
Review on Mycotoxin Issues in Ruminants: Occurrence in Forages, Effects of Mycotoxin Ingestion on Health Status and Animal Performance and Practical Strategies to Counteract Their Negative Effects

 ,
,  ,
,  ,
, 
Abstract
:1. Introduction

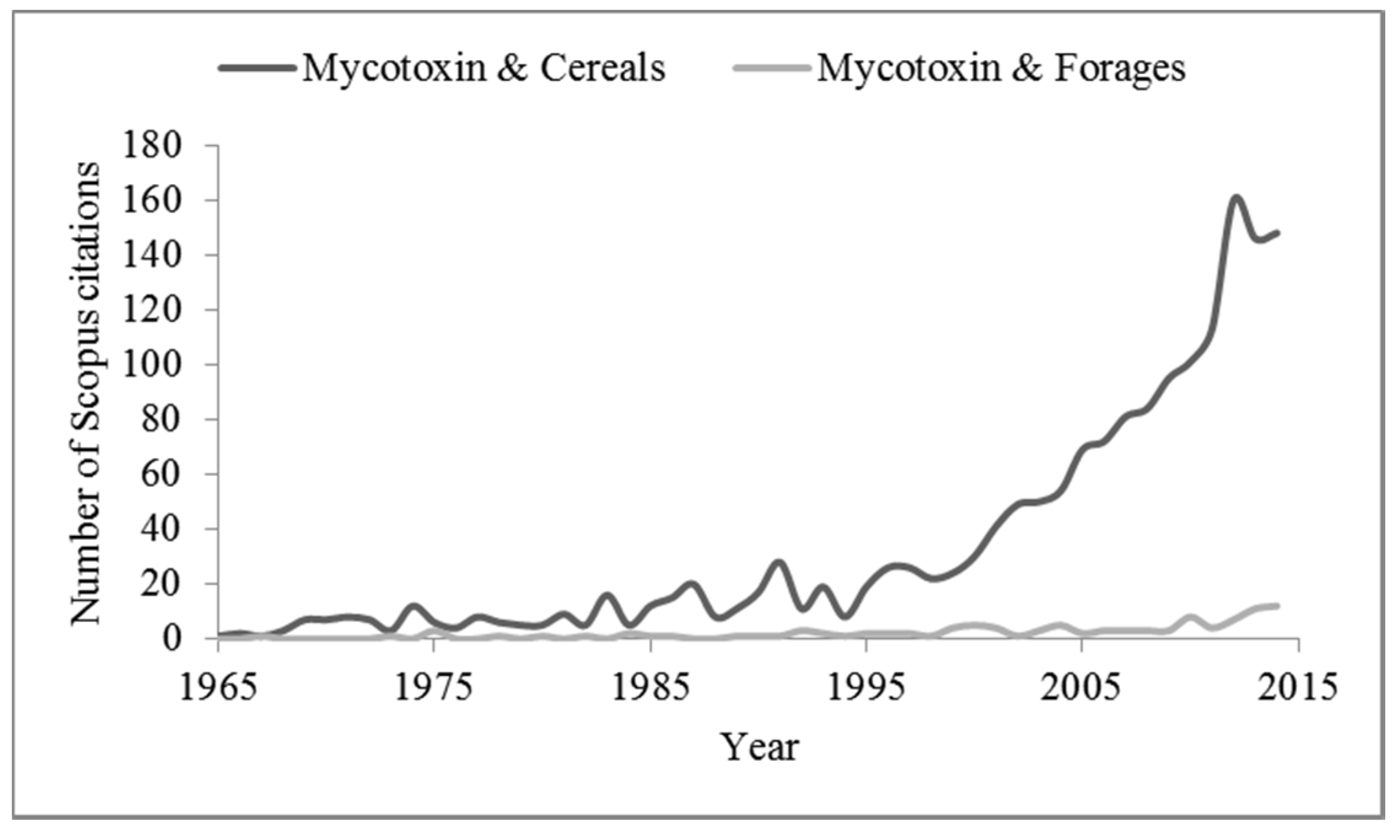{kind=link}
| Secondary Metabolites | Scopus Citation | Scientific Interest a | Secondary Metabolites | Scopus Citation | Scientific Interest a |
|---|---|---|---|---|---|
| AAL toxin | 100 | ** | Infectopyrones | 3 | * |
| Aflatoxins | 16,939 | ***** | Islanditoxin | 10 | * |
| Aflavinine | 12 | * | Luteoskyrin | 135 | ** |
| Agroclavine | 214 | *** | Marcfortine A, B and C | 38 | * |
| Alternariol | 396 | **** | Monacolins | 242 | *** |
| Andrastins | 30 | * | Moniliformin | 399 | **** |
| Aspergillic acid | 66 | * | Monoacetoxyscirpenol | 64 | * |
| Aurofusarin | 55 | * | Mycophenolic acid | 241 | *** |
| Beauvericin | 441 | **** | Neosolaniol | 242 | *** |
| β-nitropropionic acids | 4 | * | Nivalenol | 1014 | ***** |
| Botryodiploidin | 36 | * | Novae-zelandins | 1 | * |
| Butenolide | 1337 | ***** | Ochratoxins | 5162 | ***** |
| Byssochlamic acid | 31 | * | Oosporein | 45 | * |
| Chlamydosporol | 21 | * | Orsellinic acid | 205 | *** |
| Chrysogine | 18 | * | Paspalitrems | 7 | * |
| Citreoviridin | 124 | ** | Patulin | 1606 | ***** |
| Citrinin | 1994 | ***** | Penicillic acid | 437 | **** |
| Citreoisocoumarin | 9 | * | Penitrem | 202 | *** |
| Clavine alkaloids | 146 | ** | Phomopsin | 123 | ** |
| Culmorin | 33 | * | PR toxin | 320 | **** |
| Cyclopiazonic Acid | 2307 | ***** | PR-amide | 6 | * |
| Deoxynivalenol | 3720 | ***** | PR-imine | 5 | * |
| Diacetoxyscirpenol | 759 | **** | Pseurotins | 56 | * |
| Dicoumarol | 3811 | ***** | Roquefortines | 213 | *** |
| Diketopioperazines | 1 | * | Roridins | 32 | * |
| Eremofortin C | 10 | * | Rubratoxin | 191 | ** |
| Ergot toxins | 7567 | ***** | Rubrofusarin | 75 | * |
| Ergotamine | 7298 | ***** | Scirpentriol | 69 | * |
| Festuclavine | 74 | * | Slaframine | 103 | ** |
| Fumagillin | 939 | **** | Sphingofungin | 47 | * |
| Fumigatins | 23 | * | Sporidesmin | 207 | *** |
| Fumiquinazolines | 56 | * | Stachbotryotoxins | 1 | * |
| Fumitremorgen | 11 | * | Sterigmatocystin | 1000 | **** |
| Fumitremorgines | 357 | **** | T-2 & HT-2 toxin | 470 | **** |
| Fumonisins | 3542 | ***** | Tentoxin | 208 | *** |
| Fusarenone-X | 54 | * | Tenuazonic acid | 256 | *** |
| Fusaric Acid | 675 | **** | Tremorgens | 37 | * |
| Fusarins | 100 | ** | Tremorgens | 46 | * |
| Fusariocin | 2 | * | Trypacidin | 20 | * |
| Gliotoxin | 996 | **** | Verruculogen | 112 | ** |
| Helvolic acid | 89 | ** | Zearalenone | 3443 | ***** |

2. Mycotoxin Occurrence in Animal Feeds, with Special Emphasis on Their Presence in Forages
2.1. Alternaria Toxins in Forages
| Forage Products | Mycotoxins a | Number of Samples | Incidence (%) | Mean (Excluding not Detectable Data when Possible) | Range or Maximal Detected Value | Nation | References | Notes |
|---|---|---|---|---|---|---|---|---|
| Alternaria spp. derived toxin | ||||||||
| Different feeds | AAL TA toxin | 63 | 97% | 560 μg/kg | 90–1470 μg/kg | WI, US | [75] | |
| MS | AAL TA toxin | 60 | ~30% | 170 μg/kg | 200–2010 μg/kg | PA, US | [76] | |
| Hay and hay silage | AAL TA toxin | 25 | 100% | 720 μg/kg | 290–1160 μg/kg | WI, US | [75] | |
| MS | AAL TB toxin | 60 | ~15% | 50 μg/kg | 30–900 μg/kg | PA, US | [76] | |
| MS | Alternariol | 82 | 2% | 18 μg/kg | max 24 μg/kg | Denmark | [24] | |
| MS | Alternariol ME | 82 | 2% | 8 μg/kg | max 8.8 μg/kg | Denmark | [24] | |
| Aspergillus flavus and A. parasiticus derived toxin | ||||||||
| MS | AFB1 | 1 | - | 28 μg/kg | France | [77] | ||
| MS | AFB1 | 100 | 92% | - | 0.6– > 4 μg/kg | Italy | [78] | only core samples |
| MS | AFB1 | 116 | 13% | 33 μg/kg | 2–54 μg/kg | Brazil | [62] | core samples |
| MS | AFB1 | 9 | - | - | 4–34 μg/kg | France | [79] | from 1 farm |
| Silages | β-nitropropionic acid | 3 | 33% | 1360 μg/kg | - | Netherlands | [16] | |
| Various Aspergillus and Penicillium spp. derived toxin | ||||||||
| Different feeds | Cyclopiazonic acid | 63 | 87% | 340 μg/kg | 120–1820 μg/kg | WI, US | [75] | |
| Hay and hay silage | Cyclopiazonic acid | 25 | 80% | 390 μg/kg | 120–1820 μg/kg | WI, US | [75] | |
| MS | Cyclopiazonic acid | 120 | 37% | 120 μg/kg | 20–1430 μg/kg | PA, US | [80] | 4 samples from 30 bunkers |
| Silages | Cyclopiazonic acid | 3 | 33% | 55 μg/kg | - | Netherlands | [16] | |
| Aspergillus fumigatus derived toxin | ||||||||
| MS | Gliotoxin | 1 | - | 4 μg/kg | France | [77] | ||
| MS | Gliotoxin | 90 | - | 5130 μg/kg | 5100–6500 μg/kg | Argentina | [81] | |
| Silages | Gliotoxin | 3 | 33% | 1870 μg/kg | - | Netherlands | [16] | |
| MS | Gliotoxin | 196 | <1% | 140 μg/kg | max 600 μg/kg | Italy | [69] | 3 samples from 68 silos |
| Fusarium spp. derived toxin Trichothecenes type A | ||||||||
| MS | 15-acetyl DON | 140 | <1% | 901 μg/kg | max 1013 μg/kg | Netherlands | [82] | over three years |
| MS | 15-acetyl DON | 5 | 100% | 59 μg/kg | max 127 μg/kg | Germany | [83] | |
| MS | 3-acetyl DON | 20 | 0% | - | - | Denmark | [84] | |
| Hays | 3-acetyl DON | 28 | 4% | 20 μg/kg | - | Germany | [83] | |
| MS | 3-&5-acetyl DON | 19 | 21% | 217 μg/kg | 135–300 μg/kg | Switzerland | [85] | |
| Different feeds | DON | 63 | 100% | 730 μg/kg | 340–6020 μg/kg | WI, US | [75] | |
| Compound feed | DON | 72 | 54% | 433 μg/kg | max 2408 μg/kg | Netherlands | [36] | |
| MS | DON | 20 | 100% | 1056 μg/kg | 160–5094 μg/kg | Denmark | [84] | |
| MS | DON | 140 | 72% | 854 μg/kg | max 3142 μg/kg | Netherlands | [82] | over three years |
| MS | DON | 82 | 6% | 1629 μg/kg | max 2974 μg/kg | Denmark | [24] | Quantitative analysis |
| MS | DON | 1 | - | 146 μg/kg | - | France | [77] | |
| MS | DON | 196 | 8% | 280 μg/kg | max 560 μg/kg | Italy | [69] | 3 samples from 68 silos |
| MS | DON | 9 | - | - | 100–213 μg/kg | France | [79] | from 1 farm |
| MS | DON | 5 | 100% | 2919 μg/kg | max 3944 μg/kg | Germany | [83] | |
| MS | DON | 19 | 100% | 1356 μg/kg | 780–2990 μg/kg | Switzerland | [85] | |
| MS | DON | 116 | 24% | 1610 μg/kg | 150–3420 μg/kg | Brazil | [62] | core samples |
| Silages | DON | 3 | 100% | 396 μg/kg | max 761 μg/kg | Netherlands | [16] | |
| Ensiled by-products | DON | 29 | 0% | - | - | Netherlands | [36] | |
| Feed commodities | DON | 8 | 38% | 1019 μg/kg | max 1811 μg/kg | Netherlands | [36] | |
| Forage products | DON | 13 | 15% | 348 μg/kg | max 489 μg/kg | Netherlands | [36] | |
| Hay and hay silage | DON | 25 | 100% | 610 μg/kg | 510–720 μg/kg | WI, US | [75] | |
| Hays | DON | 28 | 14% | 41 μg/kg | max 69 μg/kg | Germany | [83] | |
| Silage | DON | 47 | 53% | 550 μg/kg | max 1250 μg/kg | Netherlands | [36] | |
| SGS (Wheat) | DON | 30 | 10% | 621 μg/kg | max 1165 μg/kg | Netherlands | [36] | over two years |
| MS | DON 2000 | 196 | 59% | 1290 μg/kg DM | 240–12,890 μg/kg DM | Germany | [86] | ELISA method |
| MS | DON 2002 | 182 | 89% | 2100 μg/kg DM | 260–14,290 μg/kg DM | Germany | [86] | ELISA method |
| MS | DON 2001 | 32 | 86% | 800 μg/kg | max 3700 μg/kg | PA, US | [87] | over two years |
| MS | DON 2002 | 39 | 66% | 1100 μg/kg | max 5100 μg/kg | PA, US | [87] | over two years |
| MS | Fusarenon X | 20 | 20% | 4 μg/kg | 8–14 μg/kg | Denmark | [84] | |
| MS | Nivalenol | 5 | 100% | 1612 μg/kg | max 2809 μg/kg | Germany | [83] | |
| MS | Nivalenol | 19 | 42% | 521 μg/kg | 190–760 μg/kg | Switzerland | [85] | |
| Hays | Nivalenol | 28 | 4% | 131 μg/kg | max 222 μg/kg | Germany | [83] | |
| MS | Nivalenol | 82 | 13% | 266 μg/kg | max 758 μg/kg | Denmark | [24] | Quantitative analysis |
| Fusarium spp. derived toxin: Trichothecenes type B | ||||||||
| MS | 15monoacetoxyscirpenol | 5 | 60% | 30 μg/kg | max 49 μg/kg | Germany | [83] | |
| MS | HT-2 toxin | 20 | 60% | 104 μg/kg | 2–327 μg/kg | Denmark | [84] | |
| MS | HT-2 toxin | 5 | 100% | 18 μg/kg | max 26 μg/kg | Germany | [83] | |
| MS | HT-2 toxin | 19 | 26% | 95 μg/kg | 76–120 μg/kg | Switzerland | [85] | |
| MS | T-2 toxin | 20 | 5% | 2 μg/kg | - | Denmark | [84] | |
| MS | T-2 toxin | 19 | 42% | 36 μg/kg | 14–84 μg/kg | Switzerland | [85] | |
| Fusarium spp. derived toxin: Fumonisins | ||||||||
| Different feeds | FB1 | 63 | 37% | 280 μg/kg | 20–2120 μg/kg | WI, US | [75] | |
| MS | FB1 | 140 | 1% | 17,000 μg/kg | max 26,200 μg/kg | Netherlands | [82] | over three years |
| MS | FB1 | 86 | 97% | 615 μg/kg | 21–1824 μg/kg | IL, US | [88] | |
| MS | FB1 | 60 | ~75% | 2020 μg/kg | 200–10,100 μg/kg | PA, US | [76] | |
| MS | FB1 | 116 | 15% | 5440 μg/kg | 300–3400 μg/kg | Brazil | [62] | core samples |
| MS | FB1 | 100 | 88% | - | 900– > 10,000 μg/kg | Italy | [78] | only core samples |
| Hay and hay silage | FB1 | 25 | 52% | 120 μg/kg | 20–450 μg/kg | WI, US | [75] | |
| Silages | FB1 | 3 | 33% | 21 μg/kg | - | Netherlands | [16] | |
| MS | FB2 | 64 | 72% | 93 μg/kg | 21–276 μg/kg | IL, US | [88] | |
| MS | FB2 | 60 | ~40% | 980 μg/kg | 200–20,300 μg/kg | PA, US | [76] | |
| MS | FB3 | 51 | 57% | 51 μg/kg | 16–161 μg/kg | IL, US | [88] | |
| Fusarium spp. derived toxin: other Fusarium toxins | ||||||||
| GS (bunkers) | Beauvericin | 88 | - | ~30 μg/kg DM | - | Ireland | [89] | |
| GS (round bale) | Beauvericin | 56 | - | ~30 μg/kg DM | - | Ireland | [89] | |
| MS | Enniatin A1 | 6 | - | ~120 μg/kg DM | - | Ireland | [89] | |
| GS (bunkers) | Enniatin A1 | 88 | - | ~40 μg/kg DM | - | Ireland | [89] | |
| GS (bunkers) | Enniatin A1 | 88 | - | ~20 μg/kg DM | - | Ireland | [89] | |
| GS (round bale) | Enniatin A1 | 56 | - | ~25 μg/kg DM | - | Ireland | [89] | |
| MS | Enniatin B | 82 | 24% | 53 μg/kg | max 152 μg/kg | Denmark | [24] | |
| GS (bunkers) | Enniatin B | 88 | - | ~60 μg/kg DM | - | Ireland | [89] | |
| GS (round bale) | Enniatin B | 56 | - | ~250 μg/kg DM | - | Ireland | [89] | |
| MS | Enniatin B1 | 6 | - | ~160 μg/kg DM | - | Ireland | [89] | |
| GS (bunkers) | Enniatin B1 | 88 | - | ~180 μg/kg DM | - | Ireland | [89] | |
| GS (round bale) | Enniatin B1 | 56 | - | ~80 μg/kg DM | - | Ireland | [89] | |
| Fusarium spp. derived toxin: Zearalenone | ||||||||
| MS | α-ZOL | 5 | 20% | 15 μg/kg | - | Germany | [83] | |
| MS | β-ZOL | 5 | 20% | 116 μg/kg | - | Germany | [83] | |
| Different feeds | ZEA | 63 | 32% | 220 μg/kg | 120–310 μg/kg | WI, US | [75] | |
| Compound feed | ZEA | 28% | 80 μg/kg | max 363 μg/kg | Netherlands | [36] | ||
| MS | ZEA | 140 | 49% | 174 μg/kg | max 943 μg/kg | Netherlands | [82] | over three years |
| MS | ZEA | 82 | 28% | 66 μg/kg | max 311 μg/kg | Denmark | [24] | Quantitative analysis |
| MS | ZEA | 9 | - | - | 23–41 μg/kg | France | [79] | from 1 farm |
| MS | ZEA | 5 | 100% | 432 μg/kg | max 1790 μg/kg | Germany | [83] | |
| MS | ZEA | 19 | 79% | 180 μg/kg | 83–430 μg/kg | Switzerland | [85] | |
| MS | ZEA | 85 | 15% | - | >50 μg/kg | Italy | [90] | |
| MS | ZEA | 100 | 60% | - | 30–>300 μg/kg | Italy | [78] | only core samples |
| Silages | ZEA | 3 | 100% | 145 μg/kg | max 240 μg/kg | Netherlands | [16] | |
| Ensiled by-products | ZEA | - | - | - | Netherlands | [36] | ||
| Feed commodities | ZEA | 38% | 80 μg/kg | max 108 μg/kg | Netherlands | [36] | ||
| Forage products | ZEA | 8% | 82 μg/kg | - | Netherlands | [36] | ||
| GS | ZEA | 120 | 6% | 936 μg/kg | max 308 μg/kg | Netherlands | [82] | over three years |
| Hay and hay silage | ZEA | 25 | 0% | - | - | WI, US | [75] | |
| Hays | ZEA | 28 | 43% | 24 μg/kg | max 115 μg/kg | Germany | [83] | |
| Hays | ZEA | 44 | 21% | - | - | Ireland | [91] | |
| Haylages | ZEA | 40 | 8% | - | - | Ireland | [91] | |
| Hays | ZEA | 65 | 8% | - | - | Canada | [91] | |
| Silage | ZEA | 17% | 125 μg/kg | max 273 μg/kg | Netherlands | [36] | ||
| Penicillium spp. derived toxin | ||||||||
| Different feeds | PR toxin | 63 | 76% | 130 μg/kg | 50–260 μg/kg | WI, US | [75] | |
| Hay and hay silage | PR toxin | 25 | 80% | 15 μg/kg | 50–260 μg/kg | WI, US | [75] | |
| GS (round bale) | 16-OH-roquefortine C | 5 | 20% | - | range 100–1000 μg/kg | Ireland | [10] | |
| MS | Andrastin A | 82 | 18% | 169 μg/kg | max 691 μg/kg | Denmark | [24] | Quantitative analysis |
| GS (round bale) | Andrastin A | 56 | - | ~500 μg/kg DM | - | Ireland | [89] | |
| GS (round bale) | Andrastin A | 5 | 100% | - | range trace-20,000 μg/kg | Ireland | [10] | |
| MS | Citreoisocoumarin | 82 | 8% | - | - | Denmark | [24] | Qualitative analysis |
| GS (round bale) | Citreoisocuomarin | 5 | 40% | - | trace | Ireland | [10] | |
| MS | Marcfortine A | 82 | 7% | - | - | Denmark | [24] | Qualitative analysis |
| GS (round bale) | Marcfortine A | 5 | 60% | - | range 100–1000 μg/kg | Ireland | [10] | |
| MS | Marcfortine B | 82 | 1% | Denmark | [24] | Qualitative analysis | ||
| GS (round bale) | Agroclavine | 5 | 40% | - | range 100–1000 μg/kg | Ireland | [10] | from A. fumigatus too [92] |
| GS (round bale) | Festuclavine | 5 | 40% | - | range 100–1000 μg/kg | Ireland | [10] | from A. fumigatus too [92] |
| MS | Mycophenolic Acid | 135 | 28% | 690 μg/kg | 20–23,000 μg/kg | Germany | [93] | |
| MS | Mycophenolic acid | 120 | 42% | 160 μg/kg | 20–1300 μg/kg | PA, US | [80] | 4 samples from 30 bunkers |
| MS | Mycophenolic acid | 82 | 2% | 8 μg/kg | max 8.8 μg/kg | Denmark | [24] | Quantitative analysis |
| MS | Mycophenolic Acid | 196 | 8% | 1760 μg/kg | max 48,000 μg/kg | Italy | [69] | Three samples from 68 silos |
| Silages | Mycophenolic Acid | 3 | 100% | 4244 μg/kg | max 7565 μg/kg | Netherlands | [16] | |
| Ensiled by-products | Mycophenolic acid | 10% | 66 μg/kg | max 83 μg/kg | Netherlands | [36] | ||
| GS (bunkers) | Mycophenolic Acid | 88 | - | ~250 μg/kg DM | - | Ireland | [89] | |
| GS (round bale) | Mycophenolic Acid | 56 | - | ~1250 μg/kg DM | - | Ireland | [89] | |
| GS | Mycophenolic Acid | 98 | 37% | 2200 μg/kg | 20–35,000 μg/kg | Germany | [93] | |
| GS (round bale) | Mycophenolic acid | 5 | 100% | - | range trace-20,000 μg/kg | Ireland | [10] | |
| Silage | Mycophenolic acid | 13% | 524 μg/kg | max 2630 μg/kg | Netherlands | [36] | ||
| MS | Roquefortine C | 12 | 8% | 200 μg/kg DM | - | Germany | [94] | molded silages |
| MS | Roquefortine C | 12 | 100% | 17,000 μg/kg DM | 700–36,000 μg/kg DM | Germany | [94] | unmolded samples |
| MS | Roquefortine C | 60 | 30% | 5470 μg/kg | 50–28,000 μg/kg DM | Germany | [95] | data of Armbruster, 1994 |
| MS | Roquefortine C | 120 | 60% | 380 μg/kg | 10–5710 μg/kg | PA, US | [80] | 4 samples from 30 bunkers |
| MS | Roquefortine C | 82 | 2% | 173 μg/kg | max 189 μg/kg | Denmark | [24] | Quantitative analysis |
| MS | Roquefortine C | 196 | 5% | 740 μg/kg | max 32,000 μg/kg | Italy | [69] | 3 samples from 68 silos |
| Ensiled by-products | Roquefortine C | 7% | 123 μg/kg | max 170 μg/kg | Netherlands | [36] | ||
| GS (bunkers) | Roquefortine C | 88 | - | ~500 μg/kg DM | - | Ireland | [89] | |
| GS (round bale) | Roquefortine C | 56 | - | ~280 μg/kg DM | - | Ireland | [89] | |
| GS | Roquefortine C | 24 | 13% | - | range 10–580 μg/kg | Germany | [10] | From Ambruster, 2008 PhD thesis |
| GS | Roquefortine C | 20 | 15% | 280 μg/kg | range 10–580 μg/kg | Germany | [95] | From Ambruster, 2008 PhD thesis |
| GS | Roquefortine C | 120 | <1% | 81 μg/kg | - | Netherlands | [82] | over three years |
| GS (round bale) | Roquefortine C | 5 | 40% | - | range 1000–20,000 μg/kg | Ireland | [10] | |
| Silage | Roquefortine C | 19% | 778 μg/kg | max 3160 μg/kg | Netherlands | [36] | ||
| GS (wilted) | Roquefortine C | 12 | 75% | 200 μg/kg DM | 100–300 μg/kg DM | Germany | [94] | molded silages |
| GS (wilted) | Roquefortine C | 12 | 42% | 600 μg/kg DM | 200–15,000 μg/kg DM | Germany | [94] | unmolded samples |
| MS | Roquefortine A | 82 | 11% | - | - | Denmark | [24] | Qualitative analysis |
| GS (round bale) | Roquefortine A | 5 | 40% | - | range 100–1000 μg/kg | Ireland | [10] | |
| GS (round bale) | Roquefortine B | 5 | 40% | - | range 100–1000 μg/kg | Ireland | [10] | |
| GS (round bale) | Roquefortine D | 5 | 40% | - | range 100–1000 μg/kg | Ireland | [10] | |
| MS | Patulin | 120 | 23% | 80 μg/kg | 10–1210 μg/kg | PA, US | [80] | 4 samples from 30 bunkers |
| Silages | Patulin | 3 | 100% | 153 μg/kg | max 211 μg/kg | Netherlands | [16] | |
| Monascus ruber derived toxin | ||||||||
| Silages | Monacolin KB | 233 | 21% | 6161 μg/kg | 28–65,400 μg/kg | Germany | [96] | |
| Silages | Monacolin KL | 233 | 19% | 1767 μg/kg | 25–15,600 μg/kg | Germany | [96] | |
| MS | Citrinin | 1 | - | 12 μg/kg | France | [77] | ||
| MS | Citrinin | 9 | 4–25 μg/kg | France | [79] | from 1 farm | ||
| Silages | Citrinin | 233 | 6% | 9 μg/kg | 2–64 μg/kg | Germany | [96] | |
| Forage Products | Mycotoxins a not Detected | References |
|---|---|---|
| MS | AFB1, AFB2, AFG1, AFG2, 3-acetyl-DON, DAS, ergotamin, FB2, fusarenon-X, OTA, mycophenolic acid, penicillic acid, roquefortin C, sterigmatocystin, T-2 toxin, HT-2 toxin | [82] |
| MS | AFB1, AFB2, AFG1, AFG2, OTA, T-2 toxin, HT-2 toxin, 3-acetyl-DON, 15-acetyl-DON, DAS, sterigmatocystin, fusarenon-X, ergotamine, penicillic acid | [36] |
| MS | Cyclopiazonic acid, fumitremorgin A, gliotoxin, OTA, patulin, penitrem A, sterigmatocystin, T-2 toxin, tenuazonic acid, altersetin, fumigaclavine A, fumigaclavine C, PR toxin | [24] |
| MS | ZEA, PR toxin | [69] |
| MS | 3-acetyldeoxynivalenol, DAS, fusarenon-X, T-2 toxin, HT-2 toxin, neosolaniol, scirpentriol | [83] |
| Hays | 15-monoacetoxyscirpenol, 15-acetyldeoxynivealenol, DAS, fusarenon-X, T-2 toxin, neosolaniol, scirpentriol, α-ZOL, β-ZOL | [83] |
| MS | FB1, OTA, ZEA | [77] |
| MS | Gliotoxin, OTA | [79] |
| Hays and haylages | FBs, AFs, T-2 toxin, OTA | [91] |
2.2. Aspergillus Toxins in Forages
2.3. Fusarium Toxins in Forages
2.4. Penicillium Toxins in Forages
2.5. Monascus Ruber Toxins in Forages
2.6. Zygomycetes Fungi in Forages
3. Effect of Mycotoxins Ingestion on Ruminants: In Vitro and in Vivo Experiences
| Mycotoxins a | Media | Tested Dosages | Effects | References |
|---|---|---|---|---|
| AFB1 | rumen fluid | 0, 300, 600, 900 ng AFB1/mL buffered rumen fluid | ↓ gas production, ↓ dry matter digestibility, ↓ NH3-N concentrations | [141] |
| AFB1 | rumen fluid | 1, 10 μg AFB1/mL buffered rumen fluid | ↓ dry matter digestibility | [142] |
| AFB1 | rumen fluid | 9.5 ng AFB1/mL buffered rumen fluid | no effects | [143] |
| AFB1 | rumen fluid | 0, 320, 640, 960 ng AFB1/mL buffered rumen fluid | ↓ final gas production, ↓ rate of degradation, ↓ NH3-N concentrations, ↑ isobutyrate, valerate and isovalerate molar proportions | [144] |
| DON | rumen fluid | 0.36/0.46 or 5.76/6.90 mg of DON/kg diet | None, expect ↓ NDF digestibility | [145] |
| DON | rumen fluid | 0.3 or 3.4/4.4 mg of DON/kg diet | None, expect ↓ NDF digestibility | [146] |
| DON | rumen fluid | 40 μg DON/mL of rumen fluid | ↓ gas production, ↓ VFA and NH3-N concentrations | [147] |
| DON and fusaric acid | culture media | antimicrobial activity of fusaric acid against Ruminococcus albus and Methanobrevibacter ruminantium. No effect of DON | [148] | |
| Gliotoxin | rumen fluid | 0, 1, 2, 5, 10, 20, 40, 80 μg/mL buffered rumen fluid | < 80 μg/mL no effects. At 80 μg/mL ↓ DM degradation, gas and VFA productions | [149] |
| FB1 | rumen fluid | 0, 50 or 100 mg/kg rumen fluid | none | [150] |
| OTA | rumen fluid | 200 μg of OTA/l of rumen fluid | none | [151] |
| Patulin | rumen fluid | 20, 100 and 300 μg of Patulin/mL rumen liquid | ↓ Acetic acid production within 4 h and Inhibition of protein synthesis | [152] |
| Patulin | rumen fluid | 0, 10, 20 and 40 mg of Patulin/mL rumen fluid | ↓ dDM, VFA production, dNDF, dADF, dCHO, dCP and bacterial N flows ↑ NH3-N | [153] |
| Mycopenolic acid, Roquefortine C and PR toxin | rumen fluid | 0.01, 0.30, 1.01, 1.71 and 2.00 μg of each mycotoxin/mL buffered rumen fluid | Mychopenolic acid and roquefortine C ↓ gas production, VFA production. No effect of PR toxin | [130] |
| Citrinin, Monacolin K, Pravastatin and Mevastatin | rumen fluid | 5 or 20 μg of monacolin/mL rumen fluid; 5 or 20 μg of citrinin/mL rumen fluid; Monascus spp. contaminated rice | none, ↓Methane production | [154] |
| Mycotoxins a | Study | Animals | Tested Dosages | Reported Effects | References | Notes |
|---|---|---|---|---|---|---|
| AFB1 | FT | Beef | 0.2, 0.4, 0.6 or 0.8 mg of AFB1/kg of BW | ↓ rumen mobility | [155] | |
| AFB1 | FT | Beef | 0, 100, 300, 700 and 1000 μg AFB1/kg diet | For levels 700 and 1000 μg/kg: Growth inhibition, ↓ feed efficiencies, ↑ liver and kidney weights | [156] | |
| AFB1 | FT | Lactating dairy cows | 20 μg AFB1/kg diet | ↓ feed consumption, ↓ milk production | [157] | |
| AFB1 | FT | Lactating dairy cows | 120 μg AFB1/kg diet | ↓ reproductive efficiency, ↓ milk production | [158] | |
| AFB1 | FT | Lactating dairy cows | 100 μg AFB1/kg diet | ↓ milk production | [159] | |
| AFB1 | ET | Lactating dairy cows | 100 and 300 μg of AFB1/kg of BW | ↓ feed intake → ↓ milk production | [160] | |
| AFB1 | ET | Lactating dairy cows | 13 mg of AFB1 (pure and impure from Aspergillus parasiticus in culture) | ↓ milk production | [161] | |
| AFB1 | ET | Sheep | 1.8 and 2.4 mg of AFB1/kg diet | none | [162] | Exposition period of 5 years |
| AFB1 | FT | Sheep | 0.75 mg of AFB1/kg diet | Inappetence, apathy, hepatic lesion, neurological signs and death. | [163] | |
| AFB1 | ET | Lambs | 2.6 mg of AFB1/kg diet | ↓ BW ↑ AST, GGT, prothrombin time, cholesterol, uric acid and triglyceride values ↓ albumin, glucose and urea nitrogen and urea-to-creatine ratio | [164] | |
| AFB1 | ET | Lambs | 2 mg of AFB1/kg diet | = BW ↓ ADG, immune response | [165] | |
| AFB1 | ET | Lambs | 350 μg AFB1/kg diet | = ADI and blood parameters ↓ ADG gain, serum Ca and P | [166] | Exposition period of 150 days |
| AFB1 | ET | Lambs | 0, 5.9, 11.8, 17.7, 23.5 μg AFB1/kg diet | = DMI, cellular immunity ↓ ADW | [167] | |
| AFB1 | ET | Lambs | 2.5 mg of AFB1/kg diet | ↓ feed intakes, daily gain, and gain/feed ↑ AST, GGT, total protein, cholesterol | [168] | |
| AFB1 | ET | Lactating dairy cows | 96 μg/cow/day | slightly ↑ GGT and serum protein | [169] | |
| AFB1&FB1+FB2 | ET | Heifers | C (1.9 μg of AFB1 and 3.8 mg of FBs/kg diet), A (12.0 μg of AFB1 and 6.6 mg of FBs/kg diet), A-F (19.9 μg of AFB1 and 23.2 mg of FBs/kg diet) diets | = BW, DMI ↑GGT delay in reproductive career | [170] | |
| AFB1, DON, ZEA, FB1, OTA, T-2 toxin | ET | Lactating dairy cows | 38 AFB1 and 270 T-2 μg/kg; 720 DON, 701 FB1, 541 ZEA, 501 OTA mg/kg | ↓ DMI, milk yield, CP and NDF digestibilities, impact on haematological parameters and immunosuppression | [171] | |
| Maltoryzine | Lactating dairy cows | unknown | general poison | [172] | ||
| β-nitropropionic acid | Sheep and Cattle | unknown | emphysema and difficulty in locomotion | [173] | ||
| DON | ET | Lactating and no lactating dairy cows | 0.3 or 3.4/4.4 mg of DON/kg diet | ↓ NDF digestibility and slightly ↓ in microbial crude protein | [146] | Two level of F:C ratio, being 40:60 or 70:30 |
| DON | ET | Lactating dairy cows | 4.4 or 5.3 mg DON/kg DM | ↑ DMI ↓ Milk Fat | [174] | |
| DON | ET | Lactating dairy cows | 4.4 or 5.3 mg DON/kg DM | ↑ valerate ↓ pH, acetate and isobutyrate | [175] | |
| DON | ET | Lactating dairy cows | 0.59, 42, and 104 mg of DON/cow/day | none | [176] | |
| DON | ET | Lactating dairy cows | 8 mg of DON/kg diet | none | [177] | |
| DON | ET | Non lactating cows | about 8 or 35 mg of DON/cow/day | none, except slightly ↓ ingestion of contaminated feed | [178] | |
| DON | ET | Lactating dairy cows | 66 mg of DON/kg diet | none | [176] | |
| DON | ET | Non lactating cows | 4 or 3.6 mg of DON/kg diet and 0.13 or 0.05 mg of ZEA/kg in experiments 1 and 2, respectively | = rumen pH and VFA production ↓ microbial protein and ↑rumen NH3-N concentration and | [179] | |
| DON | ET | Lactating dairy cows | 3.5 mg of DON/kg diet and 0.24 mg of ZEA/kg diet | = DMI and milk production; Influence on metabolic parameters and immune response | [180,181] | |
| DON | ET | Lactating dairy cows | Group CON (0.02 mg ZEA and 0.07 mg DON/ kg DM), group FUS-50 (0.33 mg ZEA and 2.62 mg DON/kg DM), group FUS-100 (0.66 mg ZEA and 5.24 mg DON/ kg DM) | none | [182] | |
| DON | ET | Lactating dairy cows | The average daily intake of DON in group K was 12.4 mg, in group T 14.1 mg and in group M 14.3 mg and ZEA in group K was 12.4 mg, in group T 0.67 mg and in group M 0.68 mg | slightly ↑ in AST and LDH | [183] | |
| AFB1 & DAS | ET | Lambs | Group control (uncontaminated), group AFB1-contaminated (2.5 mg /kg), group DAS-contaminated (5 mg/kg from chemical standard) and group AFB1/DAS co-contaminated (2.5 mg of AFB1 and 5 mg of diet/kg) diets | ↓ Feed ingestion, BW | [184] | |
| FBs | ET | Lactating dairy cows | 75 mg of FBs/kg and 3 mg FB1/kg BW | none | [185] | |
| FB1 | ET | Steers | 94 mg FB1/kg diet | ↑ AST, GGT, hepatocellular injury and biliary epithelial hyperplasia | [23] | Exposition period of 253 days |
| FBs | ET | Claves | 15, 31 or 148 mg FBs/kg diet | = Feed ingestion, BW ↑AST, GGT, LDH, bilirubin and cholesterol | [186] | |
| FB1 | ET | Milk-fed calves | 1 mg of FB1/kg BW intravenous administered | Liver and kidney lesions ↑ serum AST, ALP, GGT and sorbitol dehydrogenase | [187] | |
| FBs | ET | Lambs | 0, 11.1, 22.2 or 45.5 mg of FBs/kg BW | Death, ↑ alkaline phosphatase, GGT, AST, cholesterol, triglyceride, urea nitrogen and creatinine | [188] | |
| ZEA | ET | Heifers | 250 mg ZEA/heifer | ↓ Conception rate, no other effects | [189] | |
| ZEA | ET | Dairy cow | from 0 to 500 mg ZEA/cow | None | [190] | |
| DON & ZEA | FT | Heifers | About 500 μg of DON/kg diet and 750 μg of ZEA/kg diet | unsynchronized ovarian cycles, vaginitis and early development of mammary gland in the prepubertal heifers | [191] | |
| ZEA | ET | Ewes | 1.5, 3, 6, 12, or 24 mg ZEA/ewe | reproductive disorders, lower lambing percentages and infertility. | [192] | |
| OTA | ET | Sheep | 0, 1.4, or 3.5 mg of OTA/kg diet | =feed intake and nutrient utilization | [193] | |
| OTA | ET | Sheep | 14 mg of OTA/kg diet | ↓ feed intake | [193] | Preliminary ET |
| Mychopenolic acid | ET | Sheep (male) | 0, 10, 70, 300 mg of MPA/sheep/day | none | [194] | |
| Mychopenolic acid | ET | Sheep | 300 mg of MPA/sheep/day | Slightly signs of immunosuppression in jejunum, white blood cells, ileum | [195] | |
| Mychopenolic acid | ET | Sheep | 300 mg of MPA/sheep/day | none | [196] | |
| Roquefortine C | FT | Cow | about 4–8 mg of RC/kg diet | Reversible paralytic effects | [197] | |
| Roquefortine C | ET | Sheep | 0, 10 and 50 mg of RC/sheep/day | None ↓ rumen pH | [95] | |
| Citrinin | FT | Sheep | Presence of visible moldy feeds in diets contaminated by both citrinin (2–10 mg/kg) and OTA (0–20 mg/kg) | fever, diarrhea and uraemia | [198] | |
| Citrinin, monacolin K, pravastatin and mevastatin | ET | Sheep | Monascus fermented rice | None ↓ rumen methane production | [154] | |
| Patulin | FT | Beef | Suspected Patulin | neurotoxicosis, comprising tremors, ataxia, paresis, recumbency and death | [199] |
3.1. Alternaria Derived Toxins
3.2. Aspergillus Derived Toxins
3.3. Fusarium Derived Toxins
3.4. Penicillium Derived Toxins
3.5. Monascus Ruber Derived Toxins
3.6. Endophytic Fungal Toxins
3.7. Intestinal Modulation of Mycotoxins
4. On Farm Strategies to Minimize Risk of Mycotoxin Contaminations in Forages
4.1. Prevention of Mycotoxin Contaminations of Crops in Field and during Storage
4.2. Detoxification and Biodegradation of Mycotoxins on Farm Conditions
5. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Zain, M.E. Impact of mycotoxins on humans and animals. J. Saudi Chem. Soc. 2011, 15, 129–144. [Google Scholar] [CrossRef]
- Zinedine, A.; Soriano, J.M.; Moltó, J.C.; Mañes, J. Review on the toxicity, occurrence, metabolism, detoxification, regulations and intake of zearalenone: An oestrogenic mycotoxin. Food Chem. Toxicol. 2007, 45, 1–18. [Google Scholar] [CrossRef]
- Fink-Gremmels, J. The role of mycotoxins in the health and performance of dairy cows. Vet. J. 2008, 176, 84–92. [Google Scholar] [CrossRef] [PubMed]
- Council for Agricultural Science and Technology (CAST). Mycotoxins: Risks in Plant, Animal, and Human Systems; CAST: Ames, IA, USA, 2003. [Google Scholar]
- Scudamore, K.A.; Livesey, C.T. Occurrence and Significance of Mycotoxins in Forage Crops and Silage: A Review. J. Sci. Food Agric. 1998, 77, 1–17. [Google Scholar] [CrossRef]
- Kabak, B.; Dobson, A.D.W.; Var, I. Strategies to prevent mycotoxin contamination of food and animal feed: A review. Crit. Rev. Food Sci. Nutr. 2006, 46, 593–619. [Google Scholar] [CrossRef] [PubMed]
- Bryden, W.L. Mycotoxin contamination of the feed supply chain: Implications for animal productivity and feed security. Anim. Feed Sci. Technol. 2012, 173, 134–158. [Google Scholar] [CrossRef]
- Storm, I.M.L.D.; Sørensen, J.L.; Rasmussen, R.R.; Nielsen, K.F.; Thrane, U. Mycotoxins in silage. Stewart Postharvest Rev. 2008, 4, 1–12. [Google Scholar] [CrossRef]
- Jennessen, J.; Nielsen, K.F.; Houbraken, J.; Lyhne, E.K.; Schnorer, J.; Frisvad, J.C.; Samson, R.A. Secondary metabolite and mycotoxin production by the Rhizopus microsporus group. J. Agric. Food Chem. 2005, 53, 1833–1840. [Google Scholar] [CrossRef] [PubMed]
- O’Brien, M.; Nielsen, K.F.; O’Kiely, P.; Forristal, P.D.; Fuller, H.T.; Frisvad, J.C. Mycotoxins and other secondary metabolites produced in vitro by Penicillium paneum Frisvad and Penicillium roqueforti Thom isolated from baled grass silage in Ireland. J. Agric. Food Chem. 2006, 54, 9268–9276. [Google Scholar] [CrossRef] [PubMed]
- Nielsen, K.F.; Sumarah, M.W.; Frisvad, J.C.; Miller, J.D. Production of metabolites from the Penicillium roqueforti complex. J. Agric. Food Chem. 2006, 54, 3756–3763. [Google Scholar] [CrossRef] [PubMed]
- Streit, E.; Schatzmayr, G.; Tassis, P.; Tzika, E.; Marin, D.; Taranu, I.; Tabuc, C.; Nicolau, A.; Aprodu, I.; Puel, O.; et al. Current situation of mycotoxin contamination and co-occurrence in animal feed focus on Europe. Toxins 2012, 4, 788–809. [Google Scholar] [CrossRef] [PubMed]
- Magan, N.; Medina, A.; Aldred, D. Possible climate-change effects on mycotoxin contamination of food crops pre- and postharvest. Plant Pathol. 2011, 60, 150–163. [Google Scholar] [CrossRef]
- Cole, R.J.; Cox, R.H. Handbook of Toxic Fungal Metabolites; Academic Press, INC.: New York, NY, USA, 1981. [Google Scholar]
- Richard, J.L. Some major mycotoxins and their mycotoxicoses-An overview. Int. J. Food Microbiol. 2007, 119, 3–10. [Google Scholar] [CrossRef] [PubMed]
- Santos, R.R.; Fink-Gremmels, J. Mycotoxin syndrome in dairy cattle: Characterisation and intervention results. World Mycotoxin J. 2014, 7, 357–366. [Google Scholar] [CrossRef]
- Giovati, L.; Gallo, A.; Masoero, F.; Cerioli, C.; Ciociola, T.; Conti, S.; Magliani, W.; Polonelli, L. Vaccination of heifers with anaflatoxin improves the reduction of aflatoxin B1 carry over in milk of lactating dairy cows. PLoS ONE 2014, 9, e94440. [Google Scholar] [CrossRef] [PubMed]
- Murugesan, G.R.; Ledoux, D.R.; Naehrer, K.; Berthiller, F.; Applegate, T.J.; Grenier, B.; Phillips, T.D.; Schatzmayr, G. Prevalence and effects of mycotoxins on poultry health and performance, and recent development in mycotoxin counteracting strategies. Poult. Sci. 2015, 94, 1298–1315. [Google Scholar] [CrossRef] [PubMed]
- Jarvis, B.B. Advances in Experimental Medicine and Biology. In Mycotoxins and Food Safety; deVries, J.W., Trucksess, M.W., Jackson, L.S., Eds.; Springer US: Boston, MA, USA, 2002; Volume 504. [Google Scholar]
- Vieira, M.L.A.; Johann, S.; Hughes, F.M.; Rosa, C.A.; Rosa, L.H. The diversity and antimicrobial activity of endophytic fungi associated with medicinal plant Baccharis trimera (Asteraceae) from the Brazilian savannah. Can. J. Microbiol. 2014, 60, 847–856. [Google Scholar] [CrossRef] [PubMed]
- Rizzo, I.; Varsavky, E.; Haidukowski, M.; Frade, H. Macrocyclic trichothecenes in Baccharis coridifolia plants and endophytes and Baccharis artemisioides plants. Toxicon 1997, 35, 753–757. [Google Scholar] [CrossRef]
- Jouany, J.P. Methods for preventing, decontaminating and minimizing the toxicity of mycotoxins in feeds. Anim. Feed Sci. Technol. 2007, 137, 342–362. [Google Scholar] [CrossRef]
- Baker, D.C.; Rottinghaus, G.E. Chronic experimental fumonisin intoxication of calves. J. Vet. Diagn. Investig. 1999, 11, 289–292. [Google Scholar] [CrossRef]
- Storm, I.M.L.D.; Rasmussen, R.R.; Rasmussen, P.H. Occurrence of Pre- and Post-Harvest Mycotoxins and Other Secondary Metabolites in Danish Maize Silage. Toxins 2014, 6, 2256–2269. [Google Scholar] [CrossRef] [PubMed]
- Fink-Gremmels, J.; Malekinejad, H. Clinical effects and biochemical mechanisms associated with exposure to the mycoestrogen zearalenone. Anim. Feed Sci. Technol. 2007, 137, 326–341. [Google Scholar] [CrossRef]
- Pestka, J.J. Deoxynivalenol: Toxicity, mechanisms and animal health risks. Anim. Feed Sci. Technol. 2007, 137, 283–298. [Google Scholar] [CrossRef]
- Mobashar, M.; Hummel, J.; Blank, R.; Südekum, K.-H. Ochratoxin A in Ruminants—A Review on Its Degradation by Gut Microbes and Effects on Animals. Toxins 2010, 2, 809–839. [Google Scholar] [CrossRef] [PubMed]
- Cheli, F.; Campagnoli, A.; Dell’Orto, V. Fungal populations and mycotoxins in silages: From occurrence to analysis. Anim. Feed Sci. Technol. 2013, 183, 1–16. [Google Scholar] [CrossRef]
- Gallo, A.; Masoero, F. In vitro models to evaluate the capacity of different sequestering agents to adsorb aflatoxins. Ital. J. Anim. Sci. 2009, 9, 109–116. [Google Scholar] [CrossRef]
- Niderkorn, V.; Morgavi, D.P.; Pujos, E.; Tissandier, A.; Boudra, H. Screening of fermentative bacteria for their ability to bind and biotransform deoxynivalenol, zearalenone and fumonisins in an in vitro simulated corn silage model. Food Addit. Contam. 2007, 24, 406–415. [Google Scholar] [CrossRef] [PubMed]
- Fink-Gremmels, J. Mycotoxins in cattle feeds and carry-over to dairy milk: A review. Food Addit. Contam. A Chem. Anal. Control Expo. Risk Assess. 2008, 25, 172–180. [Google Scholar] [CrossRef] [PubMed]
- Gallo, A.; Moschini, M.; Cerioli, C.; Masoero, F. Use of principal component analysis to classify forages and predict their calculated energy content. Animal 2013, 7, 930–939. [Google Scholar] [CrossRef] [PubMed]
- Giuberti, G.; Gallo, A.; Masoero, F.; Ferraretto, L.F.; Hoffman, P.C.; Shaver, R.D. Factors affecting starch utilization in large animal food production system: A review. Starch/Stärke 2014, 66, 72–90. [Google Scholar] [CrossRef]
- Boysen, M.E.; Jacobsson, K.-G.; Schnurer, J. Molecular Identification of Species from the Penicillium roqueforti Group Associated with Spoiled Animal Feed. Appl. Environ. Microbiol. 2000, 66, 1523–1526. [Google Scholar] [CrossRef] [PubMed]
- Sumarah, M.W.; Miller, J.D.; Blackwell, B.A. Isolation and metabolite production by Penicillium roqueforti, P. paneum and P. crustosum isolated in Canada. Mycopathologia 2005, 159, 571–577. [Google Scholar] [CrossRef] [PubMed]
- Driehuis, F.; Spanjer, M.C.; Scholten, J.M.; te Giffel, M.C. Occurrence of mycotoxins in feedstuffs of dairy cows and estimation of total dietary intakes. J. Dairy Sci. 2008, 91, 4261–4671. [Google Scholar] [CrossRef] [PubMed]
- Driehuis, F. Silage and the safety and quality of dairy foods: A review. Agric. Food Sci. 2013, 22, 16–34. [Google Scholar]
- Casteel, S.W.; Rottinghaus, G.E.; Johnson, G.C.; Wicklow, D.T. Liver disease in cattle induced by consumption of moldy hay. Vet. Hum. Toxicol. 1995, 37, 248–251. [Google Scholar] [PubMed]
- Fink-Gremmels, J.; Diaz, D.E. Mycotoxins in forages. In The Mycotoxin Blue Book; Diaz, D.E., Ed.; Nottingham University Press: Thrumpton, Nottingham, UK, 2005; pp. 249–268. [Google Scholar]
- Dell’Orto, V.; Baldi, G.; Cheli, F. Mycotoxins in silage: Checkpoints for effective management and control. World Mycotoxin J. 2015. [Google Scholar] [CrossRef]
- Placinta, C.M.; D’Mello, J.P.F.; MacDonald, A.M.C. A review of worldwide contamination of cereal grains and animal feed with Fusarium mycotoxins. Anim. Feed Sci. Technol. 1999, 78, 21–37. [Google Scholar] [CrossRef]
- Scott, P.M. Mycotoxins in feeds and ingredients and their origin. J. Food Prot. 1978, 14, 385–398. [Google Scholar]
- Filtenborg, O.; Frisvad, J.C.; Thrane, U. Moulds in food spoilage. Int. J. Food Microbiol. 1996, 33, 85–102. [Google Scholar] [CrossRef]
- Meister, U.; Springer, M. Mycotoxins in cereals and cereal products: Occurrence and changes during processing. J. Appl. Bot. Food Qual. 2004, 78, 168–173. [Google Scholar]
- Leung, M.C.K.; Díaz-Llano, G.; Smith, T.K. Mycotoxins in pet food: A review on worldwide prevalence and preventative strategies. J. Agric. Food Chem. 2006, 54, 9623–9635. [Google Scholar] [CrossRef] [PubMed]
- Binder, E.M.; Tan, L.M.; Chin, L.J.; Handl, J.; Richard, J. Worldwide occurrence of mycotoxins in commodities, feeds and feed ingredients. Anim. Feed Sci. Technol. 2007, 137, 265–282. [Google Scholar] [CrossRef]
- Reddy, K.R.N.; Abbas, H.K.; Abel, C.A.; Shier, W.T.; Oliveira, C.A.F.; Raghavender, C.R. Mycotoxin contamination of commercially important agricultural commodities. Toxin Rev. 2009, 28, 154–168. [Google Scholar] [CrossRef]
- Streit, E.; Naehrer, K.; Rodrigues, I.; Schatzmayr, G. Mycotoxin occurrence in feed and feed raw materials worldwide: Long-term analysis with special focus on Europe and Asia. J. Sci. Food Agric. 2013, 93, 2892–2899. [Google Scholar] [CrossRef] [PubMed]
- Miller, J.D. Mycotoxins in small grains and maize: Old problems, new challenges. Food Addit. Contam. A Chem. Anal. Control Expo. Risk Assess. 2008, 25, 219–230. [Google Scholar] [CrossRef] [PubMed]
- Lazzaro, I.; Moretti, A.; Giorni, P.; Brera, C.; Battilani, P. Organic vs conventional farming: Differences in infection by mycotoxin-producing fungi on maize and wheat in Northern and Central Italy. Crop Prot. 2015, 72, 22–30. [Google Scholar] [CrossRef]
- Prandini, A.; Sigolo, S.; Filippi, L.; Battilani, P.; Piva, G. Review of predictive models for Fusarium head blight and related mycotoxin contamination in wheat. Food Chem. Toxicol. 2009, 47, 927–931. [Google Scholar] [CrossRef] [PubMed]
- European Commission (EC). Directive 2002/32/EC of the European Parliament and of the Council of 7 May 2002 on undesirable substances in animal feed. Off. J. Eur. Union 2002, L140, 10–21. [Google Scholar]
- European Commission (EC). Commission recommendation (2006/576/EU) of 17 August 2006 on the presence of deoxynivalenol, zearalenone, ochratoxin A, T-2 and HT-2 and fumonisins in products intended for animal feeding. Off. J. Eur. Union 2006, L229, 7–9. [Google Scholar]
- Food and Drug Administration (FDA). Mycotoxin Regulatory Guidance. A Guide for Grain Elevators, Feed Manufacturers, Grain Processors and Exporters; National Grain and Feed Association: Washington, DC, USA, 2011.
- Ashiq, S. Natural Occurrence of Mycotoxins in Food and Feed: Pakistan Perspective. Compr. Rev. Food Sci. Food Saf. 2015, 14, 159–175. [Google Scholar] [CrossRef]
- European Commission (EC). Commission Regulation (EU) 2015/786 of 19 May 2015 defining acceptability criteria for detoxification processes applied to products intended for animal feed as provided for in Directive 2002/32/EC of the European Parliament and of the Council. Off. J. Eur. Union 2015, L125, 10–14. [Google Scholar]
- Cheeke, P.R. Endogenous toxins and mycotoxins in forage grasses and their effects on livestock. J. Anim. Sci. 1995, 73, 909–918. [Google Scholar] [PubMed]
- Cheeke, P.R. Natural Toxicants in Feeds, Forages, and Poisonous Plants, 2nd ed.; Interstate Publishers, Inc.: Danville, IL, USA, 1998. [Google Scholar]
- Alonso, V.A.; Pereyra, C.M.; Keller, L.A.M.; Dalcero, A.M.; Rosa, C.A.R.; Chiacchiera, S.M.; Cavaglieri, L.R. Fungi and mycotoxins in silage: An overview. J. Appl. Microbiol. 2013, 115, 637–643. [Google Scholar] [CrossRef] [PubMed]
- Mostrom, M.S.; Jacobsen, B.J. Ruminant mycotoxicosis. Vet. Clin. N. Am. Food Anim. Pract. 2011, 27, 315–344. [Google Scholar] [CrossRef] [PubMed]
- Skládanka, J.; Nedělník, J.; Adam, V.; Doležal, P.; Moravcová, H.; Dohnal, V. Forage as a primary source of mycotoxins in animal diets. Int. J. Environ. Res. Public Health 2011, 8, 37–50. [Google Scholar] [CrossRef] [PubMed]
- Keller, L.A.M.; González Pereyra, M.L.; Keller, K.M.; Alonso, V.A.; Oliveira, A.A.; Almeida, T.X.; Barbosa, T.S.; Nunes, L.M.T.; Cavaglieri, L.R.; Rosa, C.A.R. Fungal and mycotoxins contamination in corn silage: Monitoring risk before and after fermentation. J. Stored Prod. Res. 2013, 52, 42–47. [Google Scholar] [CrossRef]
- Medina, A.; Rodriguez, A.; Magan, N. Effect of climate change on Aspergillus flavus and aflatoxin B1 production. Front. Microbiol. 2014, 5, 348–366. [Google Scholar] [CrossRef] [PubMed]
- Battilani, P.; Camardo Leggieri, M.; Rossi, V.; Giorni, P. AFLA-maize, a mechanistic model for Aspergillus flavus infection and aflatoxin B1 contamination in maize. Comput. Electron. Agric. 2013, 94, 38–46. [Google Scholar] [CrossRef]
- Mansfield, M.A.; Kuldau, G.A. Microbiological and molecular determination of mycobiota in fresh and ensiled maize silage. Mycologia 2007, 99, 269–278. [Google Scholar] [CrossRef] [PubMed]
- Pelhate, J. Maize silage: Incidence of moulds during conservation. Folia Vet. Lat. 1977, 7, 1–16. [Google Scholar] [PubMed]
- Vissers, M.M.M.; Driehuis, F.; Te Giffel, M.C.; de Jong, P.; Lankveld, J.M.G. Concentrations of butyric acid bacteria spores in silage and relationships with aerobic deterioration. J. Dairy Sci. 2007, 90, 928–936. [Google Scholar] [CrossRef]
- Borreani, G.; Tabacco, E. The relationship of silage temperature with the microbiological status of the face of corn silage bunkers. J. Dairy Sci. 2010, 93, 2620–2629. [Google Scholar] [CrossRef] [PubMed]
- Gallo, A.; Bertuzzi, T.; Giuberti, G.; Moschini, M.; Bruschi, S.; Cerioli, C.; Masoero, F. New assessment based on the use of principal factor analysis to investigate corn silage quality from nutritional traits, fermentation end products and mycotoxins. J. Sci. Food Agric. 2015. [Google Scholar] [CrossRef] [PubMed]
- Andersen, B.; Nielsen, K.F.; Fernández Pinto, V.; Patriarca, A. Characterization of Alternaria strains from Argentinean blueberry, tomato, walnut and wheat. Int. J. Food Microbiol. 2015, 196, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Andersen, B.; Krøger, E.; Roberts, R.G. Chemical and morphological segregation of Alternaria arborescens, A. infectoria and A. tenuissima species-groups. Mycol. Res. 2002, 106, 170–182. [Google Scholar] [CrossRef]
- Andersen, B.; Krøger, E.; Roberts, R.G. Chemical and morphological segregation of Alternaria alternata, A. gaisen and A. longipes. Mycol. Res. 2001, 105, 291–299. [Google Scholar] [CrossRef]
- Christensen, K.B.; van Klink, J.W.; Weavers, R.T.; Larsen, T.O.; Andersen, B.; Phipps, R.K. Novel chemotaxonomic markers of the Alternaria infectoria species-group. J. Agric. Food Chem. 2005, 53, 9431–9435. [Google Scholar] [CrossRef] [PubMed]
- European Food Safety Authority (EFSA). Scientific Opinion on the risks for animal and public health related to the presence of Alternaria toxins in feed and food. EFSA J. 2011, 9, 1–97. [Google Scholar]
- Yu, W.; Yu, F.-Y.; Undersander, D.J.; Chu, F.S. Immunoassays of Selected Mycotoxins in Hay, Silage and Mixed Feed. Food Agric. Immunol. 1999, 11, 307–319. [Google Scholar] [CrossRef]
- Mansfield, M.A.; Archibald, D.D.; Jones, A.D.; Kuldau, G.A. Relationship of sphinganine analog mycotoxin contamination in maize silage to seasonal weather conditions and to agronomic and ensiling practices. Phytopathology 2007, 97, 504–511. [Google Scholar] [CrossRef] [PubMed]
- Richard, E.; Heutte, N.; Bouchart, V.; Garon, D. Evaluation of fungal contamination and mycotoxin production in maize silage. Anim. Feed Sci. Technol. 2009, 148, 309–320. [Google Scholar] [CrossRef]
- Cavallarin, L.; Borreani, G.; Tabacco, E.; Lúscher, A.; Jeangros, B.; Kessler, W.; Huguenin, O.; Lobsiger, M.; Millar, N.; Suter, D. Mycotoxin occurrence in farm maize silages in northern Italy. In Land Use Systems in Grassland Dominated Regions; Proceedings of the 20th General Meeting of the European Grassland Federation, Luzern, Switzerland, 21–24 June 2004; Lüscher, A., Jeangros, B., Huguenin, O., Lobsiger, M., Millar, N., Suter, D., Eds.; Swiss Grassland Society: Zürich, Switzerland, 2004; pp. 1023–1025. [Google Scholar]
- Garon, D.; Richard, E.; Sage, L.; Bouchart, V.; Pottier, D.; Lebailly, P. Mycoflora and multimycotoxin detection in corn silage: Experimental study. J. Agric. Food Chem. 2006, 54, 3479–3484. [Google Scholar] [CrossRef] [PubMed]
- Mansfield, M.A.; Jones, A.D.; Kuldau, G.A. Contamination of fresh and ensiled maize by multiple penicillium mycotoxins. Phytopathology 2008, 98, 330–336. [Google Scholar] [CrossRef] [PubMed]
- Pereyra, C.M.; Alonso, V.A.; Rosa, C.A.R.; Chiacchiera, S.M.; Dalcero, A.M.; Cavaglieri, L.R. Gliotoxin natural incidence and toxigenicity of Aspergillus fumigatus isolated from corn silage and ready dairy cattle feed. World Mycotoxin J. 2008, 1, 457–462. [Google Scholar] [CrossRef]
- Driehuis, F.; Spanjer, M.C.; Scholten, J.M.; Te Giffel, M.C. Occurrence of mycotoxins in maize, grass and wheat silage for dairy cattle in the Netherlands. Food Addit. Contam. B 2008, 1, 41–50. [Google Scholar] [CrossRef] [PubMed]
- Schollenberger, M.; Müller, H.M.; Rüfle, M.; Suchy, S.; Plank, S.; Drochner, W. Natural occurrence of 16 Fusarium toxins in grains and feedstuffs of plant origin from Germany. Mycopathologia 2006, 161, 43–52. [Google Scholar] [CrossRef] [PubMed]
- Storm, I.M.L.D.; Kristensen, N.B.; Raun, B.M.L.; Smedsgaard, J.; Thrane, U. Dynamics in the microbiology of maize silage during whole-season storage. J. Appl. Microbiol. 2010, 109, 1017–1026. [Google Scholar] [CrossRef] [PubMed]
- Eckard, S.; Wettstein, F.E.; Forrer, H.R.; Vogelgsang, S. Incidence of Fusarium species and mycotoxins in silage maize. Toxins 2011, 3, 949–967. [Google Scholar] [CrossRef] [PubMed]
- Oldenburg, E.; Eiiner, F. Fusarium mycotoxins in forage maize—Detection and evaluation. Mycotoxin Res. 2005, 21, 105–107. [Google Scholar] [CrossRef] [PubMed]
- Mansfield, M.A.; de Wolf, E.D.; Kuldau, G.A. Relationships Between Weather Conditions, Agronomic Practices, and Fermentation Characteristics with Deoxynivalenol Content in Fresh and Ensiled Maize. Plant Dis. 2005, 89, 1151–1157. [Google Scholar] [CrossRef]
- Kim, E.-K.; Maragos, C.M.; Kendra, D.F. Liquid chromatographic determination of fumonisins B1, B2, and B3 in corn silage. J. Agric. Food Chem. 2004, 52, 196–200. [Google Scholar] [CrossRef] [PubMed]
- McElhinney, C.; Danaher, M.; Elliott, C.; O’Kiely, P. Identification and quantification of mycotoxins in silages: An Irish national survey. In The world Mycotoxin Forum; IBERS, Aberystwyth University, 2014; pp. 87–88. [Google Scholar]
- Borreani, G.; Tabacco, E.; Antoniazzi, S.; Cavallarin, L. Zearalenone contamination in farm maize silage. Ital. J. Anim. Sci. 2005, 4, 162–165. [Google Scholar] [CrossRef]
- Buckley, T.; Creighton, A.; Fogarty, U. Analysis of Canadian and Irish forage, oats and commercially available equine concentrate feed for pathogenic fungi and mycotoxins. Ir. Vet. J. 2007, 60, 231–236. [Google Scholar] [CrossRef] [PubMed]
- Frisvad, J.C.; Rank, C.; Nielsen, K.F.; Larsen, T.O. Metabolomics of Aspergillus fumigatus. Med. Mycol. 2009, 47 (Suppl. 1), S53–S71. [Google Scholar] [CrossRef] [PubMed]
- Schneweis, I.; Meyer, K.; Hormansdorfer, S.; Bauer, J. Mycophenolic acid in silage. Appl. Environ. Microbiol. 2000, 66, 3639–3641. [Google Scholar] [CrossRef] [PubMed]
- Auerbach, H.; Oldenburg, E.; Weissbach, F. Incidence of penicillium roqueforti and roquefortine C in silages. J. Sci. Food Agric. 1998, 76, 565–572. [Google Scholar] [CrossRef]
- Tüller, G.; Armbruster, G.; Wiedenmann, S.; Hänichen, T.; Schams, D.; Bauer, J. Occurrence of roquefortine in silage—toxicological relevance to sheep. J. Anim. Physiol. Anim. Nutr. Berl 1998, 80, 246–249. [Google Scholar] [CrossRef]
- Schneweis, I.; Meyer, K.; Hörmansdorfer, S.; Bauer, J. Metabolites of Monascus ruber in silages. J. Anim. Physiol. Anim. Nutr. Berl 2001, 85, 38–44. [Google Scholar] [CrossRef] [PubMed]
- Sassahara, M.; Pontes Netto, D.; Yanaka, E.K. Aflatoxin occurrence in foodstuff supplied to dairy cattle and aflatoxin M1 in raw milk in the North of Paraná state. Food Chem. Toxicol. 2005, 43, 981–984. [Google Scholar] [CrossRef] [PubMed]
- Cheli, F.; Campagnoli, A.; Pinotti, L.; Fusi, E.; Dell’Orto, V. Sampling feed for mycotoxins: Acquiring knowledge from food. Ital. J. Anim. Sci. 2010, 8, 5–22. [Google Scholar] [CrossRef]
- Miraglia, M.; de Santis, B.; Minardi, V.; Debegnach, F.; Brera, C. The role of sampling in mycotoxin contamination: An holistic view. Food Addit. Contam. 2005, 22 (Suppl. 1), 31–36. [Google Scholar] [PubMed]
- Köppen, R.; Koch, M.; Siegel, D.; Merkel, S.; Maul, R.; Nehls, I. Determination of mycotoxins in foods: Current state of analytical methods and limitations. Appl. Microbiol. Biotechnol. 2010, 86, 1595–1612. [Google Scholar] [CrossRef] [PubMed]
- Brera, C.; de Santis, B.; Prantera, E.; Debegnach, F.; Pannunzi, E.; Fasano, F.; Berdini, C.; Slate, A.B.; Miraglia, M.; Whitaker, T.B. Effect of sample size in the evaluation of “in-field” sampling plans for aflatoxin B1 determination in corn. J. Agric. Food Chem. 2010, 58, 8481–8489. [Google Scholar] [CrossRef] [PubMed]
- Rivas Casado, M.; Parsons, D.J.; Weightman, R.M.; Magan, N.; Origgi, S. Modelling a two-dimensional spatial distribution of mycotoxin concentration in bulk commodities to design effective and efficient sample selection strategies. Food Addit. Contam. A 2009, 26, 1298–1305. [Google Scholar] [CrossRef]
- European Commission (EC). Commission Regulation (EC) No 401/2006 of 23 February 2006 laying down the methods of sampling and analysis for the official control of the levels of mycotoxins in foodstuffs. Off. J. Eur. Union 2006, L70, 12–34. [Google Scholar]
- European Commission (EC). Recomendations on the presence of T-2 and HT-2 toxin in cereals and cereal products. Off. J. Eur. Union 2013, 9, 12–15. [Google Scholar]
- Chang, P.-K.; Ehrlich, K.C.; Fujii, I. Cyclopiazonic acid biosynthesis of Aspergillus flavus and Aspergillus oryzae. Toxins 2009, 1, 74–99. [Google Scholar] [CrossRef] [PubMed]
- Alonso, V.; Díaz Vergara, L.; Aminahuel, C.; Pereyra, C.; Pena, G.; Torres, A.; Dalcero, A.; Cavaglieri, L. Physiological behaviour of gliotoxigenic Aspergillus fumigatus sensu stricto isolated from maize silage under simulated environmental conditions. Food Addit. Contam. A 2015, 32, 236–244. [Google Scholar] [CrossRef] [PubMed]
- Bok, J.W.; Chung, D.; Balajee, S.A.; Marr, K.A.; Andes, D.; Nielsen, K.F.; Frisvad, J.C.; Kirby, K.A.; Keller, N.P. GliZ, a transcriptional regulator of gliotoxin biosynthesis, contributes to Aspergillus fumigatus virulence. Infect. Immun. 2006, 74, 6761–6768. [Google Scholar] [CrossRef] [PubMed]
- Boudra, H.; Morgavi, D.P. Mycotoxin risk evaluation in feeds contaminated by Aspergillus fumigatus. Anim. Feed Sci. Technol. 2005, 120, 113–123. [Google Scholar] [CrossRef]
- Frisvad, J.C.; Frank, J.M.; Kuijpers, A.F.A.; Houbraken, J.A.M.P.; Samson, R.A. New ochratoxin A or sclerotium producing species in Aspergillus section Nigri. Stud. Mycol. 2004, 50, 45–61. [Google Scholar]
- Frisvad, J.C.; Samson, R.A. Polyphasic taxonomy of Penicillium sbgenus Pemicillium: A guide to identification of food and air-bone teverticilliate penicillia and their mycotoxins. Stud. Mycol. 2004, 49, 1–173. [Google Scholar]
- Frisvad, J.C.; Thrane, U.; Samson, R.A.; Hoekstra, E.S. Mycotoxin production by common filamentous fungi. In Introduction to Food- and Airborne Fungi; Samson, R.A., Hoekstra, E.S., Frisvad, J.C., Eds.; Centraalbureau voor Schimmelcultures (CBS): Utrecht, The Netherlands, 2004; pp. 321–331. [Google Scholar]
- Meca, G.; Ritieni, A. Production and analysis of ochratoxin A produced by Aspergillus ochraceus ITEM 5137 in submerged culture. Food Chem. 2009, 117, 470–472. [Google Scholar] [CrossRef]
- Samson, R.A.; Houbraken, J.; Thrane, J.; Frisvad, J.C.; Andersen, B. Food and Indoor Fungi; CBS-KNAW Fungal Biodiversity Centre: Utrecht, The Netherlands, 2010. [Google Scholar]
- McCormick, S.P.; Stanley, A.M.; Stover, N.A.; Alexander, N.J. Trichothecenes: From simple to complex mycotoxins. Toxins 2011, 3, 802–814. [Google Scholar] [CrossRef] [PubMed]
- Chelkowski, J. Fusarium: Mycotoxins, Taxonomy, Pathogenicity; Elsevier Science Publishers B.V.: Amsterdam, The Netherlands, 2014. [Google Scholar]
- European Food Safety Authority (EFSA). Deoxynivalenol in food and feed: Occurrence and exposure. EFSA J. 2013, 11. [Google Scholar] [CrossRef]
- Mansfield, M.A. Fungi and mycotoxins in fresh and ensiled maize and the effects of agronomic practices, wheatear conditions and silage characteristics. Ph.D. Thesis, Penn State University, University Park, PA, USA, 2005. [Google Scholar]
- Dall’Asta, C.; Falavigna, C.; Galaverna, G.; Battilani, P. Role of maize hybrids and their chemical composition in Fusarium infection and fumonisin production. J. Agric. Food Chem. 2012, 60, 3800–3808. [Google Scholar] [CrossRef] [PubMed]
- Kim, S.C.; Adesogan, A.T. Influence of ensiling temperature, simulated rainfall, and delayed sealing on fermentation characteristics and aerobic stability of corn silage. J. Dairy Sci. 2006, 89, 3122–3132. [Google Scholar] [CrossRef] [PubMed]
- Gallo, A.; Giuberti, G.; Bruschi, S.; Fortunati, P.; Masoero, F. Use of principal factor analysis to generate a corn silage fermentative quality index to rank well- or poorly-preserved forages. J. Sci. Food Agric. 2015. [Google Scholar] [CrossRef] [PubMed]
- Sørensen, J.L.; Nielsen, K.F.; Thrane, U. Analysis of moniliformin in maize plants using hydrophilic interaction chromatography. J. Agric. Food Chem. 2007, 55, 9764–9768. [Google Scholar] [CrossRef] [PubMed]
- Boudra, H.; Morgavi, D.P. Reduction in Fusarium toxin levels in corn silage with low dry matter and storage time. J. Agric. Food Chem. 2008, 56, 4523–4528. [Google Scholar] [CrossRef] [PubMed]
- Berthiller, F.; Dall’Asta, C.; Schuhmacher, R.; Lemmens, M.; Adam, G.; Krska, R. Masked mycotoxins: Determination of a deoxynivalenol glucoside in artificially and naturally contaminated wheat by liquid chromatography-tandem mass spectrometry. J. Agric. Food Chem. 2005, 53, 3421–3425. [Google Scholar] [CrossRef] [PubMed]
- Berthiller, F.; Crews, C.; Dall’Asta, C.; de Saeger, S.; Haesaert, G.; Karlovsky, P.; Oswald, I.P.; Seefelder, W.; Speijers, G.; Stroka, J. Masked mycotoxins: A review. Mol. Nutr. Food Res. 2013, 57, 165–186. [Google Scholar] [CrossRef] [PubMed]
- Nagl, V.; Woechtl, B.; Schwartz-Zimmermann, H.E.; Hennig-Pauka, I.; Moll, W.-D.; Adam, G.; Berthiller, F. Metabolism of the masked mycotoxin deoxynivalenol-3-glucoside in pigs. Toxicol. Lett. 2014, 229, 190–197. [Google Scholar] [CrossRef] [PubMed]
- Rychlik, M.; Humpf, H.-U.; Marko, D.; Dänicke, S.; Mally, A.; Berthiller, F.; Klaffke, H.; Lorenz, N. Proposal of a comprehensive definition of modified and other forms of mycotoxins including “masked” mycotoxins. Mycotoxin Res. 2014, 30, 197–205. [Google Scholar] [CrossRef] [PubMed]
- European Food Safety Authority (EFSA). Scientific Opinion on the risks for public health related to the presence of zearalenone in food. EFSA J. 2011, 9, 1–124. [Google Scholar]
- Nout, M.J.R.; Bouwmeester, H.M.; Haaksma, J.; van Dijk, H. Fungal growth in silages of sugarbeet press pulp and maize. J. Agric. Sci. 1993, 121, 323–326. [Google Scholar] [CrossRef]
- Jensen, H.E.; Olsen, S.N.; Aalbaek, B. Gastrointestinal aspergillosis and zygomycosis of cattle. Vet. Pathol. 1994, 31, 28–36. [Google Scholar] [CrossRef] [PubMed]
- Gallo, A.; Giuberti, G.; Bertuzzi, T.; Moschini, M.; Masoero, F. Study of the effects of PR toxin, mycophenolic acid and roquefortine C on in vitro gas production parameters and their stability in the rumen environment. J. Agric. Sci. 2015, 153, 163–176. [Google Scholar] [CrossRef]
- Kiessling, K.H.; Pettersson, H.; Sandholm, K.; Olsen, M. Metabolism of aflatoxin, ochratoxin, zearalenone, and three trichothecenes by intact rumen fluid, rumen protozoa, and rumen bacteria. Appl. Environ. Microbiol. 1984, 47, 1070–1073. [Google Scholar] [PubMed]
- Yiannikouris, A.; Jouany, J.-P. Mycotoxins in feeds and their fate in animals: A review. Anim. Res. 2002, 51, 81–99. [Google Scholar] [CrossRef]
- Jouany, J.P.; Ushida, K. The Role of Protozoa in Feed Digestion—Review -. Asian Australas. J. Anim. Sci. 1999, 12, 113–128. [Google Scholar] [CrossRef]
- Jouany, J.P.; Diaz, D.E. Effects of mycotoxins in ruminants. In Mycotoxins Blue Book; Nottingham University Press: Thrumpton, Nottingham, UK, 2005; pp. 295–321. [Google Scholar]
- Hussein, H. Toxicity, metabolism, and impact of mycotoxins on humans and animals. Toxicology 2001, 167, 101–134. [Google Scholar] [CrossRef] [PubMed]
- Rasmussen, R.R.; Rasmussen, P.H.; Larsen, T.O.; Bladt, T.T.; Binderup, M.L. In vitro cytotoxicity of fungi spoiling maize silage. Food Chem. Toxicol. 2011, 49, 31–44. [Google Scholar] [CrossRef] [PubMed]
- Whitlow, L.; Hagler, W. Mold and mycotoxin issues in dairy cattle: Effects, prevention and treatment. Extention 2010, 1–22. [Google Scholar]
- Santos, R.R.; Schoevers, E.J.; Roelen, B.A.J.; Fink-Gremmels, J. Mycotoxins and female reproduction: In vitro approaches. World Mycotoxin J. 2013, 6, 245–253. [Google Scholar] [CrossRef]
- Pulina, G.; Battacone, G.; Brambilla, G.; Cheli, F.; Danieli, P.P.; Masoero, F.; Pietri, A.; Ronchi, B. An update on the safety of foods of animal origin and feeds. Ital. J. Anim. Sci. 2014, 13, 845–856. [Google Scholar] [CrossRef]
- Dunière, L.; Sindou, J.; Chaucheyras-Durand, F.; Chevallier, I.; Thévenot-Sergentet, D. Silage processing and strategies to prevent persistence of undesirable microorganisms. Anim. Feed Sci. Technol. 2013, 182, 1–15. [Google Scholar] [CrossRef]
- Mojtahedi, M. Effect of aflatoxin B1 on in vitro rumen microbial fermentation responses using batch culture. Annu. Rev. Res. Biol. 2013, 3, 686–693. [Google Scholar]
- Westlake, K.; Mackie, R.I.; Dutton, M.F. In vitro metabolism of mycotoxins by bacterial, protozoal and ovine ruminal fluid preparations. Anim. Feed Sci. Technol. 1989, 25, 169–178. [Google Scholar] [CrossRef]
- Auerbach, H.; Maas, R.F.M.; Op Den Camp, H.J.M.; Pol, A.; Fink Gremmels, J. Biodegradation of aflatoxin B1 by bovine rumen microorganisms in vitro and its effects on rumen fermentation. In Proceedings of the Mycotox 98. Mycotoxins in Food Chain: Processing and toxicological aspects (Mycotox 98. Les Mycotoxines dans la Chaine Alimentaire: Aspects toxicologiques et technologiques), Toulouse, France, 2–4 July 1998.
- Jiang, Y.H.; Yang, H.J.; Lund, P. Effect of aflatoxin B1 on in vitro ruminal fermentation of rations high in alfalfa hay or ryegrass hay. Anim. Feed Sci. Technol. 2012, 175, 85–89. [Google Scholar] [CrossRef]
- Boguhn, J.; Neumann, D.; Helm, A.; Strobel, E.; Tebbe, C.C.; Dänicke, S.; Rodehutscorda, M. Effects of concentrate proportion in the diet with or without Fusarium toxin-contaminated triticale on ruminal fermentation and the structural diversity of rumen microbial communities in vitro. Arch. Anim. Nutr. 2010, 64, 467–483. [Google Scholar] [CrossRef] [PubMed]
- Hildebrand, B.; Boguhn, J.; Dänicke, S.; Rodehutscord, M. Effect of Fusarium toxin-contaminated triticale and forage-to-concentrate ratio on fermentation and microbial protein synthesis in the rumen. J. Anim. Physiol. Anim. Nutr. Berl 2012, 96, 307–318. [Google Scholar] [CrossRef] [PubMed]
- Jeong, J.S.; Lee, J.H.; Simizu, Y.; Tazaki, H.; Itabashi, H.; Kimura, N. Effects of the Fusarium mycotoxin deoxynivalenol on in vitro rumen fermentation. Anim. Feed Sci. Technol. 2010, 162, 144–148. [Google Scholar] [CrossRef]
- May, H.D.; Wu, Q.; Blake, C.K. Effects of the Fusarium spp. mycotoxins fusaric acid and deoxynivalenol on the growth of Ruminococcus albus and Methanobrevibacter ruminantium. Can. J. Microbiol. 2000, 46, 692–699. [Google Scholar] [CrossRef] [PubMed]
- Morgavi, D.P.; Boudra, H.; Jouany, J.P.; Michalet-Doreau, B. Effect and stability of gliotoxin, an Aspergillus fumigatus toxin, on in vitro rumen fermentation. Food Addit. Contam. 2004, 21, 871–878. [Google Scholar] [CrossRef] [PubMed]
- Gurung, N.K.; Rankins, D.L.; Shelby, R.A. In vitro ruminal disappearance of fumonisin B1 and its effects on in vitro dry matter disappearance. Vet. Hum. Toxicol. 1999, 41, 196–199. [Google Scholar] [PubMed]
- Ozpinar, H.; Augonyte, G.; Drochner, W. Inactivation of ochratoxin in ruminal fluid with variation of pH-value and fermentation parameters in an in vitro system. Environ. Toxicol. Pharmacol. 1999, 7, 1–9. [Google Scholar] [CrossRef]
- Escoula, L. Patulin production by Penicillium granulatum and inhibition of ruminal flora. J. Environ. Pathol. Toxicol. Oncol. 1992, 11, 45–48. [Google Scholar] [PubMed]
- Tapia, M.O.; Stern, M.D.; Soraci, A.L.; Meronuck, R.; Olson, W.; Gold, S.; Koski-Hulbert, R.L.; Murphy, M.J. Patulin-producing molds in corn silage and high moisture corn and effects of patulin on fermentation by ruminal microbes in continuous culture. Anim. Feed Sci. Technol. 2005, 119, 247–258. [Google Scholar] [CrossRef]
- Morgavi, D.P.; Martin, C.; Boudra, H. Fungal secondary metabolites from Monascus spp. reduce rumen methane production in vitro and in vivo. J. Anim. Sci. 2013, 91, 848–860. [Google Scholar] [CrossRef] [PubMed]
- Cook, W.O.; Richard, J.L.; Osweiler, G.D.; Trampel, D.W. Clinical and pathologic changes in acute bovine aflatoxicosis: Rumen motility and tissue and fluid concentrations of aflatoxins B1 and M1. Am. J. Vet. Res. 1986, 47, 1817–1825. [Google Scholar] [PubMed]
- Garrett, W.N.; Heitman, H.; Booth, A.N. Aflatoxin Toxicity in Beef Cattle. Exp. Biol. Med. 1968, 127, 188–190. [Google Scholar] [CrossRef]
- Jones, M.G.; Ewart, J.M. Effects on milk production associated with consumption of decorticated extracted groundnut meal contaminated with aflatoxin. Vet. Rec. 1979, 105, 492–493. [Google Scholar] [CrossRef] [PubMed]
- Guthrie, L.D.; Bedell, D.M. Effects of aflatoxin in corn on production and reproduction in dairy cattle. Proc. Annu. Meet. U. S. Anim. Health Assoc. 1979, 83, 202–204. [Google Scholar] [PubMed]
- Patterson, D.S.; Anderson, P.H. Recent aflatoxin feeding experiments in cattle. Vet. Rec. 1982, 110, 60. [Google Scholar] [CrossRef] [PubMed]
- Mertens, D.R.; Wyatt, R.D. Acute aflatoxicosis in lactating dairy cows. J. Dairy Sci. 1977, 60, 153–154. [Google Scholar]
- Applebaum, R.S.; Brackett, R.E.; Wiseman, D.W.; Marth, E.H. Responses of dairy cows to dietary aflatoxin: Feed intake and yield, toxin content, and quality of milk of cows treated with pure and impure aflatoxin. J. Dairy Sci. 1982, 65, 1503–1508. [Google Scholar] [CrossRef]
- Lewis, G.; Markson, L.M.; Allcroft, R. The effect of feeding toxic groundnut meal to sheep over a period of five years. Vet. Proc. 1967, 80, 312–314. [Google Scholar] [CrossRef]
- Suliman, H.B.; Mohamed, A.F.; Awadelsied, N.A.; Shommein, A.M. Acute mycotoxicosis in sheep: Field cases. Vet. Hum. Toxicol. 1987, 29, 241–243. [Google Scholar] [PubMed]
- Harvey, R.B.; Kubena, L.F.; Phillips, T.D.; Corrier, D.E.; Elissalde, M.H.; Huff, W.E. Diminution of aflatoxin toxicity to growing lambs by dietary supplementation with hydrated sodium calcium aluminosilicate. Am. J. Vet. Res. 1991, 52, 152–156. [Google Scholar]
- Fernàndez, A.; Hernaindez, M.; Verde, M.; Sanz, M. Effect of aflatoxin on performance, hematology, and clinical immunology in lambs. Can. J. Vet. Res. 2000, 64, 53–58. [Google Scholar]
- Gowda, N.K.S.; Suganthi, R.U.; Malathi, V.; Raghavendra, A. Efficacy of heat treatment and sun drying of aflatoxin-contaminated feed for reducing the harmful biological effects in sheep. Anim. Feed Sci. Technol. 2007, 133, 167–175. [Google Scholar] [CrossRef]
- Tripathi, M.K.; Mondal, D.; Karim, S.A. Growth, haematology, blood constituents and immunological status of lambs fed graded levels of animal feed grade damaged wheat as substitute of maize. J. Anim. Physiol. Anim. Nutr. Berl 2008, 92, 75–85. [Google Scholar] [CrossRef] [PubMed]
- Edrington, T.S.; Harvey, R.B.; Kubena, L.F. Effect of aflatoxin in growing lambs fed ruminally degradable or escape protein sources. J. Anim. Sci. 1994, 72, 1274–1281. [Google Scholar] [PubMed]
- Masoero, F.; Gallo, A.; Moschini, M.; Piva, G.; Diaz, D. Carryover of aflatoxin from feed to milk in dairy cows with low or high somatic cell counts. Animal 2007, 1, 1344–1350. [Google Scholar] [CrossRef] [PubMed]
- Abeni, F.; Migliorati, L.; Terzano, G.M.; Capelletti, M.; Gallo, A.; Masoero, F.; Pirlo, G. Effects of two different blends of naturally mycotoxin-contaminated maize meal on growth and metabolic profile in replacement heifers. Animal 2014, 8, 1667–1676. [Google Scholar] [CrossRef] [PubMed]
- Kiyothong, K.; Rowlinson, P.; Wanapat, M.; Khampa, S. Effect of mycotoxin deactivator product supplementation on dairy cows. Anim. Prod. Sci. 2012, 52, 832–841. [Google Scholar] [CrossRef]
- U.S. Enviromental Protection Agency (EPA). Aspergillus oryzae Final Risk Assessment, Biotechnology Program Under Toxic Substances Control Act (TSCA). Available online: http://www.epa.gov/biotech_rule/pubs/fra/fra007.htm (accessed on 5 May 2015).
- James, L.F.; Hartley, W.J.; Williams, M.C.; van Kampen, K.R. Field and experimental studies in cattle and sheep poisoned by nitro-bearing Astragalus or their toxins. Am. J. Vet. Res. 1980, 41, 377–382. [Google Scholar] [PubMed]
- Keese, C.; Meyer, U.; Rehage, J.; Spilke, J.; Boguhn, J.; Breves, G.; Dänicke, S. On the effects of the concentrate proportion of dairy cow rations in the presence and absence of a fusarium toxin-contaminated triticale on cow performance. Arch. Anim. Nutr. 2008, 62, 241–262. [Google Scholar] [CrossRef] [PubMed]
- Keese, C.; Meyer, U.; Rehage, J.; Spilke, J.; Boguhn, J.; Breves, G.; Dänicke, S. Ruminal fermentation patterns and parameters of the acid base metabolism in the urine as influenced by the proportion of concentrate in the ration of dairy cows with and without Fusarium toxin-contaminated triticale. Arch. Anim. Nutr. 2008, 62, 287–302. [Google Scholar] [CrossRef] [PubMed]
- Charmley, E.; Trenholm, H.L.; Thompson, B.K.; Vudathala, D.; Nicholson, J.W.; Prelusky, D.B.; Charmley, L.L. Influence of level of deoxynivalenol in the diet of dairy cows on feed intake, milk production, and its composition. J. Dairy Sci. 1993, 76, 3580–3587. [Google Scholar] [CrossRef]
- Ingalls, J.R. Influence of deoxynivalenol on feed consumption by dairy cows. Anim. Feed Sci. Technol. 1996, 60, 297–300. [Google Scholar] [CrossRef]
- Trenholm, H.L.; Thompson, B.K.; Martin, K.E.; Greenhalgh, R.; McAllister, A.J. Ingestion of vomitoxin (Deoxynivalenol)-contaminated wheat by nonlactating dairy cows. J. Dairy Sci. 1985, 68, 1000–1005. [Google Scholar] [CrossRef]
- Dänicke, S.; Matthäus, K.; Lebzien, P.; Valenta, H.; Stemme, K.; Ueberschär, K.-H.; Böhm, J.; Razzazi-Fazeli, E.; Flachowsky, G. Effects of Fusarium toxin-contaminated wheat grain on nutrient turnover, microbial protein synthesis and metabolism of deoxynivalenol and zearalenone in the rumen of dairy cows. J. Anim. Physiol. Anim. Nutr. Berl 2005, 89, 303–315. [Google Scholar] [CrossRef] [PubMed]
- Korosteleva, S.N.; Smith, T.K.; Boermans, H.J. Effects of feedborne Fusarium mycotoxins on the performance, metabolism, and immunity of dairy cows. J. Dairy Sci. 2007, 90, 3867–3873. [Google Scholar] [CrossRef] [PubMed]
- Korosteleva, S.N.; Smith, T.K.; Boermans, H.J. Effects of feed naturally contaminated with Fusarium mycotoxins on metabolism and immunity of dairy cows. J. Dairy Sci. 2009, 92, 1585–1593. [Google Scholar] [CrossRef] [PubMed]
- Winkler, J.; Kersten, S.; Meyer, U.; Engelhardt, U.; Dänicke, S. Residues of zearalenone (ZEN), deoxynivalenol (DON) and their metabolites in plasma of dairy cows fed Fusarium contaminated maize and their relationships to performance parameters. Food Chem. Toxicol. 2014, 65, 196–204. [Google Scholar] [CrossRef] [PubMed]
- Hochsteiner, W.; Schuh, M.; Luger, K.; Baumgartner, W. Effect of mycotoxin contaminated feed on production parameters of dairy cows. Berl. Munchener Tierarztliche Wochenschr. 2000, 113, 14–21. [Google Scholar]
- Harvey, R.B.; Edrington, T.S.; Kubena, L.F.; Elissalde, M.H.; Corrier, D.E.; Rottinghaus, G.E. Effect of aflatoxin and diacetoxyscirpenol in ewe lambs. Bull. Environ. Contam. Toxicol. 1995, 54, 325–330. [Google Scholar] [CrossRef] [PubMed]
- Richard, J.L.; Meerdink, G.; Maragos, C.M.; Tumbleson, M.; Bordson, G.; Rice, L.G.; Ross, P.F. Absence of detectable fumonisins in the milk of cows fed Fusarium proliferatum (Matsushima) Nirenberg culture material. Mycopathologia 1996, 133, 123–126. [Google Scholar] [CrossRef] [PubMed]
- Osweiler, G.D.; Kehrli, M.E.; Stabel, J.R.; Thurston, J.R.; Ross, P.F.; Wilson, T.M. Effects of fumonisin-contaminated corn screenings on growth and health of feeder calves. J. Anim. Sci. 1993, 71, 459–466. [Google Scholar] [PubMed]
- Mathur, S. Fumonisin B1 is hepatotoxic and nephrotoxic in milk-fed calves. Toxicol. Sci. 2001, 60, 385–396. [Google Scholar] [CrossRef] [PubMed]
- Edrington, T.S.; Harvey, R.B.; Kubena, L.F.; Elissalde, M.H.; Rottinghaus, G.E.; Rottinghaust, G.E. Acute hepatic and renal toxicity in lambs dosed with culture material. J. Anim. Sci. 1995, 73, 508–515. [Google Scholar] [PubMed]
- Weaver, G.A.; Kurtz, H.J.; Behrens, J.C.; Robison, T.S.; Seguin, B.E.; Bates, F.Y.; Mirocha, C.J. Effect of zearalenone on the fertility of virgin dairy heifers. Am. J. Vet. Res. 1986, 47, 1395–1397. [Google Scholar] [PubMed]
- Weaver, G.A.; Kurtz, H.J.; Behrens, J.C.; Robison, T.S.; Seguin, B.E.; Bates, F.Y.; Mirocha, C.J. Effect of zearalenone on dairy cows. Am. J. Vet. Res. 1986, 47, 1826–1828. [Google Scholar] [PubMed]
- Coppock, R.W.; Mostrom, M.S.; Sparling, C.G.; Jacobsen, B.; Ross, S.C. Apparent zearalenone intoxication in a dairy herd from feeding spoiled acid-treated corn. Vet. Hum. Toxicol. 1990, 32, 246–248. [Google Scholar] [PubMed]
- Smith, J.F.; di Menna, M.E.; McGowan, L.T. Reproductive performance of Coopworth ewes following oral doses of zearalenone before and after mating. J. Reprod. Fertil. 1990, 89, 99–106. [Google Scholar] [CrossRef] [PubMed]
- Höhler, D.; Südekum, K.H.; Wolffram, S.; Frohlich, A.; Marquardt, R.R.; Ho, D. Metabolism and excretion of ochratoxin A fed to sheep. J. Anim. Sci. 1999, 77, 1217–1223. [Google Scholar] [PubMed]
- Mohr, A.I.; Lorenz, I.; Baum, B.; Hewicker-Trautwein, M.; Pfaffl, M.; Džidić, A.; Meyer, H.H.D.; Bauer, J.; Meyer, K. Influence of oral application of mycophenolic acid on the clinical health status of sheep. J. Vet. Med. Ser. A Physiol. Pathol. Clin. Med. 2007, 54, 76–81. [Google Scholar] [CrossRef] [PubMed]
- Dzidic, A.; Prgomet, C.; Mohr, A.; Meyer, K.; Bauer, J.; Meyer, H.H.D.; Pfaffl, M.W. Effects of mycophenolic acid on inosine monophosphate dehydrogenase I and II mRNA expression in white blood cells and various tissues in sheep. J. Vet. Med. Ser. A 2006, 53, 163–169. [Google Scholar] [CrossRef] [PubMed]
- Dzidic, A.; Meyer, H.H.D.; Bauer, J.; Pfaffl, M.W. Long-term effects of mycophenolic acid on the immunoglobulin and inflammatory marker-gene expression in sheep white blood cells. Mycotoxin Res. 2010, 26, 235–240. [Google Scholar] [CrossRef] [PubMed]
- Haggblom, P. Isolation of roquefortine C from feed grain. Appl. Environ. Microbiol. 1990, 56, 2924–2926. [Google Scholar] [PubMed]
- Lloyd, W.E. Citrinin and ochratoxin toxicoses in cattle in the United States. In Proceedings of the 2nd International Symposium of Veterinary Laboratory Diagnosticians, Lucerne, Switzerland, 24–26 June 1980; pp. 435–439.
- Sabater-Vilar, M.; Maas, R.F.M.; de Bosschere, H.; Ducatelle, R.; Fink-Gremmels, J. Patulin produced by an Aspergillus clavatus isolated from feed containing malting residues associated with a lethal neurotoxicosis in cattle. Mycopathologia 2004, 158, 419–426. [Google Scholar] [CrossRef] [PubMed]
- Pedrosa, K.; Borutova, R. Synergistic effects of mycotoxins discussed. Feedstuffs Repr. 2011, 83, 1–3. [Google Scholar]
- Speijers, G.J.A.; Speijers, M.H.M. Combined toxic effects of mycotoxins. Toxicol. Lett. 2004, 153, 91–98. [Google Scholar] [CrossRef] [PubMed]
- Ostry, V. Alternaria mycotoxins: An overview of chemical characterization, producers, toxicity, analysis and occurrence in foodstuffs. World Mycotoxin J. 2008, 1, 175–188. [Google Scholar] [CrossRef]
- Wang, W.; Jones, C.; Ciacci-Zanella, J.; Holt, T.; Gilchrist, D.G.; Dickman, M.B. Fumonisins and Alternaria alternata lycopersici toxins: Sphinganine analog mycotoxins induce apoptosis in monkey kidney cells. Proc. Natl. Acad. Sci. USA 1996, 93, 3461–3465. [Google Scholar] [CrossRef] [PubMed]
- Uhlig, S.; Eriksen, G.S.; Hofgaard, I.S.; Krska, R.; Beltrán, E.; Sulyok, M. Faces of a changing climate: Semi-quantitative multi-mycotoxin analysis of grain grown in exceptional climatic conditions in Norway. Toxins 2013, 5, 1682–1697. [Google Scholar] [CrossRef] [PubMed]
- Streit, E.; Schwab, C.; Sulyok, M.; Naehrer, K.; Krska, R.; Schatzmayr, G. Multi-mycotoxin screening reveals the occurrence of 139 different secondary metabolites in feed and feed ingredients. Toxins 2013, 5, 504–523. [Google Scholar] [CrossRef] [PubMed]
- Santos, R.R.; Schoevers, E.J.; Wu, X.; Roelen, B.A.J.; Fink-Gremmels, J. The protective effect of follicular fluid against the emerging mycotoxins alternariol and beauvericin. World Mycotoxin J. 2015, 1, 1–6. [Google Scholar] [CrossRef]
- Siciliano, I.; Ortu, G.; Gilardi, G.; Gullino, M.; Garibaldi, A. Mycotoxin Production in Liquid Culture and on Plants Infected with Alternaria spp. Isolated from Rocket and Cabbage. Toxins 2015, 7, 743–754. [Google Scholar] [CrossRef] [PubMed]
- International Agency for Research on Cancer (IARC). Aflatoxins. IARC Summ. Evaluation 2002, 82, 1–171. [Google Scholar]
- Gallo, A.; Moschini, M.; Masoero, F. Aflatoxins absorption in the gastro-intestinal tract and in the vaginal mucosa in lactating dairy cows. Ital. J. Anim. Sci. 2008, 7, 53–63. [Google Scholar] [CrossRef]
- Battacone, G.; Nudda, A.; Rassu, S.P.G.; Decandia, M.; Pulina, G. Excretion pattern of aflatoxin M1 in milk of goats fed a single dose of aflatoxin B1. J. Dairy Sci. 2012, 95, 2656–2661. [Google Scholar] [CrossRef] [PubMed]
- Battacone, G.; Nudda, A.; Palomba, M.; Mazzette, A.; Pulina, G. The transfer of aflatoxin M1 in milk of ewes fed diet naturally contaminated by aflatoxins and effect of inclusion of dried yeast culture in the diet. J. Dairy Sci. 2009, 92, 4997–5004. [Google Scholar] [PubMed]
- Veldman, A.; Meijs, J.A.C.; Borggreve, G.J.; Heeres-van der Tol, J.J. Carry-over of aflatoxin from cows’ food to milk. Anim. Prod. 1992, 55, 163–168. [Google Scholar] [CrossRef]
- European Food Safety Authority (EFSA). Opinion of the Scientific Panel on Contaminants in the Food Chain on a request from the Commission related to Aflatoxin B1 as undesirable substance in animal feed. EFSA J. 2004, 39, 1–27. [Google Scholar]
- Miller, D.M.; Wilson, D.M.; Eaton, D.L.; Groopman, J.D. Veterinary diseases related to aflatoxins. In The Toxicology of Aflatoxins: Human Health, Veterinary, and Agricultural Significance; Eaton, D.L., Groopman, J.D., Eds.; Academic Press: San Diego, CA, USA, 1993; pp. 347–364. [Google Scholar]
- Pier, A.C. Major biological consequences of aflatoxicosis in animal production. J. Anim. Sci. 1992, 70, 3964–3967. [Google Scholar] [PubMed]
- Bernabucci, U.; Colavecchia, L.; Danieli, P.P.; Basiricò, L.; Lacetera, N.; Nardone, A.; Ronchi, B. Aflatoxin B1 and fumonisin B1 affect the oxidative status of bovine peripheral blood mononuclear cells. Toxicol. Vitr. 2011, 25, 684–691. [Google Scholar] [CrossRef] [PubMed]
- Flores-Flores, M.E.; Lizarraga, E.; López de Cerain, A.; González-Peñas, E. Presence of mycotoxins in animal milk: A review. Food Control 2015, 53, 163–176. [Google Scholar] [CrossRef]
- Moschini, M.; Gallo, A.; Piva, G.; Masoero, F. The effects of rumen fluid on the in vitro aflatoxin binding capacity of different sequestering agents and in vivo release of the sequestered toxin. Anim. Feed Sci. Technol. 2008, 147, 292–309. [Google Scholar] [CrossRef]
- Wu, Q.; Jezkova, A.; Yuan, Z.; Pavlikova, L.; Dohnal, V.; Kuca, K. Biological degradation of aflatoxins. Drug Metab. Rev. 2009, 41, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Diaz, D.E.; Smith, T.K. Mycotoxin sequestering agents: Practical tools for the neutralisation of mycotoxins. In The Mycotoxin Blue Book; Nottingham University Press: Thrumpton, Nottingham, UK, 2005; pp. 323–339. [Google Scholar]
- Kroll, M.; Arenzana-Seisdedos, F.; Bachelerie, F.; Thomas, D.; Friguet, B.; Conconi, M. The secondary fungal metabolite gliotoxin targets proteolytic activities of the proteasome. Chem. Biol. 1999, 6, 689–698. [Google Scholar] [CrossRef] [PubMed]
- Tsunawaki, S.; Yoshida, L.S.; Nishida, S.; Kobayashi, T.; Shimoyama, T. Fungal metabolite gliotoxin inhibits assembly of the human respiratory burst NADPH oxidase. Infect. Immun. 2004, 72, 3373–3382. [Google Scholar] [CrossRef] [PubMed]
- Whitlow, L.; Hagler, W.M. Mycotoxins in dairy cattle: Occurrence, toxicity, prevention and treatment. Proc. Southwest Nutr. Conf 2005, 124–138. [Google Scholar]
- Huang, X.; Chu, F.S. Production and characterization of monoclonal and polyclonal antibodies against the mycotoxin cyclopiazonic acid. J. Agric. Food Chem. 1993, 41, 329–333. [Google Scholar] [CrossRef]
- European Food Safety Authority (EFSA). Opinion of the Scientific Panel on contaminants in the food chain [CONTAM] related to ochratoxin A (OTA) as undesirable substance in animal feed. EFSA J. 2004, 1–36. [Google Scholar]
- Hult, K.; Teiling, A.; Gatenbeck, S. Degradation of ochratoxin A by a ruminant. Appl. Environ. Microbiol. 1976, 32, 443–444. [Google Scholar] [PubMed]
- Battacone, G.; Nudda, A.; Pulina, G. Effects of ochratoxin a on livestock production. Toxins 2010, 2, 1796–1824. [Google Scholar] [CrossRef] [PubMed]
- Mobashar, M.; Blank, R.; Hummel, J.; Westphal, A.; Tholen, E.; Südekum, K.-H. Ruminal ochratoxin A degradation—Contribution of the different microbial populations and influence of diet. Anim. Feed Sci. Technol. 2012, 171, 85–97. [Google Scholar] [CrossRef]
- Müller, H.M.; Lerch, C.; Müller, K.; Eggert, W. Kinetic profiles of ochratoxin A and ochratoxin alpha during in vitro incubation in buffered forestomach and abomasal contents from cows. Nat. Toxins 1998, 6, 251–258. [Google Scholar] [CrossRef] [PubMed]
- Sreemannarayana, O.; Frohlich, A.A.; Vitti, T.G.; Marquardt, R.R.; Abramson, D. Studies of the Tolerance and Disposition of Ochratoxin a in Young Calves. J. Anim. Sci. 1988, 66, 1703–1711. [Google Scholar] [PubMed]
- Blank, R.; Rolfs, J.-P.; Südekum, K.-H.; Frohlich, A.A.; Marquardt, R.R.; Wolffram, S. Effects of chronic ingestion of ochratoxin a on blood levels and excretion of the mycotoxin in sheep. J. Agric. Food Chem. 2003, 51, 6899–6905. [Google Scholar] [CrossRef] [PubMed]
- Denli, M.; Perez, J.F. Ochratoxins in feed, a risk for animal and human health: Control strategies. Toxins 2010, 2, 1065–1077. [Google Scholar] [CrossRef] [PubMed]
- Boudra, H.; Saivin, S.; Buffiere, C.; Morgavi, D.P. Short communication: Toxicokinetics of ochratoxin A in dairy ewes and carryover to milk following a single or long-term ingestion of contaminated feed. J. Dairy Sci. 2013, 96, 6690–6696. [Google Scholar] [CrossRef] [PubMed]
- Ribelin, W.E.; Fukushima, K.; Still, P.E. The toxicity of ochratoxin to ruminants. Can. J. Comp. Med. 1978, 42, 172–176. [Google Scholar] [PubMed]
- Niederberger, M.; Oevermann, A.; Kirscher, F.; Meylan, M. Tremorgenic syndrome in a cattle herd after feeding silage contaminated with A. clavatus. Schweiz. Arch. Tierheilkd. 2011, 153, 105–110. [Google Scholar] [CrossRef] [PubMed]
- Suzuki, T.; Takeda, M.; Tanabe, H. A New Mycotoxin produced by Aspergillus clavatus. Chem. Pharm. Bull. Tokyo 1971, 19, 1786–1788. [Google Scholar] [CrossRef] [PubMed]
- Voss, K.A.; Smith, G.W.; Haschek, W.M. Fumonisins: Toxicokinetics, mechanism of action and toxicity. Anim. Feed Sci. Technol. 2007, 137, 299–325. [Google Scholar] [CrossRef]
- Guerre, P. Fusariotoxins in avian species: Toxicokinetics, metabolism and persistence in tissues. Toxins 2015, 7, 2289–2305. [Google Scholar] [CrossRef] [PubMed]
- Antonissen, G.; Martel, A.; Pasmans, F.; Ducatelle, R.; Verbrugghe, E.; Vandenbroucke, V.; Li, S.; Haesebrouck, F.; van Immerseel, F.; Croubels, S. The impact of Fusarium Mycotoxins on human and animal host susceptibility to infectious diseases. Toxins 2014, 6, 430–452. [Google Scholar] [CrossRef] [PubMed]
- Marczuk, J.; Obremski, K.; Lutnicki, K.; Gajecka, M.; Gajecki, M. Zearalenone and deoxynivalenol mycotoxicosis in dairy cattle herds. Pol. J. Vet. Sci. 2012, 15, 365–372. [Google Scholar] [CrossRef] [PubMed]
- Morgavi, D.P.; Riley, R.T. An historical overview of field disease outbreaks known or suspected to be caused by consumption of feeds contaminated with Fusarium toxins. Anim. Feed Sci. Technol. 2007, 137, 201–212. [Google Scholar] [CrossRef]
- European Food Safety Authority (EFSA). Opinion of the Scientific Panel on contaminants in the food chain [CONTAM] related to Deoxynivalenol (DON) as undesirable substance in animal feed. EFSA J. 2004, 73, 1–42. [Google Scholar]
- Côté, L.M.; Dahlem, A.M.; Yoshizawa, T.; Swanson, S.P.; Buck, W.B. Excretion of deoxynivalenol and its metabolite in milk, urine, and feces of lactating dairy cows. J. Dairy Sci. 1986, 69, 2416–2423. [Google Scholar] [CrossRef]
- Seeling, K.; Lebzien, P.; Dänicke, S.; Spilke, J.; Südekum, K.-H.; Flachowsky, G. Effects of level of feed intake and Fusarium toxin-contaminated wheat on rumen fermentation as well as on blood and milk parameters in cows. J. Anim. Physiol. Anim. Nutr. Berl 2006, 90, 103–115. [Google Scholar] [CrossRef] [PubMed]
- Hedman, R.; Pettersson, H. Transformation of nivalenol by gastrointestinal microbes. Arch. Tierernahr. 1997, 50, 321–329. [Google Scholar] [CrossRef] [PubMed]
- European Food Safety Authority (EFSA). Scientific Opinion on risks for animal and public health related to the presence of nivalenol in food and feed1. EFSA J. 2013, 11, 1–119. [Google Scholar]
- European Food Safety Authority (EFSA). Scientific Opinion on the risks for animal and public health related to the presence of T-2 and HT-2 toxin in food and feed. EFSA J. 2011, 9, 1–187. [Google Scholar]
- Hsu, I.C.; Smalley, E.B.; Strong, F.M.; Ribelin, W.E. Identification of T-2 toxin in moldy corn associated with a lethal toxicosis in dairy cattle. Appl. Microbiol. 1972, 24, 684–690. [Google Scholar] [PubMed]
- Weaver, G.A.; Kurtz, H.J.; Mirocha, C.J.; Bates, F.Y.; Behrens, J.C.; Robison, T.S.; Swanson, S.P. The failure of purified T-2 mycotoxin to produce hemorrhaging in dairy cattle. Can. Vet. J. 1980, 21, 210–213. [Google Scholar] [PubMed]
- Mann, D.D.; Buening, G.M.; Hook, B.; Osweiler, G.D. Effects of T-2 mycotoxin on bovine serum proteins. Am. J. Vet. Res. 1983, 44, 1757–1759. [Google Scholar] [PubMed]
- Mann, D.D.; Buening, G.M.; Osweiler, G.D.; Hook, B.S. Effect of subclinical levels of T-2 toxin on the bovine cellular immune system. Can. J. Comp. Med. 1984, 48, 308–312. [Google Scholar] [PubMed]
- Buening, G.M.; Mann, D.D.; Hook, B.; Osweiler, G.D. The effect of T-2 toxin on the bovine immune system: Cellular factors. Vet. Immunol. Immunopathol. 1982, 3, 411–417. [Google Scholar] [CrossRef]
- Yoshizawa, T.; Mirocha, C.J.; Behrens, J.C.; Swanson, S.P. Metabolic fate of T-2 toxin in a lactating cow. Food Cosmet. Toxicol. 1981, 19, 31–39. [Google Scholar] [CrossRef]
- Alm, K.; Dahlbom, M.; Säynäjärvi, M.; Andersson, M.A.; Salkinoja-Salonen, M.S.; Andersson, M.C. Impaired semen quality of AI bulls fed with moldy hay: A case report. Theriogenology 2002, 58, 1497–1502. [Google Scholar] [CrossRef]
- Spotti, M.; Caloni, F.; Fracchiolla, L.; Pompa, G.; Vigo, D.; Maffeo, G. Fumonisin B1 carry-over into milk in the isolated perfused bovine udder. Vet. Hum. Toxicol. 2001, 43, 109–111. [Google Scholar] [PubMed]
- European Food Safety Authority (EFSA). Opinion of the Scientific Panel on contaminants in the food chain [CONTAM] related to fumonisins as undesirable substances in animal feed. EFSA J. 2005, 235, 1–32. [Google Scholar]
- Caloni, F.; Spotti, M.; Auerbach, H.; Op den Camp, H.; Gremmels, J.F.; Pompa, G. In vitro metabolism of fumonisin B1 by ruminal microflora. Vet. Res. Commun. 2000, 24, 379–387. [Google Scholar] [CrossRef] [PubMed]
- Kennedy, D.G.; Hewitt, S.A.; McEvoy, J.D.; Currie, J.W.; Cannavan, A.; Blanchflower, W.J.; Elliot, C.T. Zeranol is formed from Fusarium spp. toxins in cattle in vivo. Food Addit. Contam. 1998, 15, 393–400. [Google Scholar] [CrossRef] [PubMed]
- Minervini, F.; Dell’Aquila, M.E. Zearalenone and reproductive function in farm animals. Int. J. Mol. Sci. 2008, 9, 2570–2584. [Google Scholar] [CrossRef] [PubMed]
- European Food Safety Authority (EFSA). Opinion of the Scientific Panel on contaminants in the food chain [CONTAM] related to Zearalenone as undesirable substance in animal feed. EFSA J. 2004, 89, 1–35. [Google Scholar]
- Fushimi, Y.; Takagi, M.; Hasunuma, H.; Uno, S.; Kokushi, E.; Watanabe, U.; Liu, J.; Marey, M.A.; Miyamoto, A.; Otoi, T.; et al. Application of mycotoxin adsorbent to cattle feed contaminated with zearalenone: Urinary zearalenone excretion and association with anti-Müllerian hormone. World Mycotoxin J. 2014, 7, 367–378. [Google Scholar] [CrossRef]
- Takagi, M.; Uno, S.; Kokushi, E.; Shiga, S.; Mukai, S.; Kuriyagawa, T.; Takagaki, K.; Hasunuma, H.; Matsumoto, D.; Okamoto, K.; et al. Measurement of urinary zearalenone concentrations for monitoring natural feed contamination in cattle herds: On-farm trials. J. Anim. Sci. 2011, 89, 287–296. [Google Scholar] [CrossRef] [PubMed]
- Takagi, M.; Hirai, T.; Shiga, S.; Uno, S.; Kokushi, E. Relationship between urinary zearalenone concentration and embryo production in superovulated cattle. Arch. Tierernahr. 2013, 56, 360–366. [Google Scholar]
- Pereyra, M.L.G.; Alonso, V.A.; Sager, R.; Morlaco, M.B.; Magnoli, C.E.; Astoreca, A.L.; Rosa, C.A.R.; Chiacchiera, S.M.; Dalcero, A.M.; Cavaglieri, L.R. Fungi and selected mycotoxins from pre- and postfermented corn silage. J. Appl. Microbiol. 2008, 104, 1034–1041. [Google Scholar] [CrossRef]
- Veselý, D.; Veselá, D.; Adámková, M. Occurrence of PR-toxin-producing Penicillium roqueforti in corn silage. Vet. Med. Praha 1981, 26, 109–115. [Google Scholar] [PubMed]
- Oh, S.Y.; Balch, C.G.; Cliff, R.L.; Sharma, B.S.; Boermans, H.J.; Swamy, H.V.L.N.; Quinton, V.M.; Karrow, N.A. Exposure to Penicillium mycotoxins alters gene expression of enzymes involved in the epigenetic regulation of bovine macrophages (BoMacs). Mycotoxin Res. 2013, 29, 235–243. [Google Scholar] [CrossRef] [PubMed]
- Solfrizzo, M.; Gambacorta, L.; Lattanzio, V.M.T.; Powers, S.; Visconti, A. Simultaneous LC-MS/MS determination of aflatoxin M1, ochratoxin A, deoxynivalenol, de-epoxydeoxynivalenol, α and β-zearalenols and fumonisin B1 in urine as a multi-biomarker method to assess exposure to mycotoxins. Anal. Bioanal. Chem. 2011, 401, 2831–2841. [Google Scholar] [CrossRef] [PubMed]
- Meky, F.; Turner, P.C.; Ashcroft, A.E.; Miller, J.D.; Qiao, Y.-L.; Roth, M.J.; Wild, C.P. Development of a urinary biomarker of human exposure to deoxynivalenol. Food Chem. Toxicol. 2003, 41, 265–273. [Google Scholar] [CrossRef]
- Ediage, E.N.; di Mavungu, J.D.; Song, S.; Wu, A.; van Peteghem, C.; de Saeger, S. A direct assessment of mycotoxin biomarkers in human urine samples by liquid chromatography tandem mass spectrometry. Anal. Chim. Acta 2012, 741, 58–69. [Google Scholar] [CrossRef] [PubMed]
- Choi, J.; Mørck, T.A.; Polcher, A.; Knudsen, L.E.; Joas, A. Review of the state of the art of human biomonitoring for chemical substances and its application to human exposure assessment for food. EFSA J. 2015, 1–321. [Google Scholar]
- Solfrizzo, M.; Gambacorta, L.; Visconti, A. Assessment of multi-mycotoxin exposure in southern Italy by urinary multi-biomarker determination. Toxins 2014, 6, 523–538. [Google Scholar] [CrossRef] [PubMed]
- Tiwary, A.K.; Puschner, B.; Poppenga, R.H. Using roquefortine C as a biomarker for penitrem A intoxication. J. Vet. Diagn. Investig. 2009, 21, 237–239. [Google Scholar] [CrossRef]
- Pedrosa, K.; Griessler, K. Toxicity, occurrence and negative effects of PR toxin—The hidden enemy. Available online: https://www.yumpu.com/en/document/view/34544639/download (accessed on 13 May 2015).
- European Food Safety Authority (EFSA). Scientific Opinion on the risks for public and animal health related to the presence of citrinin in food and feed. EFSA J. 2012, 10, 1–82. [Google Scholar]
- Bouslimi, A.; Bouaziz, C.; Ayed-Boussema, I.; Hassen, W.; Bacha, H. Individual and combined effects of ochratoxin A and citrinin on viability and DNA fragmentation in cultured Vero cells and on chromosome aberrations in mice bone marrow cells. Toxicology 2008, 251, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Griffiths, I.; Done, S. Citrinin as a possible cause of the pruritis, pyrexia, haemorrhagic syndrome in cattle. Vet. Rec. 1991, 129, 113–117. [Google Scholar] [CrossRef] [PubMed]
- Stec, J.A.N.; Rachubik, J.Ł.A.W.; Szczotka, M.; Mak, J.K.U.Ź. Effects of penicillium mycotoxins: Citrin, ochratoxin A, and patulin on in vitro proliferation of bovine lymphocytes. Bull. Vet. Inst. Pulawy 2008, 52, 163–167. [Google Scholar]
- Grenier, B.; Applegate, T.J. Modulation of intestinal functions following mycotoxin ingestion: Meta-analysis of published experiments in animals. Toxins 2013, 5, 396–430. [Google Scholar] [CrossRef] [PubMed]
- Bouhet, S.; Oswald, I.P. The effects of mycotoxins, fungal food contaminants, on the intestinal epithelial cell-derived innate immune response. Vet. Immunol. Immunopathol. 2005, 108, 199–209. [Google Scholar] [CrossRef] [PubMed]
- Maresca, M.; Fantini, J. Some food-associated mycotoxins as potential risk factors in humans predisposed to chronic intestinal inflammatory diseases. Toxicon 2010, 56, 282–294. [Google Scholar] [CrossRef] [PubMed]
- Cleveland, T.E.; Dowd, P.F.; Desjardins, A.E.; Bhatnagar, D.; Cotty, P.J. United States Department of Agriculture—Agricultural Research Service research on pre-harvest prevention of mycotoxins and mycotoxigenic fungi in US crops. Pest Manag. Sci. 2003, 59, 629–642. [Google Scholar] [CrossRef] [PubMed]
- Shinha, K.K.; Bhatnagar, D. Mycotoxins in Agriculture and Food Safety; Marcel Dekker, Inc.: New York, NY, USA, 1998. [Google Scholar]
- Eeckhout, M.; Landschoot, S.; Deschuyffeleer, N.; de Laethauwer, S.; Haesaert, G. Guidelines for prevention and control of mould growth and mycotoxin production in cereals. Available online: http://synagra.be/Download.ashx?ID=6421 (accessed on 14 June 2015).
- Food and Agriculture Organization of the United Nations (FAO). Available online: http://www.codexalimentarius.org/input/download/standards/10099/CXP_056e.pdf (accessed on 14 July 2004).
- Jard, G.; Liboz, T.; Mathieu, F.; Guyonvarc’h, A.; Lebrihi, A. Review of mycotoxin reduction in food and feed: From prevention in the field to detoxification by adsorption or transformation. Food Addit. Contam. A 2011, 28, 1590–1609. [Google Scholar] [CrossRef] [PubMed]
- Edwards, S.G. Influence of agricultural practices on fusarium infection of cereals and subsequent contamination of grain by trichothecene mycotoxins. Toxicol. Lett. 2004, 153, 29–35. [Google Scholar] [CrossRef] [PubMed]
- Schrödter, R. Influence of harvest and storage conditions on trichothecenes levels in various cereals. Toxicol. Lett. 2004, 153, 47–49. [Google Scholar] [CrossRef] [PubMed]
- Chulze, S.N. Strategies to reduce mycotoxin levels in maize during storage: A review. Food Addit. Contam. A 2010, 27, 651–657. [Google Scholar] [CrossRef] [PubMed]
- Sabatini, A.; Danieli, P.P.; Bernabucci, U.; Ronchi, B. Evaluation of mycotoxins contamination in intensive beef cattle production system. Italy J. Anim. Sci. 2010, 6, 466–468. [Google Scholar]
- Oude Elferink, S.J.W.H.; Driehuis, F.; Gottschal, J.; Spoelstra, S. Paper 2.0: Silage fermentation processes and their manipulation. FAO Plant P 2000, 161, 17–30. [Google Scholar]
- Muck, R. Recent advances in silage microbiology. Agric. Food Sci. 2013, 22, 3–15. [Google Scholar]
- Ali, M.; Cone, J.W.; Khan, N.A.; Hendriks, W.H.; Struik, P.C. Effect of temperature and duration of ensiling on in vitro degradation of maize silages in rumen fluid. J. Anim. Physiol. Anim. Nutr. Berl 2015, 99, 251–257. [Google Scholar] [CrossRef] [PubMed]
- Orosz, S.; Wilkinson, J.M.; Wigley, S.; Bíró, Z.; Galló, J. Microbial status, aerobic stability and fermentation of maize silage sealed with an oxygen barrier film or standard polyethylene film. Agric. Food Sci. 2013, 22, 182–188. [Google Scholar]
- Kung, L.; Sheperd, A.C.; Smagala, A.M.; Endres, K.M.; Bessett, C.A.; Ranjit, N.K.; Glancey, J.L. The effect of preservatives based on propionic acid on the fermentation and aerobic stability of corn silage and a total mixed ration. J. Dairy Sci. 1998, 81, 1322–1330. [Google Scholar] [CrossRef]
- Ranjit, N.K.; Kung, L. The effect of Lactobacillus buchneri, Lactobacillus plantarum, or a chemical preservative on the fermentation and aerobic stability of corn silage. J. Dairy Sci. 2000, 83, 526–535. [Google Scholar] [CrossRef]
- European Food Safety Authority (EFSA). Scientific Opinion on the safety and efficacy of propionic acid, sodium propionate, calcium propionate and ammonium propionate for all animal species. EFSA J. 2011, 9, 1–22. [Google Scholar]
- Tabacco, E.; Piano, S.; Revello-Chion, A.; Borreani, G. Effect of Lactobacillus buchneri LN4637 and Lactobacillus buchneri LN40177 on the aerobic stability, fermentation products, and microbial populations of corn silage under farm conditions. J. Dairy Sci. 2011, 94, 5589–5598. [Google Scholar] [CrossRef] [PubMed]
- Rust, S.R.; Kim, H.S.; Enders, G.L. Effects of a microbial inoculant on fermentation characteristics and nutritional value of corn silage. J. Prod. Agric. 1989, 2, 235–241. [Google Scholar] [CrossRef]
- Weinberg, Z.G.; Ashbell, G.; Hen, Y.; Azrieli, A. The effect of applying lactic acid bacteria at ensiling on the aerobic stability of silages. J. Appl. Bacteriol. 1993, 75, 512–518. [Google Scholar] [CrossRef]
- EFSA. Scientific Opinion on the safety and efficacy of Lactobacillus brevis (DSM 23231), Lactobacillus buchneri (DSM 22501), Lactobacillus buchneri (NCIMB 40788—CNCM I—4323), Lactobacillus buchneri (ATCC PTA—6138) and Lactobacillus buchneri (ATCC PPTA—2). EFSA J. 2013, 11, 1–16. [Google Scholar]
- Woolford, M.K. The Silage Fermentation; Marcel Dekker, Inc.: NewYork, NY, USA, 1984. [Google Scholar]
- González Pereyra, M.L.; Chiacchiera, S.M.; Rosa, C.A.; Sager, R.; Dalcero, A.M.; Cavaglieri, L. Comparative analysis of the mycobiota and mycotoxins contaminating corn trench silos and silo bags. J. Sci. Food Agric. 2011, 91, 1474–1481. [Google Scholar] [CrossRef] [PubMed]
- Review of mycotoxin-detoxifying agents used as feed additives: Mode of action, efficacy and feed/food safety. Available online: http://www.efsa.europa.eu/it/supporting/doc/22e.pdf (accessed on 14 June 2009).
- Awad, W.A.; Ghareeb, K.; Bohm, J.; Zentek, J. Decontamination and detoxification strategies for the Fusarium mycotoxin deoxynivalenol in animal feed and the effectiveness of microbial biodegradation. Food Addit. Contam. A Chem. Anal. Control Expo. Risk Assess. 2010, 27, 510–520. [Google Scholar] [CrossRef] [PubMed]
- Galvano, F.; Piva, A.; Ritieni, A.; Galvano, G. Dietary Strategies to Counteract the Effects of Mycotoxins: A Review. J. Food Prot. 2001, 12, 120–131. [Google Scholar]
- Gallo, A.; Masoero, F.; Bertuzzi, T.; Piva, G.; Pietri, A. Effect of the inclusion of adsorbents on aflatoxin B1 quantification in animal feedstuffs. Food Addit. Contam. A 2010, 27, 54–63. [Google Scholar] [CrossRef] [PubMed]
- Ramos, A.J.; Hernández, E. Prevention of aflatoxicosis in farm animals by Means of hydrated sodium calcium aluminosilicate addition to feedstuffs: A review. Anim. Feed Sci. Technol. 1997, 65, 197–206. [Google Scholar] [CrossRef]
- Masoero, F.; Gallo, A.; Diaz, D.; Piva, G.; Moschini, M. Effects of the procedure of inclusion of a sequestering agent in the total mixed ration on proportional aflatoxin M1 excretion into milk of lactating dairy cows. Anim. Feed Sci. Technol. 2009, 150, 34–45. [Google Scholar] [CrossRef]
- Smith, E.E.; Phillips, T.D.; Ellis, J.A.; Harvey, R.B.; Kubena, L.F.; Thompson, J.; Newton, G. Dietary hydrated sodium calcium aluminosilicate reduction of aflatoxin M1 residue in dairy goat milk and effects on milk production and components. J. Anim. Sci. 1994, 72, 677–682. [Google Scholar] [PubMed]
- Phillips, T.D.; Clement, B.A.; Kubena, L.F.; Harvey, R.B. Detection and detoxification of aflatoxins: Prevention of aflatoxicosis and aflatoxin residues with hydrated sodium calcium aluminosilicate. Vet. Hum. Toxicol. 1990, 32 (Suppl.), 15–19. [Google Scholar] [PubMed]
- Kutz, R.E.; Sampson, J.D.; Pompeu, L.B.; Ledoux, D.R.; Spain, J.N.; Vázquez-Añón, M.; Rottinghaus, G.E. Efficacy of Solis, NovasilPlus, and MTB-100 to reduce aflatoxin M1 levels in milk of early to mid lactation dairy cows fed aflatoxin B1. J. Dairy Sci. 2009, 92, 3959–3963. [Google Scholar] [CrossRef] [PubMed]
- Queiroz, O.C.M.; Han, J.H.; Staples, C.R.; Adesogan, A.T. Effect of adding a mycotoxin-sequestering agent on milk aflatoxin M1 concentration and the performance and immune response of dairy cattle fed an aflatoxin B1 contaminated diet. J. Dairy Sci. 2012, 95, 5901–5908. [Google Scholar] [CrossRef] [PubMed]
- Galvano, F.; Pietri, A.; Bertuzzi, T.; Bognanno, M.; Chies, L.; de Angelis, A.; Galvano, M. Activated Carbons: In Vitro Affinity for Fumonisin B1 and Relation of Adsorption Ability to Physicochemical Parameters. J. Food Prot. 1997, 7, 985–991. [Google Scholar]
- Diaz, D.E.; Hagler, W.M.; Blackwelder, J.T.; Eve, J.A.; Hopkins, B.A.; Anderson, K.L.; Jones, F.T.; Whitlow, L.W. Aflatoxin Binders II: Reduction of aflatoxin M1 in milk by sequestering agents of cows consuming aflatoxin in feed. Mycopathologia 2004, 157, 233–241. [Google Scholar] [CrossRef] [PubMed]
- Pietri, A.; Bertuzzi, T.; Piva, G.; Binder, E.M.; Schatzmayr, D.; Rodrigues, I. Aflatoxin Transfer from Naturally Contaminated Feed to Milk of Dairy Cows and the Efficacy of a Mycotoxin Deactivating Product. Int. J. Dairy Sci. 2009, 4, 34–42. [Google Scholar]
- Grenier, B.; Bracarense, A.-P.F.L.; Schwartz, H.E.; Lucioli, J.; Cossalter, A.-M.; Moll, W.-D.; Schatzmayr, G.; Oswald, I.P. Biotransformation approaches to alleviate the effects induced by fusarium mycotoxins in swine. J. Agric. Food Chem. 2013, 61, 6711–6719. [Google Scholar] [CrossRef] [PubMed]
- Polonelli, L.; Giovati, L.; Magliani, W.; Conti, S.; Sforza, S.; Calabretta, A.; Casoli, C.; Ronzi, P.; Grilli, E.; Gallo, A.; et al. Vaccination of lactating dairy cows for the prevention of aflatoxin B 1 carry over in milk. PLoS One 2011, 6, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Döll, S.; Dänicke, S.; Valenta, H.; Flachowsky, G. ro studies on the evaluation of mycotoxin detoxifying agents for their efficacy on deoxynivalenol and zearalenone. Arch. Anim. Nutr. 2007, 58, 311–324. [Google Scholar]
- Kong, C.; Shin, S.; Kim, B. Evaluation of mycotoxin sequestering agents for aflatoxin and deoxynivalenol: An in vitro approach. Springerplus 2014, 3, 346–350. [Google Scholar] [CrossRef] [PubMed]
- Tomašević-Čanović, M.; Daković, A.; Rottinghaus, G.; Matijašević, S.; Duričić, M. Surfactant modified zeolites-new efficient adsorbents for mycotoxins. Microporous Mesoporous Mater. 2003, 61, 173–180. [Google Scholar] [CrossRef]
- Hahn, I.; Kunz-Vekiru, E.; Twarużek, M.; Grajewski, J.; Krska, R.; Berthiller, F. Aerobic and anaerobic in vitro testing of feed additives claiming to detoxify deoxynivalenol and zearalenone. Food Addit. Contam. A 2015, 1–12. [Google Scholar]
- Avantaggiato, G.; Solfrizzo, M.; Visconti, A. Recent advances on the use of adsorbent materials for detoxification of Fusarium mycotoxins. Food Addit. Contam. 2005, 22, 379–388. [Google Scholar] [CrossRef] [PubMed]
© 2015 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gallo, A.; Giuberti, G.; Frisvad, J.C.; Bertuzzi, T.; Nielsen, K.F. Review on Mycotoxin Issues in Ruminants: Occurrence in Forages, Effects of Mycotoxin Ingestion on Health Status and Animal Performance and Practical Strategies to Counteract Their Negative Effects. Toxins 2015, 7, 3057-3111. https://doi.org/10.3390/toxins7083057
Gallo A, Giuberti G, Frisvad JC, Bertuzzi T, Nielsen KF. Review on Mycotoxin Issues in Ruminants: Occurrence in Forages, Effects of Mycotoxin Ingestion on Health Status and Animal Performance and Practical Strategies to Counteract Their Negative Effects. Toxins. 2015; 7(8):3057-3111. https://doi.org/10.3390/toxins7083057
Chicago/Turabian StyleGallo, Antonio, Gianluca Giuberti, Jens C. Frisvad, Terenzio Bertuzzi, and Kristian F. Nielsen. 2015. "Review on Mycotoxin Issues in Ruminants: Occurrence in Forages, Effects of Mycotoxin Ingestion on Health Status and Animal Performance and Practical Strategies to Counteract Their Negative Effects" Toxins 7, no. 8: 3057-3111. https://doi.org/10.3390/toxins7083057
APA StyleGallo, A., Giuberti, G., Frisvad, J. C., Bertuzzi, T., & Nielsen, K. F. (2015). Review on Mycotoxin Issues in Ruminants: Occurrence in Forages, Effects of Mycotoxin Ingestion on Health Status and Animal Performance and Practical Strategies to Counteract Their Negative Effects. Toxins, 7(8), 3057-3111. https://doi.org/10.3390/toxins7083057