
Botulinum Neurotoxins: Qualitative and Quantitative Analysis Using the Mouse Phrenic Nerve Hemidiaphragm Assay (MPN)


Abstract
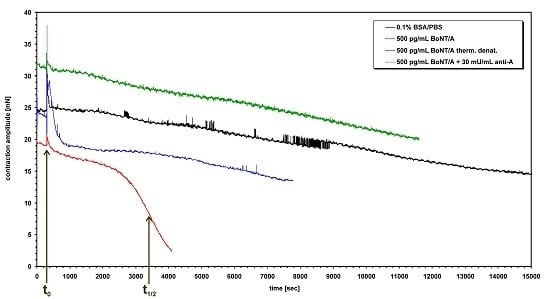
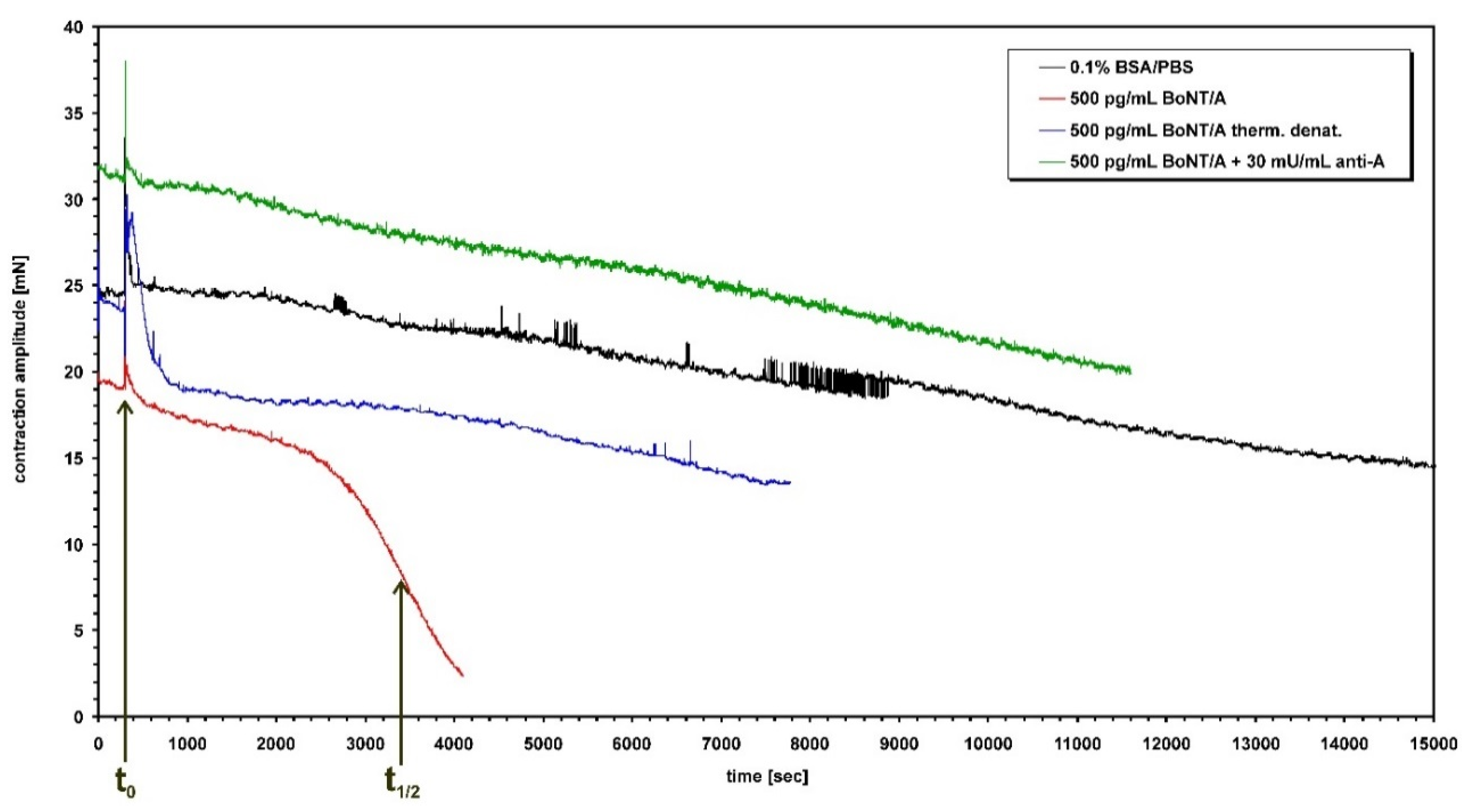
:
1. Introduction
2. Materials and Methods
2.1. Materials

{kind=link}
{kind=link}
{kind=link}
| Sample | Serotype | Matrix | Dilution Factor of Sample | BoNT Concentration | z (BoNT) | Q (BoNT) | (%) | |||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 10,000 | 2000 | 1000 | 500 | 200 | 100 | 50 | 40 | 13 | X1 (ng/mL) | X2 (ng/mL) | N | X (ng/mL) | Xa (ng/mL) | D (ng/mL) | ||||||
| Measured Paralysis Time t1/2 (min) | ||||||||||||||||||||
| S1 | BoNT/A | Meat extract | 142 | 69/78 | 44.0 | 27.0 | 2 | 35.5 | 10.5 | 25.0 | 9.3 | 2.39 | 239 | |||||||
| S2 | BoNT/A | 0.1% BSA/PBS | 120 | 88/83 | 17.4 | 21.8 | 2 | 19.6 | 9.9 | 9.7 | 3.9 | 0.98 | 98 | |||||||
| S3 | - | 0.1% BSA/PBS | >160 | >180 | >180 | >180 | 4 | - | - | - | - | - | - | |||||||
| S4 | BoNT/E | 0.1% BSA/PBS | 154 | 116 | 118 | 92/82 | 12.5 | 18.3 | 2 | 15.4 | 10.9 | 4.5 | 1.6 | 0.42 | 42 | |||||
| S5 | BoNT/A | Meat extract | 65/70 | 277 | 209 | 2 | 243 | 108 | 135 | 4.9 | 1.25 | 125 | ||||||||
| S6 | BoNT/B | 0.1% BSA/PBS | >180 | 87/88 | 10.0 | 9.0 | 2 | 9.5 | 9.0 | 0.5 | 0.2 | 0.06 | 5.7 | |||||||
| S7 | BoNT/A | 0.1% BSA/PBS | 65/60 | 277 | 376 | 2 | 327 | 100 | 227 | 8.9 | 2.27 | 227 | ||||||||
| S8A | BoNT/A | 0.1% BSA/PBS | 138 | 83/79 | 22.0 # § | 26.3 # § | 2 | 24.2 | 4.7 | 19 | 16 | 4.11 | 411 | |||||||
| 11.4 * § | 8.7 * § | 2 | 10.0 | 4.7 | 5.3 | |||||||||||||||
| S8B | BoNT/B | 0.1% BSA/PBS | 5.60 § | 4.30 § | 2 | 4.95 | 4.5 | 0.4 | 0.4 | 0.1 | 10 | |||||||||
| S9 | BoNT/A | 0.1% BSA/PBS | >180 | 121/120 | 1.08 | 1.04 | 2 | 1.06 | 0.5 | 0.6 | 4.5 | 1.14 | 114 | |||||||
| S10 | BoNT/A | Milk | 139 | 69/85 | 44.0 | 19.8 | 2 | 31.9 | 10.3 | 22 | 8.2 | 2.09 | 209 | |||||||
| S11 | BoNT/A | Serum | 121 | 74/81 | 22.6 | 24.0 | 2 | 23.3 | 9.8 | 13 | 5.4 | 1.37 | 137 | |||||||
| S12 | BoNT/A | 0.1% BSA/PBS | 66/64 | 36 | 35 | 2.617 # | 2.942 # | 2 | 2.75 | 1001 | −998 | −3.9 | −1 | −100 | ||||||
| 2617 * | 2942 * | 2 | 2780 | 1001 | 1779 | |||||||||||||||
| S13 | BoNT/A | Milk | 59/54 | 401 | 562 | 2 | 482 | 112 | 370 | 13 | 3.31 | 331 | ||||||||
| Anti-BoNT Antiserum | A | B | E | A + B | A + E | B + E | BoNT Serotype | |
|---|---|---|---|---|---|---|---|---|
| Sample # | Paralysis Time t1/2 (min) | Reported | Assigned | |||||
| S1 | >180 | A | A | |||||
| S2 | >180 | A | A | |||||
| S4 | 85 | 74 | >180 | E | E | |||
| S5 | >180 | A | A | |||||
| S6 | 109/103 | >180 | 91 | B | B | |||
| S7 | >180 | A | A | |||||
| S8 | 134/121 | 82 | 80 | >180 | 116 | 88 | A + B | A + B |
| S9 | >180 | A | A | |||||
| S10 | >180 | A | A | |||||
| S11 | >180 | A | A | |||||
| S12 | >180 | A | A | |||||
| S13 | >180 | A | A | |||||
2.2. Experimental Procedures
3. Results
4. Discussion
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Dorner, M.B.; Schulz, K.M.; Kull, S.; Dorner, B.G. Complexity of botulinum neurotoxins: Challenges for detection technology. Curr. Top. Microbiol. Immunol. 2013, 364, 219–255. [Google Scholar] [PubMed]
- Lindström, M.; Korkeala, H. Laboratory diagnostics of botulism. Clin. Microbiol. Rev. 2006, 19, 298–314. [Google Scholar] [CrossRef] [PubMed]
- Reed, L.J.; Muench, H. A simple method of estimating 50% endpoints. Am. J. Hyg. 1938, 27, 493–497. [Google Scholar]
- Pearce, L.B.; Borodic, G.E.; First, E.R.; MacCallum, R.D. Measurement of botulinum toxin activity: Evaluation of the lethality assay. Toxicol. Appl. Pharmacol. 1994, 128, 69–77. [Google Scholar] [CrossRef] [PubMed]
- Pearce, L.B.; Borodic, G.E.; Johnson, E.A.; First, E.R.; MacCallum, R. The median paralysis unit: A more pharmacologically relevant unit of biologic activity for botulinum toxin. Toxicon 1995, 33, 217–227. [Google Scholar] [CrossRef]
- Pearce, L.B.; First, E.R.; Borodic, G.E. Botulinum toxin potency: A mystery resolved by the median paralysis. J. R. Soc. Med. 1994, 87, 571–572. [Google Scholar] [PubMed]
- Deutsches Institut für Normung. Microbiological Analysis of Meat and Meat Products; Detection of Clostridium Botulinum and Botulinum Toxin; Beuth Verlag: Berlin, Germany, 1988; Volume 10102, pp. 1–5. [Google Scholar]
- Anon. AOAC official method. In Clostridium Botulinum and Its Toxins in Food; AOAC International: Rockville, MD, USA, 1977; Volume 977, p. 26. [Google Scholar]
- Adler, S.; Bicker, G.; Bigalke, H.; Bishop, C.; Blumel, J.; Dressler, D.; Fitzgerald, J.; Gessler, F.; Heuschen, H.; Kegel, B.; et al. The current scientific and legal status of alternative methods to the LD50 test for botulinum neurotoxin potency testing. The report and recommendations of a ZEBET Expert Meeting. Altern. Lab. Anim. 2010, 38, 315–330. [Google Scholar] [PubMed]
- Boroff, D.A.; Fleck, U. Statistical analysis of a rapid in vivo method for the titration of the toxin of Clostridium botulinum. J. Bacteriol. 1966, 92, 1580–1581. [Google Scholar] [PubMed]
- Bülbring, E. Observations on the isolated phrenic nerve diaphragm preparation of the rat. Br. J. Pharmacol. Chemother. 1946, 1, 38–61. [Google Scholar] [CrossRef] [PubMed]
- Habermann, E.; Dreyer, F.; Bigalke, H. Tetanus toxin blocks the neuromuscular transmission in vitro like botulinum A toxin. Naunyn Schmiedeberg’s Arch. Pharmacol. 1980, 311, 33–40. [Google Scholar] [CrossRef]
- Simpson, L.L.; Rapport, M.M. The binding of botulinum toxin to membrane lipids: Sphingolipids, steroids and fatty acids. J. Neurochem. 1971, 18, 1751–1759. [Google Scholar] [CrossRef] [PubMed]
- Rummel, A.; Häfner, K.; Mahrhold, S.; Darashchonak, N.; Holt, M.; Jahn, R.; Beermann, S.; Karnath, T.; Bigalke, H.; Binz, T. Botulinum neurotoxins C, E and F bind gangliosides via a conserved binding site prior to stimulation-dependent uptake with botulinum neurotoxin F utilising the three isoforms of SV2 as second receptor. J. Neurochem. 2009, 110, 1942–1954. [Google Scholar] [CrossRef] [PubMed]
- Wohlfarth, K.; Göschel, H.; Frevert, J.; Dengler, R.; Bigalke, H. Botulinum A toxins: Units versus units. Naunyn Schmiedeberg’s Arch. Pharmacol. 1997, 355, 335–340. [Google Scholar] [CrossRef]
- Rasetti-Escargueil, C.; Jones, R.G.; Liu, Y.; Sesardic, D. Measurement of botulinum types A, B and E neurotoxicity using the phrenic nerve-hemidiaphragm: Improved precision with in-bred mice. Toxicon 2009, 53, 503–511. [Google Scholar] [CrossRef] [PubMed]
- EDQM. Botulinum Toxin Type A for Injection. In European Pharmacopoeia, 5th ed.; Council of Europe; 2006; pp. 1117–1119. [Google Scholar]
- Pirazzini, M.; Henke, T.; Rossetto, O.; Mahrhold, S.; Krez, N.; Rummel, A.; Montecucco, C.; Binz, T. Neutralisation of specific surface carboxylates speeds up translocation of botulinum neurotoxin type B enzymatic domain. FEBS Lett. 2013, 587, 3831–3836. [Google Scholar] [CrossRef] [PubMed]
- Rummel, A.; Bade, S.; Alves, J.; Bigalke, H.; Binz, T. Two carbohydrate binding sites in the HCC-domain of tetanus neurotoxin are required for toxicity. J. Mol. Biol. 2003, 326, 835–847. [Google Scholar] [CrossRef]
- Rummel, A.; Eichner, T.; Weil, T.; Karnath, T.; Gutcaits, A.; Mahrhold, S.; Sandhoff, K.; Proia, R.L.; Acharya, K.R.; Bigalke, H.; et al. Identification of the protein receptor binding site of botulinum neurotoxins B and G proves the double-receptor concept. Proc. Natl. Acad. Sci. USA 2007, 104, 359–364. [Google Scholar] [CrossRef] [PubMed]
- Rummel, A.; Mahrhold, S.; Bigalke, H.; Binz, T. The HCC-domain of botulinum neurotoxins A and B exhibits a singular ganglioside binding site displaying serotype specific carbohydrate interaction. Mol. Microbiol. 2004, 51, 631–643. [Google Scholar] [CrossRef] [PubMed]
- Rummel, A.; Mahrhold, S.; Bigalke, H.; Binz, T. Exchange of the HCC domain mediating double receptor recognition improves the pharmacodynamic properties of botulinum neurotoxin. FEBS J. 2011, 278, 4506–4515. [Google Scholar] [CrossRef] [PubMed]
- Mahrhold, S.; Strotmeier, J.; Garcia-Rodriguez, C.; Lou, J.; Marks, J.D.; Rummel, A.; Binz, T. Identification of the SV2 protein receptor-binding site of botulinum neurotoxin type E. Biochem. J. 2013, 453, 37–47. [Google Scholar] [CrossRef] [PubMed]
- Strotmeier, J.; Gu, S.; Jutzi, S.; Mahrhold, S.; Zhou, J.; Pich, A.; Eichner, T.; Bigalke, H.; Rummel, A.; Jin, R.; et al. The biological activity of botulinum neurotoxin type C is dependent upon novel types of ganglioside binding sites. Mol. Microbiol. 2011, 81, 143–156. [Google Scholar] [CrossRef] [PubMed]
- Strotmeier, J.; Lee, K.; Volker, A.K.; Mahrhold, S.; Zong, Y.; Zeiser, J.; Zhou, J.; Pich, A.; Bigalke, H.; Binz, T.; et al. Botulinum neurotoxin serotype D attacks neurons via two carbohydrate-binding sites in a ganglioside-dependent manner. Biochem. J. 2010, 431, 207–216. [Google Scholar] [CrossRef] [PubMed]
- Strotmeier, J.; Mahrhold, S.; Krez, N.; Janzen, C.; Lou, J.; Marks, J.D.; Binz, T.; Rummel, A. Identification of the synaptic vesicle glycoprotein 2 receptor binding site in botulinum neurotoxin A. FEBS Lett. 2014, 588, 1087–1093. [Google Scholar] [CrossRef] [PubMed]
- Gu, S.; Rumpel, S.; Zhou, J.; Strotmeier, J.; Bigalke, H.; Perry, K.; Shoemaker, C.B.; Rummel, A.; Jin, R. Botulinum neurotoxin is shielded by NTNHA in an interlocked complex. Science 2012, 335, 977–981. [Google Scholar] [CrossRef] [PubMed]
- Sheridan, R.E.; Deshpande, S.S.; Smith, T. Comparison of in vivo and in vitro mouse bioassays for botulinum toxin antagonists. J. Appl. Toxicol. 1999, 19, S29–S33. [Google Scholar] [CrossRef]
- Sheridan, R.E.; Smith, T.J.; Adler, M. Primary cell culture for evaluation of botulinum neurotoxin antagonists. Toxicon 2005, 45, 377–382. [Google Scholar] [CrossRef] [PubMed]
- Sheridan, R.E.; Deshpande, S.S.; Nicholson, J.D.; Adler, M. Structural features of aminoquinolines necessary for antagonist activity against botulinum neurotoxin. Toxicon 1997, 35, 1439–1451. [Google Scholar] [CrossRef]
- Adler, M.; Scovill, J.; Parker, G.; Lebeda, F.J.; Piotrowski, J.; Deshpande, S.S. Antagonism of botulinum toxin-induced muscle weakness by 3,4-diaminopyridine in rat phrenic nerve-hemidiaphragm preparations. Toxicon 1995, 33, 527–537. [Google Scholar] [CrossRef]
- Eleopra, R.; Tugnoli, V.; Quatrale, R.; Gastaldo, E.; Rossetto, O.; de Grandis, D.; Montecucco, C. Botulinum neurotoxin serotypes A and C do not affect motor units survival in humans: An electrophysiological study by motor units counting. Clin. Neurophysiol. 2002, 113, 1258–1264. [Google Scholar] [CrossRef]
- Eleopra, R.; Montecucco, C.; Devigili, G.; Lettieri, C.; Rinaldo, S.; Verriello, L.; Pirazzini, M.; Caccin, P.; Rossetto, O. Botulinum neurotoxin serotype D is poorly effective in humans: An in vivo electrophysiological study. Clin. Neurophysiol. 2013, 124, 999–1004. [Google Scholar] [CrossRef] [PubMed]
- Rummel, A.; Karnath, T.; Henke, T.; Bigalke, H.; Binz, T. Synaptotagmins I and II act as nerve cell receptors for botulinum neurotoxin G. J. Boil. Chem. 2004, 279, 30865–30870. [Google Scholar] [CrossRef] [PubMed]
- Mahrhold, S.; Rummel, A.; Bigalke, H.; Davletov, B.; Binz, T. The synaptic vesicle protein 2C mediates the uptake of botulinum neurotoxin A into phrenic nerves. FEBS Lett. 2006, 580, 2011–2014. [Google Scholar] [CrossRef] [PubMed]
- Simpson, L.L. Studies on the binding of botulinum toxin type A to the rat phrenic nerve-hemidiaphragm preparation. Neuropharmacology 1974, 13, 683–691. [Google Scholar] [CrossRef]
- Göschel, H.; Wohlfarth, K.; Frevert, J.; Dengler, R.; Bigalke, H. Botulinum A toxin therapy: Neutralizing and nonneutralizing antibodies—Therapeutic consequences. Exp. Neurol. 1997, 147, 96–102. [Google Scholar] [CrossRef] [PubMed]
- Dressler, D.; Dirnberger, G.; Bhatia, K.P.; Irmer, A.; Quinn, N.P.; Bigalke, H.; Marsden, C.D. Botulinum toxin antibody testing: Comparison between the mouse protection assay and the mouse lethality assay. Mov. Disord. 2000, 15, 973–976. [Google Scholar] [CrossRef]
- Herrmann, J.; Mall, V.; Bigalke, H.; Geth, K.; Korinthenberg, R.; Heinen, F. Secondary non-response due to development of neutralising antibodies to botulinum toxin A during treatment of children with cerebral palsy. Neuropediatrics 2000, 31, 333–334. [Google Scholar] [CrossRef] [PubMed]
- Rollnik, J.D.; Wohlfarth, K.; Dengler, R.; Bigalke, H. Neutralizing botulinum toxin type a antibodies: Clinical observations in patients with cervical dystonia. Neurol. Clin. Neurophysiol. 2001, 2001, 2–4. [Google Scholar] [CrossRef] [PubMed]
- Dressler, D.; Bigalke, H. Botulinum toxin antibody type A titres after cessation of botulinum toxin therapy. Mov. Disord. 2002, 17, 170–173. [Google Scholar] [CrossRef] [PubMed]
- Dressler, D.; Bigalke, H.; Benecke, R. Botulinum toxin type B in antibody-induced botulinum toxin type A therapy failure. J. Neurol. 2003, 250, 967–969. [Google Scholar] [CrossRef] [PubMed]
- Dressler, D.; Bigalke, H. Antibody-induced failure of botulinum toxin type B therapy in de novo patients. Eur. Neurol. 2004, 52, 132–135. [Google Scholar] [CrossRef] [PubMed]
- Herrmann, J.; Geth, K.; Mall, V.; Bigalke, H.; Monting, S.J.; Linder, M.; Kirschner, J.; Berweck, S.; Korinthenberg, R.; Heinen, F.; et al. Clinical impact of antibody formation to botulinum toxin A in children. Ann. Neurol. 2004, 55, 732–735. [Google Scholar] [CrossRef] [PubMed]
- Dressler, D.; Lange, M.; Bigalke, H. Mouse diaphragm assay for detection of antibodies against botulinum toxin type B. Mov. Disord. 2005, 20, 1617–1619. [Google Scholar] [CrossRef] [PubMed]
- Lange, O.; Bigalke, H.; Dengler, R.; Wegner, F.; deGroot, M.; Wohlfarth, K. Neutralizing antibodies and secondary therapy failure after treatment with botulinum toxin type A: Much ado about nothing? Clin. Neuropharmacol. 2009, 32, 213–218. [Google Scholar] [CrossRef] [PubMed]
- Rasetti-Escargueil, C.; Liu, Y.; Rigsby, P.; Jones, R.G.; Sesardic, D. Phrenic nerve-hemidiaphragm as a highly sensitive replacement assay for determination of functional botulinum toxin antibodies. Toxicon 2011, 57, 1008–1016. [Google Scholar] [CrossRef] [PubMed]
- Miethe, S.; Rasetti-Escargueil, C.; Liu, Y.; Chahboun, S.; Pelat, T.; Avril, A.; Frenzel, A.; Schirrmann, T.; Thullier, P.; Sesardic, D.; et al. Development of neutralizing scFv-Fc against botulinum neurotoxin A light chain from a macaque immune library. mAbs 2014, 6, 446–459. [Google Scholar] [CrossRef] [PubMed]
- Sesardic, D.; Jones, R.G.; Leung, T.; Alsop, T.; Tierney, R. Detection of antibodies against botulinum toxins. Mov. Disord. 2004, 19, S85–S91. [Google Scholar] [CrossRef] [PubMed]
- Nowakowski, A.; Wang, C.; Powers, D.B.; Amersdorfer, P.; Smith, T.J.; Montgomery, V.A.; Sheridan, R.; Blake, R.; Smith, L.A.; Marks, J.D. Potent neutralization of botulinum neurotoxin by recombinant oligoclonal antibody. Proc. Natl. Acad. Sci. USA 2002, 99, 11346–11350. [Google Scholar] [CrossRef] [PubMed]
- Amersdorfer, P.; Wong, C.; Smith, T.; Chen, S.; Deshpande, S.; Sheridan, R.; Marks, J.D. Genetic and immunological comparison of anti-botulinum type A antibodies from immune and non-immune human phage libraries. Vaccine 2002, 20, 1640–1648. [Google Scholar] [CrossRef]
- Amersdorfer, P.; Wong, C.; Chen, S.; Smith, T.; Deshpande, S.; Sheridan, R.; Finnern, R.; Marks, J.D. Molecular characterization of murine humoral immune response to botulinum neurotoxin type A binding domain as assessed by using phage antibody libraries. Infect. Immun. 1997, 65, 3743–3752. [Google Scholar] [PubMed]
- Merz, B.; Bigalke, H.; Stoll, G.; Naumann, M. Botulism type B presenting as pure autonomic dysfunction. Clin. Auton. Res. 2003, 13, 337–338. [Google Scholar] [CrossRef] [PubMed]
- Worbs, S.; Fiebig, U.; Zeleny, R.; Schimmel, H.; Rummel, A.; Luginbühl, W.; Dorner, B.G. Qualitative and quantitative detection of botulinum neurotoxins from complex matrices: Results of the first international proficiency test. Toxins 2015, 7. [Google Scholar] [CrossRef]
- Weisemann, J.; Krez, N.; Fiebig, U.; Worbs, S.; Skiba, M.; Endermann, T.; Dorner, M.B.; Bergström, T.; Muñoz, A.; Zegers, I.; et al. Generation and characterisation of six recombinant botulinum neurotoxins as reference material to serve in an international proficiency. Toxins 2015, 7. [Google Scholar] [CrossRef]
- Bigalke, H.; Rummel, A. Medical aspects of toxin weapons. Toxicology 2005, 214, 210–220. [Google Scholar] [CrossRef] [PubMed]
- Bigalke, H. Botulinum toxin: Application, safety, and limitations. Curr. Top. Microbiol. Immunol. 2013, 364, 307–317. [Google Scholar] [PubMed]
- Pickett, A.M.; Hambleton, P. Dose standardisation of botulinum toxin. Lancet 1994, 344, 474–475. [Google Scholar] [CrossRef]
- Rossetto, O.; Pirazzini, M.; Montecucco, C. Botulinum neurotoxins: Genetic, structural and mechanistic insights. Nat. Rev. Microbiol. 2014, 12, 535–549. [Google Scholar] [CrossRef] [PubMed]
- Dover, N.; Barash, J.R.; Hill, K.K.; Xie, G.; Arnon, S.S. Molecular characterization of a novel botulinum neurotoxin type h gene. J. Infect. Dis. 2014, 209, 192–202. [Google Scholar] [CrossRef] [PubMed]
- Pellett, S. Progress in cell based assays for botulinum neurotoxin detection. Curr. Top. Microbiol. Immunol. 2013, 364, 257–285. [Google Scholar] [PubMed]
- Wiley, R.G.; Oeltmann, T.N. Anatomically selective peripheral nerve ablation using intraneural ricin injection. J. Neurosci. Methods 1986, 17, 43–53. [Google Scholar] [CrossRef]
- Kalb, S.R.; Baudys, J.; Wang, D.; Barr, J.R. Recommended mass spectrometry-based strategies to identify botulinum neurotoxin-containing samples. Toxins 2015, 7, 1765–1778. [Google Scholar] [CrossRef] [PubMed]
© 2015 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons by Attribution (CC-BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bigalke, H.; Rummel, A. Botulinum Neurotoxins: Qualitative and Quantitative Analysis Using the Mouse Phrenic Nerve Hemidiaphragm Assay (MPN). Toxins 2015, 7, 4895-4905. https://doi.org/10.3390/toxins7124855
Bigalke H, Rummel A. Botulinum Neurotoxins: Qualitative and Quantitative Analysis Using the Mouse Phrenic Nerve Hemidiaphragm Assay (MPN). Toxins. 2015; 7(12):4895-4905. https://doi.org/10.3390/toxins7124855
Chicago/Turabian StyleBigalke, Hans, and Andreas Rummel. 2015. "Botulinum Neurotoxins: Qualitative and Quantitative Analysis Using the Mouse Phrenic Nerve Hemidiaphragm Assay (MPN)" Toxins 7, no. 12: 4895-4905. https://doi.org/10.3390/toxins7124855
APA StyleBigalke, H., & Rummel, A. (2015). Botulinum Neurotoxins: Qualitative and Quantitative Analysis Using the Mouse Phrenic Nerve Hemidiaphragm Assay (MPN). Toxins, 7(12), 4895-4905. https://doi.org/10.3390/toxins7124855