
Further Characterization of Glycine-Containing Microcystins from the McMurdo Dry Valleys of Antarctica

 ,
, 
Abstract
:
1. Introduction
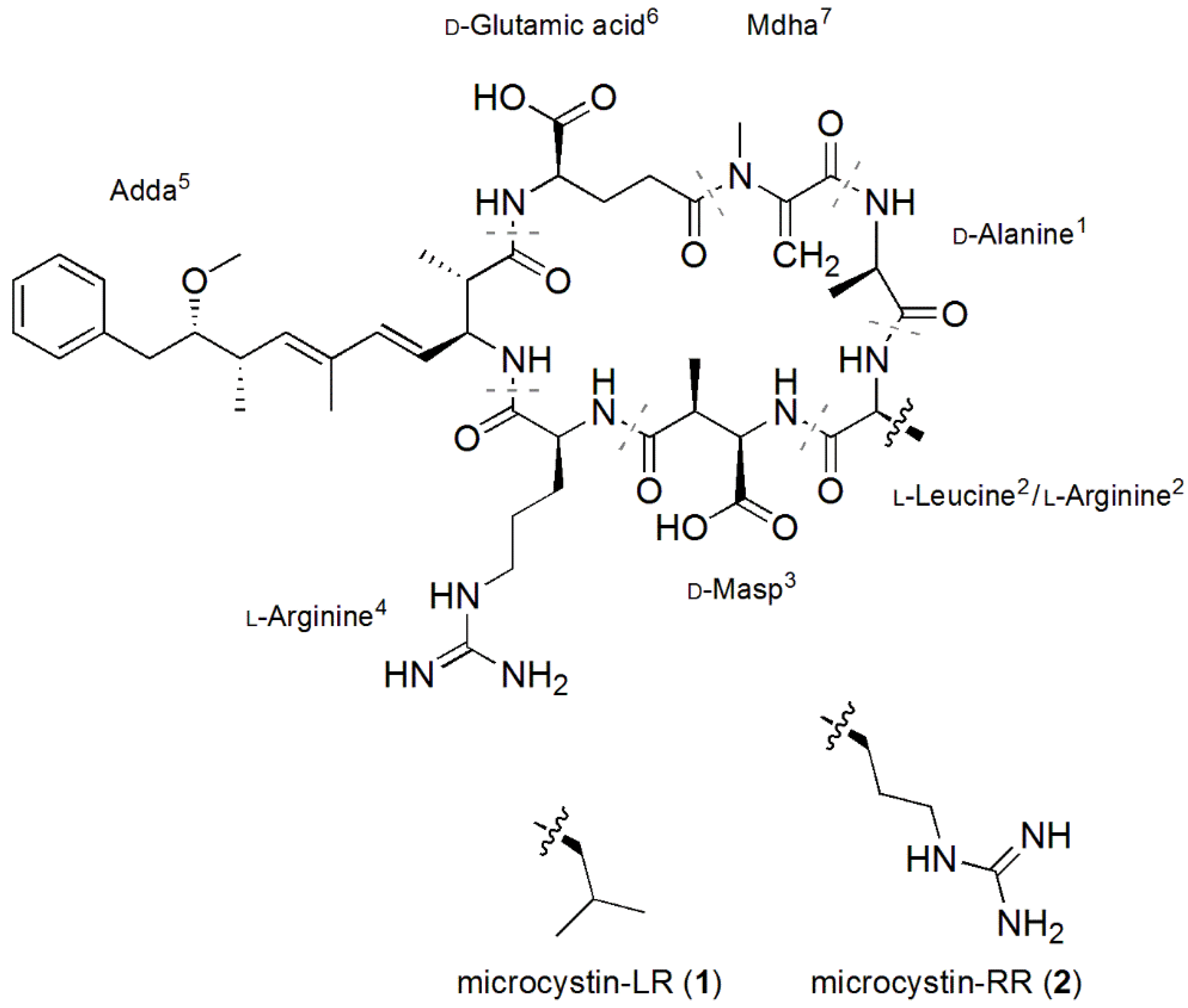
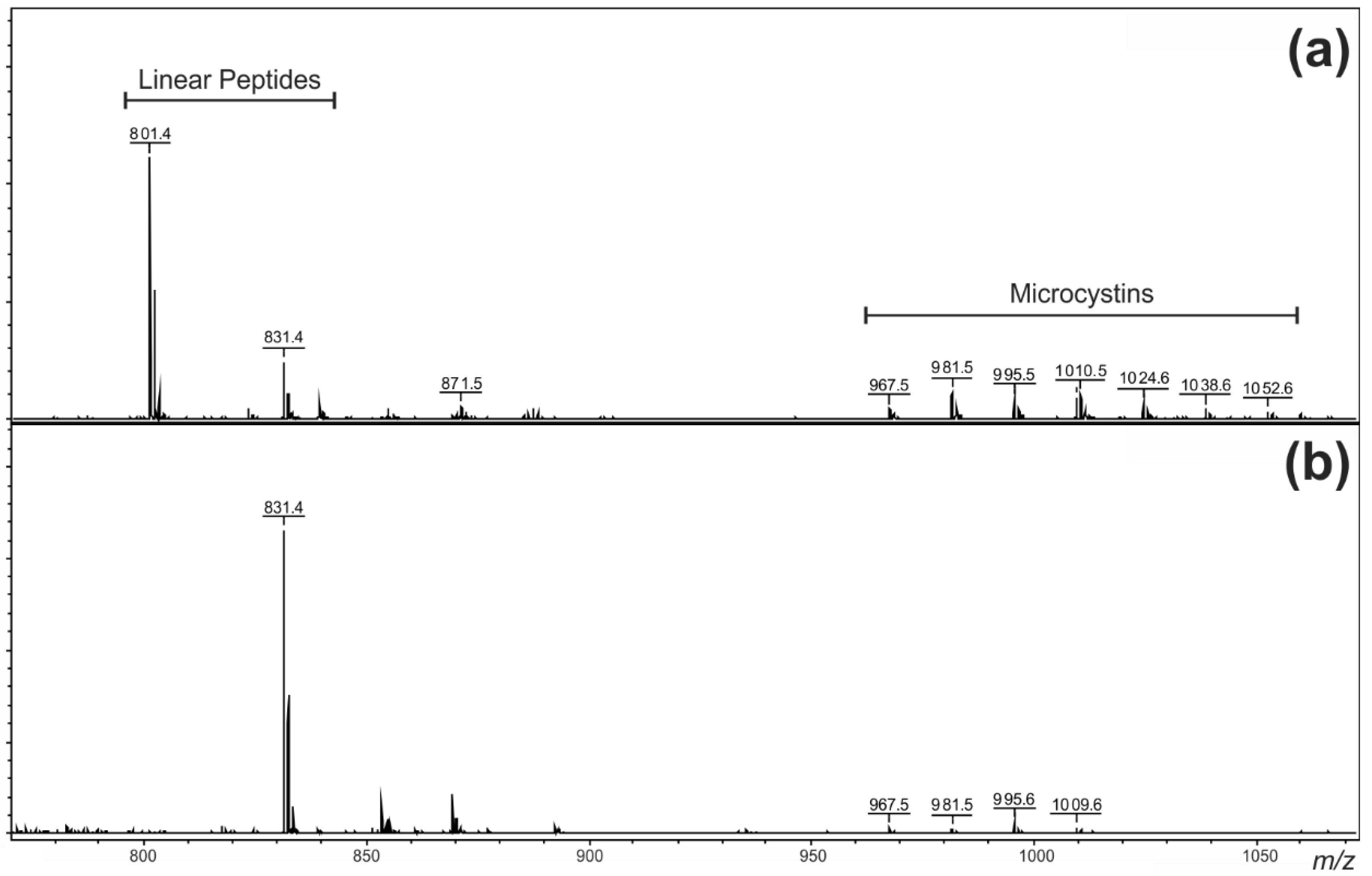
2. Results and Discussion
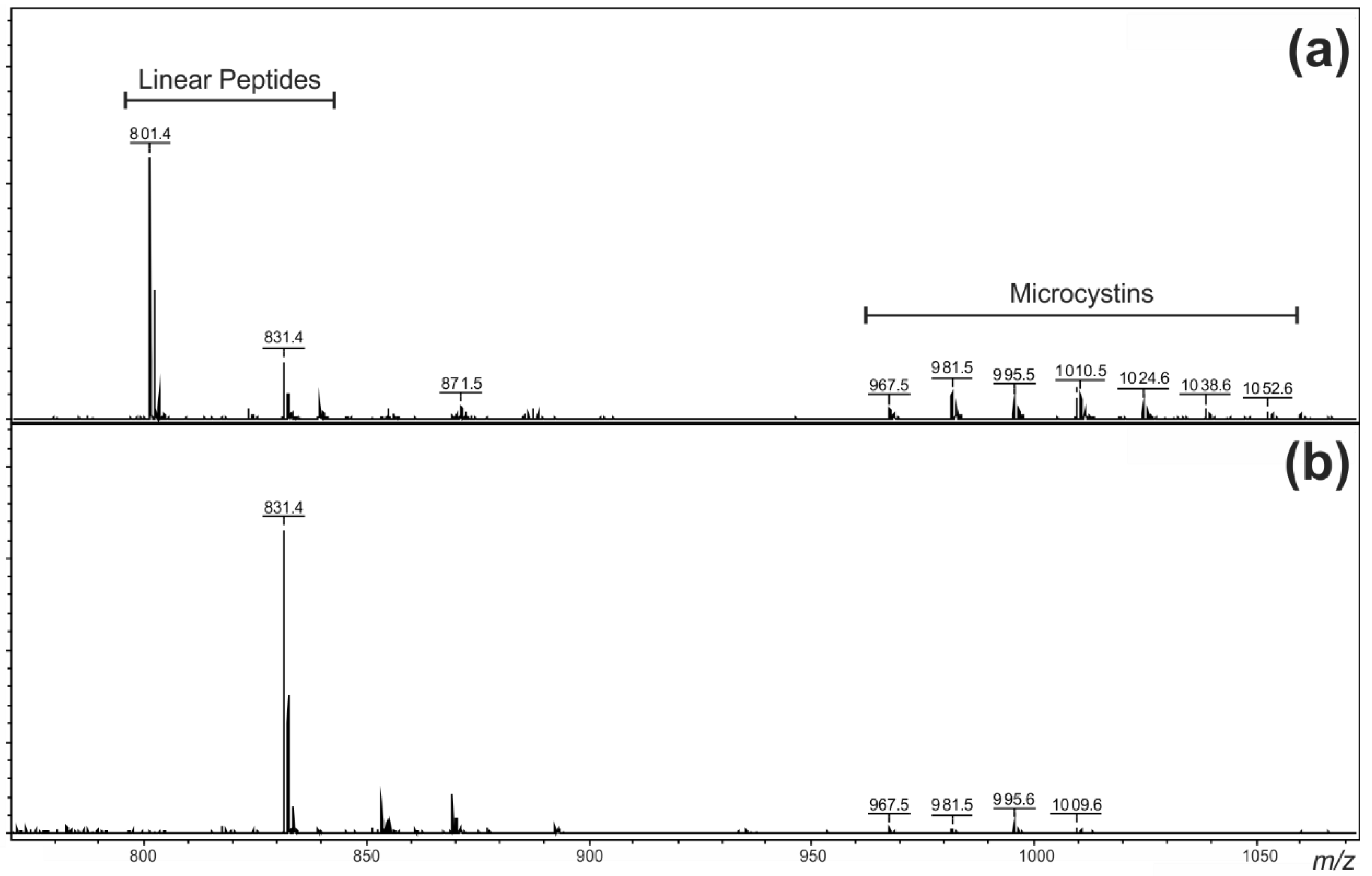
2.1. Oligopeptide Diversity in the Miers Valley Cyanobacterial Mats
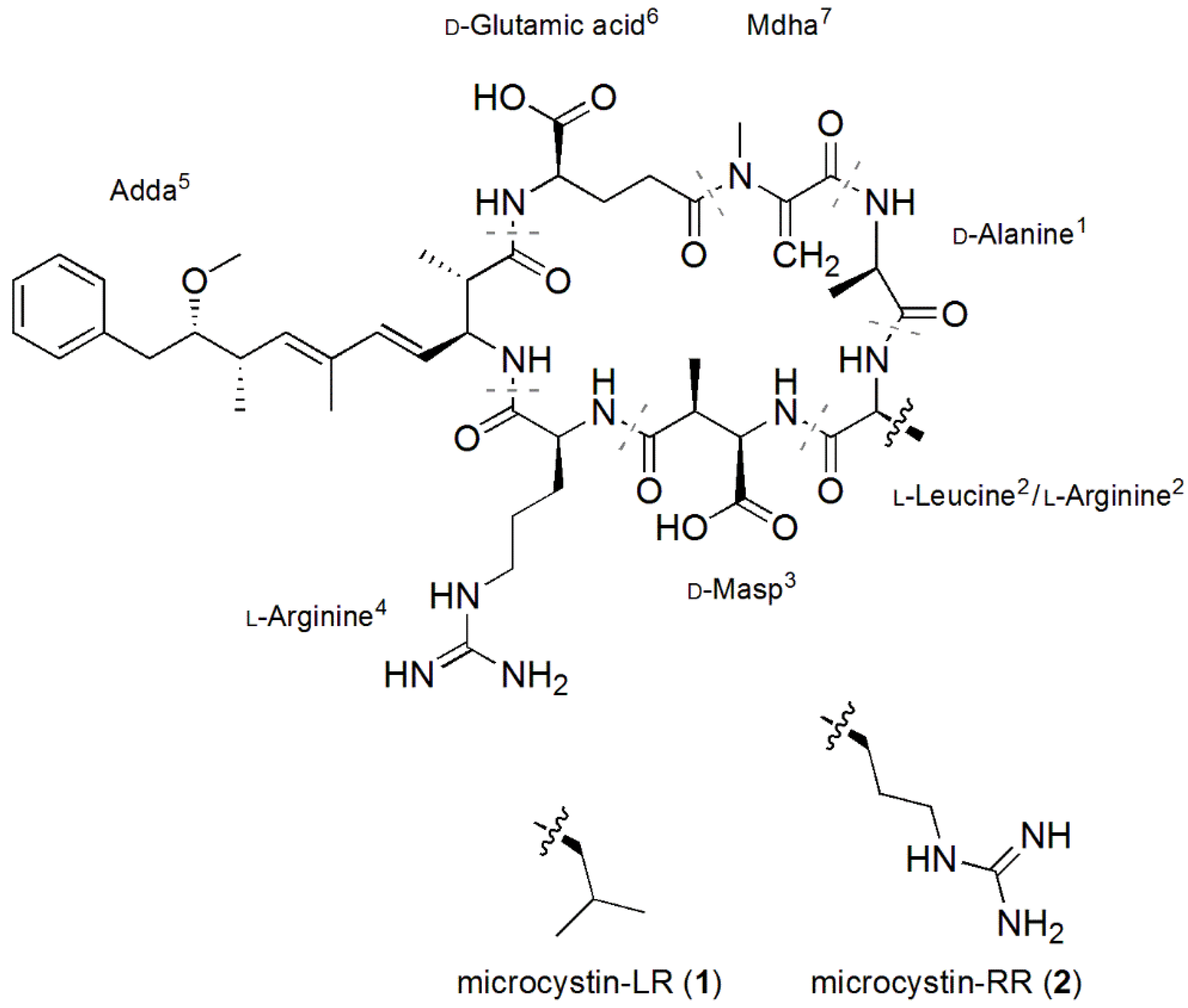
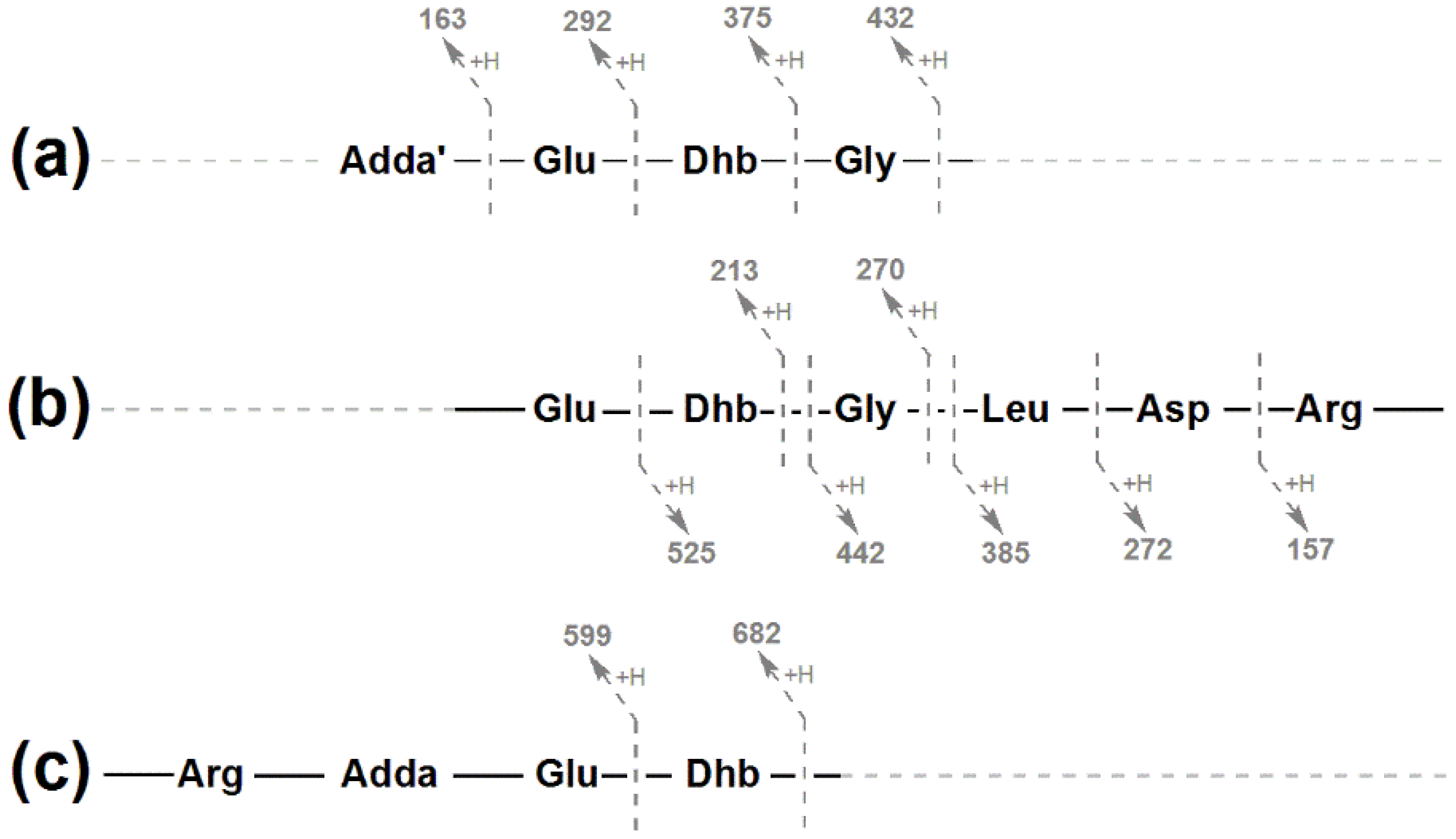
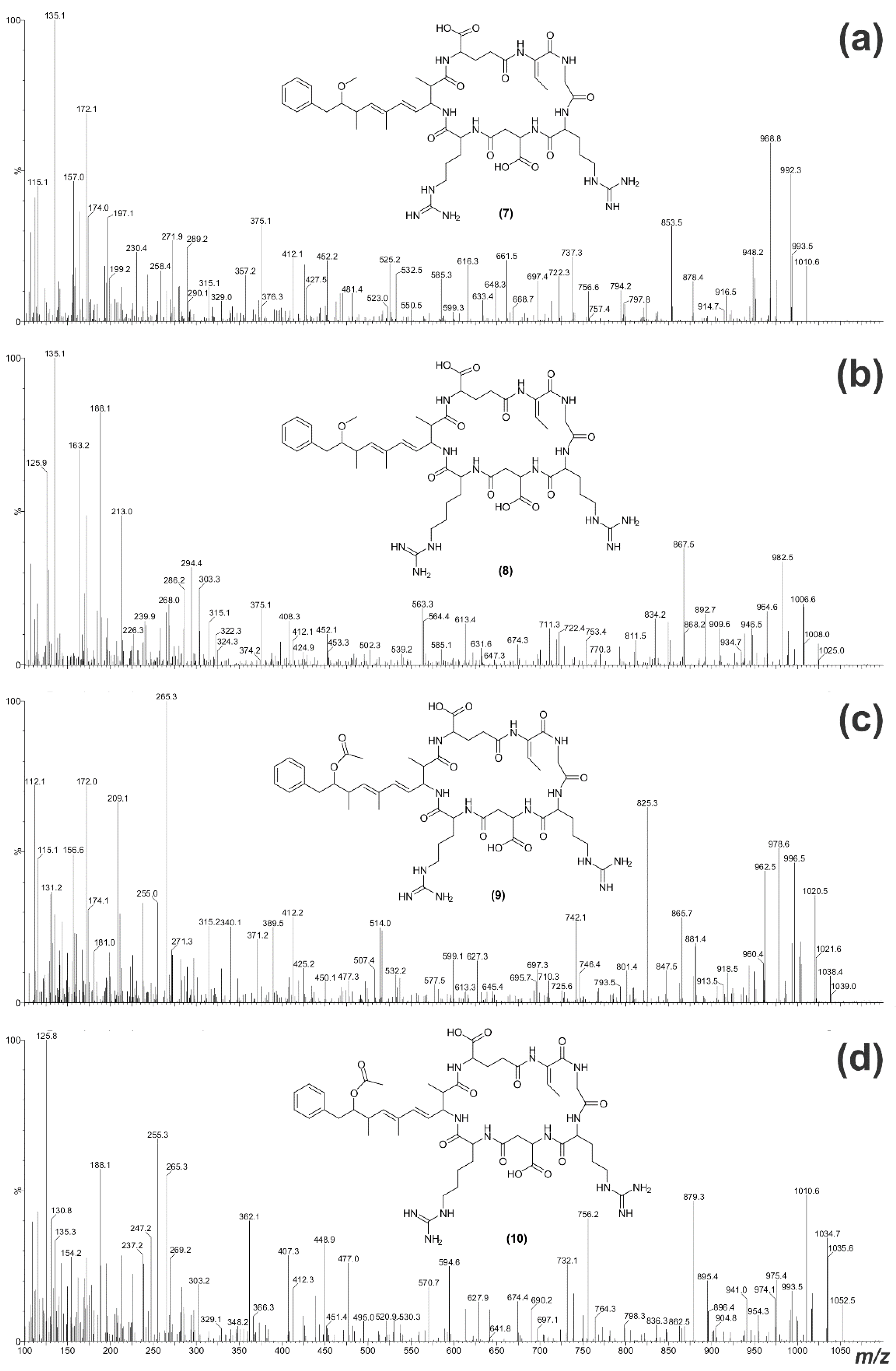
2.2. Structural Characterization of Eight Glycine-Containing Microcystins

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Microcystin | Mr a (Da) | RT b (min) |
|---|---|---|
| MC-LR (1) | 994.5 | 7.40 |
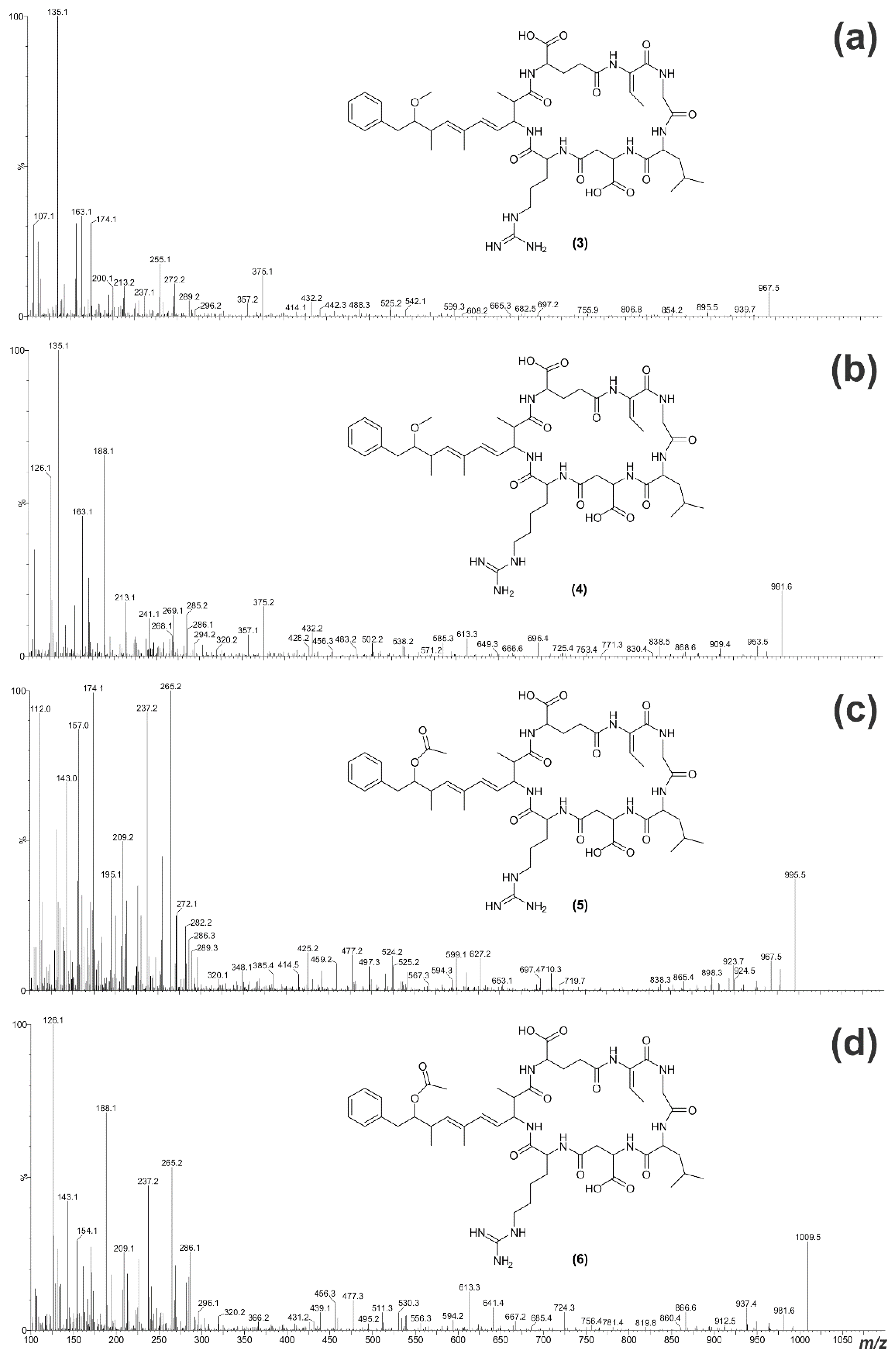
| [Gly1, Asp3, Dhb7] MC-LR (3) | 966.5 | 7.27 |
| [Gly1, Asp3, Dhb7] MC-LHar (4) | 980.5 | 7.34 |
| [Gly1, Asp3, ADMAdda5, Dhb7] MC-LR (5) | 994.5 | 7.29 |
| [Gly1, Asp3, ADMAdda5, Dhb7] MC-LHar (6) | 1008.5 | 7.36 |
| MC-RR (2) | 1023.6 | 6.48 |
| [Gly1, Asp3, Dhb7] MC-RR (7) | 1009.6 | 6.29 |
| [Gly1, Asp3, Dhb7] MC-RHar (8) | 1023.6 | 6.38 |
| [Gly1, Asp3, ADMAdda5, Dhb7] MC-RR (9) | 1037.6 | 6.33 |
| [Gly1, Asp3, ADMAdda5, Dhb7] MC-RHar (10) | 1051.6 | 6.42 |
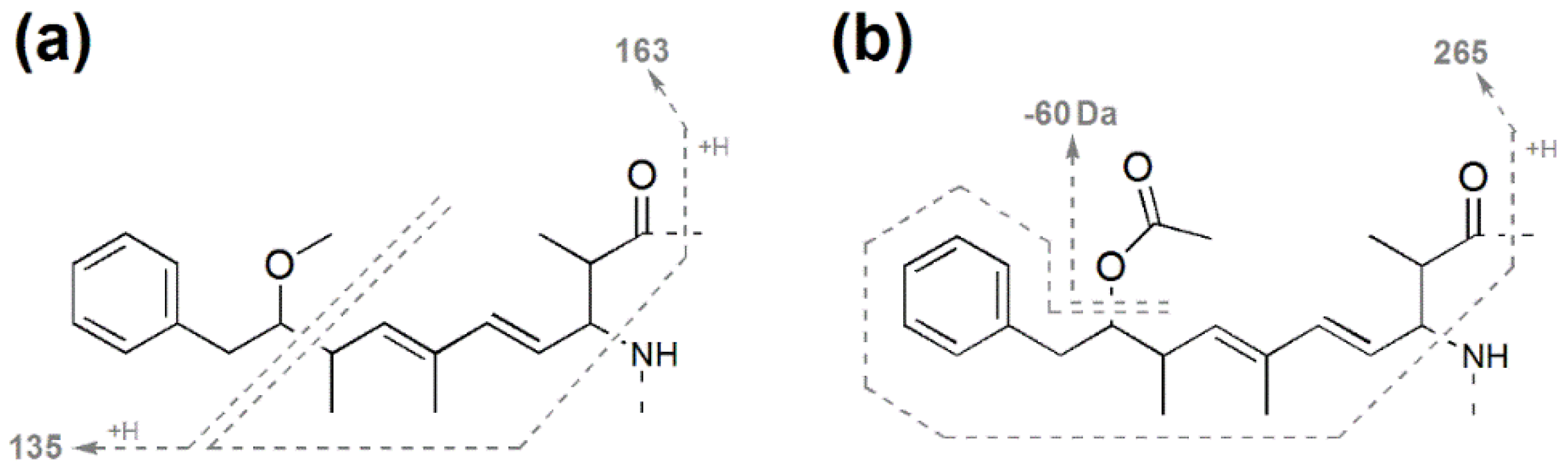
| Fragment Assignment a | MC-LR b (1) | [Gly1, Asp3, Dhb7] MC-LR (3) | [Gly1, Asp3, Dhb7] MC-LHar (4) | [Gly1, Asp3, ADMAdda5, Dhb7] MC-LR (5) | [Gly1, Asp3, ADMAdda5, Dhb7] MC-LHar (6) |
|---|---|---|---|---|---|
| M + H | 995.6 | 967.6 | 981.6 | 995.6 | 1,009.6 |
| M − HOAc + H | - | - | - | 935.5 | 949.3 |
| (Me)Asp-Arg/Har-(ADM)Adda-Glu-Mdha/Dhb-Gly/Ala + H | 882 | 868.6 | 896.9 | ||
| Arg/Har-(ADM)Adda-Glu-Mdha/Dhb-Gly/Ala + H | 753 | 753.3 | 781.4 | ||
| (Me)Asp-Arg/Har-(ADM)Adda-Glu + H | 728 | 728.4 | 756.4 | ||
| Arg/Har-(ADM)Adda-Glu-Mdha/Dhb + H | 682 | 682.5 | 696.1 | 710.3 | 724.3 |
| Glu-Mdha/Dhb-Gly/Ala-Leu-(Me)Asp-Arg/Har + H | 682 | 668.0 | |||
| Arg/Har-(ADM)Adda-Glu + H | 599 | 599.3 | 613.2 | 627.2 | 641.4 |
| (Me)Asp-Arg/Har-(ADM)Adda + H | 599 | 627.3 | |||
| (ADM)Adda’-Glu-Mdha/Dhb-Gly/Ala-Leu + H | 559 | 545.4 | |||
| Mdha/Dhb-Gly/Ala-Leu-(Me)Asp-Arg/Har + H | 553 | 525.2 | 539.4 | 525.2 | 539.3 |
| Glu-Mdha/Dhb-Gly/Ala-Leu-(Me)Asp + H | 526 | 498.2 | |||
| Arg/Har-(ADM)Adda + H | 470 | 512.3 | |||
| Gly/Ala-Leu-(Me)Asp-Arg/Har + H | 470 | 442.3 | 456.2 | 442.1 | 456.1 |
| (ADM)Adda’-Glu-Mdha/Dhb-Gly/Ala + H | 446 | 432.2 | 432.1 | ||
| Leu-(Me)Asp-Arg/Har + H | 399 | 385.1 | 399.1 | 385.4 | |
| Glu-Mdha/Dhb-Gly/Ala-Leu + H | 397 | 383.1 | 383.1 | ||
| (ADM)Adda'-Glu-Mdha/Dhb + H | 375 | 375.1 | 375.2 | 375.2 | |
| Mdha/Dhb-Gly/Ala-Leu-(Me)Asp + H | 397 | 369.2 | |||
| (ADM)Adda'-Glu + H | 292 | 292.1 | 292.2 | 292.1 | 292.1 |
| Gly/Ala-Leu-(Me)Asp + H | 314 | 286.1 | 286.3 | 286.1 | |
| ADMAdda − HOAc + H | - | - | - | 282.2 | 282.2 |
| ADMAdda − HOAc − NH3 + H | - | - | - | 265.1 | 265.2 |
| (Me)Asp-Arg/Har + H | 286 | 272.1 | 286.1 | 272.1 | 286.1 |
| Glu-Mdha/Dhb-Gly/Ala + H | 284 | 270.2 | 270.2 | 270.1 | 270.2 |
| Mdha/Dhb-Gly/Ala-Leu + H | 268 | 254.3 | 254.1 | 254.1 | 254.1 |
| Leu-(Me)Asp + H | 243 | 229.2 | 229.0 | 229.1 | |
| Glu-Mdha/Dhb + H | 213 | 213.1 | 213.1 | 213.1 | 213.1 |
| Gly/Ala-Leu + H | 185 | 171.0 | 171.1 | 171.0 | 171.1 |
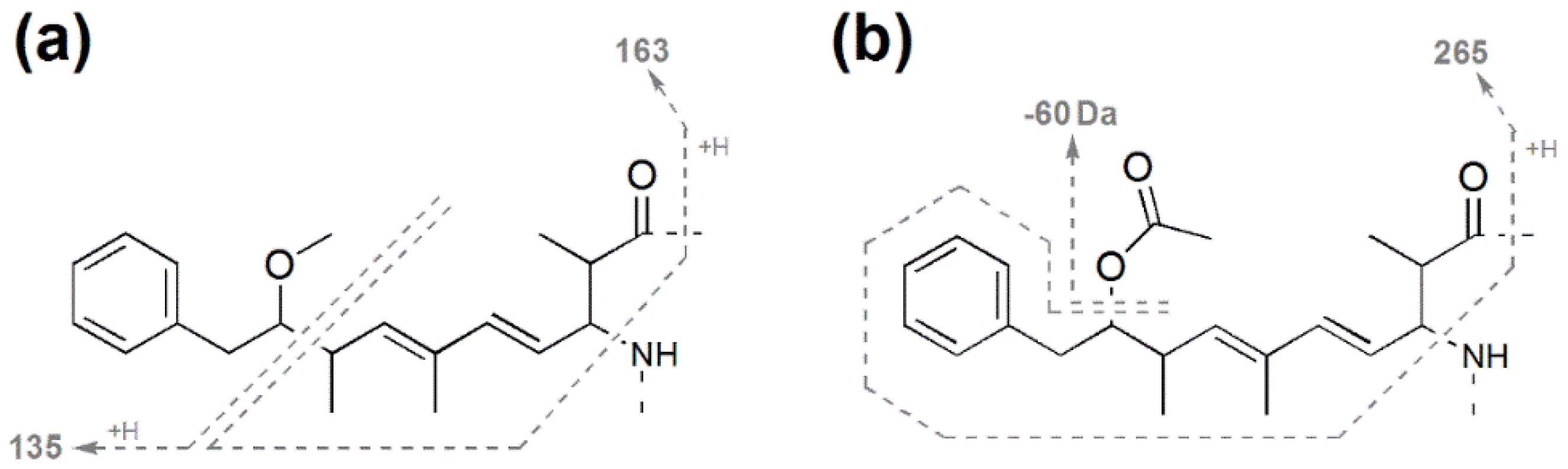
| (ADM)Adda' + H | 163 | 163.1 | 163.1 | 163.1 | 163.0 |
| Arg/Har + H | 157 | 157.0 | 171.1 | 157.1 | 171.1 |
| Mdha/Dhb-Gly/Ala + H | 155 | 141.0 | 141.0 | 141.1 | 141.1 |
| (ADM)Adda sidechain | 135 | 135.1 | 135.1 | 163.1 | 163.0 |
| Arg/Har immonium | 129 | 129.2 | 143.1 | 129.1 | 143.1 |
| Arg/Har fragment | 112 | 112.1 | 112.0/126.1 | 112.0 | 111.9/126.1 |
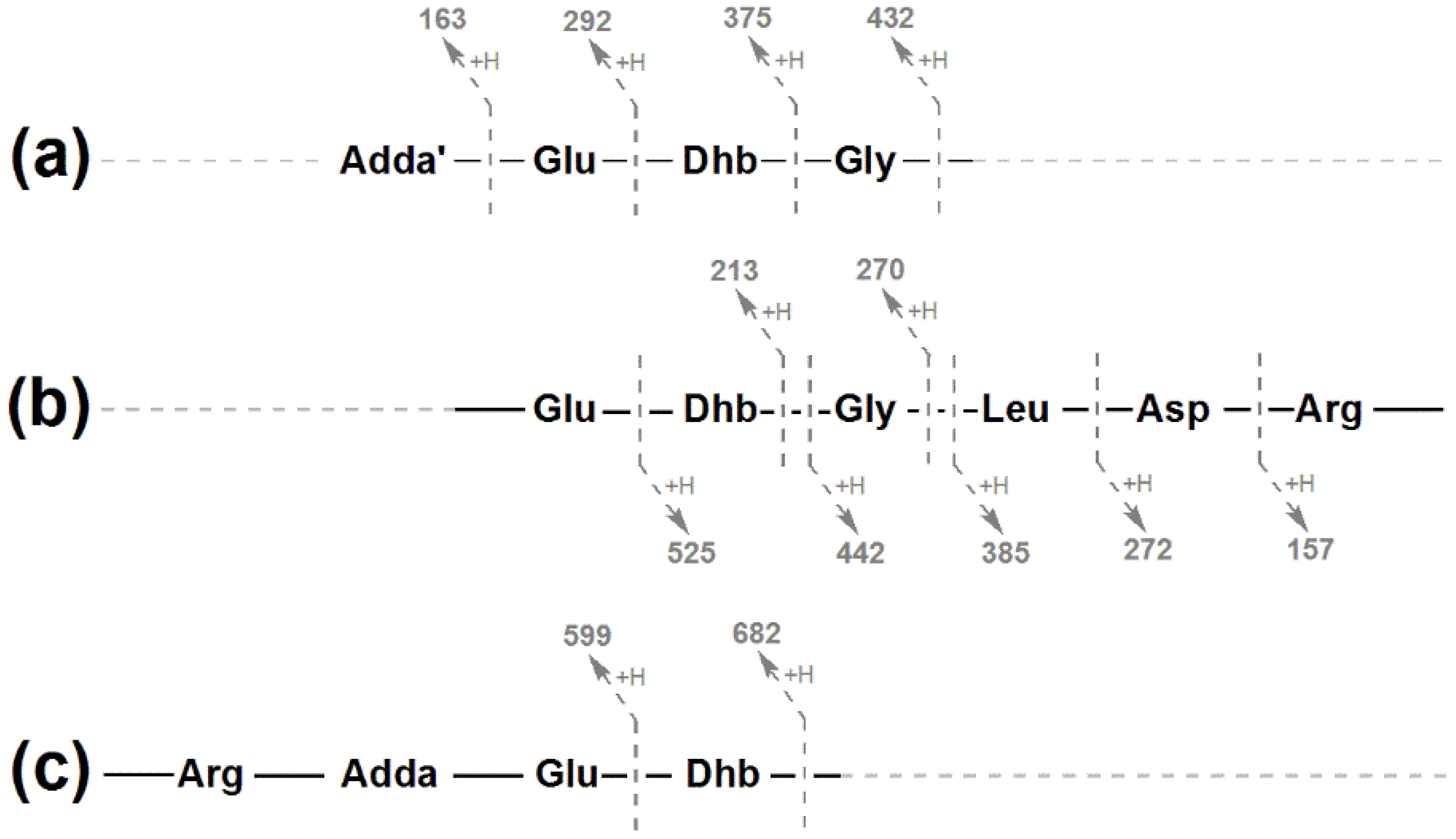
| Fragment Assignment a | MC-RR b (2) | [Gly1, Asp3, Dhb7] MC-RR (7) | [Gly1, Asp3, Dhb7] MC-RHar (8) | [Gly1, Asp3, ADMAdda5, Dhb7] MC-RR (9) | [Gly1, Asp3, ADMAdda5, Dhb7] MC-RHar (10) |
|---|---|---|---|---|---|
| M + H | 1024.7 | 1010.7 | 1024.7 | 1038.7 | 1052.7 |
| M − CN2H2 + H | 982 | 968.9 | 982.5 | 996.5 | 1010.8 |
| M − HOAc + H | - | - | - | 978.6 | 992.5 |
| Arg/Har-(ADM)Adda-Glu-Mdha/Dhb-Gly/Ala-Arg + H | 909 | 895.3 | 909.6 | ||
| (Me)Asp-Arg/Har-(ADM)Adda-Glu-Mdha/Dhb-Gly/Ala + H | 882 | 868.6 | 896.4 | ||
| (Me)Asp-Arg/Har-(ADM)Adda-Glu-Mdha/Dhb + H | 811 | 811.5 | 825.1 | 839.8 | |
| Arg/Har-(ADM)Adda-Glu-Mdha/Dhb-Gly/Ala + H | 753 | 739.2 | 753.4 | 781.0 | |
| (Me)Asp-Arg/Har-(ADM)Adda-Glu + H | 728 | 714.3 | 728.3 | 742.1 | 756.2 |
| Arg/Har-(ADM)Adda-Glu-Mdha/Dhb + H | 682 | 682.2 | 710.2 | 724.2 | |
| Glu-Mdha/Dhb-Gly/Ala-Arg-(Me)Asp-Arg + H | 725 | 697.5 | 697.3 | ||
| Arg/Har-(ADM)Adda-Glu + H | 599 | 613.5 | 641.2 | ||
| (Me)Asp-Arg/Har-(ADM)Adda + H | 599 | 585.2 | 599.2 | 613.4 | 627.9 |
| Mdha/Dhb-Gly/Ala-Arg-(Me)Asp-Arg/Har + H | 596 | 582.4 | 568.7 | ||
| Har-ADMAdda − HOAC + H | - | - | - | 452.5 | |
| (ADM)Adda'-Glu-Mdha/Dhb-Gly/Ala + H | 446 | 432.2 | |||
| Glu-Mdha/Dhb-Gly/Ala-Arg + H | 440 | 426.4 | 426.2 | ||
| Mdha/Dhb-Gly/Ala-Arg-(Me)Asp + H | 440 | 412.1 | 412.3 | 412.2 | 412.3 |
| (ADM)Adda'-Glu-Mdha/Dhb + H | 375 | 375.1 | 375.2 | ||
| Gly/Ala-Arg-(Me)Asp + H | 357 | 329.1 | 329.1 | 329.1 | |
| Mdha/Dhb-Gly/Ala-Arg + H | 311 | 297.1 | 296.9 | 297.0 | 297.0 |
| (ADM)Adda'-Glu + H | 292 | 292.1 | |||
| ADMAdda − HOAc + H | - | - | - | 282.3 | 282.1 |
| ADMAdda − HOAc − NH3 + H | - | - | - | 265.4 | 265.3 |
| (Me)Asp-Arg/Har + H | 286 | 272.3 | 286.2 | 286.1 | |
| Arg-(Me)Asp + H | 286 | 272.3 | 272.1 | 272.2 | 272.5 |
| Glu-Mdha/Dhb-Gly/Ala + H | 284 | 270.0 | 270.1 | ||
| Gly/Ala-Arg + H | 228 | 214.1 | 214.0 | 214.1 | 214.2 |
| Glu-Mdha/Dhb + H | 213 | 213.1 | 212.9 | 213.1 | 213.2 |
| (ADM)Adda' + H | 163 | 163.0 | 162.9 | 163.0 | |
| Har + H | - | - | - | 171.0 | |
| Arg + H | 157 | 157.0 | 157.1 | 156.6 | 156.9 |
| Mdha/Dhb-Gly/Ala + H | 155 | 141.5 | 140.9 | 141.2 | 140.7 |
| (ADM)Adda sidechain | 135 | 135.1 | 135.1 | 163.0 | |
| Har immonium | - | - | 143.1 | - | 143.1 |
| Arg immonium | 129 | 129.5 | 129.1 | 129.1 | |
| Har fragment | - | - | 112.0/126.0 | - | 111.8/125.8 |
| Arg fragment | 112 | 112.2 | 112.0 | 112.1 | 111.8 |
3. Experimental Section
3.1. General Experimental Procedures
3.2. Sample Collection
3.3. Matrix-Assisted Laser Desorption/Ionization-Time of Flight Mass Spectrometry Analysis
3.4. Liquid Chromatography-Mass Spectrometry Analyses
3.5. Isolation of Semi-Pure Mixtures of the Antarctic Microcystins
3.6. Advanced Marfey’s Amino Acid Analysis
3.7. β-Mercaptoethanol Derivatization for Mdha/Dhb Determination
4. Conclusions
Supplementary Materials
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Doran, P.T.; McKay, C.P.; Clow, G.D.; Dana, G.L.; Fountain, A.G.; Nylen, T.; Lyons, W.B. Valley floor climate observations from the McMurdo dry valleys, Antarctica, 1986–2000. J. Geophys. Res. 2002, 107, 4772. [Google Scholar] [CrossRef]
- Cowan, D.; Russell, N.; Mamais, A.; Sheppard, D. Antarctic Dry Valley mineral soils contain unexpectedly high levels of microbial biomass. Extremophiles 2002, 6, 431–436. [Google Scholar] [CrossRef] [PubMed]
- Wynn-Williams, D.D. Ecological aspects of Antarctic microbiology. Adv. Microb. Ecol. 1990, 11, 71–146. [Google Scholar]
- Vishniac, H.S. The microbiology of Antarctic soils. In Antarctic microbiology; Friedmann, I.E., Ed.; Wiley-Liss: New York, NY, USA, 1993; pp. 297–341. [Google Scholar]
- Taton, A.; Grubisic, S.; Balthasart, P.; Hodgson, D.A.; Laybourn-Parry, J.; Wilmotte, A. Biogeographical distribution and ecological ranges of benthic cyanobacteria in east Antarctic lakes. FEMS Microbiol. Ecol. 2006, 57, 272–289. [Google Scholar] [CrossRef] [PubMed]
- Cavacini, P. Soil algae from northern Victoria Land (Antarctica). Polar Biosci. 2001, 14, 45–60. [Google Scholar]
- Fumanti, B.; Cavacini, P.; Alfinito, S. Benthic algal mats of some lakes of Inexpressible Island (northern Victoria Land, Antarctica). Polar Biol. 1996, 17, 25–30. [Google Scholar] [CrossRef]
- Sivonen, K.; Jones, G. Cyanobacterial toxins. In Toxic Cyanobacteria in Water: A Guide to Their Public Health Consequences, Monitoring and Management; Chorus, I., Bartram, J., Eds.; E & FN Spon: London, UK, 1999; pp. 55–124. [Google Scholar]
- Hitzfeld, B.C.; Lampert, C.S.; Spaeth, N.; Mountfort, D.; Kaspar, H.; Dietrich, D.R. Toxin production in cyanobacterial mats from ponds on the McMurdo Ice Shelf, Antarctica. Toxicon 2000, 38, 1731–1748. [Google Scholar] [CrossRef] [PubMed]
- Jungblut, A.-D.; Hawes, I.; Mountfort, D.; Hitzfeld, B.; Dietrich, D.R.; Burns, B.P.; Neilan, B.A. Diversity within cyanobacterial mat communities in variable salinity meltwater ponds of McMurdo Ice Shelf, Antarctica. Environ. Microbiol. 2005, 7, 519–529. [Google Scholar] [CrossRef] [PubMed]
- Jungblut, A.-D.; Hoeger, S.J.; Mountfort, D.; Hitzfeld, B.C.; Dietrich, D.R.; Neilan, B.A. Characterization of microcystin production in an Antarctic cyanobacterial mat community. Toxicon 2006, 47, 271–278. [Google Scholar] [CrossRef] [PubMed]
- Kleinteich, J.; Wood, S.A.; Puddick, J.; Schleheck, D.; Kupper, F.C.; Dietrich, D. Potent toxins in Arctic environments—Presence of saxitoxins and an unusual microcystin variant in Arctic freshwater ecosystems. Chem. Biol. Interact. 2013, 206, 423–431. [Google Scholar] [CrossRef] [PubMed]
- Kleinteich, J.; Wood, S.A.; Kupper, F.C.; Camacho, A.; Quesada, A.; Frickey, T.; Dietrich, D.R. Temperature-related changes in polar cyanobacterial mat diversity and toxin production. Nat. Clim. Chang. 2012, 2, 356–360. [Google Scholar] [CrossRef]
- Niedermeyer, T. Microcystin congeners described in the literature. Available online: http://dx.doi.org/10.6084/m9.figshare.880756 (accessed on 21 October 2014).
- Rinehart, K.; Namikoshi, M.; Choi, B. Structure and biosynthesis of toxins from blue-green algae (cyanobacteria). J. Appl. Phycol. 1994, 6, 159–176. [Google Scholar] [CrossRef]
- Sivonen, K.; Namikoshi, M.; Evans, W.R.; Fardig, M.; Carmichael, W.W.; Rinehart, K.L. Three new microcystins, cyclic heptapeptide hepatotoxins, from Nostoc sp. strain 152. Chem. Res. Toxicol. 1992, 5, 464–469. [Google Scholar] [CrossRef] [PubMed]
- Park, H.; Namikoshi, M.; Brittain, S.M.; Carmichael, W.W.; Murphy, T. [D-Leu1] microcystin-LR, a new microcystin isolated from waterbloom in a Canadian prairie lake. Toxicon 2001, 39, 855–862. [Google Scholar] [CrossRef] [PubMed]
- Matthiensen, A.; Beattie, K.A.; Yunes, J.S.; Kaya, K.; Codd, G.A. [D-Leu1]Microcystin-LR, from the cyanobacterium Microcystis RST 9501 and from a Microcystis bloom in the Patos Lagoon estuary, Brazil. Phytochemistry 2000, 55, 383–387. [Google Scholar] [CrossRef] [PubMed]
- Shishido, T.K.; Kaasalainen, U.; Fewer, D.P.; Rouhiainen, L.; Jokela, J.; Wahlsten, M.; Fiore, M.F.; Yunes, J.S.; Rikkinen, J.; Sivonen, K. Convergent evolution of [D-Leucine1] microcystin-LR in taxonomically disparate cyanobacteria. BMC Evol. Biol. 2013, 13, 86. [Google Scholar] [CrossRef] [PubMed]
- Qi, Y.; Rosso, L.; Sedan, D.; Giannuzzi, L.; Andrinolo, D.; Volmer, D.A. Seven new microcystin variants discovered from a native Microcystis aeruginosa strain–unambiguous assignment of product ions by tandem mass spectrometry. Rapid Commun. Mass Spectrom. 2015, 29, 1–5. [Google Scholar] [CrossRef] [PubMed]
- Wood, S.A.; Mountfort, D.; Selwood, A.I.; Holland, P.T.; Puddick, J.; Cary, S.C. Widespread distribution and identification of eight novel microcystins in Antarctic cyanobacterial mats. Appl. Environ. Microbiol. 2008, 74, 7243–7251. [Google Scholar] [CrossRef] [PubMed]
- Puddick, J. Spectroscopic investigations of oligopeptides from aquatic cyanobacteria: Characterisation of new oligopeptides, development of microcystin quantification tools and investigations into microcystin production. Ph.D. Thesis, University of Waikato, Hamilton, New Zealand, February 2013. [Google Scholar]
- Puddick, J.; Prinsep, M.R.; Wood, S.A.; Miles, C.O.; Rise, F.; Cary, S.C.; Hamilton, D.P.; Wilkins, A.L. Structural characterization of new microcystins containing tryptophan and oxidized tryptophan residues. Mar. Drugs 2013, 11, 3025–3045. [Google Scholar] [CrossRef] [PubMed]
- Fujii, K.; Ikai, Y.; Mayumi, T.; Oka, H.; Suzuki, M.; Harada, K.-I. A nonempirical method using LC/MS for determination of the absolute configuration of constituent amino acids in a peptide: Elucidation of limitations of Marfey's method and of its separation mechanism. Anal. Chem. 1997, 69, 3346–3352. [Google Scholar] [CrossRef]
- Fujii, K.; Ikai, Y.; Oka, H.; Suzuki, M.; Harada, K.-I. A nonempirical method using LC/MS for determination of the absolute configuration of constituent amino acids in a peptide: Combination of Marfey’s method with mass spectrometry and its practical application. Anal. Chem. 1997, 69, 5146–5151. [Google Scholar] [CrossRef]
- Miles, C.O.; Sandvik, M.; Nonga, H.E.; Rundberget, T.; Wilkins, A.L.; Rise, F.; Ballot, A. Thiol derivatization for LC-MS identification of microcystins in complex matrices. Environ. Toxicol. 2012, 46, 8937–8944. [Google Scholar]
- Smith, J.L.; Boyer, G.L. Standardization of microcystin extraction from fish tissues: A novel internal standard as a surrogate for polar and non-polar variants. Toxicon 2009, 53, 238–245. [Google Scholar] [CrossRef] [PubMed]
- Miles, C.O.; Sandvik, M.; Haande, S.; Nonga, H.; Ballot, A. First use of LC-MS analysis with thiol derivatization to differentiate [Dhb7]- from [Mdha7]-microcystins: Analysis of cyanobacterial blooms, Planktothrix cultures and European crayfish from Lake Steinsfjorden, Norway. Environ. Sci. Technol. 2013, 47, 4080–4087. [Google Scholar] [CrossRef] [PubMed]
- Kruger, T.; Christian, B.; Luckas, B. Development of an analytical method for the unambiguous structure elucidation of cyclic peptides with special appliance for hepatotoxic desmethylated microcystins. Toxicon 2009, 54, 302–312. [Google Scholar] [CrossRef] [PubMed]
- Diehnelt, C.W.; Dugan, N.R.; Peterman, S.M.; Budde, W.L. Identification of microcystin toxins from a strain of Microcystis aeruginosa by liquid chromatography introduction into a hybrid linear ion trap-fourier transform ion cyclotron resonance mass spectrometer. Anal. Chem. 2006, 78, 501–512. [Google Scholar] [CrossRef] [PubMed]
- Hummert, C.; Dahlmann, J.; Reinhardt, K.; Dang, H.; Dang, D.; Luckas, B. Liquid chromatography-mass spectrometry identification of microcystins in Microcystis aeruginosa strain from lake Thanh Cong, Hanoi, Vietnam. Chromatographia 2001, 54, 569–575. [Google Scholar] [CrossRef]
- Yuan, M.; Namikoshi, M.; Otsuki, A.; Sivonen, K. Effect of amino acid side-chain on fragmentation of cyclic peptide ions: Differences of ESI-MS/CID mass spectra of toxic heptapeptide microcystins containing ADMAdda instead of Adda. Eur. J. Mass Spectrom. 1998, 4, 287–298. [Google Scholar] [CrossRef]
- Erhard, M.; von Döhren, H.; Jungblut, P.R. Rapid identification of the new anabaenopeptin G from Planktothrix agardhii HUB 011 using matrix-assisted laser desorption/ionization time-of-flight mass spectrometry. Rapid Commun. Mass Spectrom. 1999, 13, 337–343. [Google Scholar] [CrossRef] [PubMed]
- Mayumi, T.; Kato, H.; Imanishi, S.; Kawasaki, Y.; Hasegawa, M.; Harada, K.-I. Structural characterization of microcystins by LC/MS/MS under Ion trap conditions. J. Antibiot. 2006, 59, 710–719. [Google Scholar] [CrossRef] [PubMed]
- Frias, H.V.; Mendes, M.A.; Cardozo, K.H.M.; Carvalho, V.M.; Tomazela, D.; Colepicolo, P.; Pinto, E. Use of electrospray tandem mass spectrometry for identification of microcystins during a cyanobacterial bloom event. Biochem. Biophys. Res. Commun. 2006, 344, 741–746. [Google Scholar] [CrossRef] [PubMed]
- Bateman, K.P.; Thibault, P.; Douglas, D.J.; White, R.L. Mass spectral analyses of microcystins from toxic cyanobacteria using on-line chromatographic and electrophoretic separations. J. Chromatogr. A 1995, 712, 253–268. [Google Scholar] [CrossRef] [PubMed]
- Ferranti, P.; Fabbrocino, S.; Nasi, A.; Caira, S.; Bruno, M.; Serpe, L.; Gallo, P. Liquid chromatography coupled to quadruple time-of-flight tandem mass spectrometry for microcystin analysis in freshwaters: Method performances and characterisation of a novel variant of microcystin-RR. Rapid Commun. Mass Spectrom. 2009, 23, 1328–1336. [Google Scholar] [CrossRef] [PubMed]
- Oksanen, I.; Jokela, J.; Fewer, D.P.; Wahlsten, M.; Rikkinen, J.; Sivonen, K. Discovery of rare and highly toxic microcystins from lichen-associated cyanobacterium Nostoc sp. strain IO-102-I. Appl. Environ. Microbiol. 2004, 70, 5756–5763. [Google Scholar] [CrossRef] [PubMed]
- Laub, J.; Henriksen, P.; Brittain, S.M.; Wang, J.; Carmichael, W.W.; Rinehart, K.L.; Moestrup, Ø. [ADMAdda5]-microcystins in Planktothrix agardhii strain PH-123 (cyanobacteria)—Importance for monitoring of microcystins in the environment. Environ. Toxicol. 2002, 17, 351–357. [Google Scholar] [CrossRef] [PubMed]
- Yuan, M.; Namikoshi, M.; Otsuki, A.; Rinehart, K.L.; Sivonen, K.; Watanabe, M.F. Low-energy collisionally activated decomposition and structural characterization of cyclic heptapeptide microcystins by electrospray ionization mass spectrometry. J. Mass Spectrom. 1999, 34, 33–43. [Google Scholar] [CrossRef] [PubMed]
- Sivonen, K.; Skulberg, O.M.; Namikoshi, M.; Evans, W.R.; Carmichael, W.W.; Rinehart, K.L. Two methyl ester derivatives of microcystins, cyclic heptapeptide hepatotoxins, isolated from Anabaena flos-aquae strain CYA 83/1. Toxicon 1992, 30, 1465–1471. [Google Scholar] [CrossRef] [PubMed]
- Namikoshi, M.; Rinehart, K.L.; Sakai, R.; Sivonen, K.; Carmichael, W.W. Structures of three new cyclic heptapeptide hepatotoxins produced by the cyanobacterium (blue-green alga) Nostoc sp. strain 152. J. Org. Chem. 1990, 55, 6135–6139. [Google Scholar] [CrossRef]
- Sano, T.; Kaya, K. Two new (E)-2-amino-2-butenoic acid (Dhb)-containing microcystins isolated from Oscillatoria agardhii. Tetrahedron 1998, 54, 463–470. [Google Scholar] [CrossRef]
- Sano, T.; Takagi, H.; Kaya, K. A Dhb-microcystin from the filamentous cyanobacterium Planktothrix rubescens. Phytochemistry 2004, 65, 2159–2162. [Google Scholar] [CrossRef] [PubMed]
- Sano, T.; Beattie, K.A.; Codd, G.A.; Kaya, K. Two (Z)-dehydrobutyrine-containing microcystins from a hepatotoxic bloom of Oscillatoria agardhii from Soulseat Loch, Scotland. J. Nat. Prod. 1998, 61, 851–853. [Google Scholar] [CrossRef] [PubMed]
- Beattie, K.A.; Kaya, K.; Sano, T.; Codd, G.A. Three dehydrobutyrine-containing microcystins from Nostoc. Phytochemistry 1998, 47, 1289–1292. [Google Scholar] [CrossRef]
- Christiansen, G.; Yoshida, W.Y.; Blom, J.F.; Portmann, C.; Gademann, K.; Hemscheidt, T.; Kurmayer, R. Isolation and structure determination of two microcystins and sequence comparison of the McyABC adenylation domains in Planktothrix species. J. Nat. Prod. 2008, 71, 1881–1886. [Google Scholar] [CrossRef] [PubMed]
- Niedermeyer, T.H.J.; Daily, A.; Swiatecka-Hagenbruch, M.; Moscow, J.A. Selectivity and potency of microcystin congeners against OATP1B1 and OATP1B3 expressing cancer cells. PLoS One 2014, 9, e91476. [Google Scholar] [CrossRef] [PubMed]
- Niedermeyer, T.H.J.; Schmieder, P.; Kurmayer, R. Isolation of microcystins from the cyanobacterium Planktothrix rubescens strain No80. Nat. Prod. Bioprospect. 2014, 4, 37–45. [Google Scholar] [CrossRef] [PubMed]
- Tillett, D.; Dittmann, E.; Erhard, M.; von Döhren, H.; Börner, T.; Neilan, B.A. Structural organization of microcystin biosynthesis in Microcystis aeruginosa PCC7806: An integrated peptide-polyketide synthetase system. Chem. Biol. 2000, 7, 753–764. [Google Scholar] [CrossRef] [PubMed]
- Challis, G.L.; Ravel, J.; Townsend, C.A. Predictive, structure-based model of amino acid recognition by nonribosomal peptide synthetase adenylation domains. Chem. Biol. 2000, 7, 211–224. [Google Scholar] [CrossRef] [PubMed]
- Sivonen, K.; Carmichael, W.W.; Namikoshi, M.; Rinehart, K.L.; Dahlem, A.M.; Niemela, S.I. Isolation and characterization of hepatotoxic microcystin homologs from the filamentous freshwater cyanobacterium Nostoc sp. strain 152. Appl. Environ. Microbiol. 1990, 56, 2650–2657. [Google Scholar] [PubMed]
- Prakash, S.; Lawton, L.A.; Edwards, C. Stability of toxigenic Microcystis blooms. Harmful Algae 2009, 8, 377–384. [Google Scholar] [CrossRef]
- Kaasalainen, U.; Jokela, J.; Fewer, D.P.; Sivonen, K.; Rikkinen, J. Microcystin production in the tripartite cyanolichen Peltigera leucophlebia. Mol. Plant-Microbe Interact. 2009, 22, 695–702. [Google Scholar] [CrossRef] [PubMed]
- Puddick, J.; Prinsep, M.R.; Wood, S.A.; Kaufononga, S.A.F.; Cary, S.C.; Hamilton, D.P. High levels of structural diversity observed in microcystins from Microcystis CAWBG11 and characterization of six new microcystin congeners. Mar. Drugs 2014, 12, 5372–5395. [Google Scholar] [CrossRef] [PubMed]
- Marfey, P. Determination of D-amino acids. II. Use of a bifunctional reagent, 1,5-difluoro-2,4-dinitrobenzene. Carlsberg Res. Commun. 1984, 49, 591–596. [Google Scholar] [CrossRef]
- Puddick, J.; Prinsep, M.R.; Wood, S.A.; Cary, S.C.; Hamilton, D.P.; Wilkins, A.L. Isolation and structure determination of two new hydrophobic microcystins from Microcystis sp. (CAWBG11). Phytochem. Lett. 2013, 6, 575–581. [Google Scholar] [CrossRef]
© 2015 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Puddick, J.; Prinsep, M.R.; Wood, S.A.; Cary, S.C.; Hamilton, D.P.; Holland, P.T. Further Characterization of Glycine-Containing Microcystins from the McMurdo Dry Valleys of Antarctica. Toxins 2015, 7, 493-515. https://doi.org/10.3390/toxins7020493
Puddick J, Prinsep MR, Wood SA, Cary SC, Hamilton DP, Holland PT. Further Characterization of Glycine-Containing Microcystins from the McMurdo Dry Valleys of Antarctica. Toxins. 2015; 7(2):493-515. https://doi.org/10.3390/toxins7020493
Chicago/Turabian StylePuddick, Jonathan, Michèle R. Prinsep, Susanna A. Wood, Stephen Craig Cary, David P. Hamilton, and Patrick T. Holland. 2015. "Further Characterization of Glycine-Containing Microcystins from the McMurdo Dry Valleys of Antarctica" Toxins 7, no. 2: 493-515. https://doi.org/10.3390/toxins7020493
APA StylePuddick, J., Prinsep, M. R., Wood, S. A., Cary, S. C., Hamilton, D. P., & Holland, P. T. (2015). Further Characterization of Glycine-Containing Microcystins from the McMurdo Dry Valleys of Antarctica. Toxins, 7(2), 493-515. https://doi.org/10.3390/toxins7020493