatehouse, A.M.R. Novel biopesticide based on a spider venom peptide shows no adverse effects on honeybees. Proc. Biol. Sci. 2014, 281. [Google Scholar] [CrossRef] [PubMed]
- Mccutchen, B.F.; Choudary, P.V.; Crenshaw, R.; Maddox, D.; Kamita, S.G.; Palekar, N.; Volrath, S.; Fowler, E.; Hammock, B.D.; Maeda, S. Development of a recombinant baculovirus expressing an insect-selective neurotoxin—Potential for pest-control. Biotechnology 1991, 9, 848–852. [Google Scholar] [CrossRef] [PubMed]
- Piek, T.; Hue, B.; Mony, L.; Nakajima, T.; Pelhate, M.; Yasuhara, T. Block of synaptic transmission in insect CNS by toxins from the venom of the wasp Megascolia flavifrons (Fab.). Comp. Biochem. Physiol. C 1987, 87, 287–295. [Google Scholar] [CrossRef]
- Kishimura, H.; Yasuhara, T.; Yoshida, H.; Nakajima, T. Vespakinin-M, a novel bradykinin analogue containing hydroxyproline, in the venom of Vespa mandarinia smith. Chem. Pharm. Bull. (Tokyo) 1976, 24, 2896–2897. [Google Scholar] [CrossRef] [PubMed]
- Yasuhara, T.; Yoshida, H.; Nakajima, T. Chemical investigation of the hornet (Vespa xanthoptera cameron) venom. The structure of a new bradykinin analogue “vespakinin-X”. Chem. Pharm. Bull. 1977, 25, 936–941. [Google Scholar] [CrossRef] [PubMed]
- Gobbo, M.; Biondi, L.; Filira, F.; Rocchi, R.; Piek, T. Cyclic analogues of wasp kinins from Vespa analis and Vespa tropica. Int. J. Pept. Protein Res. 1995, 45, 282–289. [Google Scholar] [CrossRef] [PubMed]
- Yoshida, H.; Pisano, J. Vespula kinins: New carbohydrate-containing bradykinin analogues. In Animal, Plant, and Microbial Toxins; Ohsaka, A., Hayashi, K., Sawai, Y., Murata, R., Funatsu, M., Tamiya, N., Eds.; Springer: New York, NY, USA, 1976; pp. 113–121. [Google Scholar]
- Konno, K.; Hisada, M.; Naoki, H.; Itagaki, Y.; Kawai, N.; Miwa, A.; Yasuhara, T.; Morimoto, Y.; Nakata, Y. Structure and biological activities of Eumenine mastoparan-AF (EMP-AF), a new mast cell degranulating peptide in the venom of the solitary wasp (Anterhynchium flavomarginatum micado). Toxicon 2000, 38, 1505–1515. [Google Scholar] [CrossRef]
- Dohtsu, K.; Okumura, K.; Hagiwara, K.; Palma, M.S.; Nakajima, T. Isolation and sequence analysis of peptides from the venom of Protonectarina sylveirae (Hymenoptera-Vespidae). Nat. Toxins 1993, 1, 271–276. [Google Scholar] [CrossRef] [PubMed]
- Baptista-Saidemberg, N.B.; Saidemberg, D.M.; de Souza, B.M.; Cesar-Tognoli, L.M.; Ferreira, V.M.; Mendes, M.A.; Cabrera, M.P.; Ruggiero Neto, J.; Palma, M.S. Protonectin (1–6): A novel chemotactic peptide from the venom of the social wasp Agelaia pallipes pallipes. Toxicon 2010, 56, 880–889. [Google Scholar] [CrossRef] [PubMed]
- Krishnakumari, V.; Nagaraj, R. Antimicrobial and hemolytic activities of crabrolin, a 13-residue peptide from the venom of the European hornet, Vespa crabro, and its analogs. J. Pept. Res. 1997, 50, 88–93. [Google Scholar] [CrossRef] [PubMed]
- Ho, C.L.; Chen, W.C.; Lin, Y.L. Structures and biological activities of new wasp venom peptides isolated from the black-bellied hornet (Vespa basalis) venom. Toxicon 1998, 36, 609–617. [Google Scholar] [CrossRef]
© 2016 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons by Attribution (CC-BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lee, S.H.; Baek, J.H.; Yoon, K.A. Differential Properties of Venom Peptides and Proteins in Solitary vs. Social Hunting Wasps. Toxins 2016, 8, 32. https://doi.org/10.3390/toxins8020032
Lee SH, Baek JH, Yoon KA. Differential Properties of Venom Peptides and Proteins in Solitary vs. Social Hunting Wasps. Toxins. 2016; 8(2):32. https://doi.org/10.3390/toxins8020032
Chicago/Turabian StyleLee, Si Hyeock, Ji Hyeong Baek, and Kyungjae Andrew Yoon. 2016. "Differential Properties of Venom Peptides and Proteins in Solitary vs. Social Hunting Wasps" Toxins 8, no. 2: 32. https://doi.org/10.3390/toxins8020032
APA StyleLee, S. H., Baek, J. H., & Yoon, K. A. (2016). Differential Properties of Venom Peptides and Proteins in Solitary vs. Social Hunting Wasps. Toxins, 8(2), 32. https://doi.org/10.3390/toxins8020032