1. Introduction
Cyanobacteria, or blue-green algae, are microscopic organisms occurring in both freshwater and marine environments. Cyanobacteria are a significant part of the phytoplankton and the periphyton in lacustrine environments and among the largest groups of microorganisms found worldwide [
1]. Cyanobacteria can produce a wide variety of cyanotoxins including more than 80 known variations of microcystin heptapeptides and many lower molecular weight compounds [
2]. Cyanobacteria blooms may form in aquatic environments, where large populations form thick blue-green accumulations at the water surface and shoreline. Multiple types of cyanotoxins have been found in or associated with 48% of cyanobacteria with most reports (95%) describing the occurrence of hepatotoxic microcystins.
Cyanobacteria are considered a growing problem in ecosystems worldwide because of toxins that they produce [
3,
4]. Cyanotoxins or phycotoxins are organic compounds produced by cyanobacteria in the environment with biological effects on other organisms [
5,
6]. Early reports of cyanotoxin poisoning in animals (cattle, horses, and dogs) occurred in 1878 [
7]. Early reports of human poisoning from cyanotoxins in West Virginia, USA involved about 9000 people with gastroenteritis potential caused by drinking water contaminated with cyanotoxins [
8]. Algal toxins include several groups of hepatotoxins, such as microcystins, neurotoxins such as anatoxin
-a, and other compounds that can irritate the skin and gastrointestinal track such as lyngbyatoxin-
a [
4,
9] (
Table 1). Some cyanotoxins may accumulate in human and animals, causing a serious illness and poisoning [
5].
Table 1.
Some of the more important cyanotoxins and their effects on non-target organs.
Table 1.
Some of the more important cyanotoxins and their effects on non-target organs.
| Toxin | Non-target organ | Activity |
|---|
| Microcystins | Liver | Tumor Promoter |
| Nodularins | Liver | Carcinogenic |
| Cylinderospermopsin | Liver | Genotoxic |
| Anatoxin-a | Nervous System | Depolarizing neuromuscular blockers |
| Anatoxin-a (s) | Nervous System | Inhibits AchE |
| Saxitoxin | Nervous System | Na+ channel blocker |
| BMAA | Nervous System | Neurodegeneration |
| Lyngbyatoxin-a | Skin | Inflammatory agent |
| Aplysiatoxins | Skin | Inflammatory agent |
| Lipopolysaccharide | G.I.T. | Gastrointestinal irritant |
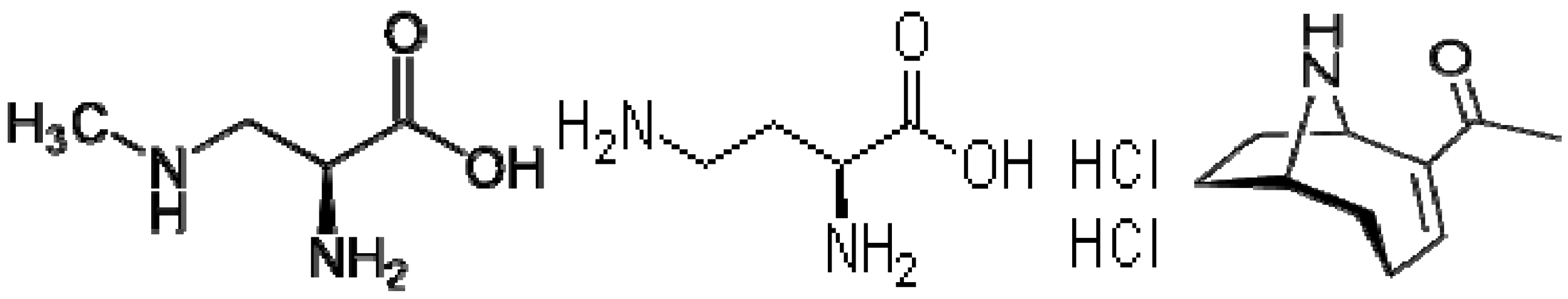
Several lower molecular weight cyanotoxins are of particular interest because they are studied less often, and have known or suspected neurotoxic effects. For example, β-
N-methylamino-
l-alanine (BMAA) (CAS number 15920-93-1) is a non-protein amino acid with the chemical name β-
N-methylamino-
l-alanine, molecular formula (C
4H
10N
2O
2) (
Figure 1) and a relatively low molecular weight of 118.13 g/mol. It has been suggested that cellular exposure to BMAA may lead to neurologic damage in the brain and central nervous system of humans and animals, potentially contributing to one of several neurodegenerative diseases [
10].
Figure 1.
Structures of β-N-methylamino-l-alanine (BMAA), 2,4-diaminobutyric acid dihydrochloride (DABA) and anatoxin-a.
Figure 1.
Structures of β-N-methylamino-l-alanine (BMAA), 2,4-diaminobutyric acid dihydrochloride (DABA) and anatoxin-a.
Polsky
et al. [
10] was the first to show BMAA neurotoxicity in extracts from cycad seeds used widely in Guam as a food and medicinal plant. Also, Karamyan and Speth [
11] mentioned that BMAA can be incorporated into plant and animal proteins. BMAA has subsequently been detected in cyanobacterial blooms and laboratory isolates from marine and freshwater sources from localities worldwide including: Iraq, Qatar, Hawaii, China, United Kingdom, South Africa, Netherlands, and Sweden [
12,
13,
14,
15,
16]. As a result, it has been hypothesized that BMAA can occur and bioaccumulate in other aquatic ecosystems where cyanobacteria are known to proliferate.
Studies have shown that BMAA occurs in zooplankton which can feed on cyanobacteria, and even higher BMAA levels were found in fish likely to feed on zooplankton [
17,
18]. Bioaccumulation of BMAA in aquatic organisms has been suggested in marine environments [
19,
20]. A particularly remarkable finding was the discovery of high BMAA levels in bottom-dwelling fish species (
S. maximus,
T. quadricornis, and
O. eperlanus) and in water filter-feeding mollusks (
M. edulis and
O. edulis) [
17]. BMAA was also detected in three aquatic animals’ species (zebra fish, brine shrimp, and the protozoan
Nassula sorex) providing additional proof that BMAA can bioaccumulate in food chains, leading to potential human exposure [
21]. BMAA was reported with a concentration as high as 7000 ng/g in South Florida samples that included water, cyanobacteria, fish (such as perch), and seafood (such as crab and shrimp) that are consumed by humans [
22]. Detectable levels of BMAA was recently reported in shark fins, which have been used widely in the well-known “shark fin soup” in Florida [
18].
Vranova
et al. (2011) [
23] showed that non-protein amino acids (NPAAs) play a significant role in plant, soil, and ecosystems functions. NPAAs are considered a store of organic nitrogen in many ecosystems, and are thought to be a toxins produced against both invertebrate and vertebrate animals, which could lead to serious human medical concerns. BMAA and DABA are NPAA toxins produced by cyanobacteria [
24]. BMAA (β-
N-methylamino-
l-alanine) and DABA (2,4-diaminobutyric) are non-protein amino acids found in both marine and fresh waters [
25]. Over the past 150 years, algal blooms, including those that are toxic, have caused problems worldwide, and now due to global warming these problems may accelerate [
26].
Metcalf
et al. [
27] detected BMAA in British waterbodies along with other cyanotoxins such as microcystin, anatoxin-
a, nodularin, and saxitoxin suggesting a health risk assessment of cyanobacterial BMAA in waterbodies. Cyanotoxins interfere with zebra fish (Danio rerio) embryo growth in an animal model system [
28]. Salierno
et al. [
29] found that fish exposed to algal neurotoxins such as domoic acid, brevetoxin, and saxitoxin, suffered from severe loss of optic region activity in the brain. Microcystin are known to accumulate in liver, muscle, and viscera of fish, affecting their growth and survival rates [
30].
DABA (
Figure 1) is also a non-protein amino acid and BMAA-isomer. DABA is chemically known as 2,4-diaminobutyric acid, with the molecular formula C
4H
10N
2O (
Figure 1), CAS Number 1883-09-6, and molecular weight of 191.06 g/mol. DABA is thought to have the same toxicological character of BMAA but has not been extensively studied [
31].
Anatoxin-
a is an alkaloid compound with potent postsynaptic and depolarizing neuromuscular blockers [
32]. The molecular formula is (C
10H
15NO) (
Figure 1), CAS number 64285-06-9, and the molecular weight is 165.232 g/mol. Clinical signs of anatoxin-
a exposure and neurotoxicity include muscular fasciculation, imbalance, and respiratory failure due to paralysis, leading to death [
33,
34,
35]. Anatoxin-
a was found to be produced by cyanobacteria isolated from Lake Biwa in Japan, along with homoanatoxin-
a [
36]. Anatoxin-
a has been detected in Wisconsin and Florida waters and algal blooms with other cyanotoxins, such as cylindrospermopsins and microcystin-LR [
33,
37]. Anatoxin-
a was also reported in cyanobacteria isolated from freshwater systems in Portugal, leading to suspicions of the growing health and ecological risk of its occurrence in fresh waters [
38].
BMAA and DABA have different mechanisms of action in comparison to anatoxin-
a. Anatoxin
-a is a cyanotoxin with acute neurotoxicity. Anatoxin-
a is produced by at least four genera of cyanobacteria and has been reported from North America, Europe, Africa, Asia, and New Zealand [
39,
40]. BMAA is produced by almost all cyanobacteria, such as the genera
Anabaena and
Nostoc which can live symbiotically in plant roots. The BMAA and DABA suggested mechanism of toxicity is though destruction of motor neurons in the brain, leading to neurodegenerative diseases likes Parkinson’s disease, Alzheimer’s, and ALS (Lou Gehrig’s disease) [
13,
20,
41,
42]. It was well explained and discussed how BMAA could be related to these neurodegenerative diseases by Papapetropoulos [
42,
43]. In one well-documented study, members of the Chamorro people on Guam who consumed the cycad seeds were found to suffer from ALS or Parkinsonism-dementia complex (PDC), which ultimately killed approximately 10% of Guam’s indigenous Chamorro population [
42]. The traditional cuisine of the Chamorro people has also included flying fox bats, which feed on cycad flowers and fruits. Thus, it has been suggested that, “the plant and animal proteins provide unrecognized reservoirs for the slow release of this toxin” BMAA were detected to be bioaccumulated in cycads seeds and flying fox bats as produced by cyanobacteria that live endosymbiosis in the roots of tree cycads in Guam [
20,
24,
44,
45,
46,
47,
48].
BMAA was also recently reported to occur in the brain tissue of nine Canadian’s Alzheimer patients [
49]. Alternative ecological pathways likely occurs leading to bioaccumulation of BMAA in aquatic and terrestrial ecosystems [
41,
47,
50]. Murch
et al. [
47] explained the mechanism of the slow release of biomagnified blue-green algal neurotoxins and neurodegenerative disease in Guam by suggesting that the BMAA neurotoxin produced by
Nostoc can be concentrated in cycads seeds. Grinding the seeds for flour, the BMAA was concentrated over time in brain tissue causing death to Chamorro people who suffered from ALS. Cox
et al. [
41] showed that the algal neurotoxin BMAA can be produced by all known groups of cyanobacteria in both symbiotic and free-living forms. Their data showed that
Nostoc can exist as symbionts with host plants such as cycads, some flowering plants such as
Gunnera, and other cyanobacteria such as
Synechocystis and
Anabaena, found in U.S. fresh waters.
Nebraska Lakes and Impoundments
Cyanotoxins have also been show to co-occur with drinking water taste-and-odor compounds, such as geosmin and 2-MIB in 91% of blooms sampled in a regional study of Midwestern lakes and impoundments [
51]. In Nebraska, algal toxin problems have become a more growing health concern with reported cases of pet, livestock and wildlife deaths, cases of GIT illness, and cutaneous rashes from contaminated lakes and ponds [
52]. In all these cases, acute exposure to toxins of cyanobacteria is suspected to be the primary cause.
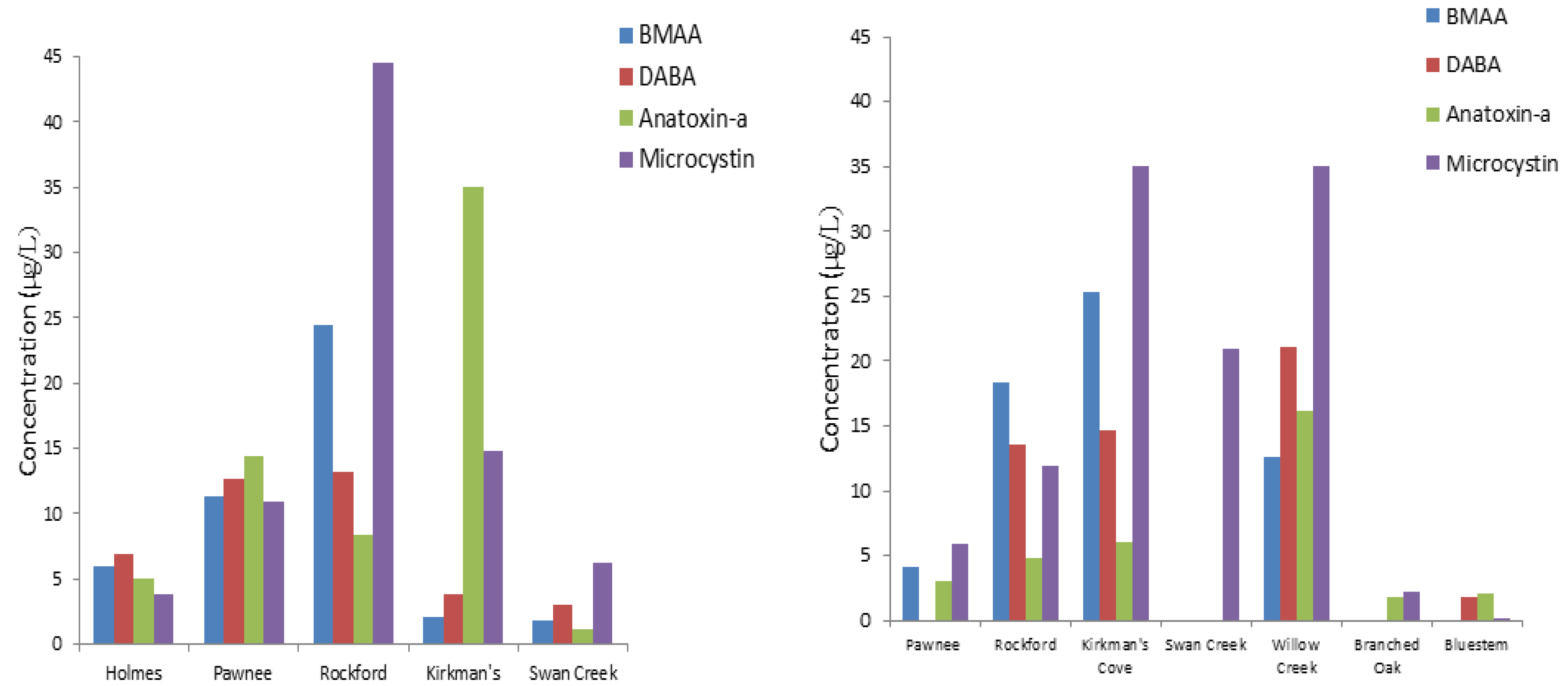
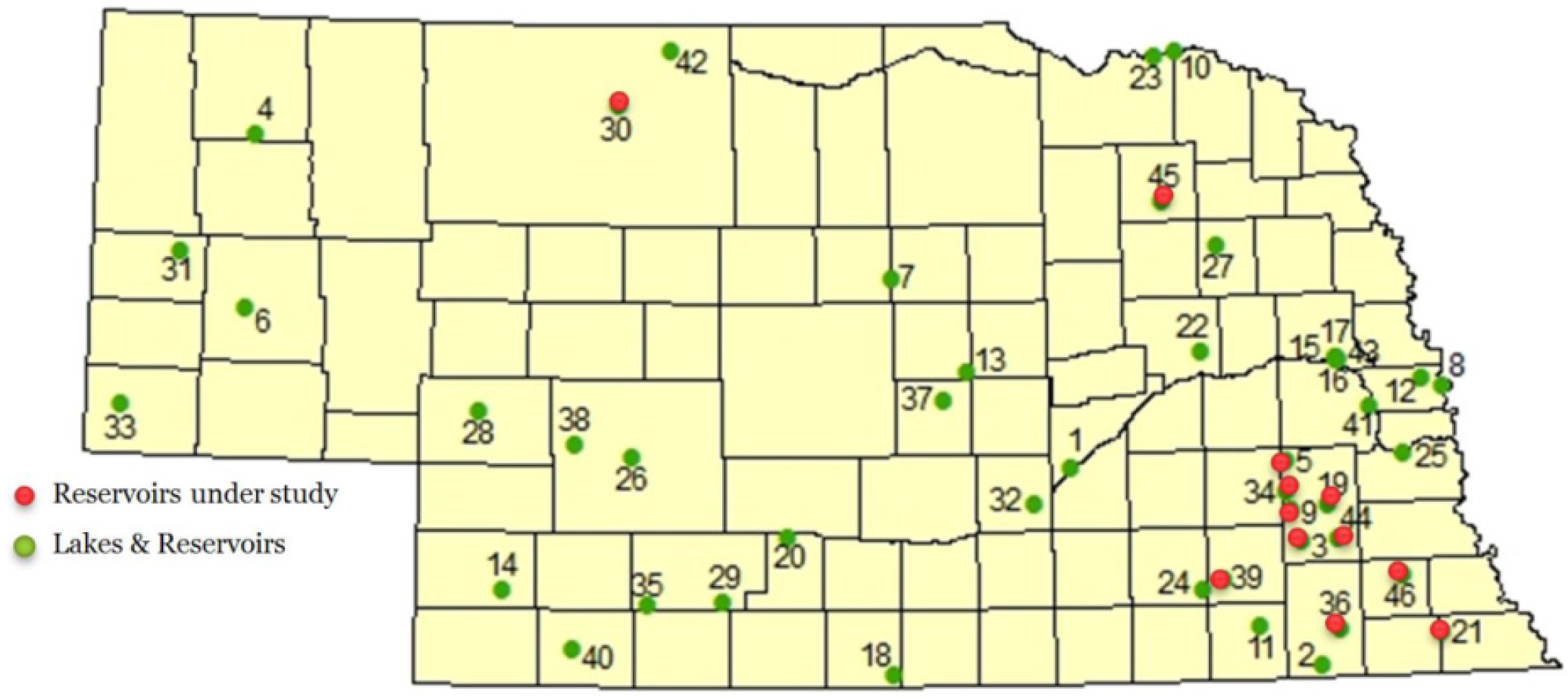
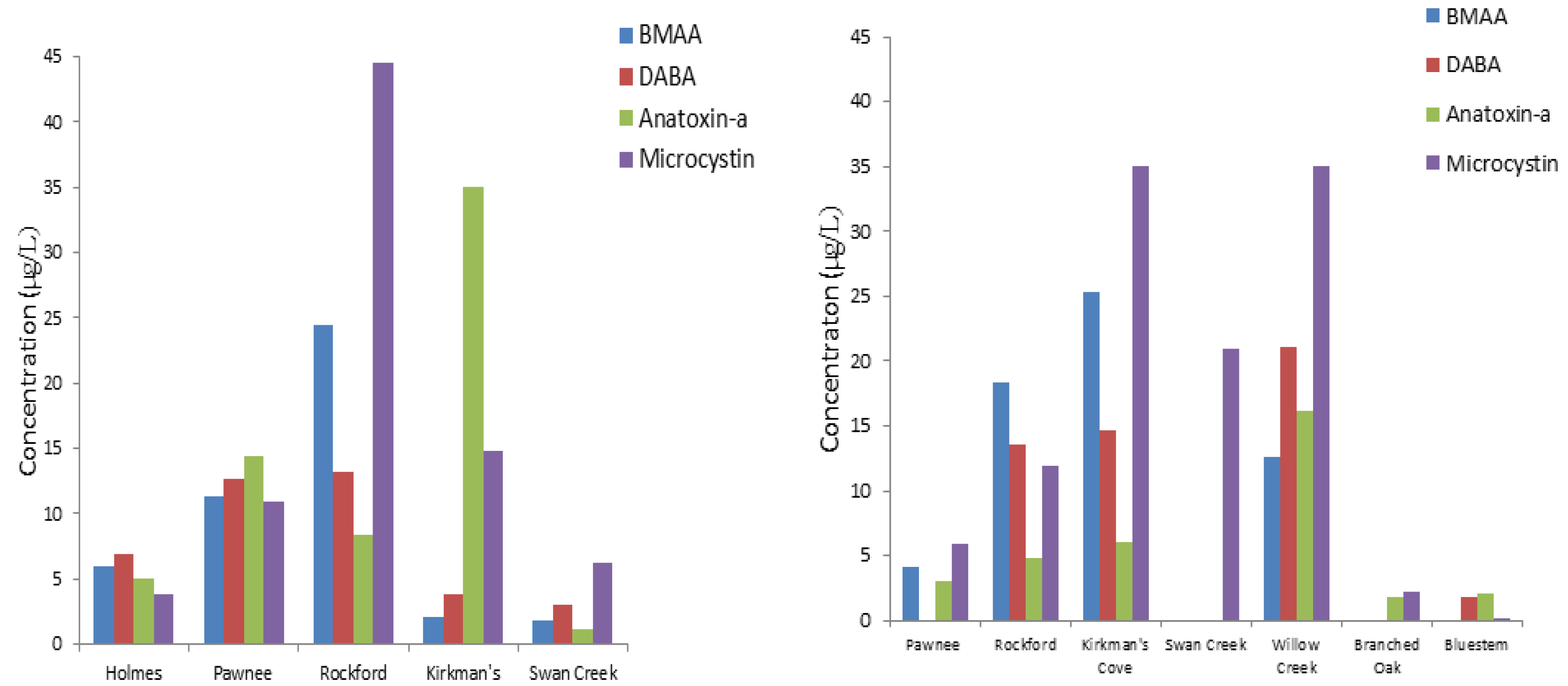
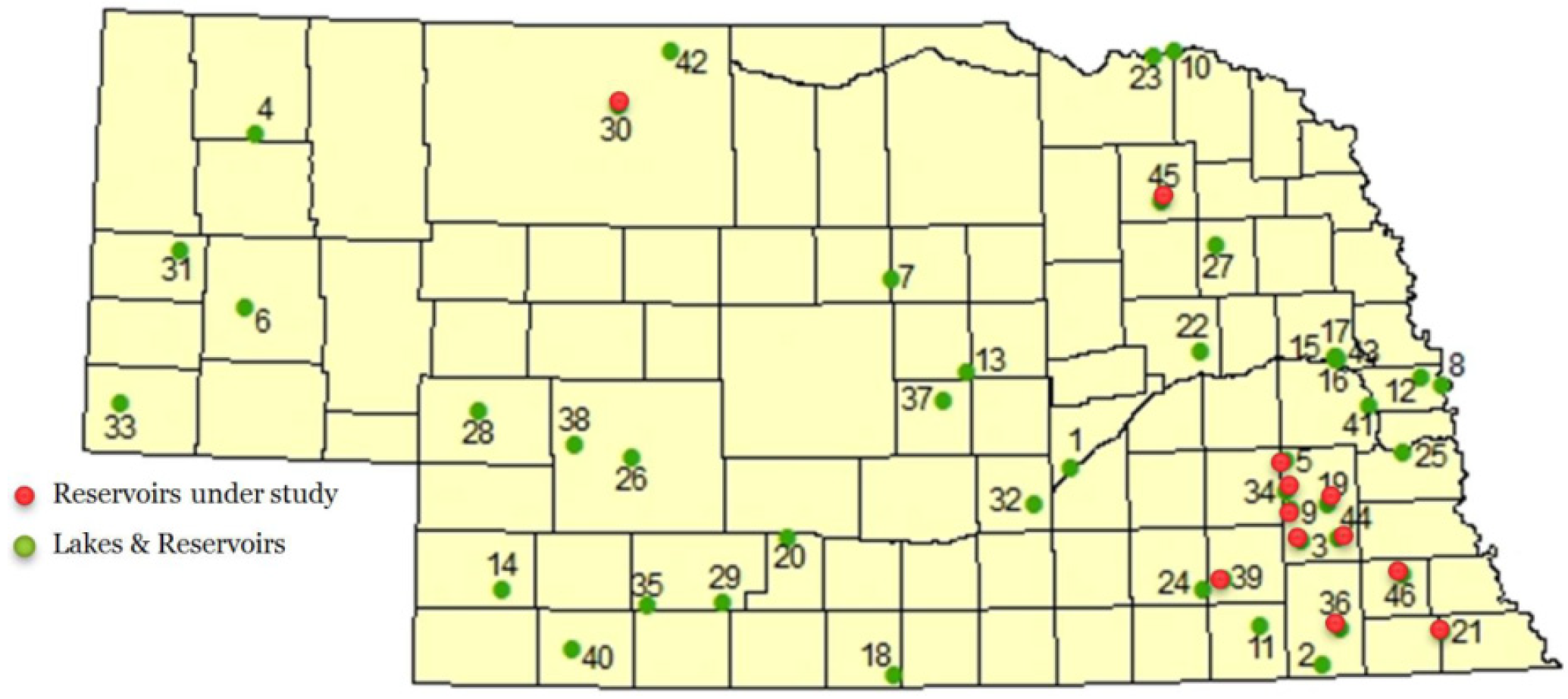
Nebraska lakes and reservoirs appear to be particularly prone to cyanobacteria blooms. Beginning in 2004, cyanotoxins were regularly monitored by the Nebraska Department of Environmental Quality. Several lakes included in this monitoring program are annually reported to be a health alert issue in Nebraska [
53]. Pawnee Reservoir #34, Fremont Lake #20, Carter Lake, Swan Creek #5A, Willow Creek Reservoir #46, and others have exceeded 20 µg/L of microcystin. WHO health advisory concentration of microcystins as determined by immunoassay [
54]. A total of about 700 water samples were collected for monitoring algal toxin levels from 34 public lakes and reservoirs between 2004 and 2010. The number of lakes monitored has increased since then [
53,
54]. According to the Nebraska Water Monitoring program report for 2010, lakes/reservoirs within Nebraska are under weekly monitoring from May 1 to September 30 each year. Weekly monitoring usually includes samples of fish and water for bacteria, and for algal toxins, namely microcystins [
54]. Health alerts are triggered in public lakes and reservoirs by NDEQ when the microcystin level reaches 20 µg/L, as measured by immunoassay. In 2004, health alerts were issued for 17 lakes/reservoirs (50% of lake samples), 12 lakes in 2005 (35%), six lakes in 2006 (12%), and six lakes in 2007 (13%) [
53,
54].
The present study was conducted to evaluate the potential for occurrence of neurotoxic cyanotoxins in Nebraska lakes and reservoirs, as well as associated plant and fish tissue collected during two summers, when cyanobacteria blooms are prevalent. The study employed a new detection method using derivatization combined with chromatography and fluorescence or mass spectrometry to simultaneously detect groups of cyanotoxins in biological and environmental samples [
55]. Few wide-spread monitoring studies of neurotoxic cyanotoxins have been conducted because of the difficulties in measuring low molecular weight and water soluble organics at trace levels in complex environmental matrices. The present study provides a snap shot of the occurrence of these compounds in lake water and in aquatic organisms likely to serve as a route of human exposure.
4. Conclusions
The cyanotoxins BMAA, DABA, and anatoxin-
a are reported for multiple Midwestern impoundments. Ninety-four environmental samples with various levels of BMAA, DABA, and anatoxin-a, out of 378 samples, were collected. Water samples showed 31 positives out of 67, fish samples showed 33 positives out of 248, and aquatic plants (water milfoil) displayed 30 positive samples out of 72 collected samples (
Table 7). Summer is the main time of the year that cyanobacteria produce/release cyanotoxins into the environment. The occurrence of cyanotoxins in lake water, aquatic plants and fish suggests potential for transfer throughout the ecosystem to higher levels in the food web. As cyanotoxins may reach higher organisms such as humans through these additional pathways, there is the potential for damage leading to neurodegenerative diseases such as ALS, PD, and AD. Thus, it is important to recognize these potential routes of exposure; directly by drinking or swimming in water or indirectly by eating contaminated fish.
Table 7.
Summary of samples with positives and undetected (ND) levels of cyanotoxins from a total of 387 samples collected between 2009 and 2010.
Table 7.
Summary of samples with positives and undetected (ND) levels of cyanotoxins from a total of 387 samples collected between 2009 and 2010.
| Samples | 2009 | 2010 | Total positive (% of total) | ND |
|---|
| Reservoir water | 13 | 18 | 31 (46.3) | 36 |
| Fish | 24 | 9 | 33 (13.3) | 215 |
| Aquatic plant | 15 | 15 | 30(41.7) | 42 |
| Total samples collected in 2009–2010 = 387 | 94 | 293 |
Of the 12 Nebraska reservoirs sampled in this study, two reservoirs did not have detectable levels of cyanotoxins and served as a control group in the study. This is consistent with previous monitoring for cyanotoxins in Nebraska. BMAA, DABA, and anatoxin-
a were detected in Nebraska reservoirs in 2009–2010 samples, including fish, aquatic plant, and lake water. BMAA and DABA were measured in 13 reservoir water samples in 2009 and 12 samples in 2010, while anatoxin-
a was measured in 13 samples in 2009 and 18 samples in 2010. BMAA was found in 22 fish tissue samples in 2009 and nine samples in 2010, while DABA was detected in 19 samples in 2009 and seven samples in 2010. Anatoxin-
a was not detected in any fish samples. BMAA, DABA, and anatoxin-
a were detected in 15 samples of aquatic plants collected in 2009 and 15 in 2010 (
Table 7 and
Table 8).
Table 8.
Cyanotoxins detection summary for Nebraska reservoir samples from 2009 and 2010.
Table 8.
Cyanotoxins detection summary for Nebraska reservoir samples from 2009 and 2010.
| Cyanotoxins | Water | Fish | Plant | Percent of detections |
|---|
| 2009 | 2010 | 2009 | 2010 | 2009 | 2010 | 2009–2010 |
|---|
| BMAA | 13 | 12 | 22 | 9 | 5 | 5 | 18% |
| DABA | 13 | 12 | 19 | 7 | 5 | 5 | 17% |
| Anatoxin-a | 13 | 18 | 0 | 0 | 5 | 5 | 12% |
| Total samples | 31 | 36 | 115 | 133 | 36 | 36 | 387 |
The results suggest that detectable reservoir water levels of BMAA, DABA, and anatoxin-a may be associated with a detectable levels of cyanotoxins in fish tissue and aquatic plants. We can also hypothesize that the presence of such cyanotoxins in Nebraska reservoir water will transfer in the food web with an increased potential for bioaccumulation. Because of variable occurrence of DABA and anatoxin-a, these cyanotoxins may not be produced by the same cyanobacteria species. BMAA concentrations were more clearly associated with detection of microcystins.




{kind=link}
{kind=link}
{kind=link}
{kind=link}