Abstract
Pufferfish poisoning has not been well documented in the South Pacific, although fish and other seafood are sources of protein in these island nations. In this study, tetrodotoxin (TTX) and its analogues in each organ of the pufferfish Arothron hispidus and A. nigropunctatus collected in the Solomon Islands were investigated using high resolution LC-MS. The toxin profiles of the same two species of pufferfish from Okinawa, Japan were also examined for comparison. TTXs concentrations were higher in the skin of both species from both regions, and relatively lower in the liver, ovary, testis, stomach, intestine, and flesh. Due to higher TTX concentrations (51.0 and 28.7 µg/g at highest) detected in the skin of the two species from the Solomon Islands (saxitoxin was <0.02 µg/g), these species should be banned from consumption. Similar results were obtained from fish collected in Okinawa, Japan: TTX in the skin of A. hispidus and A. nigropunctatus were 12.7 and 255 µg/g, respectively, at highest, and saxitoxin was also detected in the skin (2.80 µg/g at highest) and ovary of A. hispidus. TTX, 5,6,11-trideoxyTTX (with its 4-epi form), and its anhydro forms were the most abundant, and 11-oxoTTX was commonly detected in the skin.
1. Introduction
Tetrodotoxin (TTX) is a well-studied neurotoxin known for its distribution in pufferfish [1] and marine invertebrates such as snails [2], crabs [3], starfish [4], blue-ringed octopus [5], and sea slugs [6]. Recently, TTX was also detected in bivalve mollusks: New Zealand clams [7], mussels and oysters in England [8], and also in Greek shellfish [9]. Generally, TTX blocks the voltage-gated sodium ion channels, incapacitating nerve conduction and muscle action potentials, causing progressive paralysis and death due to failure of the respiratory system [10,11]. TTX, as the primary agent of pufferfish poisoning, is reported to be produced by marine bacteria and accumulated in the pufferfish via the food chain [12,13]. TTX in newts was reported to be an excellent defense strategy to ward off predators [14], and TTX is used to capture mobile prey by flatworms in Guam [15]. We have found various TTX analogues (Figure 1) which can be further classified into (1) hemilactal type analogues, (2) 5-deoxy-10,7-lactone type analogues, (3) 4,9- and 4,4a-anhydro type analogues and (4) tetrodonic acid type analogue [16,17]. Based on the structures of these analogues, we predicted a stepwise oxidative pathway of TTX in marine animals [17]. Additionally, we recently found C5–C10 directly bonded TTX analogues in newts, suggesting a monoterpene origin of TTX [18].
Toxicity experiments using mouse bioassays have established an LD50 (50% lethal dose, mice, i.p.) of TTX and few analogues. For instance, the LD50 of TTX [10], 11-deoxyTTX [19] and 6,11-dideoxyTTX [20] were 10 μg/kg, 70 μg/kg, and 420 μg/kg, respectively, while for 5,6,11-trideoxyTTX, LD99 was 750 μg/kg [21]. The toxicity of 11-oxoTTX was first reported as 120 µg/kg (minimum lethal dose, mice, i.p.) [22], but then re-examined on rat skeletal muscle fibers with an ED50 of 0.7 nM compared to TTX 4.1 nM [23], suggesting that 11-oxoTTX is more toxic than TTX. Furthermore, higher activity was also reflected in the smaller Kd values of 11-oxoTTX (to rat brain membrane) than TTX [24].
Past incidences of pufferfish food poisoning were reported in several countries including Thailand [25], Bangladesh [26], Hong Kong [27], Singapore [28], Japan [29], Suez Canal region [30], Eastern Mediterranean Sea [31], and several cases in the United States originating mainly from Asian dried pufferfish believed to be illegally imported [32,33]. Reports on these global occurrences and its potential expansion indicate that proper research on the specific toxic species and their distribution is of great importance. However, unlike ciguatera and other seafood poisoning, pufferfish poisoning is not well documented in the South Pacific region although several incidences matching its description have been reported in localized rural areas [34]. The Solomon Islands in particular does not have clear records of victims suffering from pufferfish poisoning. Past undocumented incidents may have triggered a precautionary knowledge passed down through successive generations of the potential dangers of pufferfish consumption. However, because fish and other seafood are a source of protein to the citizens of this island nation, and together with socioeconomic pressures that may affect food security and possibly alter food selectivity, we believe this investigation is imperative as well as informative to the citizens regarding potential risk of pufferfish poisoning.
Besides TTX, some pufferfish also accumulate paralytic shellfish toxins. The presence of saxitoxin (STX) and decarbamoyl STX (dcSTX) has been reported in pufferfish collected in United States [35], South-eastern Asia, and Japan [36,37,38].
In this study, the pufferfish Arothron hispidus and A. nigropunctatus collected in the Solomon Islands were investigated for TTX and its analogues, as well as STXs for the first time, using high resolution hydrophilic interaction chromatography (HR-HILIC) LC-MS. The toxin profiles of the same two species from Okinawa, Japan, were also examined and compared with those from the Solomon Islands. In addition, two specimens of Diodon holocanthus from the Solomon Islands, were also tested for the presence of TTXs, because the skin, flesh, and testis of D. holocanthus are officially allowed for consumption in Japan [39].
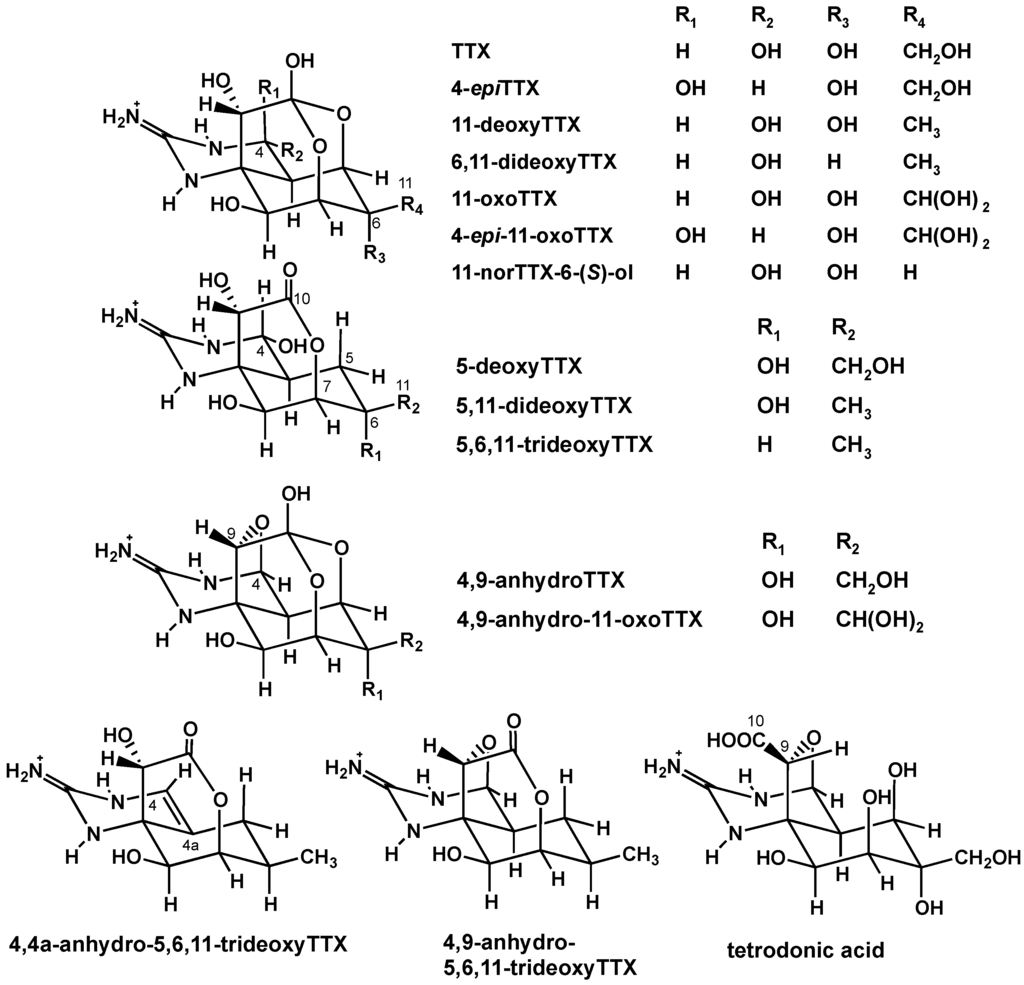
Figure 1.
The structures of TTX and its analogues.
Figure 1.
The structures of TTX and its analogues.
2. Results
2.1. Analysis of TTX and Its Analogues in the Pufferfish from the Solomon Islands
2.1.1. Preliminary Analysis of TTX and Its Analogues in the Skin of A. hispidus from the Solomon Islands using LC-FLD
Transportation of pufferfish samples from Solomon Islands to Japan under frozen condition was not possible. Therefore, immediately after collection, the pufferfish specimens were dissected into organs and soaked in ethanol-water (7:3, v/v) for shipping to Tohoku University (Sendai, Japan) at room temperature. We first examined whether TTXs in the pufferfish samples were decomposed during the transportation period of approximately two weeks. Under such conditions, certain TTX and its analogues might be hydrolyzed to tetrodonic acid (Figure 1) type compounds [16,40], which are detectable using liquid chromatography-fluorescent detection (LC-FLD) for TTXs [41,42] without specific pre-purification. TTXs in the ethanolic solution used to soak the skin of A. hispidus (three specimens), and in the skin soaked in this ethanolic solution were both analyzed by LC-FLD. The LC-FLD chromatograms thus obtained in the skin of the A. hispidus No.3 specimen are shown in Figure 2. The peak area of tetrodonic acid type analogues (peak A) was approximately 2/3 of TTX (peak C) in both the ethanolic solution (Figure 2b) and the extract of the skin (Figure 2c). The peaks of 11-oxoTTX and 4-epi-11-oxoTTX (peak B, not separated), 4,9-anhydro-11-oxoTTX (peak D), 4-epiTTX (peak E) and 4,9-anhydroTTX (peak F) are clearly observed. These data suggest that the pufferfish samples transported from the Solomon Islands are still suitable for analysis of TTXs, even though TTXs were partially transformed to tetrodonic acid type analogues.
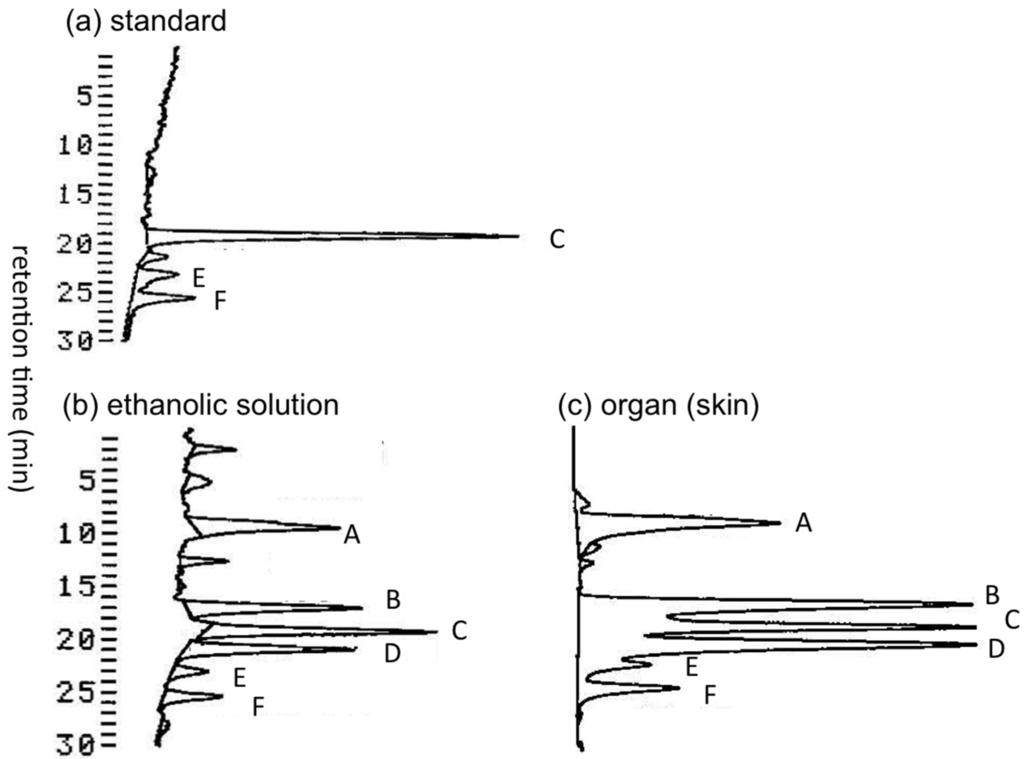
Figure 2.
The LC-FLD chromatograms of the skin extracts of the A. hispidus No.3 specimen from the Solomon Islands. (a) The TTXs standard solution (5 µL) containing TTX 1.0 µg/mL, 4-epiTTX 0.18 µg/mL, and 4,9-anhydroTTX 0.24 µg/mL (attenuation 3), (b) Aliquot sample (6 µL) prepared from the ethanol-water (7:3, v/v) solution used to soak the skin of A. hispidus No.3 specimen (10.5 mL/g) (attenuation 6), (c) The sample solution (2 µL) prepared from 0.2 M acetic acid (v/v) extract of the skin of A. hispidus No.3 specimen soaked in ethanolic solution (5.0 mL/g) (attenuation 6) (see Section 4.3). A: tetrodonic acid types, B: 11-oxoTTX and 4-epi-11-oxoTTX, C: TTX, D: 4,9-anhydro-11-oxoTTX, E: 4-epiTTX, F: 4,9-anhydroTTX. Chromatographic condition: Develosil C30 UG-5 (0.46 x25 cm) column; 30 mM ammonium heptafluorobutyrate buffer (pH 5.0) and 10 mM ammonium formate buffer (pH 5.0) containing 1% (v/v) acetonitrile as the solvent, flow rate 0.4 mL/min (see Section 4.5).
Figure 2.
The LC-FLD chromatograms of the skin extracts of the A. hispidus No.3 specimen from the Solomon Islands. (a) The TTXs standard solution (5 µL) containing TTX 1.0 µg/mL, 4-epiTTX 0.18 µg/mL, and 4,9-anhydroTTX 0.24 µg/mL (attenuation 3), (b) Aliquot sample (6 µL) prepared from the ethanol-water (7:3, v/v) solution used to soak the skin of A. hispidus No.3 specimen (10.5 mL/g) (attenuation 6), (c) The sample solution (2 µL) prepared from 0.2 M acetic acid (v/v) extract of the skin of A. hispidus No.3 specimen soaked in ethanolic solution (5.0 mL/g) (attenuation 6) (see Section 4.3). A: tetrodonic acid types, B: 11-oxoTTX and 4-epi-11-oxoTTX, C: TTX, D: 4,9-anhydro-11-oxoTTX, E: 4-epiTTX, F: 4,9-anhydroTTX. Chromatographic condition: Develosil C30 UG-5 (0.46 x25 cm) column; 30 mM ammonium heptafluorobutyrate buffer (pH 5.0) and 10 mM ammonium formate buffer (pH 5.0) containing 1% (v/v) acetonitrile as the solvent, flow rate 0.4 mL/min (see Section 4.5).
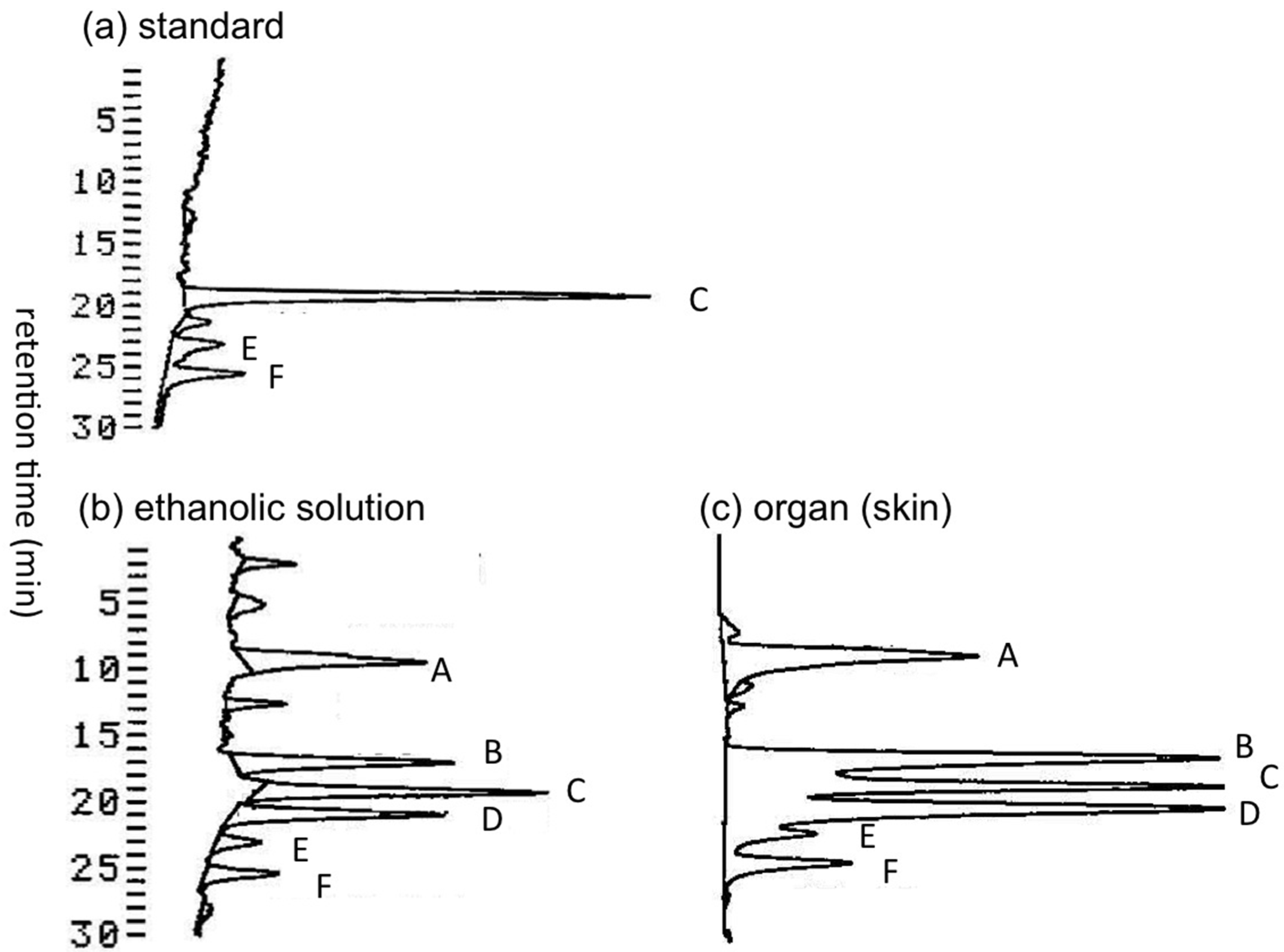
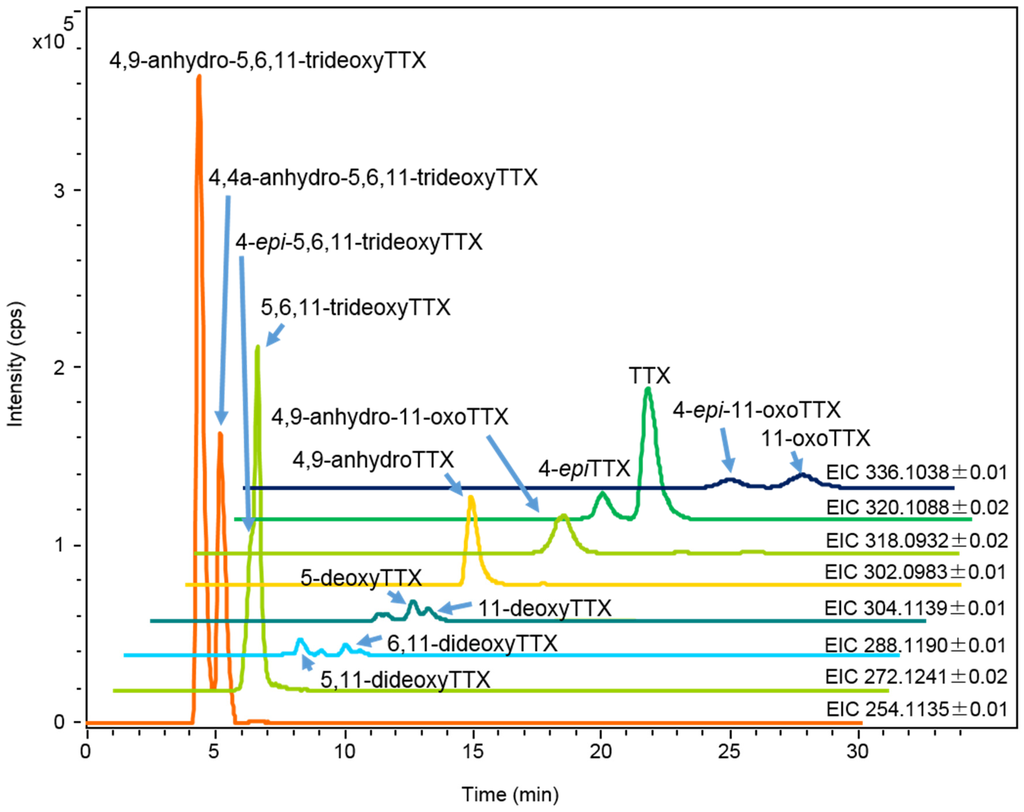
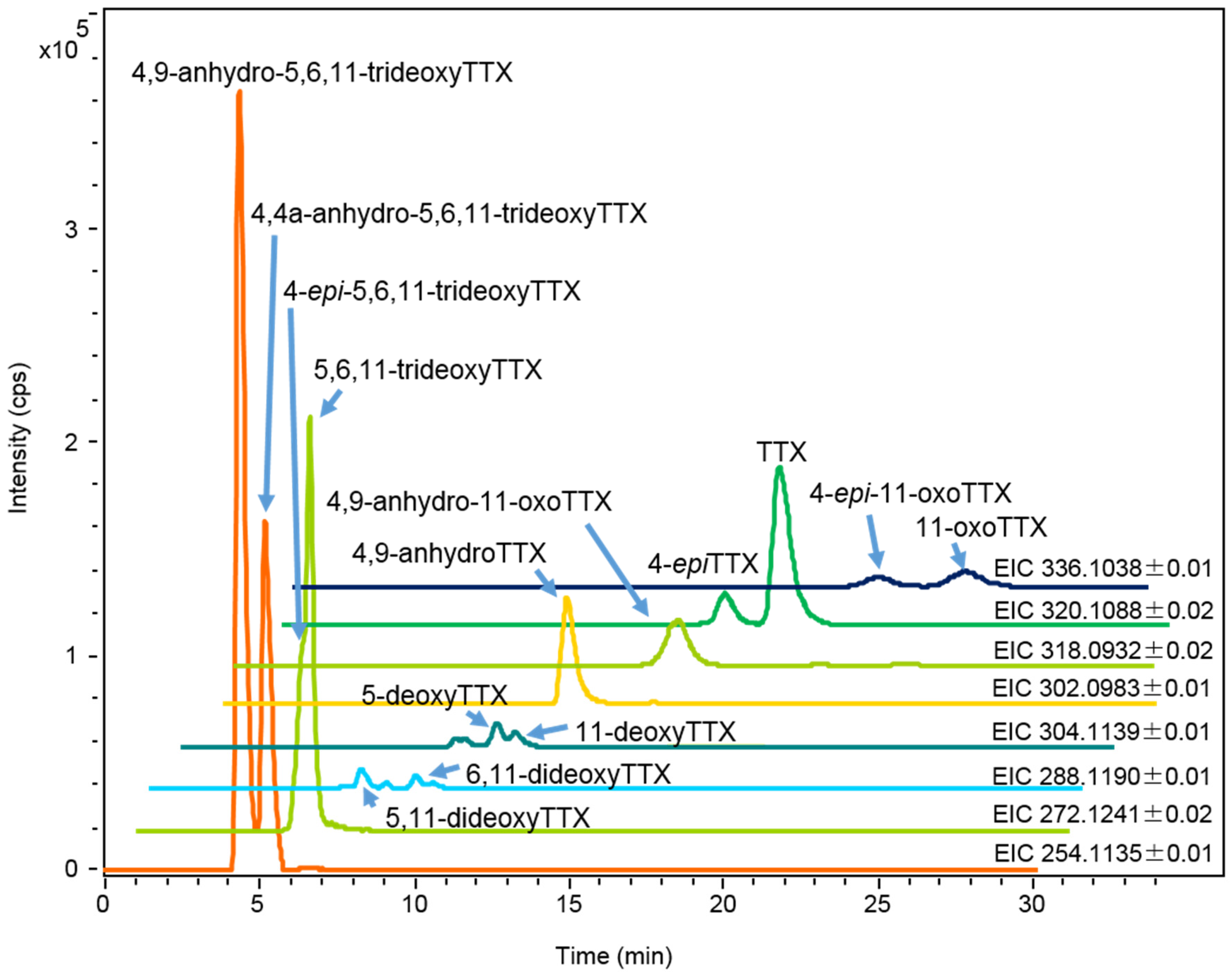
Figure 3.
HR-LC-MS chromatograms (extracted ion chromatograms: EICs) of charcoal treated sample solution (2 µL) prepared from the skin of A. hispidus No.3 specimen (25 mL/g) from the Solomon Islands soaked in ethanolic solution. The sample solution contained TTX 1.4 µg/mL, 4-epiTTX 0.30 µg/mL, 4,9-anhydroTTX 0.80 µg/mL, 5-deoxyTTX 0.13 µg/mL, 6,11-dideoxyTTX 0.03 µg/mL, 5,11-dideoxyTTX 0.09 µg/mL, 5,6,11-trideoxyTTX and 4-epi-5,6,11-trideoxyTTX (total of two analogues 1.8 µg/mL), 4,4a-anhydro-5,6,11-trideoxyTTX 2.7 µg/mL, 11-oxoTTX 0.50 µg/mL, 4-epi-11-oxoTTX 0.50 µg/mL, and 4,9-anhydro-11-oxoTTX 0.60 µg/mL.
Figure 3.
HR-LC-MS chromatograms (extracted ion chromatograms: EICs) of charcoal treated sample solution (2 µL) prepared from the skin of A. hispidus No.3 specimen (25 mL/g) from the Solomon Islands soaked in ethanolic solution. The sample solution contained TTX 1.4 µg/mL, 4-epiTTX 0.30 µg/mL, 4,9-anhydroTTX 0.80 µg/mL, 5-deoxyTTX 0.13 µg/mL, 6,11-dideoxyTTX 0.03 µg/mL, 5,11-dideoxyTTX 0.09 µg/mL, 5,6,11-trideoxyTTX and 4-epi-5,6,11-trideoxyTTX (total of two analogues 1.8 µg/mL), 4,4a-anhydro-5,6,11-trideoxyTTX 2.7 µg/mL, 11-oxoTTX 0.50 µg/mL, 4-epi-11-oxoTTX 0.50 µg/mL, and 4,9-anhydro-11-oxoTTX 0.60 µg/mL.
2.1.2. Characterization and Quantitation of TTX and Its Analogues Using High Resolution-LC-MS
Each tissue soaked in ethanolic solution was extracted with hot 0.2 M acetic acid (v/v). The extract was purified with activated charcoal and analyzed by High Resolution (HR) HILIC-LC-MS (Q-TOF MS) as previously reported [17,43,44,45]. The typical mass chromatograms of major TTX analogues in the skin of A. hispidus No.3 specimen are shown in Figure 3. All TTX analogues were quantified by HR-LC-MS based on the standard curve for TTX. However, in the samples from the Solomon Islands, the peak areas of 11-oxoTTX and 4-epi-11-oxoTTX on the extracted ion chromatograms (EIC) at m/z 336.1014 were smaller than that predicted by the LC-FLD analysis. Probably ionization was suppressed by certain compounds eluted at the same retention time. Therefore, 11-oxoTTX and 4-epi-11-oxoTTX were quantified using LC-FLD. The limit of detection (LOD) of all TTX analogues, except 11-oxoTTX and 4-epi-11-oxoTTX in the samples from the Solomon Islands, was (S/N > 3, 0.01 µg/g), while the limit of quantitation (LOQ) was (S/N > 10, 0.03 µg/g). The LOD and LOQ of 11-oxoTTX and 4-epi-11-oxoTTX in the samples from the Solomon Islands were 0.04 µg/g and 0.14 µg/g, respectively. TTX, 4,9-anhydroTTX, 5,6,11-trideoxyTTX (with its 4-epi form), 4,4a-anhydro-5,6,11-trideoxyTTX [46], and 4,9-anhydro-5,6,11-trideoxyTTX were major TTX analogues in almost all the skin samples. In addition, 11-oxoTTX which is considered more potent than TTX by some authors [23,24], was observed with its 4-epi and 4,9-anhydro forms. The MS/MS spectrum of 11-oxoTTX showed a similar fragmentation pattern to that of TTX, detecting m/z 162.0660 (C8H8N3O) and 178.0614 (C8H8N3O2) ions, corresponding to 2-aminohydroquinazoline and 2-aminodihydroquinazone, respectively [17], as the major fragment ions (Figure 4). Regarding minor analogues, monodeoxyTTX (11-deoxyTTX and 5-deoxyTTX) and dideoxyTTX (5,11-dideoxyTTX and 6,11-dideoxyTTX) were also detected.
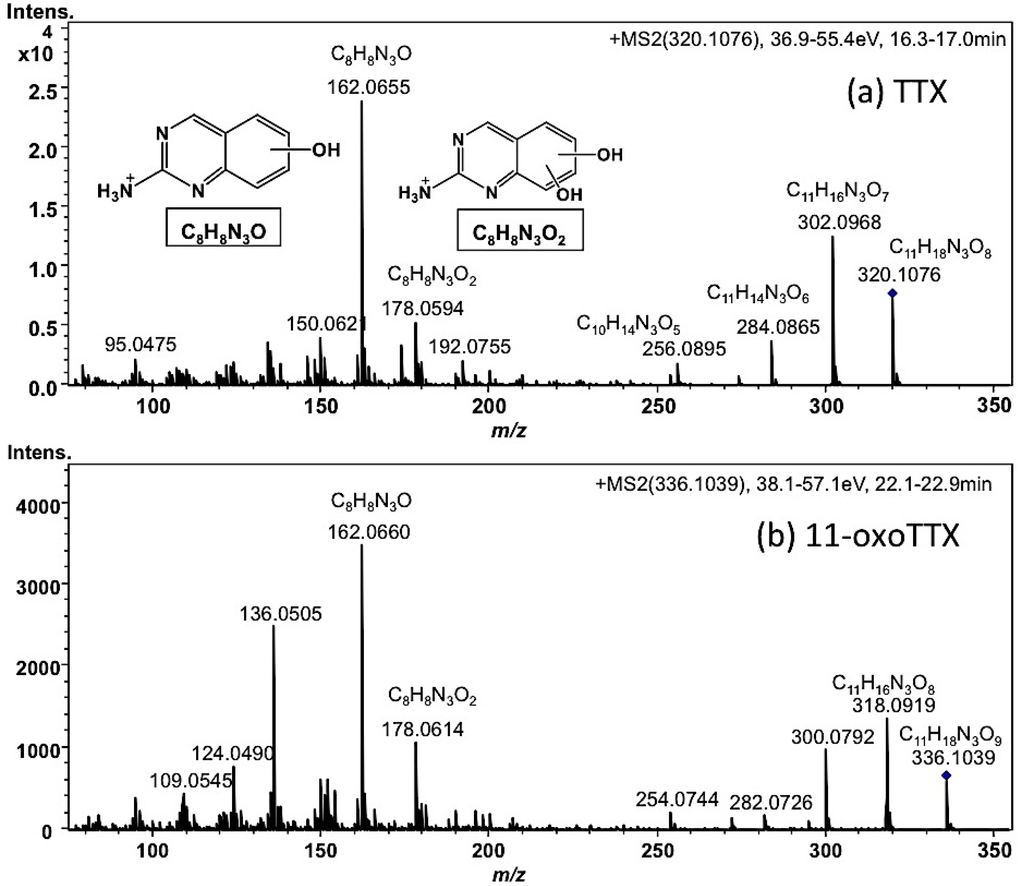
Figure 4.
HR-LC-MS/MS spectra of TTX (a) and 11-oxoTTX (b) in the skin of A. hispidus No.3 specimen from the Solomon Islands. The sample solution (2 µL) was same as shown in Figure 3. The predicted structures of the major fragment ions were superimposed in the spectra.
Figure 4.
HR-LC-MS/MS spectra of TTX (a) and 11-oxoTTX (b) in the skin of A. hispidus No.3 specimen from the Solomon Islands. The sample solution (2 µL) was same as shown in Figure 3. The predicted structures of the major fragment ions were superimposed in the spectra.
2.1.3. Estimation of the Concentrations of TTX and Its Analogues in Each Organ of A. hispidus, A. nigropunctatus, and D. holocanthus from the Solomon Islands
For three specimens of A. hispidus and A. nigropunctatus, and two specimens of D. holocanthus from the Solomon Islands, TTX and its analogues both in the ethanolic solutions and in the soaked organs were separately quantified using HR-LC-MS after prepurification. The results were combined together, according to solvent quantity and tissue weight, to estimate the real concentrations of TTXs in the organs. 5,6,11-TrideoxyTTX and 4,9-anhydro-5,6,11-trideoxyTTX were present relatively higher in the ethanolic solutions compared with the organs. The results of A. hispidus and A. nigropunctatus (three specimens for each species) from the Solomon Islands are summarized in Table 1 and Table 2, respectively. TTX, 5,6,11-trideoxyTTX, 4,4a-anhydro-5,6,11-trideoxyTTX, and 4,9-anhydro-5,6,11-trideoxyTTX were the most abundant analogues in all organs in these pufferfish species, and 11-oxoTTX which is considered a more potent analogue than TTX [23,24] was detected in the skin of all specimens. TTX concentrations in the skin were generally higher in all specimens, and relatively lower in the liver, ovary, testis, stomach, and intestine. The highest TTX concentrations in the skin of A. hispidus and A. nigropunctatus were 51.0 and 28.7 µg/g, respectively. Additionally, we confirmed that TTXs in D. holocanthus from the Solomon Islands were less than the limit of detection (LOD; S/N > 3, 0.01 µg/g) in all the organs tested (skin, liver, gonad, stomach, intestine, and flesh), even though there were only two specimens (a male and a female).

Table 1.
The concentrations of TTX and its analogues (µg/g) in the organs of A. hispidus from the Solomon Islands.
| Organ | Skin | Liver | Ovary | Testis | Stomach | Intestine | Flesh |
|---|---|---|---|---|---|---|---|
| n = 3 | n = 3 | n = 1 | n = 2 | n = 3 | n = 2 | n = 1 | |
| TTX | 7.20–51.0 | 0.03–7.99 | 1.89 | <LOD, 11.7 | 0.58–9.45 | 1.95, 5.45 | 0.07 |
| 4-epiTTX | 1.80–14.8 | <LOD-1.29 | 0.23 | <LOD, 1.93 | <LOD-1.52 | <LOD, 0.27 | <LOD |
| 4,9-anhydroTTX | 3.60–26.0 | <LOD-4.00 | <LOD | <LOD, 6.63 | <LOD-4.93 | <LOD, <LOQ(0.02) | <LOD |
| 11-deoxyTTX | 0.10–4.32 | <LOD-0.67 | <LOD | <LOD, 1.17 | <LOD-0.66 | <LOD, 0.06 | <LOD |
| 5-deoxyTTX | <LOD-5.51 | <LOD-0.21 | <LOD | <LOD, 0.35 | <LOD-0.16 | 0.10, 0.10 | <LOD |
| 6,11-dideoxyTTX | <LOD-1.47 | <LOD-0.08 | <LOD | <LOD | <LOD-0.06 | <LOD | <LOD |
| 5,11-dideoxyTTX | <LOD-3.23 | <LOD-0.14 | <LOD | <LOD | <LOD-0.56 | <LOD, 0.16 | <LOD |
| 5,6,11-trideoxyTTX | 1.47–84.3 | <LOD-16.4 | 10.1 | <LOD, 14.6 | <LOD-19.5 | 5.54, 7.20 | 0.05 |
| 4,4a-anhydro-5,6,11-trideoxyTTX | 1.36–70.1 | <LOD-10.4 | 6.74 | <LOD, 8.99 | 7.34–7.92 | 4.93, 5.36 | 0.11 |
| 4,9-anhydro-5,6,11-trideoxyTTX | 2.06–133.2 | 0.34–12.2 | 7.94 | <LOD, 9.15 | 7.71–21.6 | 5.04, 10.7 | 0.11 |
| 11-norTTX-6(S)-ol | <LOD-2.23 | <LOD-0.51 | 0.04 | <LOD, 1.02 | <LOD-0.74 | <LOD, 0.07 | <LOD |
| 11-oxoTTX | 0.32–7.07 | <LOD-0.35 | <LOD | <LOD, 0.41 | <LOD-0.31 | < LOQ(0.05), 1.68 | <LOD |
| 4-epi-11-oxoTTX | <LOQ(0.06)-3.53 | <LOD-<LOQ(0.06) | <LOD | <LOD, <LOQ(0.09) | <LOD-<LOQ(0.13) | <LOD, 0.26 | <LOD |
| 4,9-anhydro-11-oxoTTX | <LOD-13.7 | <LOD-0.18 | 0.22 | <LOD, 0.58 | <LOQ(0.08)-0.41 | 0.21, 1.72 | <LOD |
For 11-oxoTTX and 4-epi-11-oxoTTX: LOD = 0.04 µg/g (S/N > 3), LOQ = 0.14 µg/g (S/N > 10), for other analogues: LOD = 0.01 μg/g (S/N > 3), LOQ = 0.03 μg/g (S/N > 10). 5,6,11-TrideoxyTTX was measured as the mixture with its 4-epi form.
Table 2.
The concentrations of TTX and its analogues (µg/g) in the organs of A. nigropunctatus from the Solomon Islands.
| Organ | Skin | Liver | Ovary | Testis | Stomach | Intestine |
|---|---|---|---|---|---|---|
| n = 3 | n = 3 | n = 2 | n = 1 | n = 2 | n = 1 | |
| TTX | 8.51–28.7 | <LOD-19.7 | 1.14, 19.7 | <LOD | 1.57, 21.9 | 0.38 |
| 4-epiTTX | 1.61–6.68 | <LOD-3.11 | 0.28, 3.16 | <LOD | 0.41, 3.04 | 0.05 |
| 4,9-anhydroTTX | 5.15–15.9 | <LOD-10.8 | 0.91, 8.60 | <LOD | <LOD, 11.0 | 0.19 |
| 11-deoxyTTX | <LOD-0.83 | <LOD | <LOD | <LOD | <LOD | <LOD |
| 5-deoxyTTX | <LOD-0.14 | <LOD | <LOD | <LOD | <LOD | <LOD |
| 6,11-dideoxyTTX | <LOD-0.10 | <LOD | <LOD | <LOD | <LOD | <LOD |
| 5,11-dideoxyTTX | <LOD | <LOD | <LOD | <LOD | <LOD | <LOD |
| 5,6,11-trideoxyTTX | 3.99–13.0 | <LOD-10.7 | 1.12, 6.14 | <LOD | 1.25, 9.17 | 0.55 |
| 4,4a-anhydro-5,6,11-trideoxyTTX | 2.39–8.44 | 0.02–10.1 | 0.92, 5.89 | <LOD | 1.32, 8.63 | 0.53 |
| 4,9-anhydro-5,6,11-trideoxyTTX | 3.90–11.1 | <LOD-11.0 | 1.07, 6.73 | <LOD | 1.31, 8.94 | 0.54 |
| 11-norTTX-6(S)-ol | 0.10–0.84 | <LOD | <LOD | <LOD | <LOD | <LOD |
| 11-oxoTTX | 0.16–0.67 | <LOD-0.77 | <LOD, 0.56 | <LOD | <LOQ(0.05), 0.80 | <LOD |
| 4-epi-11-oxoTTX | <LOQ(0.09)-0.50 | <LOD-0.27 | <LOD, 0.22 | <LOD | <LOD, 0.26 | <LOD |
| 4,9-anhydro-11-oxo-TTX | 0.33–1.49 | <LOD-1.11 | <LOD, 0.66 | <LOD | 0.19, 0.93 | <LOQ(0.07) |
For 11-oxoTTX and 4-epi-11-oxoTTX: LOD = 0.04 µg/g (S/N > 3), LOQ =0.14 µg/g (S/N > 10), for other analogues: LOD = 0.01 μg/g (S/N > 3), LOQ = 0.03 μg/g (S/N > 10). 5,6,11-TrideoxyTTX was measured as the mixture with its 4-epi form.
2.2. The concentrations of TTX and Its Analogues in Each Organ of A. hispidus and A. nigropunctatus From Okinawa, Japan
The pufferfish specimens from Okinawa were transported live to Sendai and immediately frozen at −25 °C. TTXs in each organ of three specimens of A. hispidus and four specimens of A. nigropunctatus were extracted and pre-purified with activated charcoal, and were analyzed using HR-LC-MS. The data obtained for A. hispidus and A. nigropunctatus are shown in Table 3 and Table 4, respectively. The mass chromatograms and organ distribution of toxins in these Arothron species from Okinawa, Japan, were very similar to those from the Solomon Islands. However, one specimen of A. nigropunctatus from Okinawa (No.1 male specimen) showed very high levels of TTX and 11-oxoTTX in the skin, 255 and 42.4 µg/g, respectively. TTX concentration in the stomach of this specimen (25.5 µg/g) was higher than other organs. 11-OxoTTX was detected in the skin of all specimens of both species, same as those from the Solomon Islands.
Table 3.
The concentrations of TTX and its analogues (µg/g) in the organs of A. hispidus from Okinawa, Japan.
| Organ | Skin | Liver | Ovary | Testis | Stomach | Intestine | Flesh | Spleen |
|---|---|---|---|---|---|---|---|---|
| n = 3 | n = 3 | n = 1 | n = 2 | n = 3 | n = 3 | n = 3 | n = 3 | |
| TTX | 4.26–12.7 | 0.17–0.55 | 0.37 | 1.08, 1.11 | <LOD-2.44 | 1.61-2.05 | 0.23–0.61 | 0.15–2.09 |
| 4-epiTTX | 0.40-2.75 | <LOD-0.03 | 0.04 | 0.07, 0.17 | <LOD-0.52 | 0.10-0.36 | <LOD-0.04 | <LOD-0.25 |
| 4,9-anhydroTTX | 2.22–8.84 | <LOD-0.26 | 0.17 | 0.46, 0.52 | <LOD-1.74 | 0.59–1.36 | 0.04–0.18 | <LOD-0.74 |
| 11-deoxyTTX | 1.34–2.12 | <LOD | <LOD | 0.04, 0.18 | <LOD-0.21 | <LOD-0.32 | <LOD | <LOD-0.11 |
| 5-deoxyTTX | 0.73–1.19 | <LOD | <LOD | 0.11, 0.16 | <LOD-0.05 | <LOD-0.31 | <LOD | <LOD |
| 6,11-dideoxyTTX | 0.12–0.31 | <LOD | <LOD | <LOD, 0.05 | <LOD | <LOD-<LOQ(0.02) | <LOD | <LOD |
| 5,11-dideoxyTTX | 0.11–0.52 | <LOD | <LOD | <LOD, 0.03 | <LOD | <LOD-<LOQ(0.02) | <LOD | <LOD |
| 5,6,11-trideoxyTTX | 1.09–6.02 | < 0.06 | 0.18 | < 0.20, 0.56 | < 0.10-0.19 | 0.21–0.36 | 0.03–0.23 | <LOD-0.10 |
| 4,4a-anhydro-5,6,11-trideoxyTTX | 1.91–18.6 | 0.31–0.46 | 0.33 | 0.24, 0.67 | < 0.14–0.80 | 0.42–0.73 | 0.07–0.31 | 0.03–0.55 |
| 4,9-anhydro-5,6,11-trideoxyTTX | 7.36–46.7 | 0.44–1.66 | 1.11 | 1.70, 3.79 | < 0.46–2.23 | 1.77–2.80 | 0.21–0.45 | 0.15–1.10 |
| 11-norTTX-6(S)-ol | 0.27–0.92 | <LOD | <LOD | 0.04, 0.06 | <LOD-0.18 | 0.08–0.12 | <LOD-0.05 | <LOD-0.12 |
| 11-oxoTTX | <LOQ(0.07)-0.20 | <LOD | <LOD | <LOD, <LOQ(0.12) | <LOD | <LOD- <LOQ(0.05) | <LOD | <LOD |
| 4-epi-11-oxoTTX | <LOD | <LOD | <LOD | <LOD | <LOD | <LOD | <LOD | <LOD |
| 4,9-anhydro-11-oxoTTX | <LOQ(0.12)-0.15 | <LOD | <LOD | <LOD, <LOQ(0.07) | <LOD-< LOQ(0.08) | <LOD- <LOQ(0.09) | <LOD | <LOD |
For 11-oxoTTX, 4-epi-11-oxoTTX and 4,9-anhyydro-11-oxoTTX: LOD = 0.04 µg/g (S/N > 3), LOQ = 0.14 µg/g (S/N > 10), for other analogues: LOD = 0.01 μg/g (S/N > 3), LOQ = 0.03 μg/g (S/N > 10). 5,6,11-TrideoxyTTX was measured as the mixture with its 4-epi form.
Table 4.
The concentrations of TTX and its analogues (µg/g) in the organs of A. nigropunctatus from Okinawa, Japan.
| Organ | Skin | Liver | Ovary | Testis | Stomach | Intestine | Flesh | Spleen |
|---|---|---|---|---|---|---|---|---|
| n = 4 | n = 4 | n = 1 | n = 3 | n = 4 | n = 4 | n = 4 | n = 4 | |
| TTX | 12.6–255 | 0.29–7.46 | 1.02 | 0.26–10.1 | 0.57–25.5 | 1.22–4.51 | 0.19–4.26 | 0.85–4.19 |
| 4-epiTTX | 1.19–27.9 | <LOD-0.49 | 0.09 | <LOD-2.64 | 0.07–2.05 | 0.07–0.35 | <LOD-0.92 | 0.18–0.62 |
| 4,9-anhydroTTX | 5.39–96.6 | <LOD-2.75 | 0.50 | 0.13–6.13 | 0.24–9.65 | 0.52–2.41 | <LOD-1.82 | 0.85–2.26 |
| 11-deoxyTTX | 1.25–14.9 | <LOD-0.54 | 0.10 | 0.02–3.13 | 0.06–2.19 | 0.15–0.65 | <LOD-0.88 | 0.10–0.76 |
| 5-deoxyTTX | <LOD-4.03 | <LOD-0.23 | 0.09 | 0.10–3.80 | <LOD-5.41 | <LOD-0.54 | <LOD-0.62 | <LOD-0.80 |
| 6,11-dideoxyTTX | 0.40–1.77 | <LOD-0.07 | 0.05 | <LOD-1.31 | 0.02–0.84 | <LOD-0.27 | <LOD | <LOD-0.43 |
| 5,11-dideoxyTTX | 0.52–2.92 | <LOD-0.06 | <LOQ(0.02) | 0.10–0.94 | <LOD-1.03 | <LOD-0.12 | <LOD-0.18 | <LOD-0.14 |
| 5,6,11-trideoxyTTX | 15.3–275 | <LOD -3.69 | 0.39 | 0.61–22.3 | 0.28–25.2 | <LOD-8.40 | 0.04–3.92 | 0.74–10.0 |
| 4,4a-anhydro-5,6,11-tri-deoxyTTX | 6.81–178 | 0.34–2.69 | 0.56 | 0.45–15.3 | 0.39–10.2 | 0.74–5.31 | 0.34–3.32 | 1.01–5.69 |
| 4,9-anhydro-5,6,11-trideoxyTTX | 43–438 | 0.30–8.28 | 3.48 | 2.05–26.9 | 1.42–77.0 | 0.93–11.0 | 0.70–4.79 | 3.74–10.5 |
| 11-norTTX-6(S)-ol | 0.11–9.72 | <LOD-0.30 | <LOD | <LOD-0.32 | <LOD-0.11 | <LOD-0.19 | <LOD-0.11 | <LOD-0.15 |
| 11-oxoTTX | 3.94–42.4 | <LOQ(0.05)-0.84 | <LOQ(0.12) | <LOQ(0.07)-0.82 | <LOQ(0.05)-9.98 | <LOQ(0.12)-0.53 | <LOD-<LOQ(0.11) | 0.12–0.46 |
| 4-epi-11-oxoTTX | 0.51–6.54 | <LOD-0.16 | <LOD | <LOD-0.16 | <LOD-1.81 | <LOD-<LOQ(0.10) | <LOD | 0.05–0.16 |
| 4,9-anhydro-11-oxoTTX | 2.57–25.0 | <LOQ(0.04)-0.5 | 0.15 | < LOQ(0.05)-0.69 | <LOQ(0.11)-4.15 | 0.20–0.41 | <LOD-0.35 | 0.09–0.53 |
For 11-oxoTTX, 4-epi-11-oxoTTX and 4,9-anhydro-11-oxoTTX: LOD = 0.04 µg/g (S/N > 3), LOQ = 0.14 µg/g (S/N > 10), for other analogues: LOD = 0.01 μg/g (S/N > 3), LOQ = 0.03 μg/g (S/N > 10). 5,6,11-TrideoxyTTX was measured as the mixture with its 4-epi form.
2.3. Analysis of Saxitoxin (STX) and Its Analogues in A. hispidus and A. nigropunctatus from the Solomon Islands and Okinawa, Japan
The presence of STX, neoSTX, and dcSTX was investigated in the skin and ovary of the pufferfish from the Solomon Islands and Okinawa, Japan, using HR-LC-MS. The concentration of STX was below the LOD (0.02 µg/g) in the skin and ovary of all pufferfish from the Solomon Islands and all four specimens of A. nigropunctatus from Okinawa, Japan. However, STX was detected in the skin (0.09, 2.80, 0.34 µg/g) of all three specimens of A. hispidus from Okinawa, Japan. A female specimen A. hispidus (No. 2) from Okinawa, which possessed highest STX (2.80 µg/g) and dcSTX (0.66 µg/g) in the skin, also contained trace amount of STX (0.04 µg/g: between LOD and LOQ) in the ovary. In other samples, dcSTX was not detected. NeoSTX was not detected in all samples. LOD (S/N > 3) and LOQ (S/N > 10) for STX were 0.02 µg/g and 0.07 µg/g, neoSTX, 0.05 µg/g and 0.18 µg/g, and dcSTX, 0.07 µg/g and 0.25 µg/g, respectively.
3. Discussion
We estimated the concentrations of TTX and its analogues in the pufferfish, A. hispidus and A. nigropunctatus, from the Solomon Islands and Okinawa, Japan, using HR-LC-MS. The pufferfish organs from the Solomon Islands were transported in ethanol-water (7:3, v/v) at room temperature to Japan. Based on the LC-FLD analysis, the amounts of tetrodonic acid type analogs, likely derived from TTXs under such condition, was estimated to be almost 2/3 of total TTX. Thus, we should note that the amount of TTX in the samples from the Solomon Islands (Table 1 and Table 2) could be underestimated. However, we believe that the data obtained in this study must be reported to warn of the high toxicity of these species in the Solomon Islands. The flesh of only one specimen of A. hispidus from the Solomon Islands was analyzed, due to limited number storage tubes and volume of ethanolic solution available during pufferfish collection. TTX in that flesh was low (0.07 µg/g), however, the levels of TTX in the skin of A. hispidus and A. nigropunctatus were high in all specimens of both species. In addition, TTX was detected in the flesh of all specimens of these species from Okinawa, Japan (A. hispidus from 0.23 to 0.61 µg/g; A. nigropunctatus from 0.19 to 4.26 µg/g). Based on these data, these species, including flesh, from the Solomon Islands and Okinawa, Japan, should be banned from consumption. Khora et al. [47] and Teruya et al. [48] also indicated high toxicity in the flesh of these Arothron species from Okinawa, Japan, based on the result of a mouse bioassay.
The distribution of TTXs in the organs of A. hispidus and A. nigropunctatus from the Solomon Islands and Okinawa, Japan, revealed that the skin is the major location of TTX accumulation. The skin of A. hispidus and A. nigropunctatus from the Solomon Islands contained 7.20 to 51.0 µg/g and 8.51 to 28.7 µg/g, respectively, while those from Okinawa, Japan, were 4.26 to 12.7 µg/g (A. hispidus), and 12.6 to 255 µg/g (A. nigropunctatus). These data suggested that the concentration of TTX is highly variable among specimens. Our findings agree with previous reports that showed higher toxicity to mice in the skin tissues of the same species collected in Okinawa area, Japan [47]. A study on the Hawaiian A. hispidus suggested high levels of TTX producing bacteria isolated from the skin [49], although the details have not yet been clarified. The levels of TTX in the liver and gonads are usually high in common Japanese pufferfish species in genus Takifugu (Fugu) [50], but they were low in the Arothron species compared with those of the skin. We assume that different regions, with their different biotic and abiotic factors, may play an important role in the TTX accumulation ability of pufferfish. More specimens are needed to compare the TTX distribution profile in the organs of the pufferfish from the Pacific region. The difference of toxin distribution pattern between Takifugu and Arothron is probably caused by the difference of some proteins which are implicated in TTX accumulation system. We previously found a glycoprotein, named as PSTBP (puffer fish saxitoxin and tetrodotoxin binding protein), that binds to TTX and STX in the blood plasma of Takifugu pardalis [51], and also, it was reported that PSTBP was commonly detected in several Takifugu species [52,53]. We are planning to examine the presence of PSTBP-like protein in Arothron species to get a hint that will help explain the difference of TTX-accumulation tissues between Takifugu and Arothron genus.
We also analyzed TTX and its analogues in several organs of two specimens of D. holocanthus from the Solomon Islands, and confirmed that the concentrations of all TTX analogues were below the LOD (S/N > 3, 0.01 µg/g). We will further analyze more specimens for confirmation in the future. In Japan, the skin, flesh, and testis of D. holocanthus collected in Japan are officially allowed for consumption [39].
As previously noted [40,50], 4,9-anhydroTTX is one of the major TTX analogues in almost all toxic pufferfish tissues, because it is chemically equilibrated with TTX. Likewise, 5,6,11-trideoxyTTX, 4,4a-anhydro-5,6,11-trideoxyTTX and 4,9-anhydro-5,6,11-trideoxyTTX also showed significant concentrations in all toxic pufferfish being investigated. For instance, 4,9-anhydro-5,6,11-trideoxyTTX concentrations even surpassed those of TTX in tissues of a few individuals (Table 1, Table 3 and Table 4). Previous studies revealed that 5,6,11-trideoxyTTX, which is also a major analogue in other pufferfish species such as Japanese Takifugu pardalis [50] and European trumpet shell Charonia lampas [54], has a relatively low toxicity (LD99 720 µg/kg, mice, i.p.) [21] compared to TTX and other toxic analogues. In addition, 11-oxoTTX was commonly detected in A. nigropunctatus and A. hispidus from the Solomon Islands and Okinawa, Japan (Table 1, Table 2, Table 3 and Table 4). This analogue was originally isolated from Micronesian A. nigropunctatus [22], however, this is the first identification in A. hispidus. 11-OxoTTX has been also found in the Brazialian frog [55], the red spotted newt [56], the blue-ringed octopus [57], the marine snail [58] and the xanthid crab [59]. Because 11-oxoTTX is considered as more potent than TTX by some authors [23,24], its presence in A. hispidus and A. nigropunctatus poses a potential risk of pufferfish poisoning.
The presence of STX and its analogues were also tested in the skin and ovary of A. hispidus and A. nigropunctatus from the Solomon Islands and Okinawa, Japan. STX was detected in the skin of all three specimens of A. hispidus from Okinawa (0.09, 2.80, 0.34 µg/g), as well as in the ovary of one specimen, but not detected in the pufferfish from the Solomon Islands (< 0.02 µg/g). It is notable that the concentration of STX equivalents (STX + dcSTX) was 3.46 µg/g in the skin of an A. hispidus specimen from Okinawa. This is higher than the global regulatory limit of 800 μg of STX equivalents per kg of shellfish meat (e.g. Codex, EU, US, Japan) [60,61,62,63]. Analysis of STX in more specimens from both regions is needed, because Nakashima et al. [36] reported that STX and dcSTX were detected in the ovary of Arothron firmamentum collected in Japan at high concentration. From the results obtained in this present study, we believe this investigation is informative and also important for the people of Solomon Islands regarding potential risk of pufferfish poisoning.
4. Experimental Section
4.1. Standards and Reagents
Semi-purified TTXs mixture prepared from the ovary of Takifugu poecilonolus using a charcoal column [21] was used as the standard for TTXs as described previously [17]. The authentic 11-oxoTTX was prepared from TTX by chemical oxidation [24]. TTX used to draw the standard curve for HR-LC-MS was highly purified by our group. The standard mixture of STX, dcSTX, and neoSTX were prepared (approved under AOAC guidelines) by Oshima [64]. All of the solvents for LC and the reagents were purchased from Wako Pure Chemical Industries (Osaka, Japan), while ammonium formate for LC-MS, heptafluorobutyrate acid, and acetonitrile for LC-FLD were purchased from Sigma-Aldrich (St. Louis, MO, USA).
4.2. Sampling and Test Materials
Three specimens of Arothron hispidus, three specimens of A. nigropunctatus and two specimens of Diodon holocanthus were collected in the Tomba islets (latitude 8.427° S, longitude 157.929° W) Marovo Lagoon, Solomon Islands, in May and July 2014. The body length, body weight, and gender of the three specimens of A. hispidus were No.1 19 cm, 130 g, female; No.2 17 cm, 118 g, male; No.3 18 cm, 120 g, male, those of the three specimens of A. nigropunctatus were No.1 19 cm, 163 g, female; No.2 37 cm, 320 g, male; No.3 20 cm, 180 g, female, and those of two specimens of D. holocanthus were 30–35 cm, 300–350 g, No.1 female, No.2 male. Immediately after collection, the pufferfish were dissected into respective organs (skin, liver, gonad, stomach, intestine, flesh), and a part or whole of each organ were separately soaked in approximately 15 mL of ethanol-water (7:3, v/v) in a disposable plastic tube (TPP, Trasadingen, Switzerland). Samples were transported to Tohoku University, Sendai, Japan, at room temperature, because preserving samples at low temperature (freezing) was not possible due to the lack of proper freezing equipment and the long transit time for air-shipment from the Solomon Islands. Upon arrival samples were kept below −25 °C until use. From Okinawa, Japan, four living specimens of A. nigropunctatus (No.1 15 cm, 67 g, male; No.2 21 cm, 375 g, male; No.3 14 cm, 70 g, male; No.4 16 cm, 130 g, female) and three living specimens of A. hispidus (No.1 17 cm, 112 g, male; No.2 16.5 cm, 124.4 g, female; No.3 10.0 cm, 31.3 g, male), collected from April to June 2015, were sent to Tohoku University. After arrival, they were dissected into organs and kept frozen at −25 °C until use.
4.3. Preliminary Analysis of TTXs in the Skin of A. hispidus from the Solomon Islands Using LC-FLD for TTXs
An aliquot (0.5 mL) of the ethanol-water (7:3, v/v) solution used to soak the skin of A. hispidus three specimens from the Solomon Islands was acidified by acetic acid and dried in vacuo, and then dissolved in 0.5 mL of 0.05 M acetic acid. After centrifugation, a part of the supernatant was diluted ten-fold with 0.05 M acetic acid. An aliquot (2 µL) of this solution was applied to LC-FLD for TTXs measurement. Similarly, the skin from these specimens was removed from the ethanolic solution, and a part (0.25 g) was extracted with 1.25 mL of 0.2 M acetic acid (v/v) in boiling water. The extract was centrifuged at 15,000× g and an aliquot (2 µL) of the supernatant was applied to LC-FLD [41,42].
4.4. Sample Preparation for HR-LC-MS Analysis
4.4.1. From the Ethanolic Solution Used to Soak the Pufferfish Organs from the Solomon Islands
An aliquot (0.5 mL) of ethanol-water (7:3, v/v) solution (from a total of 14.9 mL for the skin of A. hispidus No.3 for example) was mixed with 5 µL of acetic acid, and dried in vacuo. The obtained residue was dissolved in 0.5 mL of 0.05 M acetic acid and neutralized with 1 M NaOH aqueous solution, and then applied to activated charcoal (column volume: 250 μL, for chromatography, Wako, Osaka, Japan) packed in a glass pipette equilibrated with water. After washing charcoal with 0.75 mL of water, TTXs were eluted with 1.5 mL of acetic acid-ethanol-water (2:50:48, v/v). An aliquot (50 µL) of this eluate was dried in vacuo and dissolved in 50 µL of 0.05 M acetic acid, and then filtered through Cosmospin Filter H (Nacalai Tesque, Inc., Kyoto, Japan). After dilution of the filtrate with 0.05 M acetic acid, an aliquot of this solution was used for HR-LC-MS. The recovery of TTXs in the organs from the activated charcoal was estimated at approximately 70% using LC-FLD. Therefore, the final TTXs concentrations in the organs were recalculated based on this recovery ratio.
4.4.2. From the Organs of Solomon Islands Pufferfish Soaked in Ethanolic Solution, and from the Frozen Organs of Pufferfish from Okinawa, Japan
The organ was homogenized and an aliquot (0.25 g) was mixed with 1.25 mL of acetic acid-water (1:99, v/v). The mixture was boiled for 10 minutes in a 1.5 mL micro tube, and centrifuged at 15,000× g for 15 minutes at 4 °C. Half of the supernatant was neutralized with 1 M NaOH, and then applied to the charcoal (column volume 250 µL). The sample was treated by the same procedure as described above Section (4.4.1).
4.5. LC-FLD for TTXs
Post-column LC-fluorescent detection (LC-FLD) for TTXs was performed as described previously [41,42]. Briefly, The LC condition was as follow: Develosil C30 UG-5 (0.46 cm i.d. × 25 cm) (Nomura Chemical, Seto, Japan) with 30 mM ammonium heptafluorobutyrate buffer (pH 5.0) and 10 mM ammonium formate buffer (pH 5.0) containing 1% (v/v) acetonitrile at a flow rate of 0.4 mL/min; for the post column reaction 4 M NaOH at a flow rate of 0.7 mL/min to be heated at 105 °C in the stainless tube (0.46 mm i.d. × 3.5 m). The chromatography was performed at 20 °C. The derived fluorophores by post column reaction were detected by a Jasco FP2025 plus fluoromonitor (Jasco, Tokyo, Japan) setting excitation wavelength at 365 nm and emission wavelength at 510 nm.
4.6. High Resolution LC-MS and LC-MS/MS for TTXs
HILIC LC-MS and MS/MS methods were performed as previously reported [17]. Briefly, a Shimadzu Nexera UHPLC System (Shimadzu, Kyoto, Japan) was used as the liquid chromatography system, and consisted of a LC-10AD pump (Shimadzu, Kyoto, Japan), autosampler (SIL-30AC, Shimadzu) and a TSKgel Amide-80 column (150 × 2.0 mm i.d., particle size 5 µm, Tosoh, Tokyo, Japan). The mobile phase, 16 mM ammonium formate in water/acetonitrile/formic acid (30:70:0.002, v/v) has a flow rate of 0.2 mL/min at 28 °C in the isocratic mode with an injection volume of 2–3 μL and run time of 25 min. The liquid chromatography system was connected to a Q-TOF MS spectrometer, MicrOTOFQII (Bruker Daltonics, Bremen, Germany), equipped with an ESI source. The conditions of the MS spectrometer were as follows: positive ionization mode, dry gas: nitrogen 7 L/min, dry temperature: 180 °C, nebulizer: 1.6 Bar, capillary: −4500 V. The ions at m/z 320.1088 (TTX, 4-epiTTX), 302.0983 (4,9-anhydroTTX), 304.1139 (11-deoxyTTX, 5-deoxyTTX), 288.1190 (6,11-trideoxyTTX, 5,11-dideoxyTTX), 272.1241 (5,6,11-trideoxyTTX, 4-epi-5,6,11-trideoxyTTX) and 254.1135 (4,9-anhydro-5,6,11-trideoxyTTX, 4,4a-anhydro-5,6,11-trideoxyTTX), 336.1038 (11-oxoTTX, 4-epi-11-oxoTTX), 318.0932 (4,9-anhydro-11-oxoTTX) corresponding to the [M + H]+ ions were analyzed in the extracted ion chromatograms (EIC). The mass tolerance width for the ions are as shown in Figure 3 (0.01 or 0.02). As described in the Section 2.1.2, all TTX analogues were quantified by HR-LC/MS based on the standard curve for TTX. However, in the samples from the Solomon Islands, the peak of 11-oxoTTX and 4-epi-11-oxoTTX were suppressed by certain compounds eluted at the same retention time. Therefore, in that case, 11-oxoTTX and 4-epi-11-oxoTTX were quantified using LC-FLD. The limit of detection (LOD) of all TTX analogues except 11-oxoTTX and 4-epi-11-oxoTTX were (S/N > 3, 0.01 µg/g) and the limit of quantitation (LOQ) of them were (S/N > 10, 0.03 µg/g), and LOD and LOQ of 11-oxoTTX and 4-epi-11-oxoTTX were 0.04 µg/g and 0.14 µg/g, respectively. MS/MS was performed in AutoMS/MS mode setting [M + H]+ as the precursor ions. The precursor ions and sweeping collision energy were 320.1088 ± 0.1, 36.9–55.4 eV for TTX, and 336.1038 ± 0.1, 38.1–57.1 eV for 11-oxoTTX.
4.7. High Resolution LC-MS for STXs
STX, dcSTX and neoSTX were analyzed using HR-LC-MS. LC was performed with a TSKgel Amide-80 column (150 x 2.0 mm i.d., particle size 5 µm, Tosoh, Tokyo, Japan) and the mobile phase, 2 mM ammonium formate in water/acetonitrile/formic acid (30:62:0.0125, v/v) at a flow rate of 0.2 mL/min at 25 °C in the isocratic mode with an injection volume of 2.0 µL and run time of 30 min [65]. The mass spectrometer and its condition were same as those of TTXs analysis as described above (see Section 4.4). STX, dcSTX and neoSTX were detected as [M + H]+ ions at m/z 300.1415 ± 0.05, 257.1356 ± 0.05, and 316.1365 ± 0.05, respectively. LOD (S/N > 3) and LOQ (S/N > 10) for STX were 0.02 µg/g and 0.07 µg/g, neoSTX, 0.05 µg/g and 0.18 µg/g, and dcSTX, 0.07 µg/g and 0.25 µg/g, respectively.
5. Conclusions
The concentrations of TTX and its analogues in the skin was generally higher in both A. hispidus and A. nigropunctatus both from the Solomon Islands and Okinawa, Japan, and relatively lower in the liver, ovary, testis, stomach, intestine, and flesh. Because of high concentration of TTX in the skin of these two species from the Solomon Islands (51.0 and 28.7 µg/g at highest) and Okinawa, Japan (12.7 and 255 µg/g at highest), their consumption should be banned. 11-OxoTTX, which is considered a more potent analogue than TTX by some authors, was commonly detected in the skin of all specimens of both Arothron species from these two regions, an occurrence that should warn citizens of a potentially higher toxicity. STX was detected in the skin of all three specimens of A. hispidus from Okinawa, Japan. The level of STX equivalents (STX + dcSTX) in the skin of one specimen was 3.46 µg/g, more than the global regulation level. The toxicities of other Arothron species from these regions should be continuously studied.
Acknowledgments
We would like to thank the Solomon Islands' Ministry of Fisheries and Marine Resources (MFMR) and the Japanese Government Embassy in Solomon Islands for the necessary documents needed for the Solomon Islands specimens. This work was funded by Japan Society for the Promotion of Science (JSPS) through its Funding Program for the Next Generation World-Leading Researchers (LS012) and KAKENHI Grant-in-Aid for Scientific Research no. 26292057 to M.Y.Y. C.G.P. is a Japanese Government (Monbukagakusho: MEXT) Scholarship student (no. 110429). Y.K. is a research fellow of JSPS (DC1) (no. 25-5534).
Author Contributions
Puilingi C. G. conducted collection and transfer of the pufferfish samples from the Solomon Islands to Japan. He was responsible for all the sample preparation for toxin analysis, and conducted LC-FLD analysis and data analysis of HR-LC-MS. Kudo Y. mainly performed TTXs analysis using HR-LC-MS. Cho Y. and Konoki K. were charged in analysis of the data and gave important advice. Yotsu-Yamashita, M. was responsible for collection of the samples from Okinawa, Japan, HR-LC-MS analysis of STXs, and supervision of all the experimental work and preparation of the manuscript.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Yokoo, A. Chemical studies on tetrodotoxin Rept. III. Isolation of spheroidine. J. Chem. Soc. Japan 1950, 71, 591–592. [Google Scholar]
- Noguchi, T.; Maruyama, J.; Ueda, Y.; Hashimoto, K.; Harada, T. Occurrence of tetrodotoxin in the Japanese ivory shell Babylonia japonica. Bull. Japan. Soc. Sci. Fish. 1981, 47, 901–913. [Google Scholar] [CrossRef]
- Yasumura, D.; Oshima, Y.; Yasumoto, T.; Alcala, A.C.; Alcala, L.C. Tetrodotoxin and paralytic shellfish toxins in Philippine crabs. Agric. Biol. Chem. 1986, 50, 593–598. [Google Scholar] [CrossRef]
- Maruyama, J.; Noguchi, T.; Narita, H.; Jeon, J.K.; Otsuka, M.; Hashimoto, K. Occurrence of tetrodotoxin in a starfish, Astropecten scoparius. Agric. Biol. Chem. 1985, 49, 3069–3070. [Google Scholar] [CrossRef]
- Sheumack, D.D.; Howden, M.E.H.; Spence, I.; Quinn, R.J. Maculotoxin: A Neurotoxin from the venom glands of the octopus Hapalochlaena maculosa identified as tetrodotoxin. Science 1978, 199, 188–189. [Google Scholar] [CrossRef] [PubMed]
- McNabb, P.; Selwood, A.I.; Munday, R.; Wood, S.A.; Taylor, D.I.; MacKenzie, A.L.; van Ginkel, R.; Rhodes, L.L.; Cornelisen, C.; Heasman, K.; et al. Detection of tetrodotoxin from the grey sided-gilled sea slug-Pleurobranchaea maculata, and associated dog neurotoxicosis on beaches adjacent to the Hauraki Gulf, Auckland, New Zealand. Toxicon 2010, 56, 466–473. [Google Scholar] [CrossRef] [PubMed]
- McNabb, P.S.; Taylor, D.I.; Ogilvie, S.C.; Wilkinson, L.; Anderson, A.; Hamon, D.; Wood, S.A.; Peake, B.M. First detection of tetrodotoxin in the bivalve Paphies australis by liquid chromatography coupled to triple quadrupole mass spectrometry with and without precolumn reaction. J. AOAC Int. 2014, 97, 325–333. [Google Scholar] [CrossRef] [PubMed]
- Turner, A.D.; Powell, A.; Schofield, A.; Lees, D.N.; Baker-Austin, C. Detection of the pufferfish toxin tetrodotoxin in European bivalves, England, 2013 to 2014. Euro Surveill. 2015, 20. Available online: http://www.eurosurveillance.org/ViewArticle.aspx?ArticleId=21009 (accessed on 22 August 2015). [Google Scholar]
- Vlamis, A.; Katikou, P.; Rodriguez, I.; Rey, V.; Alfonso, A.; Papazachariou, A.; Zacharaki, T.; Botana, A.M.; Botana, L.M. First Detection of tetrodotoxin in Greek shellfish by UPLC-MS/MS potentially linked to the presence of the dinoflagellate Prorocentrum minimum. Toxins 2015, 7, 1779–1807. [Google Scholar] [CrossRef] [PubMed]
- Kao, C.Y.; Furman, F.A. Pharmacological studies on tetrodotoxin, a potent neurotoxin. J. Pharmacol. Exp. Ther. 1963, 140, 31–40. [Google Scholar] [PubMed]
- Moore, J.; Anderson, N.; Narahashi, T. Tetrodoixin blocking early conductance channel on sodium. Fed. Proc. Am. Soc. Exp. Biol. 1966, 25, 569–572. [Google Scholar]
- Yasumoto, T.; Yasumura, D.; Yotsu, M.; Michishita, T.; Endo, A.; Kotaki, Y. Bacterial production of tetrodotoxin and anhydrotetrodotoxin. Agric. Biol. Chem. 1986, 50, 793–795. [Google Scholar] [CrossRef]
- Noguchi, T.; Jeon, J.K.; Arakawa, O.; Sugita, H.; Deguchi, Y.; Shida, Y.; Hashimoto, K. Occurrence of tetrodotoxin and anhydrotetrodotoxin in Vibrio sp. isolated from the intestines of axantihd crab, Atergatis floridus. J. Biochem. 1986, 99, 311–314. [Google Scholar] [PubMed]
- Hanifin, C.T. The chemical and evolutionary ecology of tetrodotoxin (TTX) toxicity in terrestrial tertebrates. Mar. Drugs 2010, 8, 577–593. [Google Scholar] [CrossRef] [PubMed]
- Ritson-Williams, R.; Yotsu-Yamashita, M.; Paul, V. Ecological functions of tetrodotoxin in a deadly polyclad flatworm. Proc. Natl. Acad. Sci. USA 2006, 103, 3176–3179. [Google Scholar] [CrossRef] [PubMed]
- Yotsu-Yamashita, M. Chemistry of puffer fish toxin. J. Toxicol. Toxin Rev. 2001, 20, 51–66. [Google Scholar] [CrossRef]
- Yotsu-Yamashita, M.; Abe, Y.; Kudo, Y.; Ritson-Williams, R.; Paul, V.J.; Konoki, K.; Cho, Y.; Adachi, M.; Imazu, T.; Nishikawa, T.; et al. First identification of 5,11-dideoxytetrodotoxin in marine animals, and characterization of major fragment ions of tetrodotoxin and its analogs by high resolution ESI-MS/MS. Mar. Drugs 2013, 11, 2799–2813. [Google Scholar] [CrossRef] [PubMed]
- Kudo, Y.; Yamashita, Y.; Mebs, D.; Cho, Y.; Konoki, K.; Yasumoto, T.; Yotsu-Yamashita, M. C5–C10 directly bonded tetrodotoxin analogues: possible biosynthetic precursors of tetrodotoxin from newts. Angew. Chem. Int. Ed. 2014, 53, 14546–14549. [Google Scholar] [CrossRef] [PubMed]
- Yasumoto, T.; Yotsu, M.; Murata, M.; Naoki, H. New tetrodotoxin analogue from the newt Cynops ensicauda. J. Am. Chem. Soc. 1988, 110, 2344–2345. [Google Scholar] [CrossRef]
- Jang, J.H.; Yotsu-Yamashita, M. 6,11-DideoxyTTX from the puffer fish, Fugu pardalis. Toxicon 2007, 50, 947–951. [Google Scholar] [CrossRef] [PubMed]
- Yotsu-Yamashita, M.; Yamagishi, Y.; Yasumoto, T. 5,6,11-Trideoxytetrodotoxin from the puffer fish, Fugu poecilonotus. Tetrahedron Lett. 1995, 36, 9329–9332. [Google Scholar] [CrossRef]
- Khora, S.S.; Yasumoto, T. Isolation of 11-oxotetrodotoxin from the puffer Arothron nigropunctatus. Tetrahedron Lett. 1989, 30, 4393–4394. [Google Scholar] [CrossRef]
- Wu, B.Q.; Yang, L.; Kao, C.Y.; Levinson, S.R.; Yotsu-Yamashita, M.; Yasumoto, T. 11-Oxo-tetrodotoxin and a specifically labelled 3H-tetrodotoxin. Toxicon 1996, 34, 407–416. [Google Scholar] [CrossRef]
- Yotsu-Yamashita, M.; Sugimoto, A.; Takai, A.; Yasumoto, T. Effects of specific modifications of several hydroxyls of tetrodotoxin on its affinity to rat brain membrane. J. Pharmacol. Exp. Ther. 1999, 289, 1688–1696. [Google Scholar] [PubMed]
- Brillantes, S.; Samosorn, W.; Faknoi, S.; Oshima, Y. Toxicity of puffers landed and marketed in Thailand. Fish. Sci. 2003, 69, 1224–1230. [Google Scholar] [CrossRef]
- Islam, Q.T.; Razzak, M.A.; Islam, M.A.; Bari, M.I.; Basher, A.; Chowdhury, F.R.; Sayeduzzaman, A.B.M.; Ahasan, H.A.M.N.; Faiz, M.A.; Arakawa, O.; et al. Puffer fish poisoning in Bangladesh: clinical and toxicological results from large out breaks in 2008. Trans. R. Soc. Trop. Med. Hyg. 2011, 105, 74–80. [Google Scholar] [CrossRef] [PubMed]
- Wan, C.K.; Tsui, S.H.; Tong, H.K. A case series of puffer fish poisoning, Hong Kong. J. Emerg. Med. 2007, 14, 215–220. [Google Scholar]
- Yong, Y.S.; Quek, L.S.; Lim, E.K.; Ngo, A. A case report of puffer fish poisoning in Singapore. Case Rep. Med. 2013, 206971. [Google Scholar] [CrossRef] [PubMed]
- Noguchi, T.; Onuki, K.; Arakawa, O. Tetrodotoxin poisoning due to pufferfish and gastropods, and their intoxication mechanism. Toxicology 2011, 2011. [Google Scholar] [CrossRef] [PubMed]
- Bentur, Y.; Ashkar, J.; Lurie, Y.; Levy, Y.; Azzam, Z.S.; Litmanovich, M.; Golik, M.; Gurevych, B.; Golani, D.; Eisenman, A. Lessepsian migration and tetrodotoxin poisoning due to Lagocephalus sceleratus in the eastern Mediterranean. Toxicon 2008, 52, 964–968. [Google Scholar] [CrossRef]
- Kheifets, J.; Rozhavsky, B.; Girsh, S.Z.; Marianna, R.; Soroksky, A. Severe tetrodotoxin poisoning after consumption of Lagocephalus sceleratus (pufferfish, fugu) fished in Mediterranean Sea, treated with cholinesterase inhibitor. Case Rep. Crit. Care 2012, 2012, 782507. [Google Scholar]
- Cohen, N.J.; Deeds, J.R.; Wong, E.S.; Hanner, R.H.; Yancy, H.F.; White, K.D.; Thompson, T.M.; Wahl, M.; Pham, T.D.; Guichard, F.M.; et al. Public health response to puffer fish (tetrodotoxin) poisoning from mislabeled product. J. Food Prot. 2009, 72, 810–817. [Google Scholar] [PubMed]
- Cole, J.B.; Heegaard, W.G.; Deeds, J.R.; McGrath, S.C.; Handy, S.M. Tetrodotoxin poisoning outbreak from imported dried puffer fish—Minneapolis, Minnesota, 2014. Morbidity Mortality Wkly Rep. 2015, 63, 1222–1225. [Google Scholar]
- Loison, G. Poisonous fish of the South Pacific. Noumea, New Caledonia: South Pacific commission. SPC Q. Bull. 1955, 5, 28–31. [Google Scholar]
- Landsberg, J.H.; Hall, S.; Johannessen, J.N.; White, K.D.; Conrad, S.M.; Abbott, J.P.; Flewelling, L.J.; Richardson, R.W.; Dickey, R.W.; Jester, E.L.; et al. Saxitoxin puffer fish poisoning in the United States, with the first report of Pyrodinium bahamense as the putative toxin source. Environ. Health Perspect. 2006, 114, 1502–1507. [Google Scholar] [CrossRef] [PubMed]
- Nakashima, K.; Arakawa, O.; Taniyama, S.; Nonaka, M.; Takatani, T.; Yamamori, K.; Fuchi, Y.; Noguchi, T. Occurrence of saxitoxins as a major toxin in the ovary of a marine puffer Arothron firmamentum. Toxicon 2004, 43, 207–212. [Google Scholar] [CrossRef] [PubMed]
- Nakamura, M.; Oshima, Y.; Yasumoto, T. Occurrence of saxitoxin in puffer fish. Toxicon 1984, 22, 381–385. [Google Scholar] [CrossRef]
- Sato, S.; Kodama, M.; Ogata, T.; Saitanu, K.; Furuya, M.; Hirayama, K.; Kakinuma, K. Saxitoxin as a toxic principle of a freshwater puffer, Tetraodon fangi, in Thailand. Toxicon 1997, 35, 137–140. [Google Scholar] [CrossRef]
- HP of Ministry of Health, Labour and Welfare of Japan. Available online: http://www.mhlw.go.jp/topics/syokuchu/poison/animal_01.html (accessed on 22 August 2015). (In Japanese)
- Nakamura, M.; Yasumoto, T. Tetrodotoxin derivatives in puffer fish. Toxicon 1985, 23, 271–276. [Google Scholar] [CrossRef]
- Yasumoto, T.; Michishita, T. Fluorometric determination of tetrodotoxin by high performance liquid chromatography. Agric. Biol. Chem. 1985, 49, 3077–3080. [Google Scholar] [CrossRef]
- Shoji, Y.; Yotsu-Yamashita, M.; Miyazawa, T.; Yasumoto, T. Electrospray ionization mass spectrometry of tetrodotoxin and its analogs: Liquid Chromatography/ Mass Spectrometry, Tandem Mass Spectrometry, and Liquid Chromatography/Tandem Mass Spectrometry. Anal. Biochem. 2001, 290, 10–17. [Google Scholar] [CrossRef] [PubMed]
- Nakagawa, T.; Jang, J.; Yotsu-Yamashita, M. Hydrophilic interaction liquid chromatography-electrospray ionization mass spectrometry of tetrodotoxin and its analogs. Anal. Biochem. 2006, 352, 142–144. [Google Scholar] [CrossRef] [PubMed]
- Jang, J.; Lee, J.S.; Yotsu-Yamashita, M. LC/MS analysis of tetrodotoxin and its deoxy analogs in the marine puffer fish Fugu niphobles from the southern coast of Korea, and in the brackish water puffer fishes Tetraodon nigroviridis and Tetraodon biocellatus from Southeast Asia. Mar. Drugs 2010, 8, 1049–1058. [Google Scholar] [CrossRef] [PubMed]
- Yotsu-Yamashita, M.; Jang, J.H.; Cho, Y.; Konoki, K. Optimization of simultaneous analysis of tetrodotoxin, 4-epitetrodotoxin, 4,9-anhydrotetrodotoxin, and 5,6,11-trideoxytetrodotoxin by hydrophilic interaction liquid chromatography–tandem mass spectrometry. Forensic Toxicol. 2011, 29, 61–64. [Google Scholar] [CrossRef]
- Kudo, Y.; Yasumoto, T.; Konoki, K.; Cho, Y.; Yotsu-Yamashita, M. Isolation and structural determination of the first 8-epi-type tetrodotoxin analogs from the newt, Cynops ensicauda popei, and comparison of tetrodotoxin analogs profiles of this newt and the puffer fish, Fugu poecilonotus. Mar. Drugs 2012, 10, 655–667. [Google Scholar] [CrossRef] [PubMed]
- Khora, S.S.; Isa, J.; Yasumoto, T. Toxicity of puffers from Okinawa, Japan. Nippon Suisan Gakkaishi 1991, 57, 163–167. [Google Scholar] [CrossRef]
- Teruya, N.; Oshiro, N.; Tamanaha, K. Toxicity of puffers from Okinawa. Available online: http://www.pref.okinawa.jp/site/hoken/eiken/syoho/documents/s40_93-98.pdf (accessed on 24 August 2015). (In Japanese)
- Campbell, S.; Harada, R.M.; DeFelice, S.V.; Bienfang, P.K.; Li, Q.X. Bacterial production of tetrodotoxin in the pufferfish Arothron hispidus. Nat. Product Res. Part A 2009, 17, 1630–1640. [Google Scholar] [CrossRef] [PubMed]
- Jang, J.; Yotsu-Yamashita, M. Distribution of tetrodotoxin, saxitoxin, and their analogs among tissues of the puffer fish Fugu pardalis. Toxicon 2006, 48, 980–987. [Google Scholar] [CrossRef] [PubMed]
- Yotsu-Yamashita, M.; Sugimoto, A.; Terakawa, T.; Shoji, Y.; Miyazawa, T.; Yasumoto, T. Purification, characterization, and cDNA cloning of a novel soluble saxitoxin and tetrodotoxin binding protein from plasma of the pufferfish, Fugu pardalis. Eur. J. Biochem. 2001, 268, 5937–5946. [Google Scholar] [CrossRef] [PubMed]
- Yotsu-Yamashita, M.; Okoshi, N.; Watanabe, K.; Araki, N.; Yamaki, H.; Shoji, Y.; Terakawa, T. Localization of pufferfish saxitoxin and tetrodotoxin binding protein (PSTBP) in the tissues of the pufferfish, Takifugu pardalis, analyzed by immunohistochemical staining. Toxicon 2013, 72, 23–28. [Google Scholar] [CrossRef] [PubMed]
- Hashiguchi, Y.; Lee, J.M.; Shiraishik, M.; Komatsu, S.; Miki, S.; Shimasaki, Y.; Mochioka, N.; Kusakabe, T.; Oshima, Y. Characterization and evolutionary analysis of tributyltin-binding protein and pufferfish saxitoxin and tetrodotoxin-binding protein genes in toxic anc nontoxic pufferfish. J. Evol. Biol. 2015. [Google Scholar] [CrossRef] [PubMed]
- Rodriguez, P.; Alfonso, A.; Vale, C.; Alfonso, C.; Vale, P.; Tellez, A.; Botana, L.M. First toxicity report of tetrodotoxin and 5,6,11-trideoxyTTX in the trumpet shell Charonia lampas lampas in Europe. Anal. Chem. 2008, 80, 5622–5629. [Google Scholar] [CrossRef] [PubMed]
- Pires, O.R.; Sebbena, A.; Schwartza, E.F.; Bloch, C.J.; Rodrigo, A.V.; Morales, R.A.V.; Schwartza, C.A. The occurrence of 11-oxotetrodotoxin, a rare tetrodotoxin analogue, in the brachycephalidae frog Brachycephalus ephippium. Toxicon 2003, 42, 563–566. [Google Scholar] [CrossRef]
- Yotsu-Yamashita, M.; Mebs, D. Occurrence of 11-oxotetrodotoxin in the red-spotted newt, Notophthalmus viridescens, and further studies on the levels of tetrodotoxin and its analogues in the newt efts. Toxicon 2003, 41, 893–897. [Google Scholar] [CrossRef]
- Kudo, Y.; Finn, J.; Fukushima, K.; Sakugawa, S.; Cho, Y.; Konoki, K.; Yotsu-Yamashita, M. Isolation of 6-deoxytetrodotoxin from the pufferfish, Takifugu pardalis, and a comparison of the effects of the C-6 and C-11 hydroxy groups of tetrodotoxin on its activity. J. Nat. Prod. 2014, 77, 1000–1004. [Google Scholar] [CrossRef] [PubMed]
- Taniyama, S.; Isami, Y.; Matsumoto, T.; Nagashima, Y.; Takatani, T.; Arakawa, O. Toxicity and toxin profile of tetrodotoxin detected in the scavenging gastropod Nassarius (Alectrion) glans. “Kinshibai”. Shokuhin Eiseigaku Zasshi 2009, 50, 22–28. (In Japanese) [Google Scholar] [CrossRef] [PubMed]
- Arakawa, O.; Noguchi, T.; Shida, Y.; Onoue, Y. Occurrence of 11-oxotetrodotoxin and 11-nortetrodotoxin-6(R)-ol in a xanthid crab Atergatis floridus collected at Kojima, Ishigaki Island. Fish. Sci. 1994, 60, 769–771. [Google Scholar]
- Codex Alimentarius, CODEX STAN 292-2008. Available online: http://www.codexalimentarius.org/input/download/standards/11109/CXS_292e.pdf (accessed on 22 August 2015).
- EU Regulation 853/2004. Regulation (EC) no. 853/2004 of the European Parliament and of the Council of 29 April 2004. Laying down specific hygiene rules for food of animal origin. Off. J. Eur. Commun. 2004, L226, 22–82. [Google Scholar]
- U.S. Food and Drug Administration. National Shellfish Sanitation Program Guide for the Control of Molluscan Shellfish 2011 Revision. Available online: http://www.fda.gov/downloads/Food/GuidanceRegulation/FederalStateFoodPrograms/UCM350344.pdf (accessed on 22 August 2015).
- Veterinary Sanitation Division Director-General. Notification No. 30 of Veterinary Sanitation Division, from the Veterinary Sanitation Division Director-General, Veterinary Sanitation Division, Environmental Health Bureau, Ministry of Health, Labour and Welfare, 1980. Available online: http://www.mhlw.go.jp/topics/syokuchu/kanren/kanshi/s58_1202-2.html (accessed on 22 August 2015). (In Japanese)
- Oshima, Y. Postcolumn derivatization liquid chromatographic method for paralytic shellfish toxins. J. AOAC Int. 1995, 78, 528–532. [Google Scholar]
- Dell’Aversano, C.; Hess, P.; Quilliam, M.A. Hydrophilic interaction liquid chromatography-mass spectrometry for the analysis of paralytic shellfish poisoning (PSP) toxins. J. Chromatogr. A 2005, 1081, 190–201. [Google Scholar] [CrossRef] [PubMed]
© 2015 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/4.0/).