
Proteomic Characterization and Comparison of Malaysian Tropidolaemus wagleri and Cryptelytrops purpureomaculatus Venom Using Shotgun-Proteomics

 ,
,  and
and
Abstract
:1. Introduction
2. Results and Discussion
2.1. The Venom Proteome of T. wagleri and C. purpureomaculatus
2.1.1. T. wagleri Venom
2.1.2. C. purpureomaculatus Venom
2.2. Venom Composition of T. wagleri and C. purpureomaculatus
2.2.1. Enzymatic Protein Families of T. wagleri and C. purpureomaculatus Venom
2.2.2. Nonenzymatic Protein Families of T. wagleri and C. purpureomaculatus
3. Conclusions
4. Materials and Methods
4.1. Materials
Snake Venom
4.2. Methods
4.2.1. In-Solution Tryptic Digestion
4.2.2. Nanoflow Liquid Chromatography Electrospray-Ionization Coupled with Tandem Mass Spectrometry (Nanoflow–ESI–LC–MS/MS)
4.2.3. Venom Protein Identification by Automated de Novo Sequencing (PEAKS Studio 7.0)
Supplementary Materials
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Das, I.; Ahmed, N.; Liat, L.B. Venomous terrestrial snakes of Malaysia: Their identity and biology. In Clinical Toxinology; Gopalakrishnakone, P., Faiz, S.M.A., Gnanathasan, A.C., Habib, G.A., Fernando, R., Yang, C.-C., Eds.; Malaysian Academic Library Institutional Repository: Kuala Lumpur, Malaysia, 2013; pp. 1–15. [Google Scholar]
- Vogel, G.; David, P.; Lutz, M.; Van Rooijen, J.; Vidal, N. Revision of the Tropidolaemus wagleri-complex (serpentes: Viperidae: Crotalinae). I. Definition of included taxa and redescription of Tropidolaemus wagleri (Boie, 1827). Zootaxa 2007, 1644, 1–40. [Google Scholar]
- Castoe, T.A.; Parkinson, C.L. Bayesian mixed models and the phylogeny of pitvipers (Viperidae: Serpentes). Mol. Phylogenet. Evol. 2006, 39, 91–110. [Google Scholar] [CrossRef] [PubMed]
- Tan, N.H.; Tan, C.S. The enzymatic activities and lethal toxins of Trimeresurus wagleri (speckled pit viper) venom. Toxicon 1989, 27, 349–357. [Google Scholar] [PubMed]
- Weinstein, S.A.; Schmidt, J.J.; Bernheimer, A.W.; Smith, L.A. Characterization and amino acid sequences of two lethal peptides isolated from venom of wagler’s pit viper, Trimeresurus wagleri. Toxicon 1991, 29, 227–236. [Google Scholar] [CrossRef]
- Molles, B.E.; Tsigelny, I.; Nguyen, P.D.; Gao, S.X.; Sine, S.M.; Taylor, P. Residues in the epsilon subunit of the nicotinic acetylcholine receptor interact to confer selectivity of waglerin-1 for the alpha-epsilon subunit interface site. Biochemistry 2002, 41, 7895–7906. [Google Scholar] [CrossRef] [PubMed]
- Molles, B.E.; Rezai, P.; Kline, E.F.; McArdle, J.J.; Sine, S.M.; Taylor, P. Identification of residues at the alpha and epsilon subunit interfaces mediating species selectivity of waglerin-1 for nicotinic acetylcholine receptors. J. Biol. Chem. 2002, 277, 5433–5440. [Google Scholar] [CrossRef] [PubMed]
- Utkin, Y.N. Three-finger toxins, a deadly weapon of elapid venom—Milestones of discovery. Toxicon 2013, 62, 50–55. [Google Scholar] [CrossRef] [PubMed]
- Das, I. A Naturalist’s Guide to the Snakes of Southeast Asia; John Beaufoy Publishing: Oxford, UK, 2012. [Google Scholar]
- David, P.; Vogel, G. Snakes of Sumatra: Annotated Checklist and Key with Natural History Notes; Bucher Kreth: Frankfurt, Germany, 1996. [Google Scholar]
- Mong, R.; Tan, H.H. Snakebite by the shore pit viper (Trimeresurus purpureomaculatus) treated with polyvalent antivenom. Wilderness Environ. Med. 2016, 27, 266–270. [Google Scholar] [CrossRef] [PubMed]
- Tan, N.H.; Armugam, A.; Tan, C.S. A comparative study of the enzymatic and toxic properties of venoms of the Asian lance-headed pit viper (genus Trimeresurus). Comp. Biochem. Phys. Part B 1989, 93, 757–762. [Google Scholar] [CrossRef]
- Gutierrez, J.M.; Lomonte, B.; Leon, G.; Alape-Giron, A.; Flores-Diaz, M.; Sanz, L.; Angulo, Y.; Calvete, J.J. Snake venomics and antivenomics: Proteomic tools in the design and control of antivenoms for the treatment of snakebite envenoming. J. Proteom. 2009, 72, 165–182. [Google Scholar] [CrossRef] [PubMed]
- UniProt. Available online: http://www.uniprot.org (accessed on 13 October 2016).
- Marcotte, E.M. How do shotgun proteomics algorithms identify proteins? Nat. Biotechnol. 2007, 25, 755–757. [Google Scholar] [CrossRef] [PubMed]
- Wu, C.C.; MacCoss, M.J. Shotgun proteomics: Tools for the analysis of complex biological systems. Curr. Opin. Mol. Ther. 2002, 4, 242–250. [Google Scholar] [PubMed]
- Rusmili, M.R.; Yee, T.T.; Mustafa, M.R.; Hodgson, W.C.; Othman, I. Proteomic characterization and comparison of malaysian Bungarus candidus and Bungarus fasciatus venoms. J. Proteom. 2014, 110, 129–144. [Google Scholar] [CrossRef] [PubMed]
- Malih, I.; Ahmad Rusmili, M.R.; Tee, T.Y.; Saile, R.; Ghalim, N.; Othman, I. Proteomic analysis of moroccan cobra Naja haje legionis venom using tandem mass spectrometry. J. Proteom. 2014, 96, 240–252. [Google Scholar] [CrossRef] [PubMed]
- Tan, N.H.; Tan, C.S. Biological properties of Trimeresurus purpureomaculatus (shore pit viper) venom and its fractions. Toxicon 1988, 26, 989–996. [Google Scholar] [CrossRef]
- Khow, O.; Chanhome, L.; Omori-Satoh, T.; Puempunpanich, S.; Sitprija, V. A hemorrhagin as a metalloprotease in the venom of Trimeresurus purpureomaculatus: Purification and characterization. Toxicon 2002, 40, 455–461. [Google Scholar] [CrossRef]
- Kamiguti, A.S.; Hay, C.R.; Theakston, R.D.; Zuzel, M. Insights into the mechanism of haemorrhage caused by snake venom metalloproteinases. Toxicon 1996, 34, 627–642. [Google Scholar] [CrossRef]
- Estevao-Costa, M.I.; Diniz, C.R.; Magalhaes, A.; Markland, F.S.; Sanchez, E.F. Action of metalloproteinases mutalysin I and II on several components of the hemostatic and fibrinolytic systems. Thromb. Res. 2000, 99, 363–376. [Google Scholar] [CrossRef]
- Loria, G.D.; Rucavado, A.; Kamiguti, A.S.; Theakston, R.D.; Fox, J.W.; Alape, A.; Gutierrez, J.M. Characterization of ‘basparin A’, a prothrombin-activating metalloproteinase, from the venom of the snake Bothrops asper that inhibits platelet aggregation and induces defibrination and thrombosis. Arch. Biochem. Biophys. 2003, 418, 13–24. [Google Scholar] [CrossRef]
- Takeda, S.; Takeya, H.; Iwanaga, S. Snake venom metalloproteinases: Structure, function and relevance to the mammalian ADAM/ADAMTS family proteins. BBA Proteins Proteom. 2012, 1824, 164–176. [Google Scholar] [CrossRef] [PubMed]
- Jeon, O.-H.; Kim, D.-S. Molecular cloning and functional characterization of a snake venom metalloprotease. Eur. J. Biochem. 1999, 263, 526–533. [Google Scholar] [CrossRef] [PubMed]
- Chen, R.Q.; Jin, Y.; Wu, J.B.; Zhou, X.D.; Lu, Q.M.; Wang, W.Y.; Xiong, Y.L. A new protein structure of P-II class snake venom metalloproteinases: It comprises metalloproteinase and disintegrin domains. Biochem. Biophys. Res. Commun. 2003, 310, 182–187. [Google Scholar] [CrossRef] [PubMed]
- Nikai, T.; Taniguchi, K.; Komori, Y.; Masuda, K.; Fox, J.W.; Sugihara, H. Primary structure and functional characterization of bilitoxin-1, a novel dimeric P-II snake venom metalloproteinase from Agkistrodon bilineatus venom. Arch. Biochem. Biophys. 2000, 378, 6–15. [Google Scholar] [CrossRef] [PubMed]
- Masuda, S.; Hayashi, H.; Araki, S. Two vascular apoptosis-inducing proteins from snake venom are members of the metalloprotease/disintegrin family. Eur. J. Biochem. 1998, 253, 36–41. [Google Scholar] [CrossRef] [PubMed]
- Masuda, S.; Ohta, T.; Kaji, K.; Fox, J.W.; Hayashi, H.; Araki, S. cDNA cloning and characterization of vascular apoptosis-inducing protein 1. Biochem. Biophys. Res. Commun. 2000, 278, 197–204. [Google Scholar] [CrossRef] [PubMed]
- Masuda, S.; Hayashi, H.; Atoda, H.; Morita, T.; Araki, S. Purification, cDNA cloning and characterization of the vascular apoptosis-inducing protein, HV1, from Trimeresurus flavoviridis. Eur. J. Biochem. 2001, 268, 3339–3345. [Google Scholar] [CrossRef] [PubMed]
- Tan, N.H.; Choy, S.K.; Chin, K.M.; Ponnudurai, G. Cross-reactivity of monovalent and polyvalent Trimeresurus antivenoms with venoms from various species of Trimeresurus (lance-headed pit viper) snake. Toxicon 1994, 32, 849–853. [Google Scholar] [CrossRef]
- Huang, H.W.; Liu, B.S.; Chien, K.Y.; Chiang, L.C.; Huang, S.Y.; Sung, W.C.; Wu, W.G. Cobra venom proteome and glycome determined from individual snakes of Naja atra reveal medically important dynamic range and systematic geographic variation. J. Proteom. 2015, 128, 92–104. [Google Scholar] [CrossRef] [PubMed]
- Gutierrez, J.M.; Sanz, L.; Flores-Diaz, M.; Figueroa, L.; Madrigal, M.; Herrera, M.; Villalta, M.; Leon, G.; Estrada, R.; Borges, A.; et al. Impact of regional variation in Bothrops asper snake venom on the design of antivenoms: Integrating antivenomics and neutralization approaches. J. Proteome Res. 2010, 9, 564–577. [Google Scholar] [CrossRef] [PubMed]
- Ponnudurai, G.; Chung, M.C.; Tan, N.H. Purification and properties of the L-amino acid oxidase from malayan pit viper (Calloselasma rhodostoma) venom. Arch. Biochem. Biophys. 1994, 313, 373–378. [Google Scholar] [CrossRef] [PubMed]
- Stabeli, R.G.; Marcussi, S.; Carlos, G.B.; Pietro, R.C.; Selistre-de-Araujo, H.S.; Giglio, J.R.; Oliveira, E.B.; Soares, A.M. Platelet aggregation and antibacterial effects of an L-amino acid oxidase purified from Bothrops alternatus snake venom. Bioorg. Med. Chem. 2004, 12, 2881–2886. [Google Scholar] [CrossRef] [PubMed]
- Ali, S.A.; Stoeva, S.; Abbasi, A.; Alam, J.M.; Kayed, R.; Faigle, M.; Neumeister, B.; Voelter, W. Isolation, structural, and functional characterization of an apoptosis-inducing L-amino acid oxidase from leaf-nosed viper (Eristocophis macmahoni) snake venom. Arch. Biochem. Biophys. 2000, 384, 216–226. [Google Scholar] [CrossRef] [PubMed]
- Zhong, S.R.; Jin, Y.; Wu, J.B.; Jia, Y.H.; Xu, G.L.; Wang, G.C.; Xiong, Y.L.; Lu, Q.M. Purification and characterization of a new L-amino acid oxidase from Daboia russellii siamensis venom. Toxicon 2009, 54, 763–771. [Google Scholar] [CrossRef] [PubMed]
- Rodrigues, R.S.; da Silva, J.F.; Boldrini Franca, J.; Fonseca, F.P.; Otaviano, A.R.; Henrique Silva, F.; Hamaguchi, A.; Magro, A.J.; Braz, A.S.; dos Santos, J.I.; et al. Structural and functional properties of BP-LAAO, a new L-amino acid oxidase isolated from Bothrops pauloensis snake venom. Biochimie 2009, 91, 490–501. [Google Scholar] [CrossRef] [PubMed]
- Rosenberg, P. The relationship between enzymatic activity and pharmacological properties of phospholipases in natural poisons. In Nature Toxins; Oxford University Press: Oxford, UK, 1986; pp. 129–174. [Google Scholar]
- Krizaj, I.; Siigur, J.; Samel, M.; Cotic, V.; Gubensek, F. Isolation, partial characterization and complete amino acid sequence of the toxic phospholipase A2 from the venom of the common viper, Vipera berus berus. Biochim. Biophys. Acta 1993, 1157, 81–85. [Google Scholar] [CrossRef]
- Kini, R.M. Excitement ahead: Structure, function and mechanism of snake venom phospholipase A2 enzymes. Toxicon 2003, 42, 827–840. [Google Scholar] [CrossRef] [PubMed]
- Tsai, I.H.; Chang, H.C.; Chen, J.M.; Cheng, A.C.; Khoo, K.H. Glycan structures and intrageneric variations of venom acidic phospholipases A2 from Tropidolaemus pitvipers. FEBS J. 2012, 279, 2672–2682. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.-M.; Huang, K.-F.; Tsai, I.-H. Snake venom glutaminyl cyclases: Purification, cloning, kinetic study, recombinant expression, and comparison with the human enzyme. Toxicon 2014, 86, 40–50. [Google Scholar] [CrossRef] [PubMed]
- Pawlak, J.; Manjunatha Kini, R. Snake venom glutaminyl cyclase. Toxicon 2006, 48, 278–286. [Google Scholar] [CrossRef] [PubMed]
- Aird, S.D. Ophidian envenomation strategies and the role of purines. Toxicon 2002, 40, 335–393. [Google Scholar] [CrossRef]
- Bernheimer, A.W.; Linder, R.; Weinstein, S.A.; Kim, K.S. Isolation and characterization of a phospholipase B from venom of collett’s snake, Pseudechis colletti. Toxicon 1987, 25, 547–554. [Google Scholar] [CrossRef]
- Longo, E.; Stamato, F.M.; Ferreira, R.; Tapia, O. The catalytic mechanism of serine proteases II: The effect of the protein environment in the alpha-chymotrypsin proton relay system. J. Theor. Biol. 1985, 112, 783–798. [Google Scholar] [CrossRef]
- Braud, S.; Bon, C.; Wisner, A. Snake venom proteins acting on hemostasis. Biochimie 2000, 82, 851–859. [Google Scholar] [CrossRef]
- Castro, H.C.; Zingali, R.B.; Albuquerque, M.G.; Pujol-Luz, M.; Rodrigues, C.R. Snake venom thrombin-like enzymes: From reptilase to now. Cell. Mol. Life Sci. 2004, 61, 843–856. [Google Scholar] [CrossRef] [PubMed]
- Tan, N.H. Isolation and characterization of the thrombin-like enzyme from Cryptelytrops purpureomaculatus venom. Comp. Biochem. Physiol. C Toxicol. Pharmacol. 2010, 151, 131–136. [Google Scholar] [CrossRef] [PubMed]
- Ouyang, C.; Huang, T.F. Alpha and beta-fibrinogenases from Trimeresurus gramineus snake venom. Biochim. Biophys. Acta 1979, 571, 270–283. [Google Scholar] [CrossRef]
- Ouyang, C.; Hwang, L.J.; Huang, T.F. Alpha-fibrinogenase from Agkistrodon rhodostoma (malayan pit viper) snake venom. Toxicon 1983, 21, 25–33. [Google Scholar] [CrossRef]
- Le Bonniec, B.F.; Libraire, J. Plasminogen activators from snake venoms. In Toxins and Hemostasis: From Bench to Bedside; Kini, M.R., Clemetson, J.K., Markland, S.F., McLane, A.M., Morita, T., Eds.; Springer: Dordrecht, The Netherlands, 2011; pp. 371–392. [Google Scholar]
- Chang, C.H.; Chung, C.H.; Kuo, H.L.; Hsu, C.C.; Huang, T.F. The highly specific platelet glycoprotein (GP) VI agonist trowaglerix impaired collagen-induced platelet aggregation ex vivo through matrix metalloproteinase-dependent GPVI shedding. J. Thromb. Haemost. 2008, 6, 669–676. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; Zheng, L.; Kong, C.; Kolatkar, P.R.; Chung, M.C. Purpureotin: A novel di-dimeric c-type lectin-like protein from Trimeresurus purpureomaculatus venom is stabilized by noncovalent interactions. Arch. Biochem. Biophys. 2004, 424, 53–62. [Google Scholar] [CrossRef] [PubMed]
- Ogawa, T.; Chijiwa, T.; Oda-Ueda, N.; Ohno, M. Molecular diversity and accelerated evolution of C-type lectin-like proteins from snake venom. Toxicon 2005, 45, 1–14. [Google Scholar] [CrossRef] [PubMed]
- Arlinghaus, F.T.; Eble, J.A. C-type lectin-like proteins from snake venoms. Toxicon 2012, 60, 512–519. [Google Scholar] [CrossRef] [PubMed]
- Schmidt, J.J.; Weinstein, S.A.; Smith, L.A. Molecular properties and structure-function relationships of lethal peptides from venom of Wagler’s pit viper, Trimeresurus wagleri. Toxicon 1992, 30, 1027–1036. [Google Scholar] [CrossRef]
- Molles, B.E.; Taylor, P. Structure and function of the waglerins, peptide toxins from the venom of Wagler’s pit viper, Tropidolaemus wagleri. J. Toxicol. Toxin Rev. 2002, 21, 273–292. [Google Scholar] [CrossRef]
- Mallick, P.; Kuster, B. Proteomics: A pragmatic perspective. Nat. Biotechnol. 2010, 28, 695–709. [Google Scholar] [CrossRef] [PubMed]
- Yamazaki, Y.; Morita, T. Structure and function of snake venom cysteine-rich secretory proteins. Toxicon 2004, 44, 227–231. [Google Scholar] [CrossRef] [PubMed]
- Yamazaki, Y.; Koike, H.; Sugiyama, Y.; Motoyoshi, K.; Wada, T.; Hishinuma, S.; Mita, M.; Morita, T. Cloning and characterization of novel snake venom proteins that block smooth muscle contraction. Eur. J. Biochem. 2002, 269, 2708–2715. [Google Scholar] [CrossRef] [PubMed]
- Yamazaki, Y.; Hyodo, F.; Morita, T. Wide distribution of cysteine-rich secretory proteins in snake venoms: Isolation and cloning of novel snake venom cysteine-rich secretory proteins. Arch. Biochem. Biophys. 2003, 412, 133–141. [Google Scholar] [CrossRef]
- Higuchi, S.; Murayama, N.; Saguchi, K.; Ohi, H.; Fujita, Y.; Camargo, A.C.M.; Ogawa, T.; Deshimaru, M.; Ohno, M. Bradykinin-potentiating peptides and C-type natriuretic peptides from snake venom. Immunopharmacology 1999, 44, 129–135. [Google Scholar] [CrossRef]
- Lameu, C.; Neiva, M.; Hayashi, M.A.F. Venom bradykinin-related peptides (BRPS) and its multiple biological roles. In An Integrated View of the Molecular Recognition and Toxinology—From Analytical Procedures to Biomedical Applications; Radis-Baptista, G., Ed.; InTech: Rijeka, Croatia, 2013. [Google Scholar]
- Vink, S.; Jin, A.H.; Poth, K.J.; Head, G.A.; Alewood, P.F. Natriuretic peptide drug leads from snake venom. Toxicon 2012, 59, 434–445. [Google Scholar] [CrossRef] [PubMed]
- Ondetti, M.A.; Rubin, B.; Cushman, D.W. Design of specific inhibitors of angiotensin-converting enzyme: New class of orally active antihypertensive agents. Science 1977, 196, 441–444. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Xin, L.; Shan, B.; Chen, W.; Xie, M.; Yuen, D.; Zhang, W.; Zhang, Z.; Lajoie, G.A.; Ma, B. Peaks db: De novo sequencing assisted database search for sensitive and accurate peptide identification. Mol. Cell. Proteom. 2012, 11. [Google Scholar] [CrossRef] [PubMed]


{kind=link}
{kind=link}
| Accession | −10lgP | Coverage (%) | #Peptides | #Unique | Description |
|---|---|---|---|---|---|
| P58930|WAG24_TROWA | 318.99 | 100 | 23 | 21 | Waglerin-4 (Tropidolaemus wagleri) |
| P24335|WAG13_TROWA | 306 | 100 | 20 | 18 | Waglerin-3 (Tropidolaemus wagleri) |
| C5H5D3|VM32_BOTAT | 247.11 | 16 | 6 | 2 | Zinc metalloproteinase-disintegrin-like batroxstatin-2 (Bothrops atrox) |
| P81382|OXLA_CALRH | 220.52 | 15 | 7 | 3 | l-amino-acid oxidase (Calloselasma rhodostoma) |
| F8S101|PLB_CROAD | 164.24 | 8 | 4 | 3 | Phospholipase B (Crotalus adamanteus) |
| P0C8J0|SLA_OPHHA | 161.12 | 85 | 4 | 4 | Snaclec ophioluxin subunit alpha (Fragment) (Ophiophagus hannah) |
| Q71RR4|SLA_TRIST | 78.55 | 17 | 2 | 2 | Snaclec coagulation factor IX/factor X-binding protein subunit A (Viridovipera stejnegeri) |
| P0DM38|SLDA_TRIAB | 65.57 | 20 | 2 | 2 | Snaclec alboaggregin-D subunit alpha (Cryptelytrops albolabris) |
| J3SBP3|PDE2_CROAD | 125.42 | 15 | 7 | 7 | Venom phosphodiesterase 2 (Crotalus adamanteus) |
| B6EWW8|V5NTD_GLOBR | 118.86 | 14 | 6 | 6 | Snake venom 5′-nucleotidase (Gloydius brevicaudus) |
| P00625|PA2A1_OVOOK | 111.35 | 16 | 4 | 3 | Acidic phospholipase A2 DE-I (Ovophis okinavensis) |
| P0DM51|PA2B1_BOTPA | 67.6 | 24 | 3 | 3 | Basic phospholipase A2 BnpTX-1 (Fragment) (Bothrops pauloensis) |
| O13069|VSP2_BOTJA | 92.72 | 21 | 4 | 3 | Thrombin-like enzyme KN-BJ 2 (Bothropoides jararaca) |
| Accession | −10lgp | Coverage (%) | #Peptides | #Unique | Description |
|---|---|---|---|---|---|
| P0C6E8|VM3G1_TRIGA | 322.01 | 38 | 29 | 5 | Zinc metalloproteinase/disintegrin (Fragment) (Trimeresurus gramineus) |
| P15503|VM2TA_TRIGA | 306.48 | 33 | 24 | 2 | Zinc metalloproteinase/disintegrin (Trimeresurus gramineus) |
| Q3HTN1|VM3SA_TRIST | 276.23 | 30 | 15 | 8 | Zinc metalloproteinase-disintegrin-like stejnihagin-A (Viridovipera stejnegeri) |
| P62383|VM2G_TRIGA | 270.15 | 85 | 6 | 2 | Disintegrin trigramin-gamma (Trimeresurus gramineus) |
| Q2LD49|VM3TM_TRIST | 255.16 | 23 | 15 | 6 | Zinc metalloproteinase-disintegrin-like TSV-DM (Viridovipera stejnegeri) |
| P0C6B6|VM2AL_TRIAB | 246.22 | 24 | 14 | 8 | Zinc metalloproteinase homolog-disintegrin albolatin (Cryptelytrops albolabris) |
| P0DM87|VM2_TRIST | 242.76 | 11 | 10 | 3 | Zinc metalloproteinase-disintegrin stejnitin (Viridovipera stejnegeri) |
| Q3HTN2|VM3SB_TRIST | 235.08 | 13 | 7 | 3 | Zinc metalloproteinase-disintegrin-like stejnihagin-B (Viridovipera stejnegeri) |
| Q8AWX7|VM2AG_GLOHA | 224.09 | 12 | 6 | 2 | Zinc metalloproteinase-disintegrin agkistin (Gloydius halys) |
| P20164|VM3HB_PROFL | 214.8 | 14 | 10 | 5 | Zinc metalloproteinase-disintegrin-like HR1b (Probothrops flavoviridis) |
| Q2UXQ5|VM3E2_ECHOC | 202.85 | 6 | 4 | 2 | Zinc metalloproteinase-disintegrin-like EoVMP2 (Echis ocellatus) |
| Q98UF9|VM3H3_BOTJA | 180.86 | 12 | 5 | 2 | Zinc metalloproteinase-disintegrin-like HF3 (Bothropoides jararaca) |
| Q0NZX6|VM2JR_BOTJA | 175.59 | 13 | 4 | 4 | Zinc metalloproteinase-disintegrin jararin (Fragment) (Bothropoides jararaca) |
| Q92043|VM3AA_CROAT | 149.13 | 14 | 7 | 5 | Zinc metalloproteinase-disintegrin-like atrolysin-A (Fragment) (Crotalus atrox) |
| P0DM90|VM32B_GLOBR | 122.17 | 5 | 3 | 2 | Zinc metalloproteinase-disintegrin-like brevilysin H2b (Gloydius brevicaudus) |
| Q7ZZS9|VM2J_PROJR | 118.04 | 14 | 6 | 2 | Zinc metalloproteinase/disintegrin (Trimeresurus jerdonii) |
| C5H5D4|VM33_BOTAT | 90.17 | 7 | 2 | 2 | Zinc metalloproteinase-disintegrin-like batroxstatin-3 (Fragment) (Bothrops atrox) |
| Q90495|VM3E_ECHCA | 55.28 | 2 | 2 | 2 | Zinc metalloproteinase-disintegrin-like ecarin (Echis carinatus) |
| P0CJ41|VSPAF_TRIAB | 304.68 | 42 | 13 | 8 | Alpha-fibrinogenase albofibrase (Cryptelytrops albolabris) |
| P0DJL2|SLA_CRYPP | 295.1 | 85 | 15 | 3 | Snaclec purpureotin subunit alpha (Cryptelytrops purpureomaculatus) |
| P0DJL3|SLB_CRYPP | 264.92 | 65 | 10 | 2 | Snaclec purpureotin subunit beta (Cryptelytrops purpureomaculatus) |
| P0DM38|SLDA_TRIAB | 257.69 | 49 | 9 | 6 | Snaclec alboaggregin-D subunit alpha (Cryptelytrops albolabris) |
| Q71RR4|SLA_TRIST | 256.32 | 58 | 10 | 8 | Snaclec coagulation factor IX/factor X-binding protein subunit A (Viridovipera stejnegeri) |
| P81115|SLBA_TRIAB | 256.21 | 61 | 11 | 3 | Snaclec alboaggregin-B subunit alpha (Cryptelytrops albolabris) |
| Q9YGP1|LECG_TRIST | 212.86 | 37 | 9 | 3 | C-type lectin TsL (Viridovipera stejnegeri) |
| Q71RQ1|SLAA_TRIST | 156.33 | 28 | 5 | 3 | Snaclec stejaggregin-A subunit alpha (Viridovipera stejnegeri) |
| Q71RQ0|SLAB1_TRIST | 115.42 | 20 | 3 | 3 | Snaclec stejaggregin-A subunit beta-1 (Viridovipera stejnegeri) |
| Q8JIV8|SL_DEIAC | 104.16 | 20 | 3 | 2 | Snaclec clone 2100755 (Deinagkistrodon acutus) |
| Q7T2Q0|SLB_ECHML | 100.26 | 22 | 3 | 2 | Snaclec EMS16 subunit beta (Echis multisquamatus) |
| Q6H3D7|PA2HH_TRIST | 254.23 | 52 | 14 | 8 | Basic phospholipase A2 homolog CTs-R6 (Viridovipera stejnegeri) |
| Q2YHJ5|PA2AB_TRIPE | 249.27 | 64 | 15 | 8 | Acidic phospholipase A2 Tpu-E6b (Trimeresurus puniceus) |
| Q6H3D6|PA2HD_TRIST | 233.17 | 59 | 13 | 7 | Basic phospholipase A2 homolog Ts-R6 (Viridovipera stejnegeri) |
| G3DT18|PA2A_BOTMO | 180.26 | 25 | 10 | 7 | Acidic phospholipase A2 BmooPLA2 (Bothrops moojeni) |
| A7LAC7|VSP2_TRIAB | 223.03 | 33 | 9 | 2 | Thrombin-like enzyme 2 (Cryptelytrops albolabris) |
| A7LAC6|VSP1_TRIAB | 200.97 | 37 | 8 | 4 | Thrombin-like enzyme 1 (Cryptelytrops albolabris) |
| Q8AY81|VSPST_TRIST | 172.15 | 23 | 6 | 2 | Thrombin-like enzyme stejnobin (Viridovipera stejnegeri) |
| J3SEZ3|PDE1_CROAD | 221.02 | 28 | 19 | 19 | Venom phosphodiesterase 1 (Crotalus adamanteus) |
| Q90W54|OXLA_GLOBL | 211.82 | 15 | 9 | 3 | l-amino-acid oxidase (Gloydius blomhoffii) |
| Q6WP39|OXLA_TRIST | 206.52 | 14 | 10 | 4 | l-amino-acid oxidase (Viridovipera stejnegeri) |
| Q6TGQ8|OXLA_BOTMO | 179.18 | 23 | 9 | 2 | l-amino-acid oxidase (Fragment) (Bothrops moojeni) |
| Q4F867|OXLA_DABSI | 178.26 | 13 | 6 | 2 | l-amino-acid oxidase (Fragments) (Daboia siamensis) |
| X2JCV5|OXLA_CERCE | 133.13 | 10 | 6 | 2 | l-amino acid oxidase (Cerastes cerastes) |
| P60623|CRVP_TRIST | 204.78 | 46 | 11 | 4 | Cysteine-rich venom protein (Fragment) (Viridovipera stejnegeri) |
| P81995|CRVP_CRYPP | 153.46 | 52 | 3 | 3 | Cysteine-rich venom protein tripurin (Fragment) (Cryptelytrops purpureomaculatus) |
| Q8JI39|CRVP_PROFL | 127.32 | 24 | 8 | 3 | Cysteine-rich venom protein triflin (Probothrops flavoviridis) |
| P0DJF5|VSPPA_TRIAB | 177.15 | 27 | 8 | 4 | Venom plasminogen activator GPV-PA (Cryptelytrops albolabris) |
| O13061|VSPB_TRIGA | 165.79 | 21 | 5 | 3 | Snake venom serine protease 2B (Trimeresurus gramineus) |
| Q90YA8|QPCT_GLOBL | 145.59 | 11 | 3 | 3 | Glutaminyl-peptide cyclotransferase (Gloydius blomhoffii) |
| F8S0Z7|V5NTD_CROAD | 106.69 | 17 | 6 | 6 | Snake venom 5′-nucleotidase (Crotalus adamanteus) |
| P0C7P6|BNP_TRIGA | 105.18 | 17 | 4 | 4 | Bradykinin-potentiating and C-type natriuretic peptides (Trimeresurus gramineus) |
| F8S101|PLB_CROAD | 89.21 | 8 | 3 | 3 | Phospholipase B (Crotalus adamanteus) |
| Enzymatic | Non-Enzymatic Protein |
|---|---|
| Snake venom metalloproteinase (SVMP) * | Snake venom C-type lectin (SNACLEC) |
| Phospholipase A2 (PLA2) | Waglerin |
| l-amino acid oxidase (LAAO) | - |
| Snake venom serine protease (SVSP) | - |
| Snake venom 5′-nucleotidase | - |
| Venom phosphodiesterase (vPDE) | - |
| Phospholipase B (PLB) | - |
| Enzymatic | Non-Enzymatic Protein |
|---|---|
| Snake venom metalloproteinase (SVMP) | Snake venom C-type lectin (SNACLEC) |
| Phospholipase A2 (PLA2) | Cysteine-rich venom protein (CRVP) |
| l-amino acid oxidase (LAAO) | Bradykinin-potentiating and C-type natriuretic peptide (BCNP) * |
| Snake venom serine protease (SVSP) | - |
| Glutaminyl-peptide cyclotransferase (QPCT) * | - |
| Snake venom 5′-nucleotidase | - |
| Venom phosphodiesterase (vPDE) | - |
| Phospholipase B (PLB) | - |
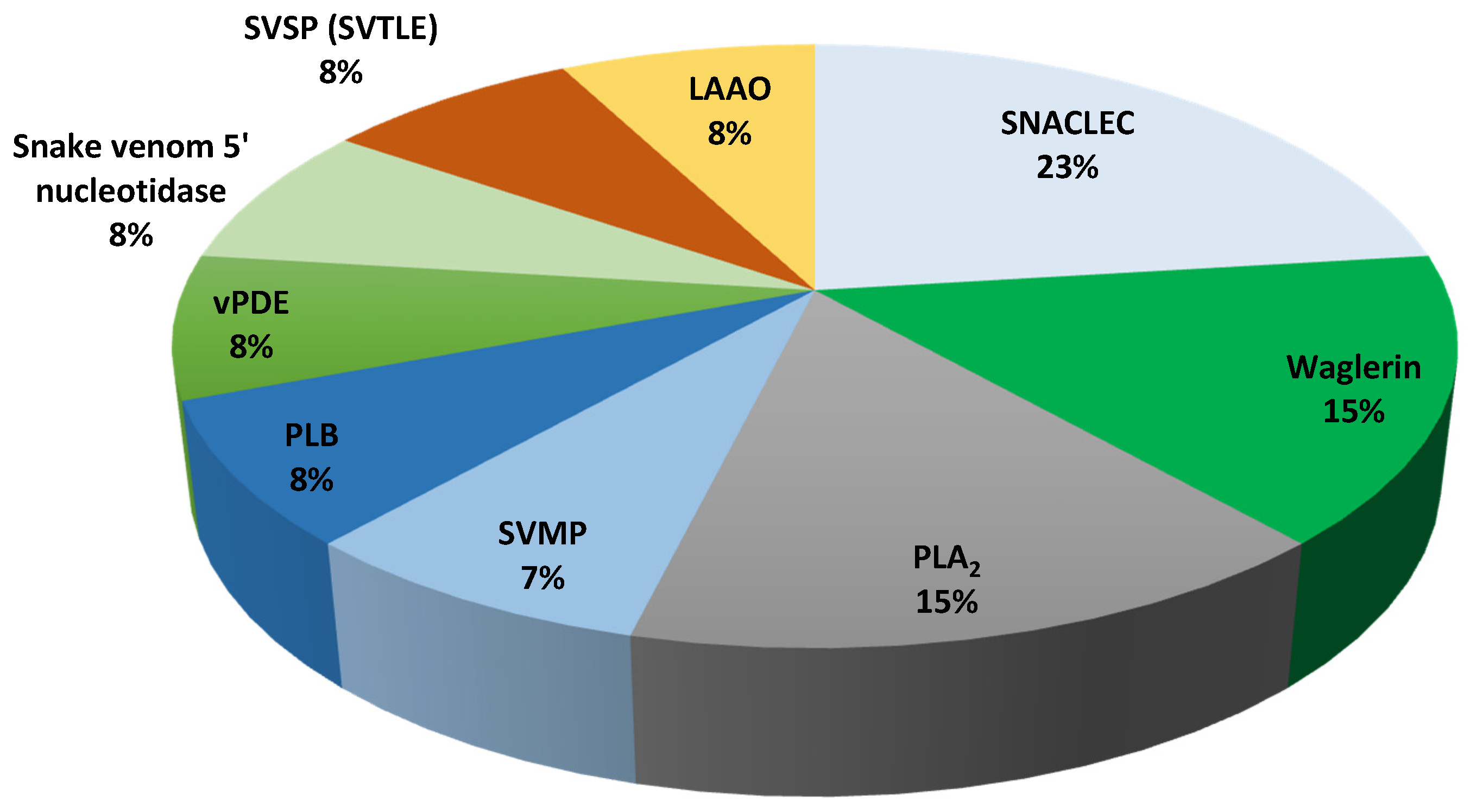
| Venom Protein Family | Tropidolaemus wagleri | Cryptelytrops purpureomaculatus |
|---|---|---|
| Snake venom C-type lectin (SNACLEC) | 23% | 19% |
| Waglerin | 15% | - |
| Phospholipase A2 (PLA2) | 15% | 8% |
| l-amino acid oxidase (LAAO) | 8% | 10% |
| Snake venom 5′-nucleotidase | 8% | 2% |
| Snake venom serine protease (SVSP) | 8% | 12% |
| Venom phosphodiesterase (vPDE) | 8% | 2% |
| Phospholipase B (PLB) | 8% | 2% |
| Snake venom metalloproteinase (SVMP) | 7% | 35% |
| Cysteine-rich venom protein (CRVP) | - | 6% |
| Bradykinin-potentiating and C-type natriuretic peptides (BCNP) | - | 2% |
| Glutaminyl-peptide cyclotransferase (QPCT) | - | 2% |
© 2016 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC-BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zainal Abidin, S.A.; Rajadurai, P.; Chowdhury, M.E.H.; Ahmad Rusmili, M.R.; Othman, I.; Naidu, R. Proteomic Characterization and Comparison of Malaysian Tropidolaemus wagleri and Cryptelytrops purpureomaculatus Venom Using Shotgun-Proteomics. Toxins 2016, 8, 299. https://doi.org/10.3390/toxins8100299
Zainal Abidin SA, Rajadurai P, Chowdhury MEH, Ahmad Rusmili MR, Othman I, Naidu R. Proteomic Characterization and Comparison of Malaysian Tropidolaemus wagleri and Cryptelytrops purpureomaculatus Venom Using Shotgun-Proteomics. Toxins. 2016; 8(10):299. https://doi.org/10.3390/toxins8100299
Chicago/Turabian StyleZainal Abidin, Syafiq Asnawi, Pathmanathan Rajadurai, Md Ezharul Hoque Chowdhury, Muhamad Rusdi Ahmad Rusmili, Iekhsan Othman, and Rakesh Naidu. 2016. "Proteomic Characterization and Comparison of Malaysian Tropidolaemus wagleri and Cryptelytrops purpureomaculatus Venom Using Shotgun-Proteomics" Toxins 8, no. 10: 299. https://doi.org/10.3390/toxins8100299
APA StyleZainal Abidin, S. A., Rajadurai, P., Chowdhury, M. E. H., Ahmad Rusmili, M. R., Othman, I., & Naidu, R. (2016). Proteomic Characterization and Comparison of Malaysian Tropidolaemus wagleri and Cryptelytrops purpureomaculatus Venom Using Shotgun-Proteomics. Toxins, 8(10), 299. https://doi.org/10.3390/toxins8100299