Downregulation and Mutation of a Cadherin Gene Associated with Cry1Ac Resistance in the Asian Corn Borer, Ostrinia furnacalis (Guenée)



Abstract
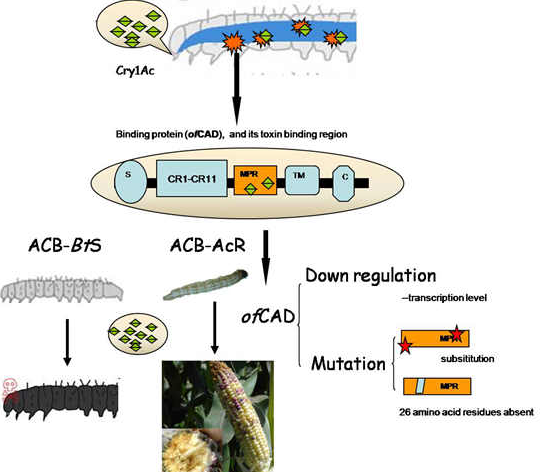
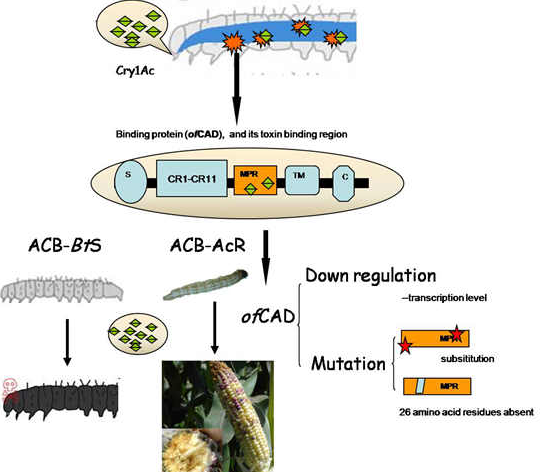
:
1. Introduction
2. Results
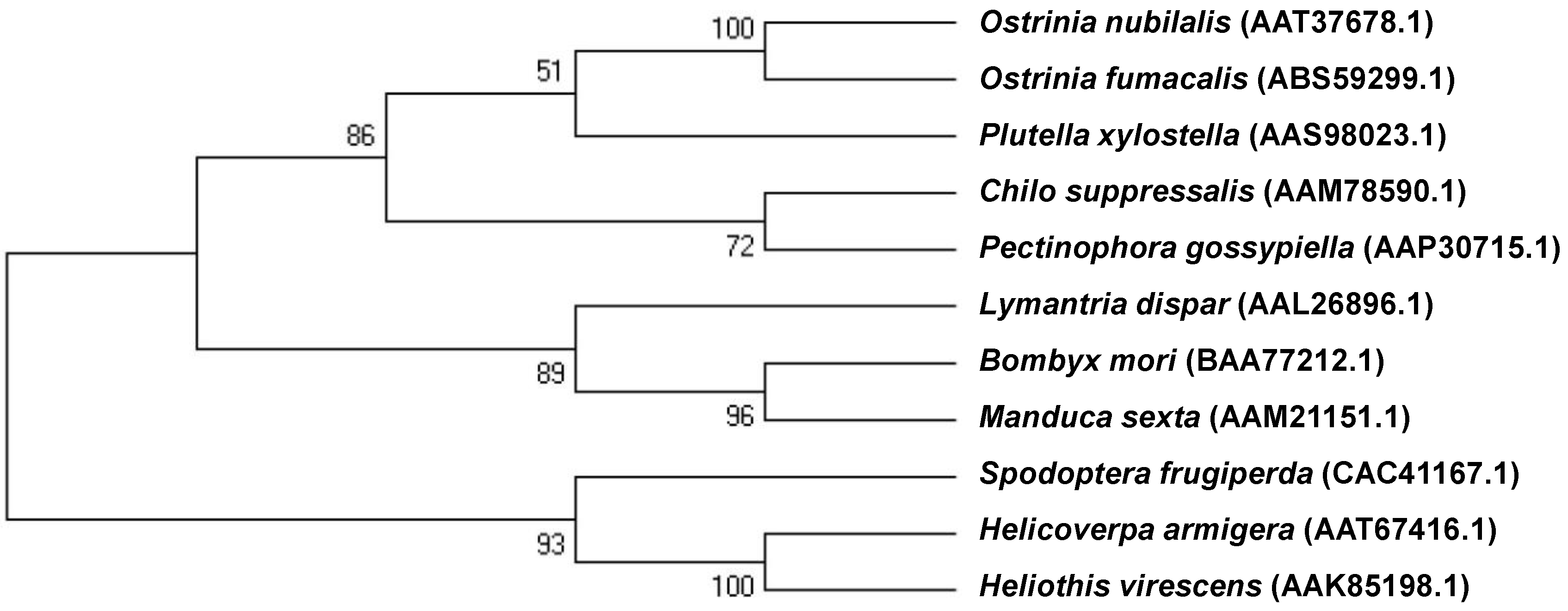
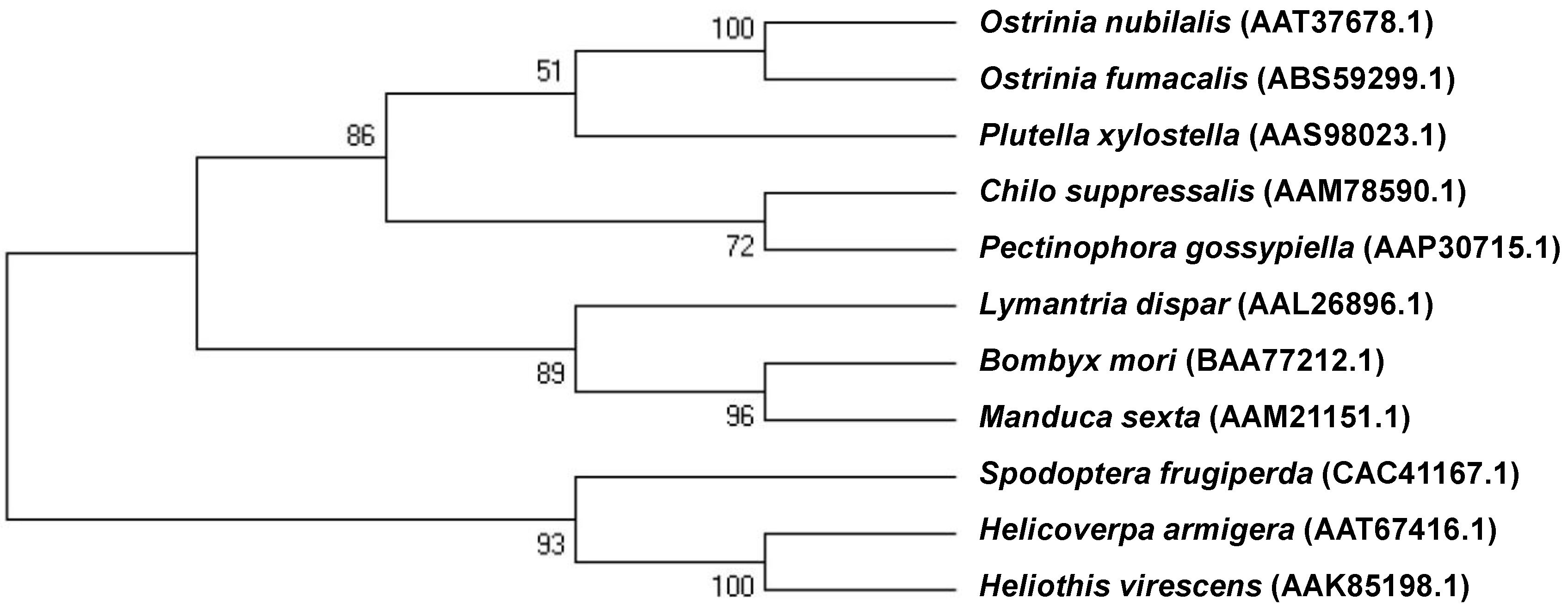
2.1. cDNA of the Cadherin-Like Protein in Asian Corn Borer (ACB)

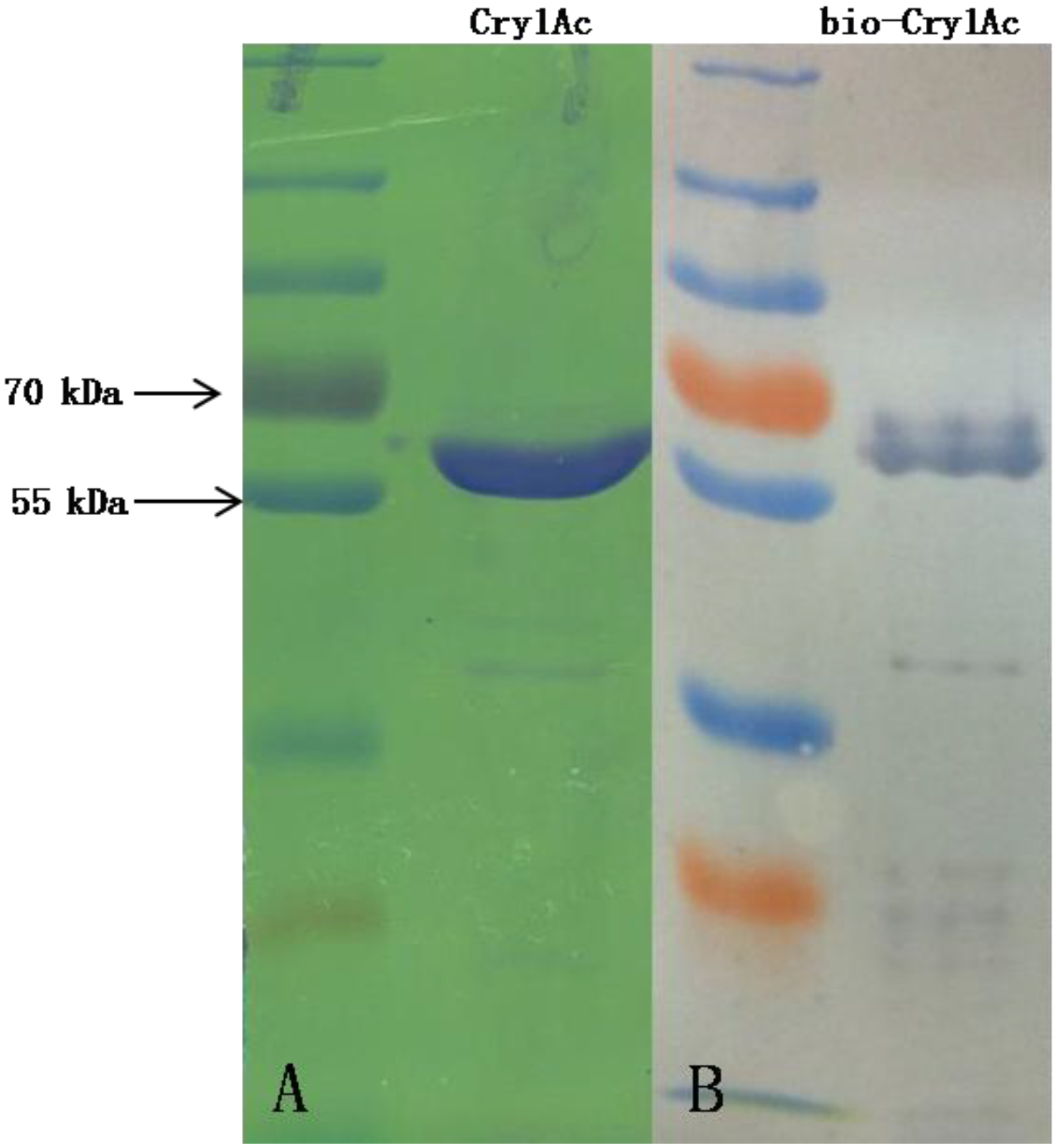
2.2. Recombinant Expression of OfCAD Internal Peptides and Binding Studies


{kind=link}
{kind=link}
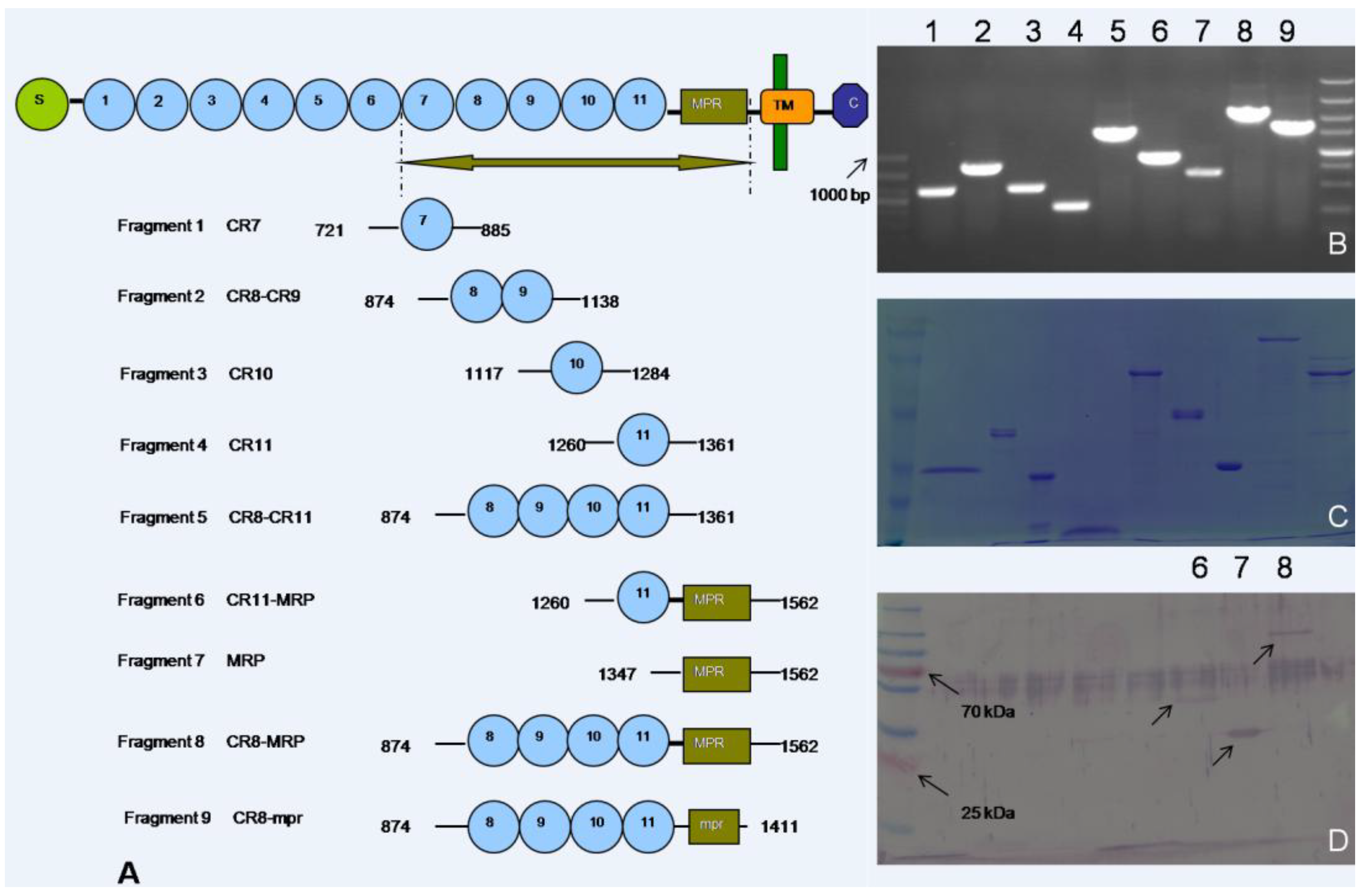{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Fragment | 1 | 2 | 3 | 4 | 5 | 6 | 7 | 8 | 9 |
|---|---|---|---|---|---|---|---|---|---|
| No. of amino acid residues | 216 | 315 | 216 | 153 | 538 | 354 | 267 | 739 | 588 |
| Molecular weight (kDa) | 24 | 36 | 24 | 17 | 60 | 39 | 29 | 82 | 66 |
| Express in supernatant | + | − | − | − | − | − | + | + | − |
| Express as inclusion body | + | + | + | + | + | + | − | + | + |
2.3. MPR Mutants in ACB-Ac200 and Their Binding Ability to Cry1Ac
| Fragment | Primers | Position | Primer DNA sequence | Product |
|---|---|---|---|---|
| For cloning of ofcad | ||||
| ofcad-F | 223 | CARSTBATMTWYRTGGAKGARGA | ||
| ofcad-R | 4870 | TCVACRGYRTGYTTGTTRGTRCC | ||
| GSP1 | 792 | CGAGTCAGAAGCTGTGACGCT | ||
| GSP2 | 536 | CAGGCGCTGACCACTTGCAGGAT | ||
| AUAP | -- | CAGGCGCTGACCACTTGCAGGAT | ||
| ofcad-3F | 4178 | AGACCGTGCGAGCATGCAGCT | ||
| 3 sites adaptor | -- | CTGATCTAGAGGTACCGGATCC | ||
| For generating OfCAD fragments | ||||
| Fragment1 | F1 | 2161 | CCGGAATTCCGCGTGAGGGACCTCAACACT | 507 |
| R1 | 2633 | CCGCTCGAGTTACACCGTCGTGTTGAACTTCTCAG | ||
| Fragment2 | F2 | 2623 | CCAGAATTCGAGCCGCTCACTGAGAAGTTCAAC | 803 |
| R2 | 3394 | GCCTCGAGTTACGCCAGAACGCCGCCTTCTGT | ||
| Fragment3 | F3 | 3358 | CCAGAATTCCCCGACTCCGTCATTCGGCTTTC | 507 |
| R3 | 3829 | CCGCTCGAGTTAGCCGTCAATAAACCTGTAGTAGAT | ||
| Fragment4 | F4 | 3778 | CCGGAATTCCCGCAAGCAGAAGACCTTAAAAACCA | 308 |
| R4 | 4063 | CCGCTCGAGTTACGTCGAAATGCCCGCTGTGTA | ||
| Fragment5 | F2 | 2623 | CCAGAATTCGAGCCGCTCACTGAGAAGTTCAAC | 1473 |
| R4 | 4063 | CCGCTCGAGTTACGTCGAAATGCCCGCTGTGTA | ||
| Fragment6 | F4 | 3778 | CCGGAATTCCCGCAAGCAGAAGACCTTAAAAACCA | 921 |
| R5 | 4667 | CCGCTCGAGTTACTCGCCTAGCGTCGGAGTGT | ||
| Fragment7 | F5 | 4039 | CCGGAATTCCGCCCAATTTTCGAGCAGGACCTTTA | 660 |
| R5 | 4667 | CCGCTCGAGTTACTCGCCTAGCGTCGGAGTGT | ||
| Fragment8 | F2 | 2623 | CCAGAATTCGAGCCGCTCACTGAGAAGTTCAAC | 2076 |
| R5 | 4667 | CCGCTCGAGTTACTCGCCTAGCGTCGGAGTGT | ||
| Fragment9 | F2 | 2623 | CCAGAATTCGAGCCGCTCACTGAGAAGTTCAAC | 1623 |
| R6 | 4213 | CCGCTCGAGTTAGGTGGTCGCATGCAGCGTGAA | ||
| For qPCR | ||||
| qcad-F | 4329 | AGCCCGTGTGAAAGTCTAC | 255 | |
| qcad-R | 4565 | ACGGCTTCGACCTCTTGTG | ||
| qβ-F | CCTCCACCTCCCTCGAGAAG | |||
| qβ-R | GGCAACGGAACCTCTCGTTA | |||

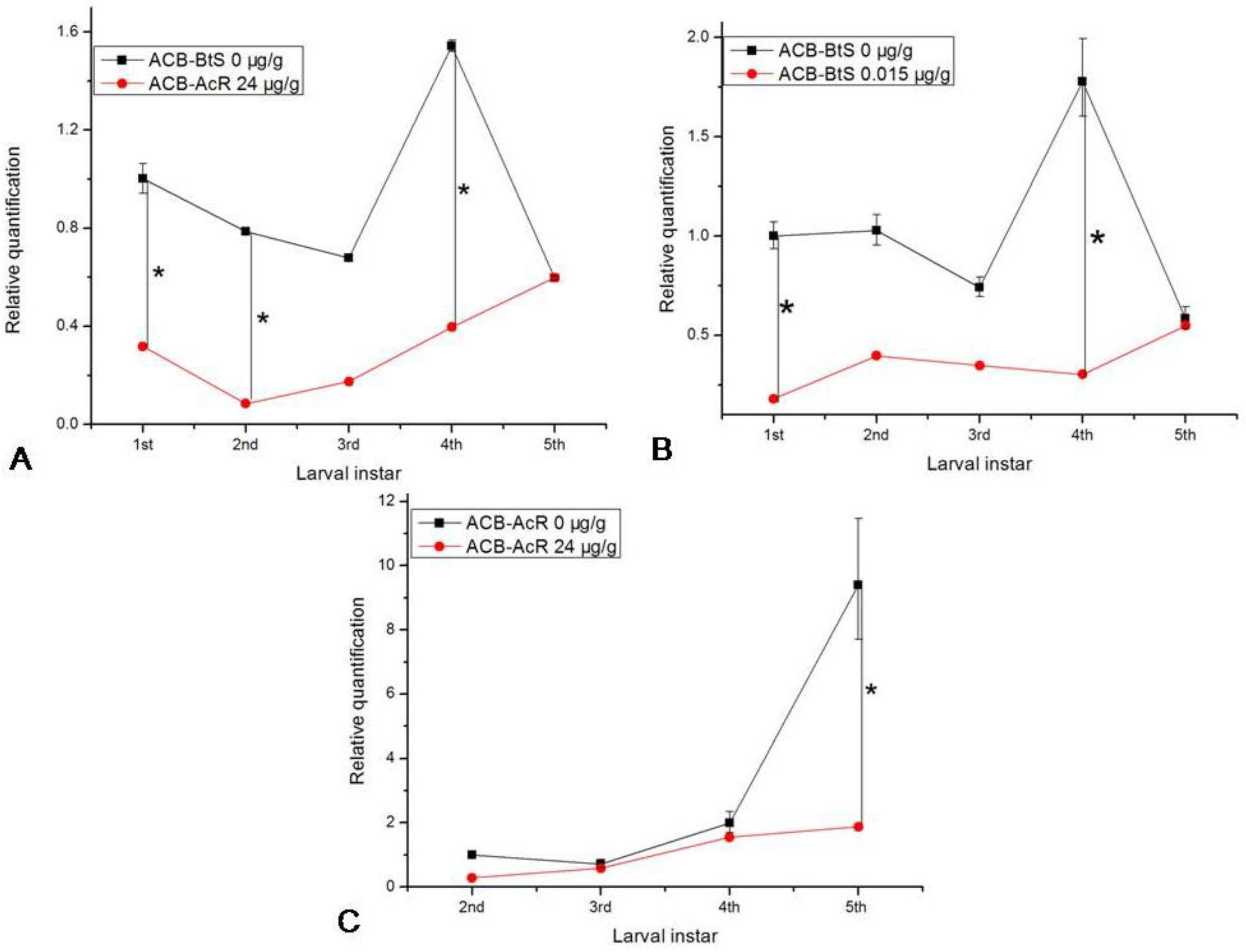
2.4. Transcription Levels of ofcad
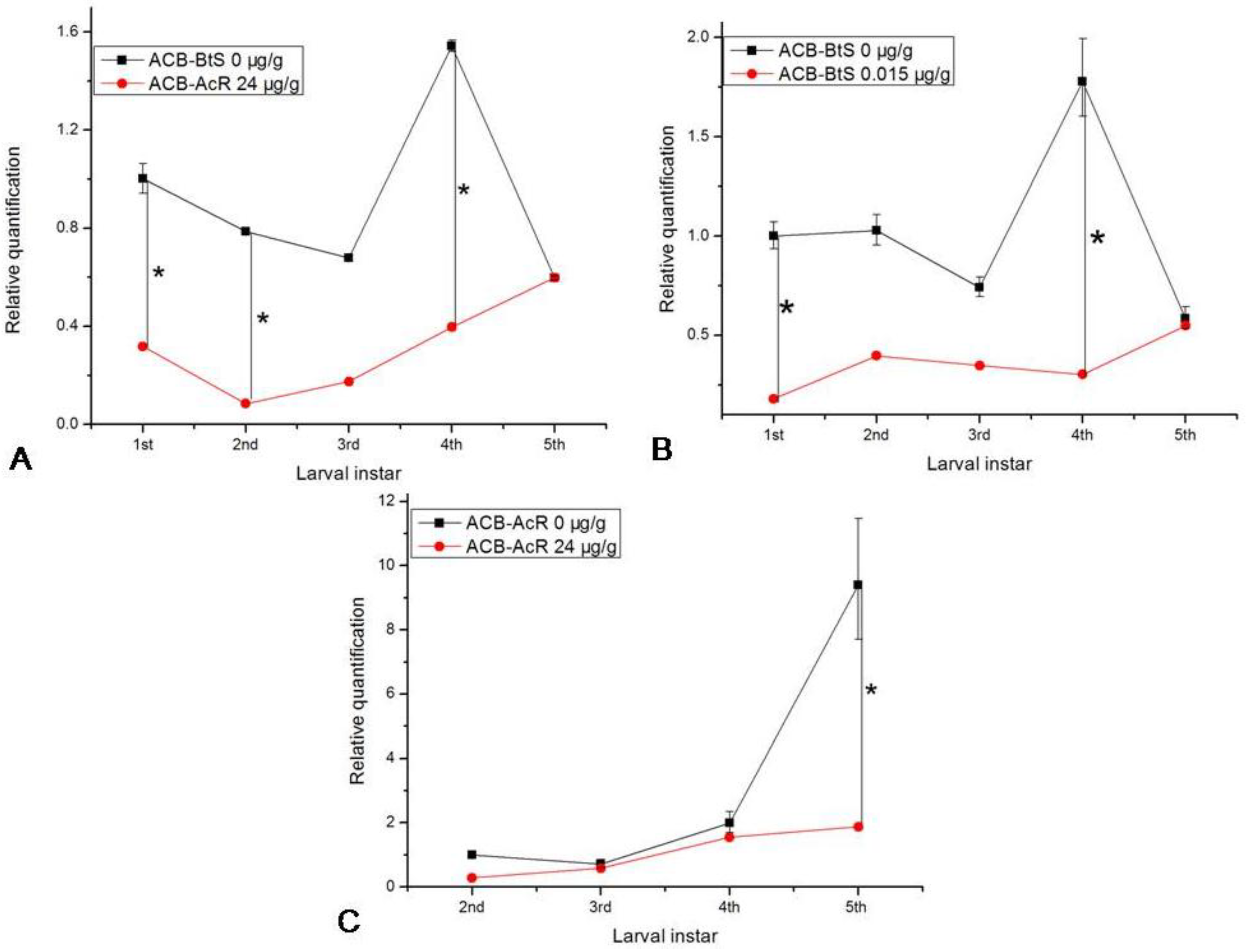
3. Discussion
4. Experimental Section
4.1. Insect Strains and Experimental Treatments
4.2. Cry1Ac Toxin
4.3. ACB Larval Midgut cDNA Synthesis
4.4. Cloning of a Full-Length cDNA Coding for Cadherin-Like Proteins in ACB
4.5. Expression and Purification of OfCAD Fragments
4.6. Ligand Blot Binding Assay
4.7. Variation of Membrane Proximal Extracellular Region (MPR) Screening in ACB-Ac200 Larvae
4.8. Binding Assay through Co-Immunoprecipitation (Co-IP)
4.9. Quantitative Real-Time PCR (qPCR)
5. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Crickmore, N.; Zeigler, D.R.; van Rie, J.; Lereclus, D.; Baum, J.; Bravo, A.; Dean, D.H. Bacillus Thuringiesis Toxin Nomenclature. Available online: http://www.lifesci.sussex.ac.uk/home/Neil_Crickmore/Bt/index.html (accessed on 2 April 2014).
- James, C. Global status of commercialized biotech/GM crops: 2010. ISAAA Briefs 2010, vii +, 280. [Google Scholar]
- Whalon, M.E.; Wingerd, B.A. Bt: Mode of action and use. Arch. Insect Biochem. 2003, 54, 200–211. [Google Scholar] [CrossRef]
- Bravo, A.; Gomez, I.; Conde, J.; Munoz-Garay, C.; Sanchez, J.; Miranda, R.; Zhuang, M.; Gill, S.S.; Soberon, M. Oligomerization triggers binding of a Bacillus thuringiensis Cry1Ab pore-forming toxin to aminopeptidase N receptor leading to insertion into membrane microdomains. Bba-Biomembranes 2004, 1667, 38–46. [Google Scholar]
- Gomez, I.; Sanchez, J.; Miranda, R.; Bravo, A.; Soberon, M. Cadherin-like receptor binding facilitates proteolytic cleavage of helix alpha-1 in domain I and oligomer pre-pore formation of Bacillus thuringiensis Cry1Ab toxin. Febs Lett. 2002, 513, 242–246. [Google Scholar] [CrossRef]
- Griffitts, J.S.; Aroian, R.V. Many roads to resistance: How invertebrates adapt to Bt toxins. Bioessays 2005, 27, 614–624. [Google Scholar] [CrossRef]
- Gasteiger, E.; Gattiker, A.; Hoogland, C.; Ivanyi, I.; Appel, R.D.; Bairoch, A. Expasy: The proteomics server for in-depth protein knowledge and analysis. Nucl. Acids Res. 2003, 31, 3784–3788. [Google Scholar] [CrossRef]
- Huang, F.N.; Higgins, R.A.; Buschman, L.L. Baseline susceptibility and changes in susceptibility to Bacillus thuringiensis subsp. kurstaki under selection pressure in European corn borer (Lepidoptera: Pyralidae). J. Econ. Entomol. 1997, 90, 1137–1143. [Google Scholar]
- Bolin, P.C.; Hutchison, W.D.; Andow, D.A. Long-term selection for resistance to Bacillus thuringiensis Cry1Ac endotoxin in a Minnesota population of European corn borer (Lepidoptera: Crambidae). J. Econ. Entomol. 1999, 92, 1021–1030. [Google Scholar]
- Chaufaux, J.; Seguin, M.; Swanson, J.J.; Bourguet, D.; Siegfried, B.D. Chronic exposure of the European corn borer (Lepidoptera: Crambidae) to Cry1Ab Bacillus thuringiensis toxin. J. Econ. Entomol. 2001, 94, 1564–1570. [Google Scholar] [CrossRef]
- Ferre, J.; van Rie, J. Biochemistry and genetics of insect resistance to Bacillus thuringiensis. Annu. Rev. Entomol. 2002, 47, 501–533. [Google Scholar] [CrossRef]
- Siqueira, H.A.A.; Moellenbeck, D.; Spencer, T.; Siegfried, B.D. Cross-resistance of crylab-selected Ostrinia nubilalis (Lepidoptera: Crambidae) to Bacillus thuringiensis delta-endotoxins. J. Econ. Entomol. 2004, 97, 1049–1057. [Google Scholar] [CrossRef]
- Pereira, E.J.G.; Lang, B.A.; Storer, N.P.; Siegfried, B.D. Selection for Cry1F resistance in the European corn borer and cross-resistance to other Cry toxins. Entomol. Exp. Appl. 2008, 126, 115–121. [Google Scholar] [CrossRef]
- Ali, M.I.; Luttrell, R.G.; Young, S.Y. Susceptibilities of Helicoverpa zea and Heliothis virescens (Lepidoptera: Noctuidae) populations to Cry1Ac insecticidal protein. J. Econ. Entomol. 2006, 99, 164–175. [Google Scholar] [CrossRef]
- Li, G.P.; Wu, K.M.; Gould, F.; Wang, J.K.; Miaoi, J.; Gao, X.W.; Guo, Y.Y. Increasing tolerance to Cry1Ac cotton from cotton bollworm, Helicoverpa armigera, was confirmed in Bt cotton farming area of China. Ecol. Entomol. 2007, 32, 366–375. [Google Scholar] [CrossRef]
- Van Rensburg, J.B.J. First report of field resistance by the stem borer, Busseola fusca (Fuller) to Bt-transgenic maize. S. Afr. J. Plant. Soil 2007, 24, 147–151. [Google Scholar]
- Tabashnik, B.E.; Gassmann, A.J.; Crowder, D.W.; Carriere, Y. Insect resistance to Bt crops: Evidence versus theory. Nat. Biotechnol. 2008, 26, 199–202. [Google Scholar] [CrossRef]
- Angst, B.D.; Marcozzi, C.; Magee, A.I. The cadherin superfamily: Diversity in form and function. J. Cell Sci 2001, 114, 629–641. [Google Scholar]
- Midboe, E.G.; Candas, M.; Bulla, L.A., Jr. Expression of a midgut-specific cadherin BT-R1 during the development of Manduca sexta larva. Comparative Biochem. Physiol. Part. B Biochem. Mol. Biol. 2003, 135, 125–137. [Google Scholar] [CrossRef]
- Vadlamudi, R.K.; Ji, T.H.; Bulla, L.A., Jr. A specific binding protein from Manduca sexta for the insecticidal toxin of Bacillus thuringiensis subsp. Berliner. J. Biol. Chem. 1993, 268, 12334–12340. [Google Scholar]
- Vadlamudi, R.K.; Weber, E.; Ji, I.; Ji, T.H.; Bulla, L.A., Jr. Cloning and expression of a receptor for an insecticidal toxin of Bacillus thuringiensis. J. Biol. Chem. 1995, 270, 5490–5494. [Google Scholar] [CrossRef]
- Griko, N.; Candas, M.; Zhang, X.; Junker, M.; Bulla, L.A., Jr. Selective antagonism to the cadherin BT-R1 interferes with calcium-induced adhesion of epithelial membrane vesicles. Biochemistry 2004, 43, 1393–1400. [Google Scholar] [CrossRef]
- Gahan, L.J.; Gould, F.; Heckel, D.G. Identification of a gene associated with BT resistance in Heliothis virescens. Science 2001, 293, 857–860. [Google Scholar] [CrossRef]
- Jurat-Fuentes, J.L.; Adang, M.J. The Heliothis virescens cadherin protein expressed in drosophila S2 cells functions as a receptor for Bacillus thuringiensis Cry1A but not Cry1Fa toxins. Biochemistry 2006, 45, 9688–9695. [Google Scholar]
- Morin, S.; Biggs, R.W.; Sisterson, M.S.; Shriver, L.; Ellers-Kirk, C.; Higginson, D.; Holley, D.; Gahan, L.J.; Heckel, D.G.; Carriere, Y.; et al. Three cadherin alleles associated with resistance to Bacillus thuringiensis in pink bollworm. Proc. Natl. Acad. Sci. USA 2003, 100, 5004–5009. [Google Scholar] [CrossRef]
- Nagamatsu, Y.; Toda, S.; Koike, T.; Miyoshi, Y.; Shigematsu, S.; Kogure, M. Cloning, sequencing, and expression of the Bombyx mori receptor for Bacillus thuringiensis insecticidal Cry1A(a) toxin. Biosci. Botechnol. Biochem. 1998, 62, 727–734. [Google Scholar] [CrossRef]
- Nagamatsu, Y.; Toda, S.; Yamaguchi, F.; Ogo, M.; Kogure, M.; Nakamura, M.; Shibata, Y.; Katsumoto, T. Identification of Bombyx mori midgut receptor for Bacillus thuringiensis insecticidal Cry1A(a) toxin. Biosci. Biotechnol. Biochem. 1998, 62, 718–726. [Google Scholar] [CrossRef]
- Wang, G.; Wu, K.; Liang, G.; Guo, Y. Gene cloning and expression of cadherin in midgut of Helicoverpa armigera and its Cry1A binding region. Sci. China Ser. C Life Sci./Chin. Acad. Sci. 2005, 48, 346–356. [Google Scholar]
- Flannagan, R.D.; Yu, C.G.; Mathis, J.P.; Meyer, T.E.; Shi, X.; Siqueira, H.A.; Siegfried, B.D. Identification, cloning and expression of a Cry1Ab cadherin receptor from European corn borer, Ostrinia nubilalis (Hübner) (Lepidoptera: Crambidae). Insect Biochem. Mol. Biol. 2005, 35, 33–40. [Google Scholar] [CrossRef]
- Chen, J.; Brown, M.R.; Hua, G.; Adang, M.J. Comparison of the localization of Bacillus thuringiensis Cry1A delta-endotoxins and their binding proteins in larval midgut of tobacco hornworm, Manduca sexta. Cell Tissue Res. 2005, 321, 123–129. [Google Scholar] [CrossRef]
- Hara, H.; Atsumi, S.; Yaoi, K.; Nakanishi, K.; Higurashi, S.; Miura, N.; Tabunoki, H.; Sato, R. A cadherin-like protein functions as a receptor for Bacillus thuringiensis Cry1Aa and Cry1Ac toxins on midgut epithelial cells of Bombyx mori larvae. FEBS Lett. 2003, 538, 29–34. [Google Scholar] [CrossRef]
- Braun, L.; Keddie, B.A. A new tissue technique for evaluating effects of Bacillus thuringiensis toxins on insect midgut epithelium. J. Invertebr. Pathol. 1997, 69, 92–104. [Google Scholar] [CrossRef]
- Nagamatsu, Y.; Koike, T.; Sasaki, K.; Yoshimoto, A.; Furukawa, Y. The cadherin-like protein is essential to specificity determination and cytotoxic action of the Bacillus thuringiensis insecticidal Cry1Aa toxin. FEBS Lett. 1999, 460, 385–390. [Google Scholar] [CrossRef]
- Dorsch, J.A.; Candas, M.; Griko, N.B.; Maaty, W.S.A.; Midboe, E.G.; Vadlamudi, R.K.; Bulla, L.A. Cry1A toxins of Bacillus thuringiensis bind specifically to a region adjacent to the membrane-proximal extracellular domain of BT-R(1) in Manduca sexta: Involvement of a cadherin in the entomopathogenicity of Bacillus thuringiensis. Insect Biochem. Mol. Biol. 2002, 32, 1025–1036. [Google Scholar] [CrossRef]
- Gomez, I.; Oltean, D.I.; Gill, S.S.; Bravo, A.; Soberon, M. Mapping the epitope in cadherin-like receptors involved in Bacillus thuringiensis Cry1A toxin interaction using phage display. J. Biol. Chem. 2001, 276, 28906–28912. [Google Scholar] [CrossRef]
- Fabrick, J.A.; Tabashnik, B.E. Binding of Bacillus thuringiensis toxin Cry1Ac to multiple sites of cadherin in pink bollworm. Insect Biochem. Mol. Biol. 2007, 37, 97–106. [Google Scholar] [CrossRef]
- Xu, X.; Yu, L.; Wu, Y. Disruption of a cadherin gene associated with resistance to Cry1Ac {delta}-endotoxin of Bacillus thuringiensis in Helicoverpa armigera. Appl. Environ. Microbiol. 2005, 71, 948–954. [Google Scholar] [CrossRef]
- Chen, J.; Hua, G.; Jurat-Fuentes, J.L.; Abdullah, M.A.; Adang, M.J. Synergism of Bacillus thuringiensis toxins by a fragment of a toxin-binding cadherin. Proc. Natl. Acad. Sci. USA 2007, 104, 13901–13906. [Google Scholar] [CrossRef]
- Hua, G.; Zhang, R.; Abdullah, M.A.; Adang, M.J. Anopheles gambiae cadherin agcad1 binds the Cry4Ba toxin of Bacillus thuringiensis israelensis and a fragment of agcad1 synergizes toxicity. Biochemistry 2008, 47, 5101–5110. [Google Scholar] [CrossRef]
- Park, Y.; Abdullah, M.A.; Taylor, M.D.; Rahman, K.; Adang, M.J. Enhancement of Bacillus thuringiensis Cry3Aa and Cry3Bb toxicities to coleopteran larvae by a toxin-binding fragment of an insect cadherin. Appl. Environ. Microbiol. 2009, 75, 3086–3092. [Google Scholar] [CrossRef]
- Peng, D.; Xu, X.; Ruan, L.; Yu, Z.; Sun, M. Enhancing Cry1Ac toxicity by expression of the Helicoverpa armigera cadherin fragment in Bacillus thuringiensis. Res. Microbiol. 2010, 161, 383–389. [Google Scholar] [CrossRef]
- Liu, C.; Wu, K.; Wu, Y.; Gao, Y.; Ning, C.; Oppert, B. Reduction of Bacillus thuringiensis Cry1Ac toxicity against Helicoverpa armigera by a soluble toxin-binding cadherin fragment. J. Insect Physiol. 2009, 55, 686–693. [Google Scholar] [CrossRef]
- Yang, Y.; Zhu, Y.C.; Ottea, J.; Husseneder, C.; Leonard, B.R.; Abel, C.; Luttrell, R.; Huang, F. Down regulation of a gene for cadherin, but not alkaline phosphatase, associated with Cry1Ab resistance in the sugarcane borer Diatraea saccharalis. PLoS One 2011, 6, e25783. [Google Scholar] [CrossRef]
- Wang, Z.Y.; Lu, X.; He, K.L.; Zhou, D.R. Review of history, present situation and prospect of the Asian maize borer research in China. J. Shenyang Agric. Univ. 2000, 31, 402–412. [Google Scholar]
- Zhou, D.R.; He, K.L.; Wang, Z.Y.; Ye, Z.H.; Wen, L.P.; Gao, Y.X.; Song, Y.Y. Asian Corn Borer and its Integrated Management; Golden Shield Press: Beijing, China, 1995. [Google Scholar]
- Wang, P.; He, K.L.; Wang, Z.Y.; Wang, Y.L. Evaluating transgenic Cry1Ac maize for resistance to Ostrinia furnacalis (gueneé). Acta Phytophylacica Sin. 2012, 39, 395–400. [Google Scholar]
- Li, N. Studies on the binding sites of Bacillus thuringiensis toxins with midgut and relationship between V-ATPase subunit A gene and Bt resistance of Ostrinia furnacalis. M.Sc. Thesis, Chinese Academy of Agricultural Sciences, Beijing, China, 2012. [Google Scholar]
- Han, H.L.; Li, G.T.; Wang, Z.Y.; Zhang, J.; He, K.L. Cross-resistance of Cry1Ac-selected Asian corn borer to other Bt toxins. Acta Phytophylacica Sin. 2009, 36, 329–334. [Google Scholar]
- ISREC ProfileScan Server. Available online: http://hits.isb-sib.ch/cgi-bin/PFSCAN (accessed on 20 May 2013).
- Tamura, K.; Peterson, D.; Peterson, N.; Stecher, G.; Nei, M.; Kumar, S. Mega5: Molecular evolutionary genetics analysis using maximum likelihood, evolutionary distance, and maximum parsimony methods. Mol. Biol. Evol. 2011, 28, 2731–2739. [Google Scholar] [CrossRef]
- Coates, B.S.; Sumerford, D.V.; Hellmich, R.L.; Lewis, L.C. Sequence variation in the cadherin gene of Ostrinia nubilalis: A tool for field monitoring. Insect Biochem. Mol. Biol. 2005, 35, 129–139. [Google Scholar] [CrossRef]
- Hua, G.; Jurat-Fuentes, J.L.; Adang, M.J. Bt-R1a extracellular cadherin repeat 12 mediates Bacillus thuringiensis Cry1Ab binding and cytotoxicity. J. Biol. Chem. 2004, 279, 28051–28056. [Google Scholar] [CrossRef]
- Xie, R.Y.; Zhuang, M.B.; Ross, L.S.; Gomez, I.; Oltean, D.I.; Bravo, A.; Soberon, M.; Gill, S.S. Single amino acid mutations in the cadherin receptor from Heliothis virescens affect its toxin binding ability to Cry1A toxins. J. Biol. Chem. 2005, 280, 8416–8425. [Google Scholar] [CrossRef]
- Storer, N.P.; Babcock, J.M.; Schlenz, M.; Meade, T.; Thompson, G.D.; Bing, J.W.; Huckaba, R.M. Discovery and characterization of field resistance to Bt maize: Spodoptera frugiperda (Lepidoptera: Noctuidae) in Puerto Rico. J. Econ. Entomol. 2010, 103, 1031–1038. [Google Scholar] [CrossRef]
- Yang, Y.J.; Chen, H.Y.; Wu, S.W.; Yang, Y.H.; Xu, X.J.; Wu, Y.D. Identification and molecular detection of a deletion mutation responsible for a truncated cadherin of Helicoverpa armigera. Insect Biochem. Mol. Biol. 2006, 36, 735–740. [Google Scholar] [CrossRef]
- Bel, Y.; Siqueira, H.A.A.; Siegfried, B.D.; Ferre, J.; Escriche, B. Variability in the cadherin gene in an Ostrinia nubilalis strain selected for Cry1Ab resistance. Insect Biochem. Mol. Biol. 2009, 39, 218–223. [Google Scholar] [CrossRef]
- Xu, X.J.; Wu, Y.D. Disruption of Ha_BtR alters binding of Bacillus thuringiensis delta-endotoxin Cry1Ac to midgut BMVs of Helicoverpa armigera. J. Invertebr. Pathol. 2008, 97, 27–32. [Google Scholar] [CrossRef]
- Yang, Y.J.; Chen, H.Y.; Wu, Y.D.; Yang, Y.H.; Wu, S.W. Mutated cadherin alleles from a field population of Helicoverpa armigera confer resistance to Bacillus thuringiensis toxin Cry1Ac. Appl. Environ. Microb. 2007, 73, 6939–6944. [Google Scholar] [CrossRef]
- Zhou, D.R.; Ye, Z.H.; Wang, Z.Y. Artificial rearing technique for Asian corn borer, Ostrinia Furnacalis (Guenée) and its applications in pest management research. In Advances in Insect Rearing for Research and Pest Management; Anderson, T.E., Leppla, N.C., Eds.; Westview Press: Boulder, CO, USA; San Francisco, CA, USA; Oxford, UK, 1992; pp. 173–193. [Google Scholar]
- SignalP Server. v 4.1. Available online: http://www.cbs.dtu.dk/services/SignalP/,v (accessed on 20 May 2013).
- ProtParam. Available online: http://web.expasy.org/protparam/ (accessed on 20 May 2013).
- NetNGlyc. v 1.0. Available online: http://www.cbs.dtu.dk/services/NetNGlyc/,v (accessed on 20 May 2013).
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2−ΔΔCT method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef]
© 2014 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Jin, T.; Chang, X.; Gatehouse, A.M.R.; Wang, Z.; Edwards, M.G.; He, K. Downregulation and Mutation of a Cadherin Gene Associated with Cry1Ac Resistance in the Asian Corn Borer, Ostrinia furnacalis (Guenée). Toxins 2014, 6, 2676-2693. https://doi.org/10.3390/toxins6092676
Jin T, Chang X, Gatehouse AMR, Wang Z, Edwards MG, He K. Downregulation and Mutation of a Cadherin Gene Associated with Cry1Ac Resistance in the Asian Corn Borer, Ostrinia furnacalis (Guenée). Toxins. 2014; 6(9):2676-2693. https://doi.org/10.3390/toxins6092676
Chicago/Turabian StyleJin, Tingting, Xue Chang, Angharad M. R. Gatehouse, Zhenying Wang, Martin G. Edwards, and Kanglai He. 2014. "Downregulation and Mutation of a Cadherin Gene Associated with Cry1Ac Resistance in the Asian Corn Borer, Ostrinia furnacalis (Guenée)" Toxins 6, no. 9: 2676-2693. https://doi.org/10.3390/toxins6092676
APA StyleJin, T., Chang, X., Gatehouse, A. M. R., Wang, Z., Edwards, M. G., & He, K. (2014). Downregulation and Mutation of a Cadherin Gene Associated with Cry1Ac Resistance in the Asian Corn Borer, Ostrinia furnacalis (Guenée). Toxins, 6(9), 2676-2693. https://doi.org/10.3390/toxins6092676