Maresin 2, a Specialized Pro-Resolution Lipid Mediator, Reduces Pain and Inflammation Induced by Bothrops jararaca Venom in Mice


 , , , ,
, , , ,  , , ,
, , ,  and
and
Abstract
1. Introduction
2. Results
2.1. BjV Induces Dose- and Time-Dependent Hyperalgesia That Is Amenable by MaR2 Treatment
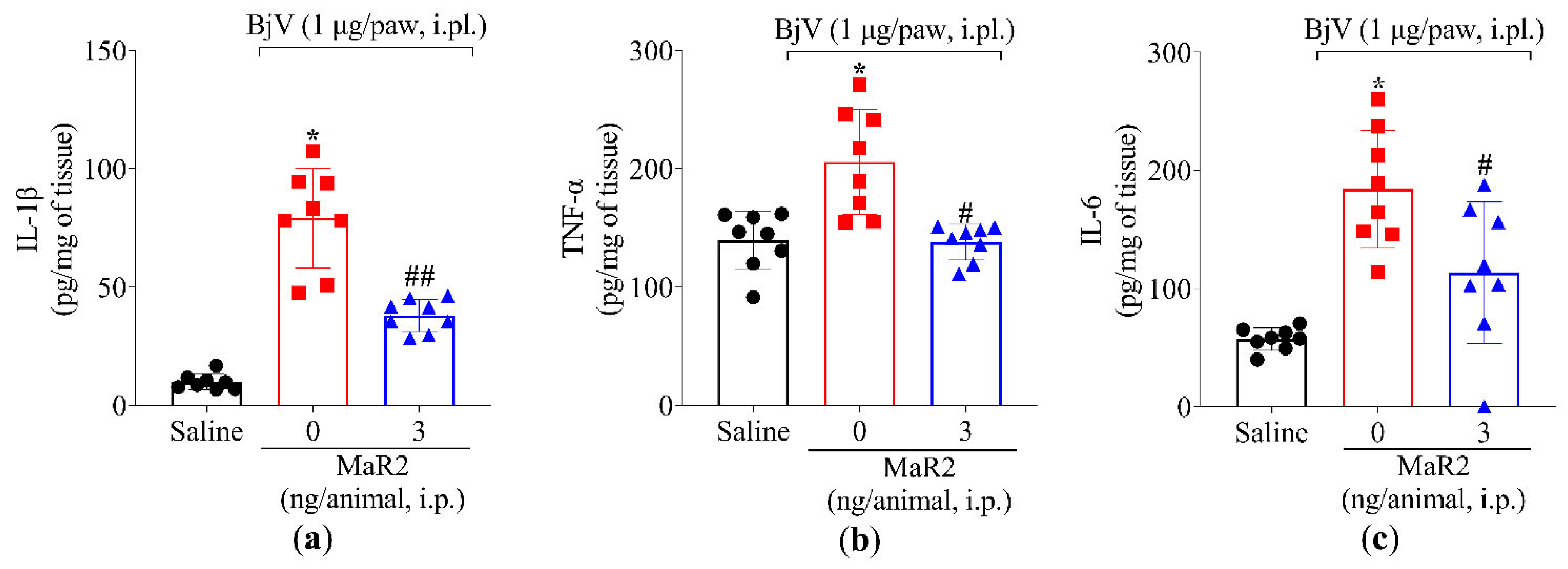
2.2. MaR2 Reduces Myeloperoxidase (MPO) Activity, Oxidative Stress, and Levels of TNF-α, IL-1β, and IL-6 in the Paws of BjV-Injected Mice
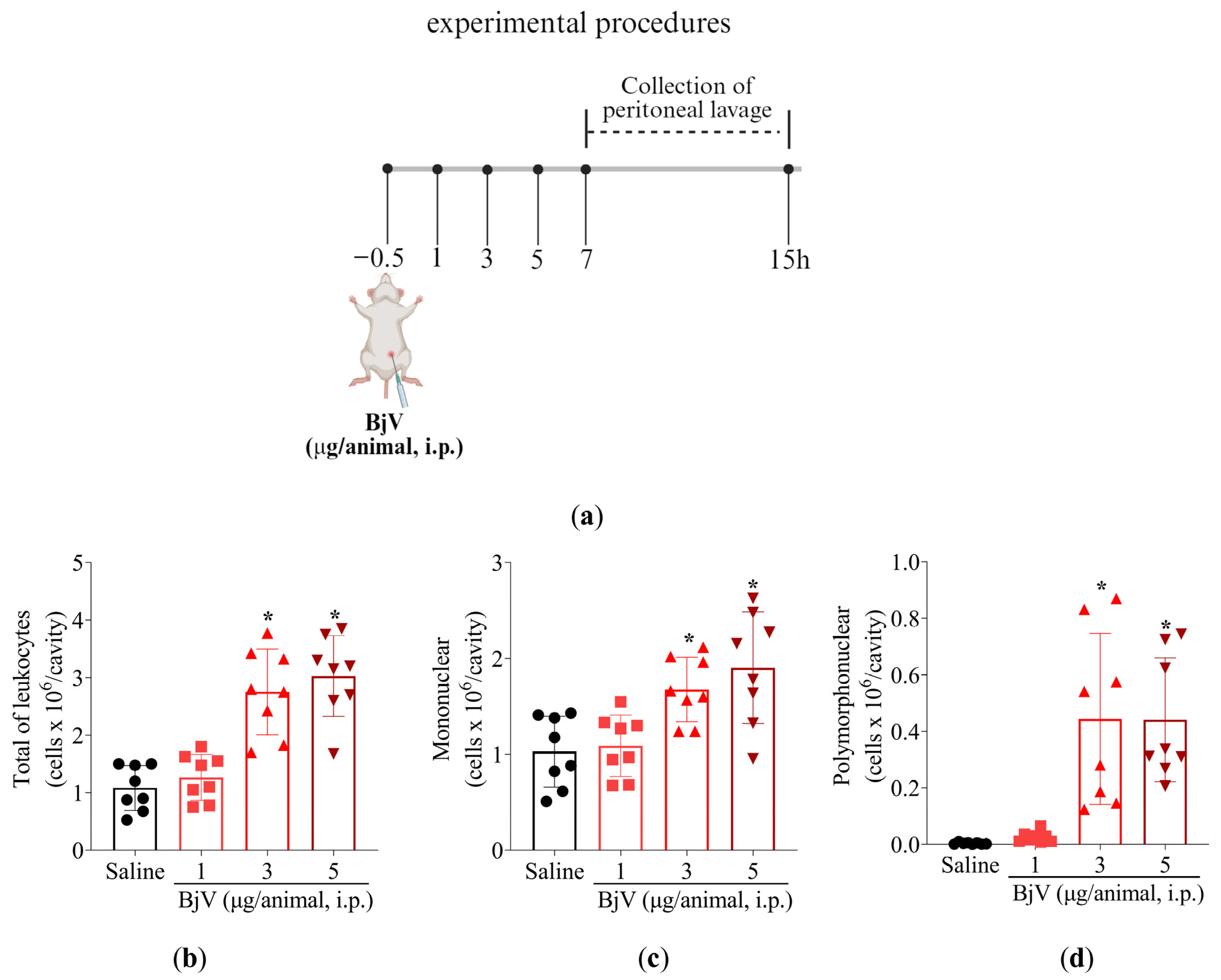
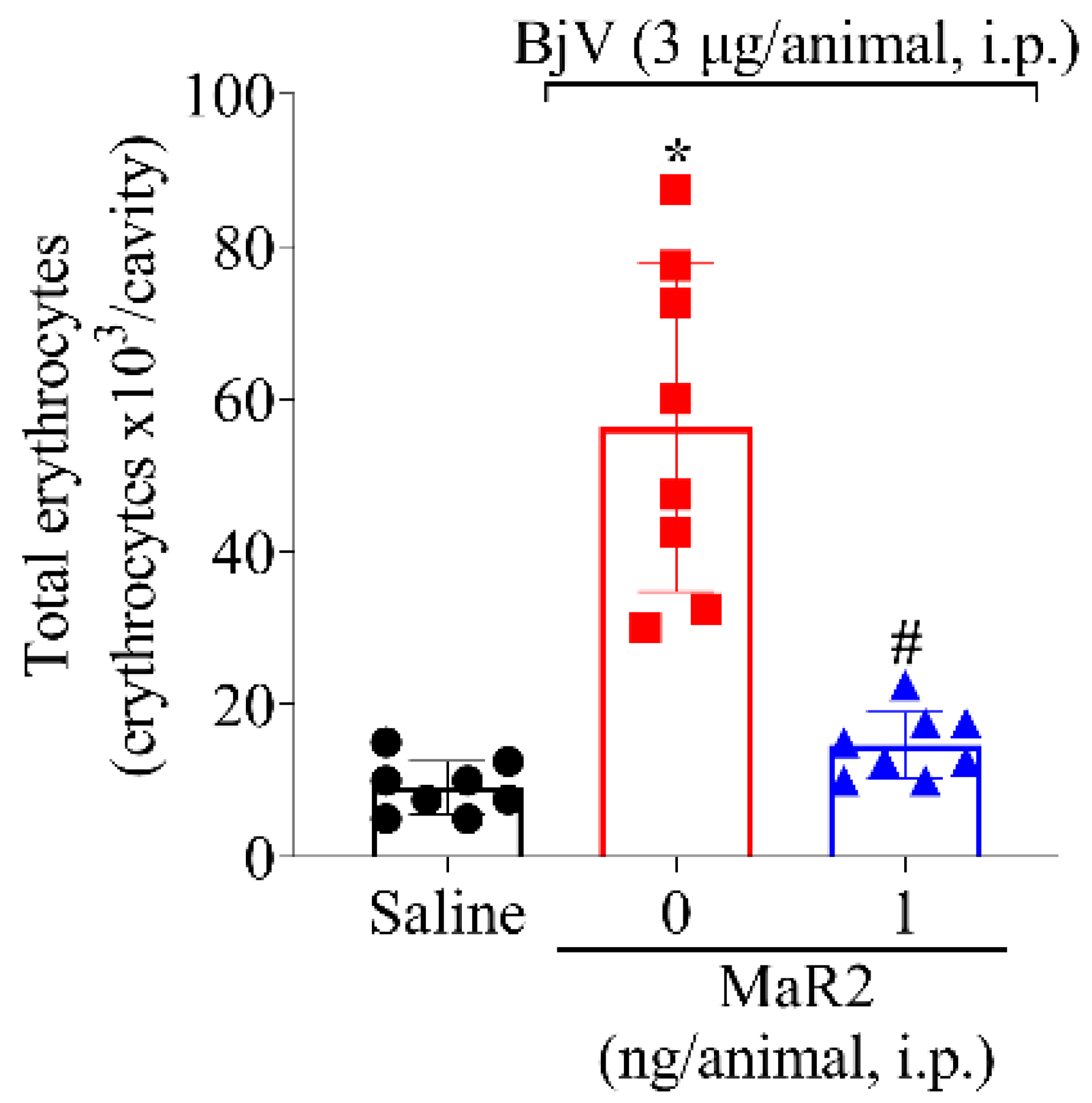
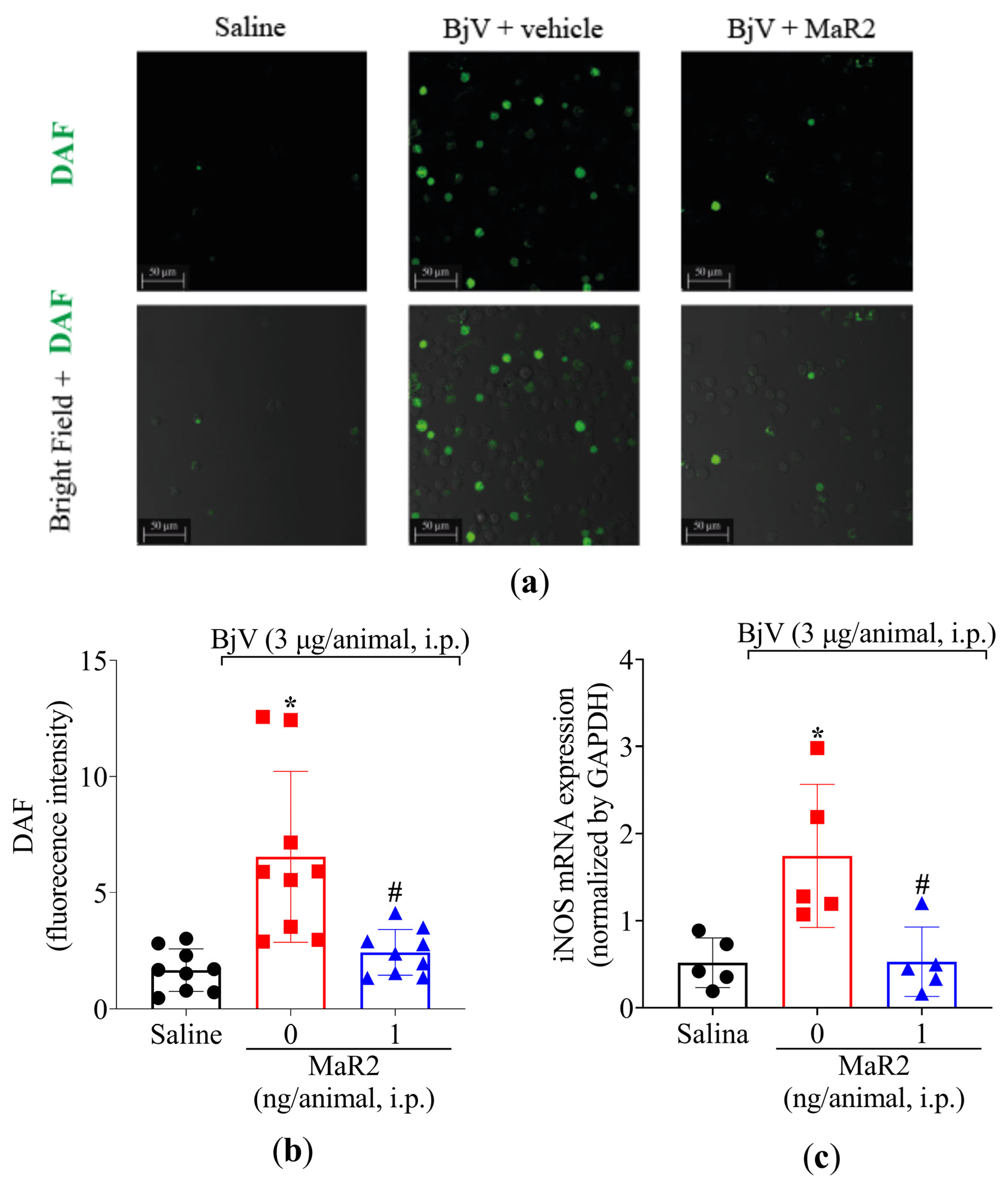
2.3. MaR2 Inhibits Leukocyte Recruitment, Hemorrhage, and Oxidative Stress in Peritonitis Induced by BjV
2.3.1. MaR2 Inhibits Leukocyte Recruitment
2.3.2. MaR2 Inhibits BjV-Induced Hemorrhage
2.3.3. MaR2 Inhibits Leukocyte Activity and Oxidative Stress Caused by BjV
3. Discussion
4. Conclusions
5. Materials and Methods
5.1. Animals
5.2. General Experimental Procedures
5.3. Test Compounds
5.4. Mechanical Hyperalgesia Assessment
5.5. Thermal Hyperalgesia Assessment
5.6. Static Weight Bearing (SWB)
5.7. Evaluation of Leukocytes Profile
5.7.1. Leukocyte Migration and Differential Cell Counts
5.7.2. Myeloperoxidase Activity (MPO)
5.8. Oxidative Stress
5.8.1. Nitroblue Tetrazolium Reduction (NBT) Assay
5.8.2. Microscopic NBT-Positive Cell Count
5.8.3. Antioxidant Efficiency by ABTS
5.8.4. Total Intracellular ROS Detection
5.8.5. Intracellular Nitric Oxide (NO) Detection
5.9. Cytokines Quantitation
5.10. Erythrocyte Count
5.11. RT-qPCR
5.12. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| ABTS | 2,2′-azino-bis(3-ethylbenzothiazoline-6-sulfonic acid |
| BjV | Bothrops jararaca venom |
| CGRP | Calcitonin gene-related peptide |
| DMSO | Dimethyl sulfoxide |
| EDTA | Ácido etilenodiaminotetracético |
| IASP | International Association for the Study of Pain |
| iNOS | Inducible nitric oxide synthase |
| HTAB | Hexadecyltrimethylammonium bromide |
| IL-1β | Interleukin-1β |
| IL-6 | Interleukin-6 |
| I.P. | Intraperitoneal |
| I.PL. | Intraplantar |
| KCl | Potassium chloride |
| K2HPO4 | Dipotassium phosphate |
| LPS | Lipopolysaccharide |
| MaR1 | Maresin 1 |
| MaR2 | Maresin 2 |
| MPO | Myeloperoxidase |
| NBT | Nitroblue tetrazolium |
| NFκB | Nuclear factor kappa-B |
| NLRP3 | Nucleotide-binding oligomerization domain (NOD)-like receptor pyrin domain containing 3 |
| NO | Nitric oxide |
| O2•− | Superoxide anion |
| PBS | Phosphate-buffered saline |
| PMN | Polymorphonuclear leukocyte |
| RNS | Reactive nitrogen species |
| ROS | Reactive oxygen species |
| SPM | Specialized pro-resolution lipid mediators |
| SWB | Static weight bearing |
| TRPA1 | Transient receptor potential ankyrin 1 |
| TRPV1 | Transient receptor potential vanilloid 1 |
| TNF-α | Tumor necrosis factor alpha |
References
- da Saúde, M. Boletim Epidemiológico Vol. 53-No 36; Secretariat of Health Surveillance, Ministry of Health: Rio de Janeiro, Brazil, 2022; Volume 53, pp. 1–19.
- Ribeiro, L.A.; Jorge, M.T. Bites by snakes in the genus Bothrops: A series of 3139 cases. Rev. Soc. Bras. Med. Trop. 1997, 30, 475–480. [Google Scholar] [CrossRef] [PubMed]
- Ferraz, C.R.; Carvalho, T.T.; Fattori, V.; Saraiva-Santos, T.; Pinho-Ribeiro, F.A.; Borghi, S.M.; Manchope, M.F.; Zaninelli, T.H.; Cunha, T.M.; Casagrande, R.; et al. Jararhagin, a Snake Venom Metalloproteinase, Induces Mechanical Hyperalgesia in Mice with the Neuroinflammatory Contribution of Spinal Cord Microglia and Astrocytes. Int. J. Biol. Macromol. 2021, 179, 610–619. [Google Scholar] [CrossRef] [PubMed]
- Ferraz, C.R.; Calixto-Campos, C.; Manchope, M.F.; Casagrande, R.; Clissa, P.B.; Baldo, C.; Verri, W.A. Jararhagin-Induced Mechanical Hyperalgesia Depends on TNF-α, IL-1β and NFκB in Mice. Toxicon 2015, 103, 119–128. [Google Scholar] [CrossRef]
- Clissa, P.B.; Laing, G.D.; Theakston, R.D.G.; Mota, I.; Taylor, M.J.; Moura-da-Silva, A.M. The Effect of Jararhagin, a Metalloproteinase from Bothrops Jararaca Venom, on pro-Inflammatory Cytokines Released by Murine Peritoneal Adherent Cells. Toxicon 2001, 39, 1567–1573. [Google Scholar] [CrossRef] [PubMed]
- Gutiérrez, J.M.; Lomonte, B. Phospholipase A2 Myotoxins from Bothrops Snake Venoms. Toxicon 1995, 33, 1405–1424. [Google Scholar] [CrossRef]
- Bjarnason, J.B.; Fox, J.W. Hemorrhagic Metalloproteinases from Snake Venoms. Pharmacol. Ther. 1994, 62, 325–372. [Google Scholar] [CrossRef]
- Petricevich, V.L.; Teixeira, C.F.; Tambourgi, D.V.; Gutiérrez, J.M. Increments in Serum Cytokine and Nitric Oxide Levels in Mice Injected with Bothrops Asper and Bothrops jararaca Snake Venoms. Toxicon 2000, 38, 1253–1266. [Google Scholar] [CrossRef]
- Júnior, F.A.N.; Jorge, A.R.C.; Marinho, A.D.; Silveira, J.A.D.M.; Alves, N.T.Q.; Costa, P.H.S.; Silva, P.L.B.E.; Chaves-Filho, A.J.M.; Lima, D.B.; Sampaio, T.L.; et al. Bothrops Alternatus Snake Venom Induces Cytokine Expression and Oxidative Stress on Renal Function. Curr. Top. Med. Chem. 2019, 19, 2058–2068. [Google Scholar] [CrossRef]
- Redza-Dutordoir, M.; Averill-Bates, D.A. Activation of Apoptosis Signalling Pathways by Reactive Oxygen Species. Biochim. Biophys. Acta 2016, 1863, 2977–2992. [Google Scholar] [CrossRef]
- Picolo, G.; Chacur, M.; Gutiérrez, J.M.; Teixeira, C.F.P.; Cury, Y. Evaluation of Antivenoms in the Neutralization of Hyperalgesia and Edema Induced by Bothrops jararaca and Bothrops Asper Snake Venoms. Brazilian J. Med. Biol. Res. Rev. Bras. Pesqui. Medicas e Biol. 2002, 35, 1221–1228. [Google Scholar] [CrossRef]
- Muniz, E.G.; Maria, W.S.; Estevão-Costa, M.I.; Buhrnheim, P.; Chávez-Olórtegui, C. Neutralizing Potency of Horse Antibothropic Brazilian Antivenom against Bothrops Snake Venoms from the Amazonian Rain Forest. Toxicon 2000, 38, 1859–1863. [Google Scholar] [CrossRef]
- Dias da Silva, W.; De Andrade, S.A.; Megale, Â.A.A.; De Souza, D.A.; Sant’Anna, O.A.; Magnoli, F.C.; Guidolin, F.R.; Godoi, K.S.; Saladini, L.Y.; Spencer, P.J.; et al. Antibodies as Snakebite Antivenoms: Past and Future. Toxins 2022, 14, 606. [Google Scholar] [CrossRef]
- Kondo, F.V.; Cabrera, W.H.K.; Ribeiro, O.G.; De Franco, M.; Jensen, J.R.; Picolo, G.; Sant’Anna, M.B.; Spadafora-Ferreira, M.; Borrego, A.; Ibañez, O.M.; et al. Pain and Cellular Migration Induced by Bothrops jararaca Venom in Mice Selected for an Acute Inflammatory Response: Involvement of Mast Cells. Front. Immunol. 2021, 12, 779473. [Google Scholar] [CrossRef]
- Rezende, R.M.; França, D.S.; Menezes, G.B.; dos Reis, W.G.P.; Bakhle, Y.S.; Francischi, J.N. Different Mechanisms Underlie the Analgesic Actions of Paracetamol and Dipyrone in a Rat Model of Inflammatory Pain. Br. J. Pharmacol. 2008, 153, 760–768. [Google Scholar] [CrossRef]
- Ghosh, M.; Acharyya, A.; Bhattacharya, P.; Chakrabortty, S. Role of steroid on management of limb swelling and local pain in haematotoxic snake bite. J. Family Med. Prim. Care. 2022, 11, 7394–7397. [Google Scholar] [CrossRef]
- Stormholt, E.R.; Steiness, J.; Derby, C.B.; Larsen, M.E.; Maagaard, M.; Mathiesen, O. Glucocorticoids Added to Paracetamol and NSAIDs for Post-Operative Pain: A Systematic Review with Meta-Analysis and Trial Sequential Analysis. Acta Anaesthesiol. Scand 2023, 67, 688–702. [Google Scholar] [CrossRef]
- Bonilla-Aldana, D.K.; Bonilla-Aldana, J.L.; Ulloque-Badaracco, J.R.; Al-Kassab-Córdova, A.; Hernandez-Bustamante, E.A.; Alarcon-Braga, E.A.; Siddiq, A.; Benites-Zapata, V.A.; Rodriguez-Morales, A.J.; Luna, C.; et al. Snakebite-Associated Infections: A Systematic Review and Meta-Analysis. Am. J. Trop. Med. Hyg. 2024, 110, 874–886. [Google Scholar] [CrossRef]
- Fetrow, K.O. The Management of Pain in Orthopaedics. Clin. J. Pain 1989, 5 (Suppl. S2), S26–S32, discussion S33–S34. [Google Scholar] [CrossRef]
- Buckley, C.D.; Gilroy, D.W.; Serhan, C.N. Proresolving Lipid Mediators and Mechanisms in the Resolution of Acute Inflammation. Immunity 2014, 40, 315–327. [Google Scholar] [CrossRef]
- Serhan, C.N.; Chiang, N. Resolution Phase Lipid Mediators of Inflammation: Agonists of Resolution. Curr. Opin. Pharmacol. 2013, 13, 632–640. [Google Scholar] [CrossRef]
- Zaninelli, T.H.; Fattori, V.; Verri, W.A.J. Harnessing Inflammation Resolution in Arthritis: Current Understanding of Specialized Pro-Resolving Lipid Mediators’ Contribution to Arthritis Physiopathology and Future Perspectives. Front. Physiol. 2021, 12, 1444. [Google Scholar] [CrossRef] [PubMed]
- Fattori, V.; Zaninelli, T.H.; Rasquel-Oliveira, F.S.; Casagrande, R.; Verri, W.A. Specialized Pro-Resolving Lipid Mediators: A New Class of Non-Immunosuppressive and Non-Opioid Analgesic Drugs. Pharmacol. Res. 2020, 151, 104549. [Google Scholar] [CrossRef] [PubMed]
- Rasquel-Oliveira, F.S.; Silva, M.D.V.D.; Martelossi-Cebinelli, G.; Fattori, V.; Casagrande, R.; Verri, W.A.J. Specialized Pro-Resolving Lipid Mediators: Endogenous Roles and Pharmacological Activities in Infections. Molecules 2023, 28, 5032. [Google Scholar] [CrossRef] [PubMed]
- Ji, R.-R. Specialized Pro-Resolving Mediators as Resolution Pharmacology for the Control of Pain and Itch. Annu. Rev. Pharmacol. Toxicol. 2023, 63, 273–293. [Google Scholar] [CrossRef]
- Fattori, V.; Pinho-Ribeiro, F.A.; Staurengo-Ferrari, L.; Borghi, S.M.; Rossaneis, A.C.; Casagrande, R.; Verri, W.A., Jr.; Verri, W.A. The Specialised Pro-Resolving Lipid Mediator Maresin 1 Reduces Inflammatory Pain with a Long-Lasting Analgesic Effect. Br. J. Pharmacol. 2019, 176, 1728–1744. [Google Scholar] [CrossRef]
- Deng, B.; Wang, C.-W.; Arnardottir, H.H.; Li, Y.; Cheng, C.-Y.C.; Dalli, J.; Serhan, C.N. Maresin Biosynthesis and Identification of Maresin 2, a New Anti-Inflammatory and pro-Resolving Mediator from Human Macrophages. PLoS ONE 2014, 9, e102362. [Google Scholar] [CrossRef]
- Miranda, J.; Brazil, J.C.; Morris, A.H.; Parkos, C.A.; Quiros, M.; Nusrat, A. Maresin-2 Promotes Mucosal Repair and Has Therapeutic Potential When Encapsulated in Thermostable Nanoparticles. Proc. Natl. Acad. Sci. USA 2023, 120, e2218162120. [Google Scholar] [CrossRef]
- Tozzi, O.N.; Jiacomini, I.G.; Bastos, T.S.B.; Nicolazzi, L.H.C.N.; Dos Santos Luz, R.B.; Paredes, L.C.; Gonçalves, L.E.; Lima, M.H.S.; Verri, W.A.J.; Camara, N.O.S.; et al. Evaluation of the Effects of Loxosceles Intermedia’s Venom in Zebrafish. Toxicol. Rep. 2022, 9, 1410–1418. [Google Scholar] [CrossRef]
- Fattori, V.; Zaninelli, T.H.; Ferraz, C.R.; Brasil-Silva, L.; Borghi, S.M.; Cunha, J.M.; Chichorro, J.G.; Casagrande, R.; Verri, W.A.J. Maresin 2 Is an Analgesic Specialized Pro-Resolution Lipid Mediator in Mice by Inhibiting Neutrophil and Monocyte Recruitment, Nociceptor Neuron TRPV1 and TRPA1 Activation, and CGRP Release. Neuropharmacology 2022, 216, 109189. [Google Scholar] [CrossRef]
- Lopes, R.V.; Baggio, D.F.; Ferraz, C.R.; Bertozzi, M.M.; Saraiva-Santos, T.; Verri Junior, W.A.; Chichorro, J.G. Maresin-2 Inhibits Inflammatory and Neuropathic Trigeminal Pain and Reduces Neuronal Activation in the Trigeminal Ganglion. Curr. Res. Neurobiol. 2023, 4, 100093. [Google Scholar] [CrossRef]
- Serhan, C.N.; Levy, B.D. Resolvins in Inflammation: Emergence of the pro-Resolving Superfamily of Mediators. J. Clin. Investig. 2018, 128, 2657–2669. [Google Scholar] [CrossRef] [PubMed]
- Kay, J.; Thadhani, E.; Samson, L.; Engelward, B. Inflammation-Induced DNA Damage, Mutations and Cancer. DNA Repair 2019, 83, 102673. [Google Scholar] [CrossRef] [PubMed]
- Klebanoff, S.J. Myeloperoxidase. Proc. Assoc. Am. Physicians 1999, 111, 383–389. [Google Scholar] [CrossRef]
- Serhan, C.N. Pro-Resolving Lipid Mediators Are Leads for Resolution Physiology. Nature 2014, 510, 92–101. [Google Scholar] [CrossRef]
- Escalante, T.; Núñez, J.; Moura da Silva, A.M.; Rucavado, A.; Theakston, R.D.G.; Gutiérrez, J.M. Pulmonary Hemorrhage Induced by Jararhagin, a Metalloproteinase from Bothrops Jararaca Snake Venom. Toxicol. Appl. Pharmacol. 2003, 193, 17–28. [Google Scholar] [CrossRef]
- Moura-da-Silva, A.M.; Ramos, O.H.P.; Baldo, C.; Niland, S.; Hansen, U.; Ventura, J.S.; Furlan, S.; Butera, D.; Della-Casa, M.S.; Tanjoni, I.; et al. Collagen Binding Is a Key Factor for the Hemorrhagic Activity of Snake Venom Metalloproteinases. Biochimie 2008, 90, 484–492. [Google Scholar] [CrossRef] [PubMed]
- Kamiguti, A.S.; Hay, C.R.; Theakston, R.D.; Zuzel, M. Insights into the Mechanism of Haemorrhage Caused by Snake Venom Metalloproteinases. Toxicon 1996, 34, 627–642. [Google Scholar] [CrossRef]
- Zuliani, J.P.; Soares, A.M.; Gutiérrez, J.M. Polymorphonuclear neutrophil leukocytes in snakebite envenoming. Toxicon 2020, 187, 188–197. [Google Scholar] [CrossRef]
- Yu, C.-X.; Shi, Z.-A.; Ou, G.-C.; Chen, X.-J.; Liu, Q.; Zeng, D.; Nie, X.-J.; Chen, J.-J. Maresin-2 Alleviates Allergic Airway Inflammation in Mice by Inhibiting the Activation of NLRP3 Inflammasome, Th2 Type Immune Response and Oxidative Stress. Mol. Immunol. 2022, 146, 78–86. [Google Scholar] [CrossRef]
- Silva, G.M.; Berto, D.H.; Lima, C.A.; Waitman, K.B.; Lima, C.F.G.; Prezoto, B.C.; Vieira, M.L.; Rocha, M.M.T.; Gonçalves, L.R.C.; Andrade, S.A. Synergistic Effect of Serine Protease Inhibitors and a Bothropic Antivenom in Reducing Local Hemorrhage and Coagulopathy Caused by Bothrops jararaca Venom. Toxicon 2021, 199, 87–93. [Google Scholar] [CrossRef]
- Fattori, V.; Hohmann, M.S.; Rossaneis, A.C.; Pinho-Ribeiro, F.A.; Verri, W.A. Capsaicin: Current Understanding of Its Mechanisms and Therapy of Pain and Other Pre-Clinical and Clinical Uses. Molecules 2016, 21, 844. [Google Scholar] [CrossRef] [PubMed]
- Verri, W.A.; Cunha, T.M.; Parada, C.A.; Wei, X.Q.; Ferreira, S.H.; Liew, F.Y.; Cunha, F.Q.; Verri, W.A., Jr.; Cunha, T.M.; Parada, C.A.; et al. IL-15 Mediates Immune Inflammatory Hypernociception by Triggering a Sequential Release of IFN-Gamma, Endothelin, and Prostaglandin. Proc. Natl. Acad. Sci. USA 2006, 103, 9721–9725. [Google Scholar] [CrossRef]
- Varela, M.L.; Mogildea, M.; Moreno, I.; Lopes, A. Acute Inflammation and Metabolism. Inflammation 2018, 41, 1115–1127. [Google Scholar] [CrossRef]
- Teixeira, C.F.P.; Chaves, F.; Zamunér, S.R.; Fernandes, C.M.; Zuliani, J.P.; Cruz-Hofling, M.A.; Fernandes, I.; Gutiérrez, J.M. Effects of Neutrophil Depletion in the Local Pathological Alterations and Muscle Regeneration in Mice Injected with Bothrops jararaca Snake Venom. Int. J. Exp. Pathol. 2005, 86, 107–115. [Google Scholar] [CrossRef] [PubMed]
- Cedro, R.C.A.; Menaldo, D.L.; Costa, T.R.; Zoccal, K.F.; Sartim, M.A.; Santos-Filho, N.A.; Faccioli, L.H.; Sampaio, S.V. Cytotoxic and Inflammatory Potential of a Phospholipase A2 from Bothrops jararaca Snake Venom. J. Venom. Anim. Toxins Incl. Trop. Dis. 2018, 24, 33. [Google Scholar] [CrossRef] [PubMed]
- Farsky, S.H.P.; Borelli, P.; Fock, R.A.; Proto, S.Z.; Ferreira, J.M.C.; Melo, S.B. V Chronic Blockade of Nitric Oxide Biosynthesis in Rats: Effect on Leukocyte Endothelial Interaction and on Leukocyte Recruitment. Inflamm. Res. 2004, 53, 442–452. [Google Scholar] [CrossRef]
- Farsky, S.H.; Walber, J.; Costa-Cruz, M.; Cury, Y.; Teixeira, C.F. Leukocyte Response Induced by Bothrops jararaca Crude Venom: In Vivo and in Vitro Studies. Toxicon 1997, 35, 185–193. [Google Scholar] [CrossRef]
- Kolaczkowska, E.; Kubes, P. Neutrophil Recruitment and Function in Health and Inflammation. Nat. Rev. Immunol. 2013, 13, 159–175. [Google Scholar] [CrossRef]
- Sikora, J.P.; Karawani, J.; Sobczak, J. Neutrophils and the Systemic Inflammatory Response Syndrome (SIRS). Int. J. Mol. Sci. 2023, 24, 13469. [Google Scholar] [CrossRef]
- El Kebir, D.; József, L.; Pan, W.; Filep, J. Myeloperoxidase delays neutrophil apoptosis through CD11b/CD18 integrins and prolongs inflammation. Circ. Res. 2008, 103, 352–359. [Google Scholar] [CrossRef]
- Lau, D.; Mollnau, H.; Eiserich, J.P.; Freeman, B.A.; Daiber, A.; Gehling, U.M.; Brümmer, J.; Rudolph, V.; Münzel, T.; Heitzer, T.; et al. Myeloperoxidase Mediates Neutrophil Activation by Association with CD11b/CD18 Integrins. Proc. Natl. Acad. Sci. USA 2005, 102, 431–436. [Google Scholar] [CrossRef] [PubMed]
- Carr, A.C.; Winterbourn, C.C. Oxidation of neutrophil glutathione and protein thiols by myeloperoxidase-derived hypochlorous acid. Biochem. J. 1997, 327, 275–281. [Google Scholar] [CrossRef] [PubMed]
- Rutgers, A.; Heeringa, P.; Giesen, J.E.H.M.; Theunissen, R.T.; Jacobs, H.; Tervaert, J.W.C. Neutrophil Myeloperoxidase Activity and the Influence of Two Single-Nucleotide Promoter Polymorphisms. Br. J. Haematol. 2003, 123, 536–538. [Google Scholar] [CrossRef] [PubMed]
- Park, M.D.; Silvin, A.; Ginhoux, F.; Merad, M. Macrophages in Health and Disease. Cell 2022, 185, 4259–4279. [Google Scholar] [CrossRef]
- Sugiyama, S.; Okada, Y.; Sukhova, G.K.; Virmani, R.; Heinecke, J.W.; Libby, P. Macrophage Myeloperoxidase Regulation by Granulocyte Macrophage Colony-Stimulating Factor in Human Atherosclerosis and Implications in Acute Coronary Syndromes. Am. J. Pathol. 2001, 158, 879–891. [Google Scholar] [CrossRef]
- Aratani, Y. Myeloperoxidase: Its Role for Host Defense, Inflammation, and Neutrophil Function. Arch. Biochem. Biophys. 2018, 640, 47–52. [Google Scholar] [CrossRef]
- Zychar, B.C.; Dale, C.S.; Demarchi, D.S.; Gonçalves, L.R.C. Contribution of Metalloproteases, Serine Proteases and Phospholipases A2 to the Inflammatory Reaction Induced by Bothrops Jararaca Crude Venom in Mice. Toxicon 2010, 55, 227–234. [Google Scholar] [CrossRef]
- Abad Ribeiro, A.B.; Santoro, M.L.; Duarte, M.R.; Virgulino, C.C.; de Oliveira, G.S.S.; França, F.O.D.S. Hemoperitoneum after a Bothrops Snakebite: Case Report. Toxicon 2024, 237, 107350. [Google Scholar] [CrossRef]
- Förstermann, U.; Sessa, W.C. Nitric Oxide Synthases: Regulation and Function. Eur. Heart J. 2012, 33, 829–837, 837a–837d. [Google Scholar] [CrossRef]
- Zamuner, S.R.; Gutiérrez, J.M.; Muscará, M.N.; Teixeira, S.A.; Teixeira, C.F. Bothrops Asper and Bothrops jararaca Snake Venoms Trigger Microbicidal Functions of Peritoneal Leukocytes in Vivo. Toxicon 2001, 39, 1505–1513. [Google Scholar] [CrossRef]
- Serhan, C.N.; Dalli, J.; Karamnov, S.; Choi, A.; Park, C.; Xu, Z.; Ji, R.; Zhu, M.; Petasis, N.A. Macrophage Proresolving Mediator Maresin 1 Stimulates Tissue Regeneration and Controls Pain. FASEB J. 2012, 26, 1755–1765. [Google Scholar] [CrossRef] [PubMed]
- Maximiano, T.K.E.; Carneiro, J.A.; Fattori, V.; Verri, W.A. TRPV1: Receptor Structure, Activation, Modulation and Role in Neuro-Immune Interactions and Pain. Cell Calcium 2024, 119, 102870. [Google Scholar] [CrossRef] [PubMed]
- Almeida, A.S.D.; Bernardes, L.D.B.; Trevisan, G. TRP Channels in Cancer Pain. Eur. J. Pharmacol. 2021, 904, 174185. [Google Scholar] [CrossRef] [PubMed]
- Lourenco-Gonzalez, Y.; Fattori, V.; Domiciano, T.P.; Rossaneis, A.C.; Borghi, S.M.; Zaninelli, T.H.; Bernardy, C.C.F.; Alves-Filho, J.C.; Cunha, T.M.; Cunha, F.Q.; et al. Repurposing of the Nootropic Drug Vinpocetine as an Analgesic and Anti-Inflammatory Agent: Evidence in a Mouse Model of Superoxide Anion-Triggered Inflammation. Mediat. Inflamm. 2019, 2019, 6481812. [Google Scholar] [CrossRef]
- Zamuner, S.R.; Teixeira, C.F.P. Cell Adhesion Molecules Involved in the Leukocyte Recruitment Induced by Venom of the Snake Bothrops Jararaca. Mediat. Inflamm. 2002, 11, 351–357. [Google Scholar] [CrossRef]
- da Silva Fernandes Ribas, A.; de Godoi, K.S.; Sant’Anna, S.S.; da Rocha, M.M.T.; da Silva, W.D. Release of Cytokines in the Peritoneal Fluid of C57BL/6 Mice After Bothrops Jararaca and Bothrops Atrox Venom Injection. Toxins 2025, 17, 164. [Google Scholar] [CrossRef]
- Fattori, V.; Rasquel-Oliveira, F.S.; Artero, N.A.; Ferraz, C.R.; Borghi, S.M.; Casagrande, R.; Verri, W.A. Diosmin Treats Lipopolysaccharide-Induced Inflammatory Pain and Peritonitis by Blocking NF-ΚB Activation in Mice. J. Nat. Prod. 2020, 83, 1018–1026. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | Sense | Antisense | Manufacturer and Prime Number |
|---|---|---|---|
| Gapdh | 5′-GCCCAGAACATCATCCCTGC-3′ | 5′-GCCTCTCTTGCTCAGTGTCC-3′ | Invitrogen Sense: I3932A11 Antisense: 13932A12 |
| Gp91phox (Cybb gene) | 5′-AGCTATGAGGTGGTGATGTTAGTGG-3′ | 5′-CACAATATTTGTACCAGACAGACTTGAG-3′ | Invitrogen Sense: I2203C11 Antisense: I2203C12 |
| iNOS (Nos2 gene) | 5′-CGAAACGCTTCACTTCCAA-3′ | 5′-TGAGCCTATATTGCTGTGGCT-3′ | Invitrogen Sense: I2203B07 Antisense: I2203B08 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Dantas, K.L.S.; Bianchini, B.H.S.; da Silva, M.D.V.; Piva, M.; da Cunha, J.M.; Zanoveli, J.M.; Cardoso, F.C.; Vicentini, F.T.M.C.; Ferraz, C.R.; Clissa, P.B.; et al. Maresin 2, a Specialized Pro-Resolution Lipid Mediator, Reduces Pain and Inflammation Induced by Bothrops jararaca Venom in Mice. Toxins 2025, 17, 367. https://doi.org/10.3390/toxins17080367
Dantas KLS, Bianchini BHS, da Silva MDV, Piva M, da Cunha JM, Zanoveli JM, Cardoso FC, Vicentini FTMC, Ferraz CR, Clissa PB, et al. Maresin 2, a Specialized Pro-Resolution Lipid Mediator, Reduces Pain and Inflammation Induced by Bothrops jararaca Venom in Mice. Toxins. 2025; 17(8):367. https://doi.org/10.3390/toxins17080367
Chicago/Turabian StyleDantas, Kassyo L. S., Beatriz H. S. Bianchini, Matheus D. V. da Silva, Maiara Piva, Joice M. da Cunha, Janaina M. Zanoveli, Fernanda C. Cardoso, Fabiana T. M. C. Vicentini, Camila R. Ferraz, Patricia B. Clissa, and et al. 2025. "Maresin 2, a Specialized Pro-Resolution Lipid Mediator, Reduces Pain and Inflammation Induced by Bothrops jararaca Venom in Mice" Toxins 17, no. 8: 367. https://doi.org/10.3390/toxins17080367
APA StyleDantas, K. L. S., Bianchini, B. H. S., da Silva, M. D. V., Piva, M., da Cunha, J. M., Zanoveli, J. M., Cardoso, F. C., Vicentini, F. T. M. C., Ferraz, C. R., Clissa, P. B., Casagrande, R., & Verri, W. A. (2025). Maresin 2, a Specialized Pro-Resolution Lipid Mediator, Reduces Pain and Inflammation Induced by Bothrops jararaca Venom in Mice. Toxins, 17(8), 367. https://doi.org/10.3390/toxins17080367