
Detection of Cyanotoxins, β-N-methylamino-L-alanine and Microcystins, from a Lake Surrounded by Cases of Amyotrophic Lateral Sclerosis

Abstract
:1. Introduction
2. Results and Discussion
2.1. Results
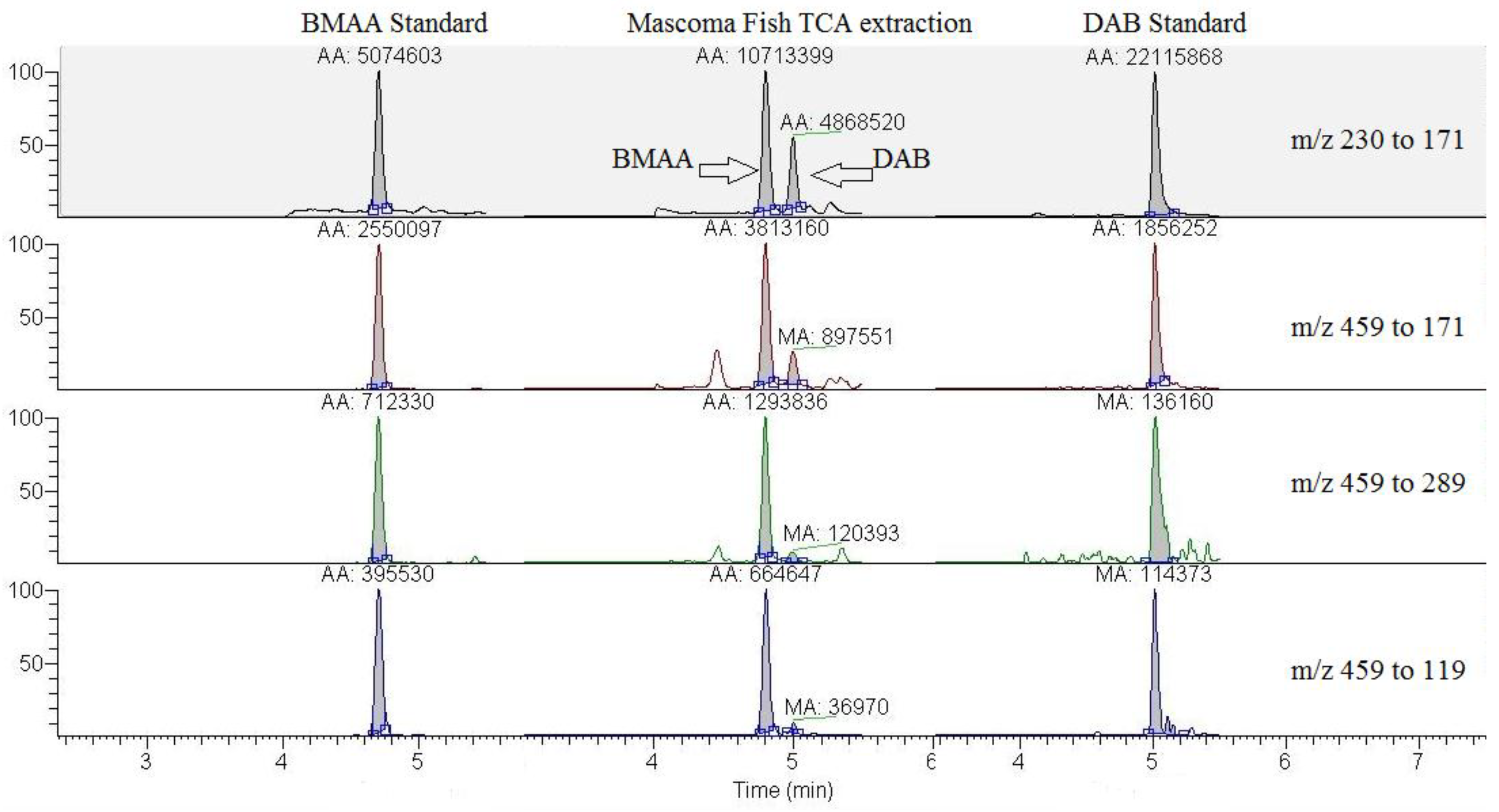

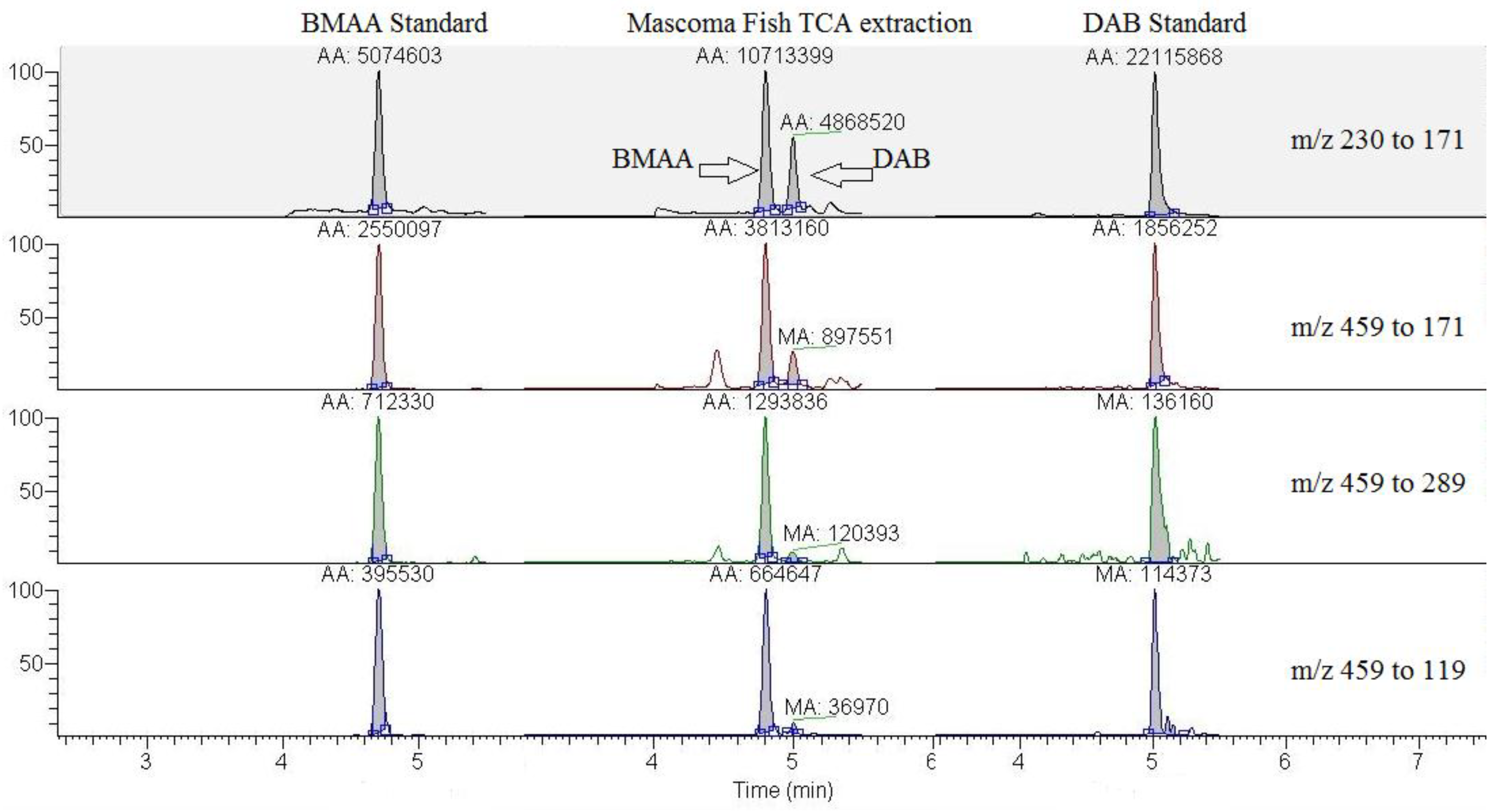{kind=link}
{kind=link}
| Tissue | Number of replicates examined | BMAA total ± SD (μg BMAA/g sample) | DAB total ± SD (μg DAB/g sample) | Ratio of m/z 289/171 from parent m/z 459 at retention time for BMAA | Ratio of m/z 119/171 from parent m/z 459 at retention time for BMAA |
|---|---|---|---|---|---|
| Carp brain | 3 | 0.43 ± 0.02 # | 0.01 ± 0.002 # | 13.2 ± 1.2 * | 28.9 ± 0.6 * |
| Carp liver | 3 | 1.28 ± 0.03 | ND | 12.5 ± 0.8 * | 29.3 ± 1.7 * |
| Carp muscle | 1 | 1.27 | ND | 13.2 | 29.4 |
| BMAA standard | 4 | - | - | 12.4 ± 0.9 | 31.0 ± 1.6 |
| Toxin Analysis | Lake | Date |
|---|---|---|
| ND | 2972 Goose Pond (cyano B + microcystin positive) | 4 July 2011 |
| DAB | 2973-1 Goose Pond (Background—No Blooms) | 24 May 2010 |
| DAB at 5 ng/cm2 filter | 2973-2 Goose Pond | 25 May 2010 |
| AEG + DAB | Mascoma Lake 4 | 19–21 September 2009 |
| AEG + DAB + BMAA | Mascoma Lake 5 | 13–15 September 2009 |

2.2. Discussion
3. Experimental Section
3.1. Fish Tissue Analysis
3.2. Microcystin Preparation and Analysis
3.3. Aerosol Collection
4. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Factor-Litvak, P.; Al-Chalabi, A.; Ascherio, A.; Bradley, W.; Chio, A.; Garruto, R.; Hardiman, O.; Kamel, F.; Kasarskis, E.; McKee, A.; et al. Current pathways for epidemiological research in Amyotrophic Lateral Sclerosis. Amyotroph. Lateral Scler. Front. Degener. 2013, 14, 33–43. [Google Scholar] [CrossRef]
- Ishimura, R.; Nagy, G.; Dotu, I.; Zhou, H.; Yang, X.L.; Schimmel, P.; Senju, S.; Nishimura, Y.; Chuang, J.H.; Ackerman, S.L. RNA function. Ribosome stalling induced by mutation of a CNS-specific tRNA causes neurodegeneration. Science 2014, 345, 455–459. [Google Scholar] [CrossRef] [PubMed]
- Pablo, J.; Banack, S.A.; Cox, P.A.; Johnson, T.E.; Papapetropoulos, S.; Bradley, W.G.; Buck, A.; Mash, D.C. Cyanobacterial neurotoxin BMAA in ALS and Alzheimer’s Disease. Acta Neurol. Scand. 2009, 120, 216–225. [Google Scholar] [CrossRef] [PubMed]
- Banack, S.A.; Caller, T.A.; Stommel, E.W. The cyanobacteria derived toxin β-N-methylamino-l-alanine and Amyotrophic Lateral Sclerosis. Toxins 2010, 2, 2837–2850. [Google Scholar] [CrossRef] [PubMed]
- Murch, S.J.; Cox, P.A.; Banack, S.A. A mechanism for slow release of biomagnified cyanobacterial neurotoxins and neurodegenerative disease in Guam. Proc. Natl. Acad. Sci. USA 2004, 101, 12228–12231. [Google Scholar] [CrossRef] [PubMed]
- Merel, S.; Walker, D.; Chicana, R.; Snyder, S.; Baures, E.; Thomas, O. State of knowledge and concerns on cyanobacterial blooms and cyanotoxins. Environ. Int. 2013, 59, 303–327. [Google Scholar] [CrossRef] [PubMed]
- Cox, P.A.; Banack, S.A.; Murch, S.J.; Rasmussen, U.; Tien, G.; Bidigare, R.R.; Metcalf, J.S.; Morrison, L.F.; Codd, G.A.; Bergman, B. Diverse taxa of cyanobacteria produce β-N-methylamino-l-alanine, a neurotoxic amino acid. Proc. Natl. Acad. Sci. USA 2005, 102, 5074–5078. [Google Scholar] [CrossRef] [PubMed]
- Funari, E.; Testai, E. Human health risk assessment related to cyanotoxins exposure. Crit. Rev. Toxicol. 2008, 38, 97–125. [Google Scholar] [CrossRef] [PubMed]
- Metcalf, J.S.; Codd, G.A. Cyanotoxins. In Ecology of cyanobacteria II: Their Diversity in Space and Time, 2nd ed.; Whitton, B.A., Ed.; Springer: Dordrecht, The Netherlands, 2012; pp. 651–675. [Google Scholar]
- Reed, D.; Plato, C.; Elizan, T.; Kurland, L.T. The amyotrophic lateral sclerosis/parkinsonism-dementia complex: A ten-year follow-up on Guam. I. Epidemiologic studies. Am. J. Epidemiol. 1966, 83, 54–73. [Google Scholar] [PubMed]
- Arnold, A.; Edgren, D.C.; Palladino, V.S. Amyotrophic lateral sclerosis; fifty cases observed on Guam. J. Nerv. Ment. Dis. 1953, 117, 135–139. [Google Scholar] [CrossRef]
- Kurland, L.T.; Mulder, D.W. Epidemiologic investigations of amyotrophic lateral sclerosis. I. Preliminary report on geographic distribution and special reference to the Mariana Islands, including clinical and pathologic observations. Neurology 1954, 4, 438–448. [Google Scholar] [CrossRef] [PubMed]
- Zimmerman, H. Monthly Report to the Medical Officer in Command; United States Navy Medical Research Unit: San Fransisco, CA, USA, 1945. [Google Scholar]
- Whiting, M. Toxicity of cycads: A literature review. Econ. Bot. 1963, 17, 270–302. [Google Scholar] [CrossRef]
- Dastur, D.K. Cycad toxicity in monkeys: Clinical, pathological, and biochemical aspects. Fed. Proc. 1964, 23, 1368–1369. [Google Scholar] [PubMed]
- Whiting, M.G. Food practices in ALS foci in Japan, the Marianas, and New Guinea. Fed. Proc. 1964, 23, 1343–1345. [Google Scholar] [PubMed]
- Spencer, P.S.; Nunn, P.B.; Hugon, J.; Ludolph, A.; Roy, D.N. Motorneurone disease on Guam: Possible role of a food neurotoxin. Lancet 1986, 1, 965. [Google Scholar] [CrossRef] [PubMed]
- Banack, S.; Cox, P. Distribution of the neurotoxic non-protein amino acid BMAA in Cycas micronesica. Bot. J. Lin. Soc. 2003, 143, 165–168. [Google Scholar] [CrossRef]
- Cox, P.A.; Banack, S.A.; Murch, S.J. Biomagnification of cyanobacterial neurotoxins and neurodegenerative disease among the chamorro people of Guam. Proc. Natl. Acad. Sci. USA 2003, 100, 13380–13383. [Google Scholar] [CrossRef] [PubMed]
- Vega, A.; Bell, E.A. α-amino-β-methylaminopropionic acid, a new amino acid from seeds of Cycas circinalis. Phytochemistry 1967, 6, 759–762. [Google Scholar] [CrossRef]
- Kisby, G.E.; Ellison, M.; Spencer, P.S. Content of the neurotoxins cycasin (methylazoxymethanol β-D-glucoside) and BMAA (β-N-methylamino-l-alanine) in cycad flour prepared by Guam Chamorros. Neurology 1992, 42, 1336–1340. [Google Scholar] [CrossRef] [PubMed]
- Banack, S.A.; Cox, P.A. Biomagnification of cycad neurotoxins in flying foxes: Implications for ALS-PDC in Guam. Neurology 2003, 61, 387–389. [Google Scholar] [CrossRef] [PubMed]
- Cheng, R.; Banack, S.A. Previous studies underestimate BMAA concentrations in cycad flour. Amyotroph. Lateral Scler. 2009, 10, 41–43. [Google Scholar] [CrossRef] [PubMed]
- Banack, S.A.; Murch, S.J.; Cox, P.A. Neurotoxic flying foxes as dietary items for the Chamorro people, Marianas Islands. J. Ethnopharmacol. 2006, 106, 97–104. [Google Scholar] [CrossRef] [PubMed]
- Murch, S.J.; Cox, P.A.; Banack, S.A.; Steele, J.C.; Sacks, O.W. Occurrence of β-N-methylamino-l-alanine (BMAA) in ALS/PDC patients from Guam. Acta Neurol. Scand. 2004, 110, 267–269. [Google Scholar] [CrossRef] [PubMed]
- Garruto, R.M.; Yanagihara, R.; Gajdusek, D.C. Disappearance of high-incidence Amyotrophic Lateral Sclerosis and Parkinsonism-Dementia on Guam. Neurology 1985, 35, 193–198. [Google Scholar] [CrossRef] [PubMed]
- Plato, C.C.; Garruto, R.M.; Galasko, D.; Craig, U.K.; Plato, M.; Gamst, A.; Torres, J.M.; Wiederholt, W. Amyotrophic lateral sclerosis and parkinsonism-dementia complex of Guam: Changing incidence rates during the past 60 years. Am. J. Epidemiol. 2003, 157, 149–157. [Google Scholar] [CrossRef] [PubMed]
- Cox, P.A.; Sacks, O.W. Cycad neurotoxins, consumption of flying foxes, and ALS-PDC disease in Guam. Neurology 2002, 58, 956–959. [Google Scholar] [CrossRef] [PubMed]
- Torres, J.; Iriarte, L.L.; Kurland, L.T. Amyotrophic lateral sclerosis among Guamanians in California. Calif. Med. 1957, 86, 385–388. [Google Scholar] [PubMed]
- Eldridge, R.; Ryan, E.; Rosario, J.; Brody, J.A. Amyotrophic lateral sclerosis and parkinsonism dementia in a migrant population from Guam. Neurology 1969, 19, 1029–1037. [Google Scholar] [CrossRef] [PubMed]
- Garruto, R.M.; Gajdusek, C.; Chen, K.M. Amyotrophic Lateral Sclerosis among Chamorro migrants from Guam. Ann. Neurol. 1980, 8, 612–619. [Google Scholar] [PubMed]
- Garruto, R.M.; Gajdusek, D.C.; Chen, K.M. Amyotrophic lateral sclerosis and parkinsonism-dementia among filipino migrants to Guam. Ann. Neurol. 1981, 10, 341–350. [Google Scholar] [CrossRef] [PubMed]
- Horner, R.D. Amyotrophic lateral sclerosis among 1991 Gulf War veterans: Evidence for a time-limited outbreak. Neuroepidemiology 2008, 31, 28–32. [Google Scholar] [CrossRef] [PubMed]
- Cox, P.A.; Richer, R.; Metcalf, J.S.; Banack, S.A.; Codd, G.A.; Bradley, W.G. Cyanobacteria and BMAA exposure from desert dust: A possible link to sporadic ALS among Gulf War veterans. Amyotroph. Lateral Scler. 2009, 10, 109–117. [Google Scholar] [CrossRef] [PubMed]
- Haley, R.W. Excess incidence of ALS in young Gulf War veterans. Neurology 2003, 61, 750–756. [Google Scholar] [CrossRef] [PubMed]
- Horner, R.D.; Kamins, K.G.; Feussner, J.R.; Grambow, S.C.; Hoff-Lindquist, J.; Harati, Y.; Mitsumoto, H.; Pascuzzi, R.; Spencer, P.S.; Tim, R.; et al. Occurrence of amyotrophic lateral sclerosis among Gulf War veterans. Neurology 2003, 61, 742–749. [Google Scholar] [CrossRef] [PubMed]
- Coffman, C.J.; Horner, R.D.; Grambow, S.C.; Lindquist, J.; Project, V.A.C.S.P. Estimating the occurrence of amyotrophic lateral sclerosis among Gulf War (1990–1991) veterans using capture-recapture methods. Neuroepidemiology 2005, 24, 141–150. [Google Scholar] [CrossRef] [PubMed]
- Caller, T.A.; Doolin, J.W.; Haney, J.F.; Murby, A.J.; West, K.G.; Farrar, H.E.; Ball, A.; Harris, B.T.; Stommel, E.W. A cluster of amyotrophic lateral sclerosis in New Hampshire: A possible role for toxic cyanobacteria blooms. Amyotroph. Lateral Scler. 2009, 10, 101–108. [Google Scholar] [CrossRef] [PubMed]
- Torbick, N.; Hession, S.; Stommel, E.; Caller, T. Mapping amyotrophic lateral sclerosis lake risk factors across northern New England. Int. J. Health Geograph. 2014, 13. [Google Scholar] [CrossRef]
- Downing, S.; Banack, S.A.; Metcalf, J.S.; Cox, P.A.; Downing, T.G. Nitrogen starvation of cyanobacteria results in the production of β-N-methylamino-l-alanine. Toxicon 2011, 58, 187–194. [Google Scholar] [CrossRef] [PubMed]
- Crivelli, A. The destruction of aquatic vegetation by carp. Hydrobiologia 1983, 106, 37–41. [Google Scholar] [CrossRef]
- Gerking, S.D. Chapter 4—Plant-eating fish. In Feeding Ecology of Fish; Gerking, S.D., Ed.; Academic Press: San Diego, CA, USA, 1994; pp. 57–88. [Google Scholar]
- Stommel, E.W.; Field, N.C.; Caller, T.A. Aerosolization of cyanobacteria as a risk factor for Amyotrophic Lateral Sclerosis. Med. Hypotheses 2013, 80, 142–145. [Google Scholar] [CrossRef] [PubMed]
- Banack, S.A.; Downing, T.G.; Spacil, Z.; Purdie, E.L.; Metcalf, J.S.; Downing, S.; Esterhuizen, M.; Codd, G.A.; Cox, P.A. Distinguishing the cyanobacterial neurotoxin β-N-methylamino-l-alanine (BMAA) from its structural isomer 2,4-diaminobutyric acid (2,4-DAB). Toxicon 2010, 56, 868–879. [Google Scholar] [CrossRef] [PubMed]
- Banack, S.A.; Metcalf, J.S.; Jiang, L.; Craighead, D.; Ilag, L.L.; Cox, P.A. Cyanobacteria produce N-(2-aminoethyl)glycine, a backbone for peptide nucleic acids which may have been the first genetic molecules for life on earth. PLoS One 2012, 7, e49043. [Google Scholar] [CrossRef] [PubMed]
- Metcalf, J.S.; Banack, S.A.; Richer, R.; Cox, P.A. Neurotoxic amino acids and their isomers in desert environments. J. Arid Environ. 2015, 112B, 140–144. [Google Scholar] [CrossRef]
- Haney, J.; University of New Hampshire, Durham, NH, USA. Unpublished work. 2014.
- Jiang, L.; Eriksson, J.; Lage, S.; Jonasson, S.; Shams, S.; Mehine, M.; Ilag, L.L.; Rasmussen, U. Diatoms: A novel source for the neurotoxin BMAA in aquatic environments. PLoS One 2014, 9, e84578. [Google Scholar] [CrossRef] [PubMed]
- Banack, S.A.; Metcalf, J.S.; Spacil, Z.; Downing, T.G.; Downing, S.; Long, A.; Nunn, P.B.; Cox, P.A. Distinguishing the cyanobacterial neurotoxin β-N-methylamino-l-alanine (BMAA) from other diamino acids. Toxicon 2011, 57, 730–738. [Google Scholar] [CrossRef] [PubMed]
- Smith, J.L.; Haney, J.F. Foodweb transfer, accumulation, and depuration of microcystins, a cyanobacterial toxin, in pumpkinseed sunfish (Lepomis gibbosus). Toxicon 2006, 48, 580–589. [Google Scholar] [CrossRef] [PubMed]
- Xie, L.; Xie, P.; Guo, L.; Li, L.; Miyabara, Y.; Park, H.D. Organ distribution and bioaccumulation of microcystins in freshwater fish at different trophic levels from the eutrophic Lake Chaohu, China. Environ. Toxicol. 2005, 20, 293–300. [Google Scholar] [CrossRef] [PubMed]
- Jiao, Y.; Chen, Q.; Chen, X.; Wang, X.; Liao, X.; Jiang, L.; Wu, J.; Yang, L. Occurrence and transfer of a cyanobacterial neurotoxin β-N-methylamino-l-alanine within the aquatic food webs of gonghu bay (Lake Taihu, China) to evaluate the potential human health risk. Sci. Total Environ. 2014, 468–469, 457–463. [Google Scholar] [CrossRef] [PubMed]
- Metcalf, J.S.; Banack, S.A.; Lindsay, J.; Morrison, L.F.; Cox, P.A.; Codd, G.A. Co-occurrence of β-N-methylamino-l-alanine, a neurotoxic amino acid with other cyanotoxins in British waterbodies, 1990–2004. Environ. Microbiol. 2008, 10, 702–708. [Google Scholar] [CrossRef] [PubMed]
- Masseret, E.; Banack, S.; Boumediene, F.; Abadie, E.; Brient, L.; Pernet, F.; Juntas-Morales, R.; Pageot, N.; Metcalf, J.; Cox, P.; et al. Dietary BMAA exposure in an Amyotrophic Lateral Sclerosis cluster from Southern France. PLoS One 2013, 8, e83406. [Google Scholar] [CrossRef] [PubMed]
- Brand, L.E.; Pablo, J.; Compton, A.; Hammerschlag, N.; Mash, D.C. Cyanobacterial blooms and the occurrence of the neurotoxin β-N-methylamino-l-alanine (BMAA) in south Florida aquatic food webs. Harmful Algae 2010, 9, 620–635. [Google Scholar] [CrossRef] [PubMed]
- Mondo, K.; Hammerschlag, N.; Basile, M.; Pablo, J.; Banack, S.A.; Mash, D.C. Cyanobacterial neurotoxin β-N-methylamino-l-alanine (BMAA) in shark fins. Mar. Drugs 2012, 10, 509–520. [Google Scholar] [CrossRef] [PubMed]
- Jonasson, S.; Eriksson, J.; Berntzon, L.; Spacil, Z.; Ilag, L.L.; Ronnevi, L.O.; Rasmussen, U.; Bergman, B. Transfer of a cyanobacterial neurotoxin within a temperate aquatic ecosystem suggests pathways for human exposure. Proc. Natl. Acad. Sci. USA 2010, 107, 9252–9257. [Google Scholar] [CrossRef] [PubMed]
- Banack, S.A.; Metcalf, J.S.; Bradley, W.G.; Cox, P.A. Detection of cyanobacterial neurotoxin β-N-methylamino-l-alanine within shellfish in the diet of an ALS patient in florida. Toxicon 2014, 90, 167–173. [Google Scholar] [CrossRef] [PubMed]
- Field, N.C.; Metcalf, J.S.; Caller, T.A.; Banack, S.A.; Cox, P.A.; Stommel, E.W. Linking β-methylamino-l-alanine exposure to sporadic Amyotrophic Lateral Sclerosis in Annapolis, MD. Toxicon 2013, 70C, 179–183. [Google Scholar] [CrossRef] [PubMed]
- Sienko, D.G.; Davis, J.P.; Taylor, J.A.; Brooks, B.R. Amyotrophic lateral sclerosis. A case-control study following detection of a cluster in a small wisconsin community. Arch. Neurol. 1990, 47, 38–41. [Google Scholar] [CrossRef] [PubMed]
- Fleming, L.E.; Kirkpatrick, B.; Backer, L.C.; Bean, J.A.; Wanner, A.; Reich, A.; Zaias, J.; Cheng, Y.S.; Pierce, R.; Naar, J.; et al. Aerosolized red-tide toxins (brevetoxins) and asthma. Chest 2007, 131, 187–194. [Google Scholar] [CrossRef] [PubMed]
- Fleming, L.E.; Bean, J.A.; Kirkpatrick, B.; Cheng, Y.S.; Pierce, R.; Naar, J.; Nierenberg, K.; Backer, L.C.; Wanner, A.; Reich, A.; et al. Exposure and effect assessment of aerosolized red tide toxins (brevetoxins) and asthma. Environ. Health Perspect. 2009, 117, 1095–1100. [Google Scholar] [CrossRef] [PubMed]
- Dondero, T.J., Jr.; Rendtorff, R.C.; Mallison, G.F.; Weeks, R.M.; Levy, J.S.; Wong, E.W.; Schaffner, W. An outbreak of legionnaires’ disease associated with a contaminated air-conditioning cooling tower. N. Engl. J. Med. 1980, 302, 365–370. [Google Scholar] [CrossRef] [PubMed]
- Nygard, K.; Werner-Johansen, O.; Ronsen, S.; Caugant, D.A.; Simonsen, O.; Kanestrom, A.; Ask, E.; Ringstad, J.; Odegard, R.; Jensen, T.; et al. An outbreak of legionnaires disease caused by long-distance spread from an industrial air scrubber in sarpsborg, norway. Clin. Infect. Dis. 2008, 46, 61–69. [Google Scholar] [CrossRef] [PubMed]
- Den Boer, J.W.; Yzerman, E.; van Belkum, A.; Vlaspolder, F.; van Breukelen, F.J. Legionnaire’s disease and saunas. Lancet 1998, 351, 114. [Google Scholar]
- Den Boer, J.W.; Yzerman, E.P.; Schellekens, J.; Lettinga, K.D.; Boshuizen, H.C.; van Steenbergen, J.E.; Bosman, A.; van den Hof, S.; van Vliet, H.A.; Peeters, M.F.; et al. A large outbreak of legionnaires’ disease at a flower show, the netherlands, 1999. Emerg. Infect. Dis. 2002, 8, 37–43. [Google Scholar]
- Morris, J.G., Jr. Pfiesteria, “The cell from hell”, and other toxic algal nightmares. Clin. Infect. Dis. 1999, 28, 1191–1196. [Google Scholar] [CrossRef] [PubMed]
- Cheng, Y.S.; Zhou, Y.; Irvin, C.M.; Kirkpatrick, B.; Backer, L.C. Characterization of aerosols containing microcystin. Mar. Drugs 2007, 5, 136–150. [Google Scholar] [CrossRef] [PubMed]
- Sharma, N.K.; Singh, S. Differential aerosolization of algal and cyanobacterial particles in the atmosphere. Indian J. Microbiol. 2010, 50, 468–473. [Google Scholar] [CrossRef] [PubMed]
- Powell, J.T.; Chatziefthimiou, A.D.; Banack, S.A.; Cox, P.A.; Metcalf, J.S. Desert crust microorganisms, their environment, and human health. J. Arid Environ. 2015, 112B, 127–133. [Google Scholar] [CrossRef]
- Richer, R.; Banack, S.A.; Metcalf, J.S.; Cox, P.A. The persistence of cyanobacterial toxins in desert soils. J. Arid Environ. 2015, 112B, 134–139. [Google Scholar] [CrossRef]
- Danielyan, L.; Schäfer, R.; von Ameln-Mayerhofer, A.; Buadze, M.; Geisler, J.; Klopfer, T.; Burkhardt, U.; Proksch, B.; Verleysdonk, S.; Ayturan, M.; et al. Intranasal delivery of cells to the brain. Eur. J. Cell Biol. 2009, 88, 315–324. [Google Scholar] [CrossRef] [PubMed]
- Boutte, C.; Grubisic, S.; Balthasart, P.; Wilmotte, A. Testing of primers for the study of cyanobacterial molecular diversity by dgge. J. Microbiol. Methods 2006, 65, 542–550. [Google Scholar] [CrossRef] [PubMed]
© 2015 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Banack, S.A.; Caller, T.; Henegan, P.; Haney, J.; Murby, A.; Metcalf, J.S.; Powell, J.; Cox, P.A.; Stommel, E. Detection of Cyanotoxins, β-N-methylamino-L-alanine and Microcystins, from a Lake Surrounded by Cases of Amyotrophic Lateral Sclerosis. Toxins 2015, 7, 322-336. https://doi.org/10.3390/toxins7020322
Banack SA, Caller T, Henegan P, Haney J, Murby A, Metcalf JS, Powell J, Cox PA, Stommel E. Detection of Cyanotoxins, β-N-methylamino-L-alanine and Microcystins, from a Lake Surrounded by Cases of Amyotrophic Lateral Sclerosis. Toxins. 2015; 7(2):322-336. https://doi.org/10.3390/toxins7020322
Chicago/Turabian StyleBanack, Sandra Anne, Tracie Caller, Patricia Henegan, James Haney, Amanda Murby, James S. Metcalf, James Powell, Paul Alan Cox, and Elijah Stommel. 2015. "Detection of Cyanotoxins, β-N-methylamino-L-alanine and Microcystins, from a Lake Surrounded by Cases of Amyotrophic Lateral Sclerosis" Toxins 7, no. 2: 322-336. https://doi.org/10.3390/toxins7020322
APA StyleBanack, S. A., Caller, T., Henegan, P., Haney, J., Murby, A., Metcalf, J. S., Powell, J., Cox, P. A., & Stommel, E. (2015). Detection of Cyanotoxins, β-N-methylamino-L-alanine and Microcystins, from a Lake Surrounded by Cases of Amyotrophic Lateral Sclerosis. Toxins, 7(2), 322-336. https://doi.org/10.3390/toxins7020322