Abstract
Natural killer (NK) cells exert important immunoregulatory functions by releasing several inflammatory molecules, such as IFN-γ and members of chemokines, which include CCL3/MIP-1α and CCL4/MIP-1β. These cells also express heptahelical receptors, which are coupled to heterotrimeric G proteins that guide them into inflamed and injured tissues. NK cells have been shown to recognize and destroy transformed cells and virally-infected cells, but their roles in neurodegenerative diseases have not been examined in detail. In this review, I will summarize the effects of NK cells in two neurodegenerative diseases, namely multiple sclerosis and globoid cell leukodystrophy. It is hoped that the knowledge obtained from these diseases may facilitate building rational protocols for treating these and other neurodegenerative or autoimmune diseases using NK cells and drugs that activate them as therapeutic tools.
1. Introduction
The immune system is composed of a network of cells and proteins that act in concert to defend the body against diseases. There is increasing evidence that key components of the innate immune system, such as natural killer (NK) cells, are instrumental in disease control or progression. These cells have both cytolytic and immunoregulatory functions [1] and play vital roles by surveilling the microenvironment searching for abnormal growth or microbial-infected cells [2]. NK cells recognize and kill these latter cells by a set of receptors known as NK cell cytotoxicity receptors [3]. Therefore, NK cells are important in protection, as well as therapy against cancer [4,5], in lysing virally-infected cells [6] and are implicated in therapy against HIV-infection [7]. More than 90% of NK cells are found in the blood circulation, whereas less than 10% are found in tissues, such as spleen, liver and lungs [8]. NK cells are not circulatory [9]; however, under pathological conditions and during inflammation, they extravasate into the lymph nodes and accumulate at sites of tumor growth [10]. In addition, NK cells with phenotypes and functional activities similar to activated NK cells can be found in the liver of normal rats [11]. These observations suggest that NK cells found in secondary lymphoid tissues or in non-lymphoid tissues have been stimulated in situ before their extravasation into these areas. It is not clear why the majority of NK cells are found in the circulation and what influence their accumulation in peripheral tissues. The majority of peripheral blood resting NK cells (about 80%–90%) do not express the CD56 molecule, and are designated as CD56dim/− cells. These cells have high cytolytic activity and low immunoregulatory capacity, whereas the other 10%–20%, designated as CD56bright/+ cells, have high immunoregulatory function and low cytolytic activity [8,12]. In addition to their role in cancer [4,5], NK cells are involved in autoimmune diseases, such as multiple sclerosis [13], systemic lupus erythematosus [14] and rheumatoid arthritis [15]. The model described by others [16], is perhaps doubtful to have any significant impact for human diseases, and will not be further discussed.
1.1. Natural Killer (NK) Cells Express G Protein-Coupled Receptors
NK cells express receptors, which bind members of the heptahelical receptors (also known as G protein-coupled receptors, “GPRC”, or seven transmembrane spanning domain receptors). Both CD56bright/+ and CD56dim/− NK cells express the sphingosine 1-phosphate receptors S1P1, S1P4 and S1P5 [17]. In addition, both subsets express LPA1, LPA2 and LPA2, the receptors for lysophosphatidic acid [18]. The expression of lysolipid receptors on the surface of these cells may facilitate their migration into various tissues, since lysophospholipids are secreted by tumor and inflammatory cells [19]. Although NK cells have been implicated in several neurodegenerative diseases, such as Alzheimer’s disease [20,21], this review will focus on two neurodegenerative diseases, where the author developed experience with, namely multiple sclerosis and globoid cell leukodystrophy.
1.2. NK Cells Contain Different Subsets
Previously, human NK cells have been classified based on the expression of the CD56 molecule, but other findings suggest that NK cells might be classified into NK1 cells producing Th1-like cytokines, such as IFN-γ, and NK2 cells producing Th2-like cytokines, such as IL-5 and IL-13 [22]. Human NK cells are also divided into cells that express the chemokine receptors, CXCR1, CXCR3 and CXCR4, while others express CCR1, CCR4, CCR5, CCR6, CCR7, CCR9, CXCR5 and CXCR6 within both CD56bright and CD56dim subsets [23]. Another set of NK cells isolated from human peyers patches or tonsils expressing NKp44 and CCR6 molecules and secreting IL-22 and CCL20/MIP-3α/LARC was discovered and the cells were designated as NK22 [24]. Yet, other cells isolated from intestinal lamina propria, expressing NKp46+ NKG2D+ NK1.1int RORγhigh and secreting IL-22, have been described [25]. Human NK cells in stage III development express CD34+CD117+2B4+ that produced IL-22 and IL-26 but not IL-17, have been isolated from tonsil tissues [26]. Further work from our group described a new subset of human NK cells isolated from the peripheral blood and activated in vitro with IL-2. The cells secrete IL-17 and IFN-γ and were designated as NK17/NK1 cells [27]. Figure 1 shows the phenotypic expression of NK22 cells and NK17/NK1 cells.
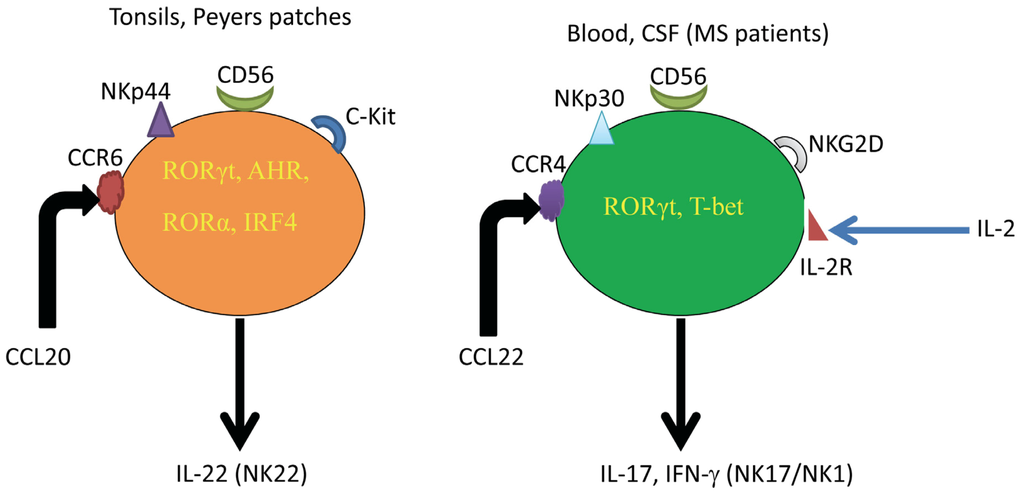
Figure 1.
Comparison among NK22 cells and NK17/NK1 cells. While NK22 cells are found in tissues, such as tonsils and peyers patches, NK17/NK1 cells are generated from human blood after activation in vitro with IL-2 and are abundant in the CSF of MS patients without activation. NK22 cells express the NK cytotoxicity receptor NKp44, whereas NK17/NK1 cells express NKp30, NKG2D, as well as NKp44, NKp46 and CD158, but to a lower extent than the other NK cell cytotoxicity receptors. Whereas NK22 cells express CCR6, secrete and respond CCL20/MIP-3α, NK17/NK1 cells express CCR4, secrete and respond to CCL22/MDC. As for the transcription factors, NK22 cells express RORγ, AHR, RORα and IRF4, while NK17/NK1 cells express RORγ and T-bet and, consequently, secrete both IL-17 and IFN-γ.
Although it is well established that IFN-γ is an inflammatory cytokine, the function of IL-17 in autoimmune diseases has not been clarified. IL-17 is considered an inflammatory molecule, because it induces the production of IL-6, TNF-α or CXCL8/IL-8 [28], stimulates the production of matrix metalloproteinases-2, 3, 9 and 13 [29], and facilitates the proliferation of endothelial cells [30], but it may relieve inflammatory colitis disease [31]. In addition, it was recently reported that this cytokine may be beneficial in myocardial infarction, due to its ability to inhibit the chemotaxis of monocytes in this disease [32]. We also reported that IL-17 tends to reduce the expression of two major chemokine receptors involved in monocyte migration into inflamed hearts, namely CCR2 and CXCR4 [32].
2. Role of NK Cells in Multiple Sclerosis (MS)
Multiple sclerosis (MS) is an inflammatory disorder that results in demyelination and destructions of neurons. The animal experimental model of MS, also known as experimental autoimmune encephalomyelitis (EAE), is an excellent model, which describes the role that autoreactive CD4+ cells play in this disease [33,34]. A hallmark of the immune response in MS is the formation of isolated areas of inflammation called MS lesions. Lesions can appear both in the white matter and in the grey matter of the brain and are often found around the ventricles, in the optic nerve, in the brain stem and in the spinal cord [35].
The role of NK cells in MS is controversial, with one school of thought suggests that NK cells ameliorate the disease, whereas another school indicates that they exacerbate the disease [36,37]. The first indicates that NK cells ameliorate EAE by lysing cells that aggravate the MS disease [38]. Consequently, depleting NK cells before immunization of sensitive mice with myelin oligodendrocyte glycoprotein (MOG35–55) peptide leads to a severe relapsing EAE, which is the result of increased T-cell proliferation and production of Th1 cytokines. NK cells, by suppressing pathogenic autoreactive T-cells, which mediate the central nervous system (CNS) inflammation, may repair the demyelination [39]. Impaired NK cell recruitment into CX3CR1-deficient mice suffering from EAE also leads to severe disease [40]. This is because recruitment of NK cells by CX3CR1/Fractalkine axis into the CNS of EAE mice inhibits the inflammatory Th17 cells [41], and hence, lack of NK cells may result in enhancing the severity of the disease.
In contrast, others suggested that NK cells exacerbate MS/EAE. For example, higher NK cell activity is correlated with a higher risk of developing active lesions in relapsing-remitting MS patients [42]. Also, IL-12 produced by astrocytes promotes NK cell developments that secrete cytokines, which enhance T-cell activation [43]. Further, IL-18 produced during the primary injection of antigens increased IFN-γ secretion by NK cells. IFN-γ in turn activates autoreactive Th1 responses, whereas an impaired capacity of NK cells to release IFN-γ is a major mechanism underlying resistance to EAE [44]. In these studies, it was observed that depleting NK cells with specific antibodies leads to diminished EAE clinical disease [45].
We examined the role of NK cells in both EAE experimental system and MS patients. Our approach was to investigate whether drugs used to treat MS patients may exert effects on NK cell biological activities. One of the drugs we used is glatiramer acetate (GA; commercial name Copaxone), which is made up of four amino acids, namely Glu, Ala, Lys and Tyr, that are found in myelin. This drug prevents the incidence of experimental autoimmune encephalomyelitis in animals and reduces relapses in patients with MS [46]. MS patients receiving GA had a steady decline in disease relapses with neurological improvement when compared to patients receiving placebo [47,48]. We recently described the activities of NK cells and dendritic cells (DCs) isolated from nine relapsing remitting MS patients. Our results demonstrate that NK cells generated from the peripheral blood of these patients have increased cytolytic activity against tumor target cells, autologous immature (i) and mature (m) DCs [49]. NK cells are known to interact with DCs leading to DCs maturation or death and/or NK cells activation [50]. It was also suggested that the interaction between NK cells and DCs is bidirectional and involves cell-to-cell contact, where DCs activate NK cells by enhancing their proliferation, cytotoxic activity and IFN-γ production, whereas activated NK cells provide either maturation signals for DCs or induce their death by direct killing (Figure 2).
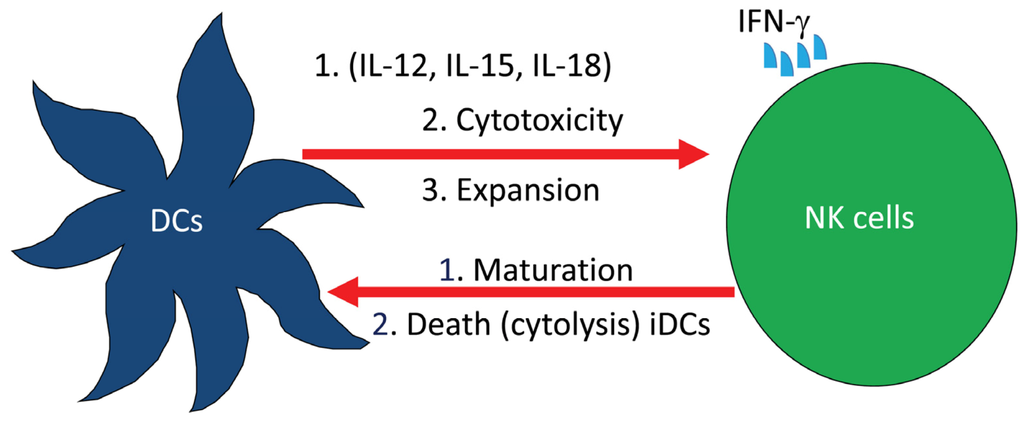
Figure 2.
Complex interactions among cells of the innate immune system. Dendritic cells (DCs) releasing IL-12, IL-15 and IL-18 activate natural killer (NK) cell cytotoxicity, expansion and the release of IFN-γ. In turn, NK cells induce either the maturation or death of DCs.
It is not yet known where in the body NK cells interact with DCs, however, there is a consensus suggesting that this interaction might take place at inflammatory sites [50]. Our results showing that NK cells lyse DCs after treatment may provide a mechanistic explanation for such an interaction, leading to a vitally important sequence of events, which culminate in benefiting MS patients. In addition, the observations provide a novel mechanism of action for drugs used to treat autoimmune diseases, showing that these drugs ablate DCs by activating NK cells, which lyse them.
In vitro work also revealed that GA enhances the cytolysis of activated human NK cells against immature and mature monocyte-derived DCs [51]. These findings were supported in mice with EAE, where administration of GA ameliorated the EAE clinical scores, corroborated with the ability of NK cells to kill both immature and mature DCs [52]. Collectively, these findings demonstrate that GA inhibition of EAE clinical disease may be due to its ability to activate NK cells to lyse DCs, among other functions. The finding that NK cells exposed to GA kill both immature DCs and mature DCs leads to the inability of presenting antigens to autoreactive T-cells. Finally, we suggest that the newly described NK17/NK1 cells may be of interest in MS disease. We are currently evaluating the effects of various drugs used to treat MS patients on the biological activities of these cells. Also, we are examining whether these cells may secrete anti-inflammatory cytokines, such as IL-10 among others, notwithstanding the perplexing and highly questionable results, showing that IL-10 is a culprit in another autoimmune disease [53].
3. Role of NK Cells in Globoid Cell Leukodystrophy (GLD)
Globoid cell leukodystrophy (GLD), or Krabbe disease, is an autosomal recessive disease that affects infants [54,55,56]. Patients who are homozygous for this disease are presented between 3 and 6 months of age with irritability, spasticity and mental deterioration. These patients died before the age of two years from hyperpyrexia or respiratory infections. The pathology of GLD is characterized by the destruction of oligodendrocytes, reduced myelin formation and the accumulation of globoid cells, not only in humans, but also in canines, domestic cats and rhesus monkeys [57,58,59,60]. Accumulation of the toxic lipid D-galactosyl-β1-1’ sphingosine (galactosylsphingosine = GalSph) in the brain is thought to cause the disease [55,61,62]. Galactosylsphingosine is virtually absent from normal brain or other tissues [63], but it accumulates at high concentrations in the brain of Krabbe patients, due to the deficiency of the enzyme galactosyl ceramidase, “GALC or GC” (Figure 3).
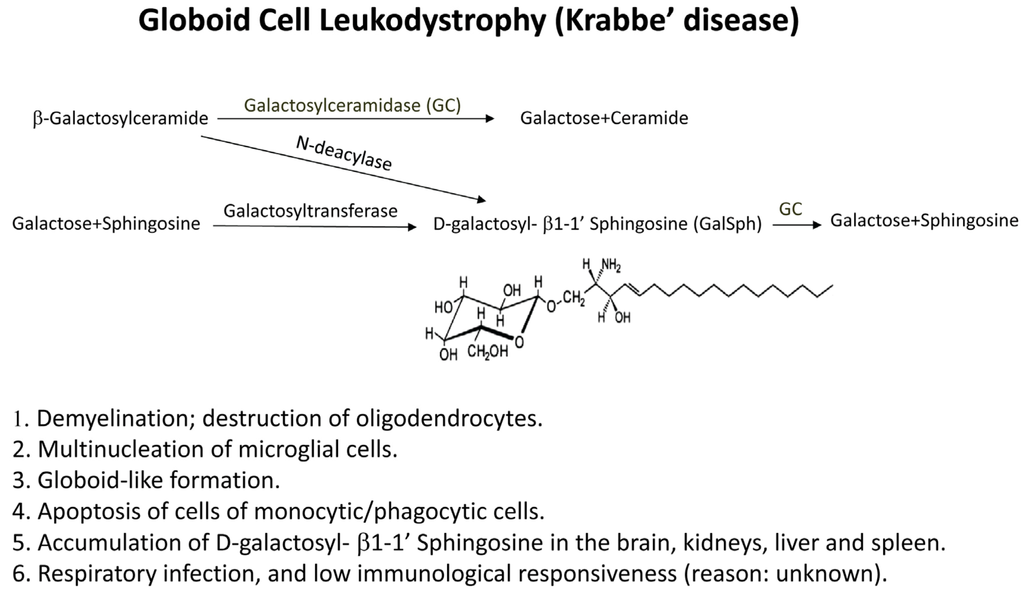
Figure 3.
Formation of glycosphingolipids. Galactosylceramidase (GALC or GC) is deficient in patients with globoid cell leukodystrophy (GLD) or in twitcher mice, resulting in the accumulation of the toxic lipid (GalSph), which, under normal conditions, is converted into galactose and sphingosine by GC. Some characteristics of GLD disease are described in [1,2,3,4,5,6].
Twitcher mice, which have mutations in their GALC gene, represent an authentic model of human Krabbe disease [64]. Galactosylsphingosine is increased in the kidneys, liver, spleen and highly accumulates in the brain of these animals. For example, the level of this lipid can reach up to 15 nM/g body weights in the brain of twitcher mice and about 60 ng/100 mg weights in the spleen of these mice [61]. Galactosylsphingosine induces the in vitro apoptosis of rat C6 glial cells [65] or the oligodendrocytes MO3.13 cell line [66]. Apoptotic cells were also observed in the brain of patients with various lysosomal storage diseases, such as Tay-Sachs and Sandhoff diseases, as well as the brain of mice with GM2 gangliosidosis [67]. A heptahelical receptor that binds heterotrimeric G proteins, named T-cell-death-associated gene 8 “TDAG8” [68,69], has been shown to bind GalSph, as well as glucosylated sphingosine “GlcSph”. Human TDAG8 exists in normal tissues and is restricted to lymphoid organs, such as spleen and lymph nodes, as well as its expression in peripheral blood lymphocytes. We reported that human NK cells express TDAG8 and demonstrated that GalSph and GlcSph damage these cells [70]. Although GalSph and GlcSph induce the apoptosis and multinucleation in NK cells, most other lipids examined did not affect the morphology or induced apoptosis of these cells [70].
Patients with storage diseases that involve the nervous system succumb to infections, which are the major cause of death in these patients [71]. The reason for increased rate of infections is not clear. Our results showing that galactosylsphingosine induces the in vitro chemotaxis of NK cells [70] and their accumulation in twitcher mice splenic white pulps [52], corroborated with induction of their apoptosis, may provide an explanation to this enigma. In fact, it was very difficult to isolate NK cells from the spleens of twitcher mice in order to examine their in vitro activities, which could be due to their apoptosis [52]. In addition to NK cells, cells with Ia+ phenotype that include macrophages and dendritic cells also disappeared from the spleens of twitcher mice. These findings may provide an explanation to the observations that twitcher mice have low immunological activity and can easily accept grafts [72]. In line with these observations is the demonstration that macrophages may counteract the demyelination in twitcher mice [73].
Transplantation of hematopoietic cells from normal mice into syngeneic twitcher mice resulted in increased survival, but in these early studies, the success of the transplantation procedures did not go beyond 100 days [74,75]. However, recent advancements revealed that administration of murine bone marrow-derived stem cells via intraperitoneal injection resulted in improved lifespan and reduced twitching severity and frequency [76]. In humans, Krivit et al. [77] reported a highly successful rate of recovery in five patients with late-onset GLD after hematopoietic stem cell transplantation, and the success of these transplantation procedures was attributed to the ability of donor macrophages to provide the enzyme necessary to hydrolyze galactosylsphingosine. Based on these observations, it was shown that administration of the enzyme GALC inserted into adeno-virus to twitcher mice resulted in the accumulation of this enzyme in the brain of these mice, which consequently led to prolong their lives [78]. Further, recent studies demonstrated that a combination of enzyme replacement therapy with bone marrow transplantation may have a great promise for treating twitcher mice [79,80]. It is not clear whether NK cells were involved in these therapeutic regiments. However, if indeed NK cells were present in the grafts, then these cells may have helped in resolving the infections associated with the disease.
4. Conclusions
Although two concepts were put forward in the literature regarding the role of NK cells in MS disease, a plausible explanation for these contradictory effects is the presence of different subsets of NK cells. It is clear that the beneficial effects of drugs used to treat MS are always associated with increased NK cytolytic activity against target cells. Therefore, it can be concluded that activation of NK cells in MS patients may be an important factor for subsiding the severity of the disease. The discovery of NK17/NK1 cells should add another important value of using these cells in treating autoimmune diseases and, in particular, MS.
Regarding another neurodegenerative disease, NK cells express TDAG8, the receptor for glycosphingolipids galactosyl sphingosine (GalSph) or glucosyl sphingosine (GluSph) and that GalSph and GluSph damage NK cells by inducing their apoptosis. In addition, GalSph induces human NK cell chemotaxis in vitro and promotes the localization of mouse NK cells in the white pulp areas of twitcher mice spleens, corroborated with the induction of their apoptosis. Whether these lipids perform similar functions in the brain of patients suffering from the disease is an issue that has not yet been investigated. The involvement of NK cells may provide an explanation to the observation that patients with lysosomal storage diseases die of respiratory infections as their NK cells are destroyed by the glycosphingolipids. Collectively, these observations should provide new insights into neurodegenerative diseases, and the knowledge can be utilized for formulating new strategies to treat these diseases.
Acknowledgments
The work done in my laboratory was supported by grants from the University of Oslo, Vivi Irene Hansens fund for SLE and Halvor Høies fund for MS.
Conflict of Interests
The author declares no conflict of interest.
References
- Cooper, M.A.; Yokoyama, W.M. Memory-like responses of natural killer cells. Immunol. Rev. 2010, 235, 297–305. [Google Scholar]
- Wood, S.M.; Ljunggren, H.G.; Bryceson, Y.T. Insights into NK cell biology from human genetics and disease associations. Cell. Mol. Life Sci. 2011, 68, 3479–3493. [Google Scholar] [CrossRef]
- Moretta, A.; Bottino, C.; Vitale, M.; Pende, D.; Cantoni, C.; Mingari, M.C.; Biassoni, R.; Moretta, L. Activating receptors and coreceptors involved in human natural killer cell-mediated cytolysis. Annu. Rev. Immunol. 2001, 19, 197–223. [Google Scholar] [CrossRef]
- Ljunggren, H.G.; Malmberg, K.J. Prospects for the use of NK cells in immunotherapy of human cancer. Nat. Rev. Immunol. 2007, 7, 329–339. [Google Scholar] [CrossRef]
- Yang, Q.; Goding, S.R.; Hokland, M.E.; Basse, P.H. Antitumor activity of NK cells. Immunol. Res. 2006, 36, 13–25. [Google Scholar] [CrossRef]
- Salazar-Mather, T.P.; Lewis, C.A.; Biron, C.A. Type I interferons regulate inflammatory cell trafficking and macrophage inflammatory protein 1α delivery to the liver. J. Clin. Invest. 2002, 110, 321–330. [Google Scholar]
- Maghazachi, A.A.; Al-Aoukaty, A. Chemokines activate natural killer cells through heterotrimeric G-proteins: Implications for the treatment of AIDS and cancer. FASEB J. 1998, 12, 913–924. [Google Scholar]
- Cooper, M.A.; Fehniger, T.A.; Caligiuri, M.A. The biology of human natural killer-cell subsets. Trends Immunol. 2001, 22, 633–640. [Google Scholar] [CrossRef]
- Rolstad, B.; Herberman, R.B.; Reynolds, C.W. Natural killer cell activity in the rat. V. The circulation patterns and tissue localization of peripheral blood large granular lymphocytes (LGL). J. Immunol. 1986, 136, 2800–2808. [Google Scholar]
- Sandel, M.H.; Speetjens, F.M.; Menon, A.G.; Albertsson, P.A.; Basse, P.H.; Hokland, M.; Nagelkerke, J.F.; Tollenaar, R.A.; van de Velde, C.J.; Kuppen, P.J. Natural killer cells infiltrating colorectal cancer and MHC class I expression. Mol. Immunol. 2005, 42, 541–546. [Google Scholar] [CrossRef]
- Kuppen, P.J.; Gorter, A.; Hagenaars, M.; Jonges, L.E.; Giezeman-Smits, K.M.; Nagelkerke, J.F.; Fleuren, G.; van de Velde, C.J. Role of NK cells in adoptive immunotherapy of metastatic colorectal cancer in a syngeneic rat model. Immunol. Rev. 2001, 184, 236–243. [Google Scholar]
- Maghazachi, A.A. Compartmentalization of human natural killer cells. Mol. Immunol. 2005, 42, 523–529. [Google Scholar] [CrossRef]
- Maghazachi, A.A. Role of natural killer cells in multiple sclerosis. ISRN Immunol. 2012. [Google Scholar] [CrossRef]
- Schleinitz, N.; Vély, F.; Harlé, J.R.; Vivier, E. Natural killer cells in human autoimmune diseases. Immunology 2010, 131, 451–458. [Google Scholar] [CrossRef]
- Dalbeth, N.; Callan, M.F.C. A subset of natural killer cells is greatly expanded within inflamed joints. Arthritis Rheum. 2002, 46, 1763–1772. [Google Scholar] [CrossRef]
- Schurgers, E.; Billiau, A.; Matthys, P. Collagen-induced arthritis as an animal model for rheumatoid arthritis: Focus on interferon-γ. J. Interferon Cytokine Res. 2011, 31, 917–926. [Google Scholar] [CrossRef]
- Kveberg, L.; Bryceson, Y.; Inngjerdingen, M.; Rolstad, B.; Maghazachi, A.A. Sphingosine 1 phosphate induces the chemotaxis of human natural killer cells. Role for heterotrimeric G proteins and phosphoinositide 3 kinases. Eur. J. Immunol. 2002, 32, 1856–1864. [Google Scholar] [CrossRef]
- Jin, Y.; Knudsen, E.; Wang, L.; Maghazachi, A.A. Lysophosphatidic acid induces human natural killer cell chemotaxis and intracellular calcium mobilization. Eur. J. Immunol. 2003, 33, 2083–2089. [Google Scholar] [CrossRef]
- Rolin, J.; Maghazachi, A.A. Effects of lysophospholipids on tumor microenvironment. Cancer Microenviron. 2011, 4, 393–403. [Google Scholar] [CrossRef]
- Martins, L.C.; Rocha, N.P.; Torres, K.C.; dos Santos, R.R.; França, G.S.; de Moraes, E.N.; Mukhamedyarov, M.A.; Zefirov, A.L.; Rizvanov, A.A.; Kiyasov, A.P.; et al. Disease-specific expression of the serotonin-receptor 5-HT(2C) in natural killer cells in Alzheimer’s dementia. J. Neuroimmunol. 2012, 251, 73–79. [Google Scholar] [CrossRef]
- Masera, R.G.; Prolo, P.; Sartori, M.L.; Staurenghi, A.; Griot, G.; Ravizza, L.; Dovio, A.; Chiappelli, F.; Angeli, A. Mental deterioration correlates with response of natural killer (NK) cell activity to physiological modifiers in patients with short history of Alzheimer’s disease. Psychoneuroendocrinology 2002, 27, 447–461. [Google Scholar] [CrossRef]
- Peritt, D.; Robertson, S.; Gri, G.; Showe, L.; Aste-Amezaga, M.; Trinchieri, G. Differentiation of human NK cells into NK1 and NK2 subsets. J. Immunol. 1998, 161, 5821–5824. [Google Scholar]
- Berahovich, R.D.; Lai, N.L.; Wei, Z.; Lanier, L.L.; Schall, T.J. Evidence for NK cell subsets based on chemokine receptor expression. J. Immunol. 2006, 177, 7833–7840. [Google Scholar]
- Cella, M.; Fuchs, A.; Vermi, W.; Facchetti, F.; Otero, K.; Lennerz, J.K.; Doherty, J.M.; Mills, J.C.; Colonna, M. A human natural killer cell subset provides an innate source of IL-22 for mucosal immunity. Nature 2009, 457, 722–725. [Google Scholar] [CrossRef]
- Sanos, S.L.; Bui, V.L.; Mortha, A.; Oberle, K.; Heners, C.; Johner, C.; Diefenbach, A. RORγt and commensal microflora are required for the differentiation of mucosal interleukin 22-producing NKp46+ cells. Nat. Immunol. 2009, 10, 83–91. [Google Scholar]
- Hughes, T.; Becknell, B.; McClory, S.; Briercheck, E.; Freud, A.G.; Zhang, X.; Mao, H.; Nuovo, G.; Yu, J.; Caligiuri, M.A. Stage 3 immature human natural killer cells found in secondary lymphoid tissue constitutively and selectively express the TH17 cytokine interleukin-22. Blood 2009, 113, 4008–4010. [Google Scholar] [CrossRef]
- Pandya, A.D.; Al-Jaderi, Z.; Høglund, R.A.; Holmøy, T.; Harbo, H.F.; Norgauer, J.; Maghazachi, A.A. Identification of human NK17/NK1 cells. PLoS One 2011, 6, e26780. [Google Scholar]
- Hwang, S.Y.; Kim, J.Y.; Kim, K.W.; Park, M.K.; Moon, Y.; Kim, W.U.; Kim, H.Y. IL-17 induces production of IL-6 and IL-8 in rheumatoid arthritis synovial fibroblasts via NF-κB- and PI3-kinase/Akt-dependent pathways. Arthritis. Res. Ther. 2004, 6, R120–R128. [Google Scholar] [CrossRef]
- Sylvester, J.; Liacini, A.; Li, W.Q.; Zafarullah, M. Interleukin-17 signal transduction pathways implicated in inducing matrix metalloproteinase-3, -13 and aggrecanase-1 genes in articular chondrocytes. Cell. Signal. 2004, 16, 469–476. [Google Scholar] [CrossRef]
- Honorati, M.C.; Neri, S.; Cattini, L.; Facchini, A. Interleukin-17, a regulator of angiogenic factor release by synovial fibroblasts. Osteoarthr. Cartil. 2006, 14, 345–352. [Google Scholar]
- O’Connor, W., Jr.; Kamanaka, M.; Booth, C.J.; Town, T.; Nakae, S.; Iwakura, Y.; Kolls, J.K.; Flavell, R.A. A protective function for interleukin 17A in T cell-mediated intestinal inflammation. Nat. Immunol. 2009, 10, 603–609. [Google Scholar] [CrossRef]
- Troitskaya, M.; Baysa, A.; Vaage, J.; Sand, K.L.; Maghazachi, A.A.; Valen, G. Interleukin-17 (IL-17) Expression is reduced during acute myocardial infarction: Role on chemokine receptor expression in monocytes and their in vitro chemotaxis towards chemokines. Toxins 2012, 4, 1427–1439. [Google Scholar] [CrossRef]
- Gold, R.; Linington, C.; Lassmann, H. Understanding pathogenesis and therapy of multiple sclerosis via animal models: 70 years of merits and culprits in experimental autoimmune encephalomyelitis. Brain 2006, 129, 1953–1971. [Google Scholar] [CrossRef]
- Friese, M.A.; Montalban, X.; Wilcox, N.; Bell, J.I.; Martin, R.; Fugger, L. The value of animal models for drug development in multiple sclerosis. Brain 2006, 129, 1940–1952. [Google Scholar] [CrossRef]
- Hestvick, A.L.K. The Double-edged sword of autoimmunity: Lessons from multiple sclerosis. Toxins 2010, 2, 856–877. [Google Scholar] [CrossRef]
- Shi, F.-D.; Zhou, Q. Natural killer cells as indispensable players and therapeutic targets in autoimmunity. Autoimmunity 2011, 44, 3–10. [Google Scholar] [CrossRef]
- Maghazachi, A.A. Role of chemokines in the biology of natural killer cells. Curr. Top. Microbiol. Immunol. 2010, 341, 37–58. [Google Scholar] [CrossRef]
- Segal, B.M. The role of natural killer cells in curbing neuroinflammation. J. Neuroimmunol. 2007, 191, 2–7. [Google Scholar] [CrossRef]
- Takahashi, K.; Aranami, T.; Endoh, M.; Miyake, S.; Yamamura, T. The regulatory role of natural killer cells in multiple sclerosis. Brain 2004, 127, 1917–1927. [Google Scholar] [CrossRef]
- Huang, D.; Shi, F.D.; Jung, S.; Pien, G.C.; Wang, J.; Salazar-Mather, T.P.; He, T.T.; Weaver, J.T.; Ljunggren, H.G.; Biron, C.A.; et al. The neuronal chemokine CX3CL1/fractalkine selectively recruits NK cells that modify experimental autoimmune encephalomyelitis within the central nervous system. FASEB J. 2006, 20, 896–905. [Google Scholar] [CrossRef]
- Hao, J.; Liu, R.; Piao, W.; Zhou, Q.; Vollmer, T.L.; Campagnolo, D.I.; Xiang, R.; la Cava, A.; Van Kaer, L.; Shi, F.D. Central nervous system (CNS)-resident natural killer cells suppress Th17 responses and CNS autoimmune pathology. J. Exp. Med. 2010, 207, 1907–1921. [Google Scholar] [CrossRef]
- Kastrukoff, L.F.; Morgan, N.G.; Zecchini, D.; White, R.; Petkau, A.J.; Satoh, J.; Paty, D.W. A role for natural killer cells in the immunopathogenesis of multiple sclerosis. J. Neuroimmunol. 1998, 86, 123–133. [Google Scholar] [CrossRef]
- Pagenstecher, A.; Lassmann, S.; Carson, M.J.; Kincaid, C.L.; Stalder, A.K.; Campbell, I.L. Astrocyte-targeted expression of IL-12 induces active cellular immune responses in the central nervous system and modulates experimental allergic encephalomyelitis. J. Immunol. 2000, 164, 4481–4492. [Google Scholar]
- Shi, F.D.; Takeda, K.; Akira, S.; Sarvetnick, N.; Ljunggren, H.G. IL-18 directs autoreactive T cells and promotes autodestruction in the central nervous system via induction of IFN-γ by NK cells. J. Immunol. 2000, 165, 3099–3104. [Google Scholar]
- Winkler-Pickett, R.; Young, H.A.; Cherry, J.M.; Diehl, J.; Wine, J.; Back, T.; Bere, W.E.; Mason, A.T.; Ortaldo, J.R. In vivo regulation of experimental autoimmune encephalomyelitis by NK cells: Alteration of primary adaptive responses. J. Immunol. 2008, 180, 4495–4506. [Google Scholar]
- Kala, M.; Miravalle, A.; Vollmer, T. Recent insights into the mechanism of action of glatiramer acetate. J. Neuroimmunol. 2011, 235, 9–17. [Google Scholar] [CrossRef]
- Johnson, K.P.; Brooks, B.R.; Ford, C.C.; Goodman, A.D.; Lisak, R.P.; Myers, L.W.; Pruitt, A.A.; Rizzo, M.A.; Rose, J.W.; Weiner, L.P.; Wolinsky, J.S. Glatiramer acetate (Copaxone): Comparison of continuous versus delayed therapy in a six-year organized multiple sclerosis trial. Multiple Sclerosis 2003, 9, 585–591. [Google Scholar] [CrossRef]
- Arnon, R.; Aharoni, R. Mechanism of action of glatiramer acetate in multiple sclerosis and its potential for the development of new applications. Proc. Natl. Acad. Sci. USA 2004, 101, 14593–14598. [Google Scholar] [CrossRef]
- Høglund, R.A.; Holmøy, T.; Harbo, H.F.; Maghazachi, A.A. A one year follow-up study of natural killer and dendritic cells activities in multiple sclerosis patients receiving glatiramer acetate (GA). PLoS One 2013, in press. [Google Scholar]
- Moretta, A. Natural killer cells and dendritic cells: Rendezvous in abused tissues. Nat. Rev. Immunol. 2002, 2, 957–964. [Google Scholar] [CrossRef]
- Sand, K.L.; Knudsen, E.; Rolin, J.; Al-Falahi, Y.; Maghazachi, A.A. Modulation of natural killer cell cytotoxicity and cytokine release by the drug glatiramer acetate. Cell. Mol. Life Sci. 2009, 66, 1446–1456. [Google Scholar] [CrossRef]
- Al-Falahi, Y.; Sand, K.L.; Knudsen, E.; Damaj, B.B.; Rolin, J.; Maghazachi, A.A. Splenic natural killer cell activity in two models of experimental neurodegenerative diseases. J. Cell. Mol. Med. 2009, 13, 2693–2703. [Google Scholar] [CrossRef]
- Hidalgo, E.; Essex, S.J.; Yeo, L.; Curnow, S.J.; Filer, A.; Cooper, M.S.; Thomas, A.M.; McGettrick, H.M.; Salmon, M.; Buckley, C.D.; et al. The response of T cells to interleukin-6 is differentially regulated by the microenvironment of the rheumatoid synovial fluid and tissue. Arthritis Rheum. 2011, 63, 3284–3293. [Google Scholar]
- Krabbe, K. A new familial, infantile form of diffuse brain sclerosis. Brain 1916, 30, 74–114. [Google Scholar] [CrossRef]
- Svennerholm, L.; Vanier, M.-T.; Månsson, J.E. Krabbe disease: A galactosylsphingosine (psychosine) lipidosis. J. Lipid Res. 1980, 21, 53–64. [Google Scholar]
- Vanier, M.-T.; Svennerholm, L. Chemical pathology of Krabbe’s disease. III. Ceramide hexosides and gangliosides of brain. Acta Paediatr. Scand. 1975, 64, 641–648. [Google Scholar] [CrossRef]
- Baskin, G.B.; Ratterree, M.; Davison, B.B.; Falkenstein, K.P.; Clarke, M.R.; England, J.D.; Vanier, M.T; Luzi, P.; Rafi, M.A.; Wenger, D.A. Genetic galactocerebrosidase deficiency (globoid cell leukodystrophy, Krabbe disease) in rhesus monkeys (Macaca mulatta). Lab. Anim. Sci. 1998, 48, 476–482. [Google Scholar]
- Wenger, D.A. Murine, canine and non-human primate models of Krabbe disease. Mol. Med. Today 2000, 6, 449–451. [Google Scholar] [CrossRef]
- Wenger, D.A.; Victoria, T.; Rafi, M.A.; Luzi, P.; Vanier, M.T.; Vite, C.; Patterson, D.F.; Haskins, M.H. Globoid cell leukodystrophy in cairn and West Highland white terriers. J. Hered. 1990, 90, 138–142. [Google Scholar]
- Sigurdson, C.J.; Basaraba, R.J.; Mazzaferro, E.M.; Gould, D.H. Globoid cell-like leukodystrophy in a domestic longhaired cat. Vet. Pathol. 2002, 39, 494–496. [Google Scholar] [CrossRef]
- Whitfield, P.D.; Sharp, P.C.; Taylor, R.; Meikle, P. Quantification of galactosylsphingosine in the twitcher mouse using electrospray ionization-tandem mass spectrometry. J. Lipid Res. 2001, 42, 2092–2095. [Google Scholar]
- White, A.B.; Givogri, M.I.; Lopez-Rosas, A.; Cao, H.; van Breemen, R.; Thinakaran, G.; Bongarzone, E.R. Psychosine accumulates in membrane microdomains in the brain of krabbe patients, disrupting the raft architecture. J. Neurosci. 2009, 29, 6068–6077. [Google Scholar]
- Duchen, L.W.; Eicher, E.M.; Jacobs, J.M.; Scaravilli, F.; Teixeira, F. Hereditary leucodystrophy in the mouse: The new mutant twitcher. Brain 1980, 103, 695–710. [Google Scholar] [CrossRef]
- Tanaka, K.; Nagara, H.; Kobayashi, T.; Goto, I. The twitcher mouse: Accumulation of galactosylsphingosine and pathology of sciatic nerve. Brain Res. 1988, 454, 340–346. [Google Scholar] [CrossRef]
- Jatana, M.; Giri, S.; Singh, A.K. Apoptotic positive cells in Krabbe brain and induction of apoptosis in rat C6 glial cells by psychosine. Neurosci. Lett. 2002, 330, 183–187. [Google Scholar] [CrossRef]
- Haq, E.; Giri, S.; Singh, I.; Singh, A.K. Molecular mechanism of psychosine-induced cell death in human oligodendrocyte cell line. J. Neurochem. 2003, 86, 1428–1440. [Google Scholar] [CrossRef]
- Huang, J.Q.; Trasler, J.M.; Igdoura, S.; Michaud, J.; Hanal, N.; Gravel, R.A. Apoptotic cell death in mouse models of GM2 gangliosidosis and observations on human Tay-Sachs and Sandhoff diseases. Hum. Mol. Genet. 1997, 6, 1879–1885. [Google Scholar] [CrossRef]
- Choi, J.W.; Lee, S.Y.; Choi, Y. Identification of a putative G protein-coupled receptor induced during activation-induced apoptosis of T cells. Cell. Immunol. 1996, 168, 78–84. [Google Scholar] [CrossRef]
- Kyaw, H.; Zeng, Z.; Su, K.; Fan, P.; Shell, B.K.; Carter, K.C.; Li, Y. loning, characterization, and mapping of human homolog of mouse T-cell death-associated gene. DNA Cell Biol. 1998, 17, 493–500. [Google Scholar] [CrossRef]
- Maghazachi, A.A.; Knudsen, E.; Jin, Y.; Jenstad, M.; Chaudhry, F.A. D-galactosyl-β1–1’ sphingosine and D-glucosyl-β1–1’ sphingosine induce human natural killer cell apoptosis. Biochem. Biophys. Res. Commun. 2004, 320, 810–815. [Google Scholar] [CrossRef]
- Sabatelli, M.; Quaranta, L.; Madia, F.; Lippi, G.; Conte, A.; lo Monaco, M.; di Trapani, G.; Rafi, M.A.; Wenger, D.A.; Vaccaro, A.M.; Tonali, P. Peripheral neuropathy with hypomyelinating features in adult-onset Krabbe’s disease. Neuromuscular Disord. 2002, 12, 386–391. [Google Scholar] [CrossRef]
- Wenger, D.A.; Coppola, S.; Liu, S.-L. Insights into the diagnosis and treatment of lysosomal storage diseases. Arch. Neurol. 2003, 60, 322–328. [Google Scholar] [CrossRef]
- Kondo, Y.; Adams, J.M.; Vanier, M.T.; Duncan, I.D. Macrophages counteract demyelination in a mouse model of globoid cell leukodystrophy. J. Neurosci. 2011, 31, 3610–3624. [Google Scholar]
- Ichioka, T.; Kishimoto, Y.; Brennan, S.; Santos, G.W.; Yeager, A.M. Hematopoietic cell transplantation in murine globoid cell leukodystrophy (the twitcher mouse): Effects on levels of galactosylceramidase, psychosine, and galactocerebrosides. Proc. Natl. Acad. Sci. USA 1987, 84, 4259–4263. [Google Scholar] [CrossRef]
- Yeager, A.M.; Brennan, S.; Tiffany, C.; Moser, H.W.; Santos, G.W. Prolonged survival and remyelination after hematopoietic cell transplantation in the twitcher mouse. Science 1984, 225, 1052–1054. [Google Scholar]
- Scruggs, B.A.; Bowles, A.C.; Zhang, X.; Semon, J.A.; Kyzar, E.J.; Myers, L.; Kalueff, A.V.; Bunnell, B.A. High-throughput screening of stem cell therapy for globoid cell leukodystrophy using automated neurophenotyping of twitcher mice. Behav. Brain Res. 2013, 236, 35–47. [Google Scholar] [CrossRef]
- Krivit, W.; Shapiro, E.G.; Peters, C.; Wagner, J.E.; Cornu, G.; Kurtzberg, J.; Wenger, D.A.; Kolodny, E.H.; Vanier, M.T.; Loes, D.J.; et al. Hematopoietic stem-cell transplantation in globoid-cell leukodystrophy. N. Engl. J. Med. 1998, 338, 1119–1126. [Google Scholar] [CrossRef]
- Rafi, M.A.; Rao, H.Z.; Luzi, P.; Curtis, M.T.; Wenger, D.A. Extended normal life after AAVrh10-mediated gene therapy in the mouse model of krabbe disease. Mol. Ther. 2012, 20, 2031–2042. [Google Scholar] [CrossRef]
- Qin, E.Y.; Hawkins-Salsbury, J.A.; Jiang, X.; Reddy, A.S.; Farber, N.B.; Ory, D.S.; Sands, M.S. Bone marrow transplantation increases efficacy of central nervous system-directed enzyme replacement therapy in the murine model of globoid cell leukodystrophy. Mol. Genet. Metab. 2012, 107, 186–196. [Google Scholar] [CrossRef]
- Reddy, A.S.; Kim, J.H.; Hawkins-Salsbury, J.A.; Macauley, S.L.; Tracy, E.T.; Vogler, C.A.; Han, X.; Song, S.K.; Wozniak, D.F.; Fowler, S.C.; Klein, R.S.; Sands, M.S. Bone marrow transplantation augments the effect of brain- and spinal cord-directed adeno-associated virus 2/5 gene therapy by altering inflammation in the murine model of globoid-cell leukodystrophy. J. Neurosci. 2011, 31, 9945–9957. [Google Scholar]
© 2013 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).