Laboratory-Scale Evaluation of a Plant-Based Algaecide for Harmful and Non-Harmful Algae


Abstract
1. Introduction
2. Results
2.1. Exposure Concentrations for the Inhibition of Photosynthesis
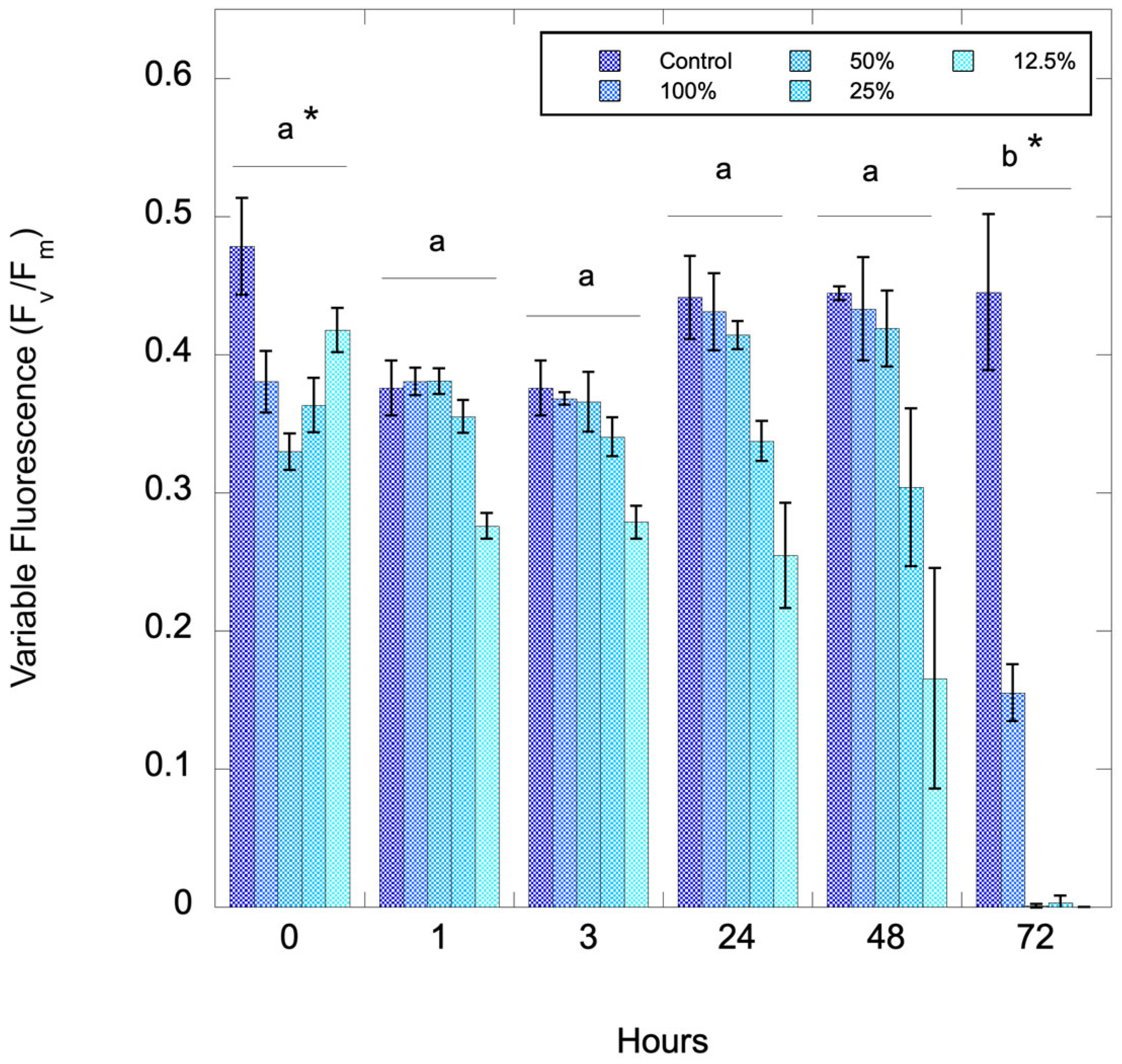
2.2. Reduction in Chlorophyll and Cell Counts
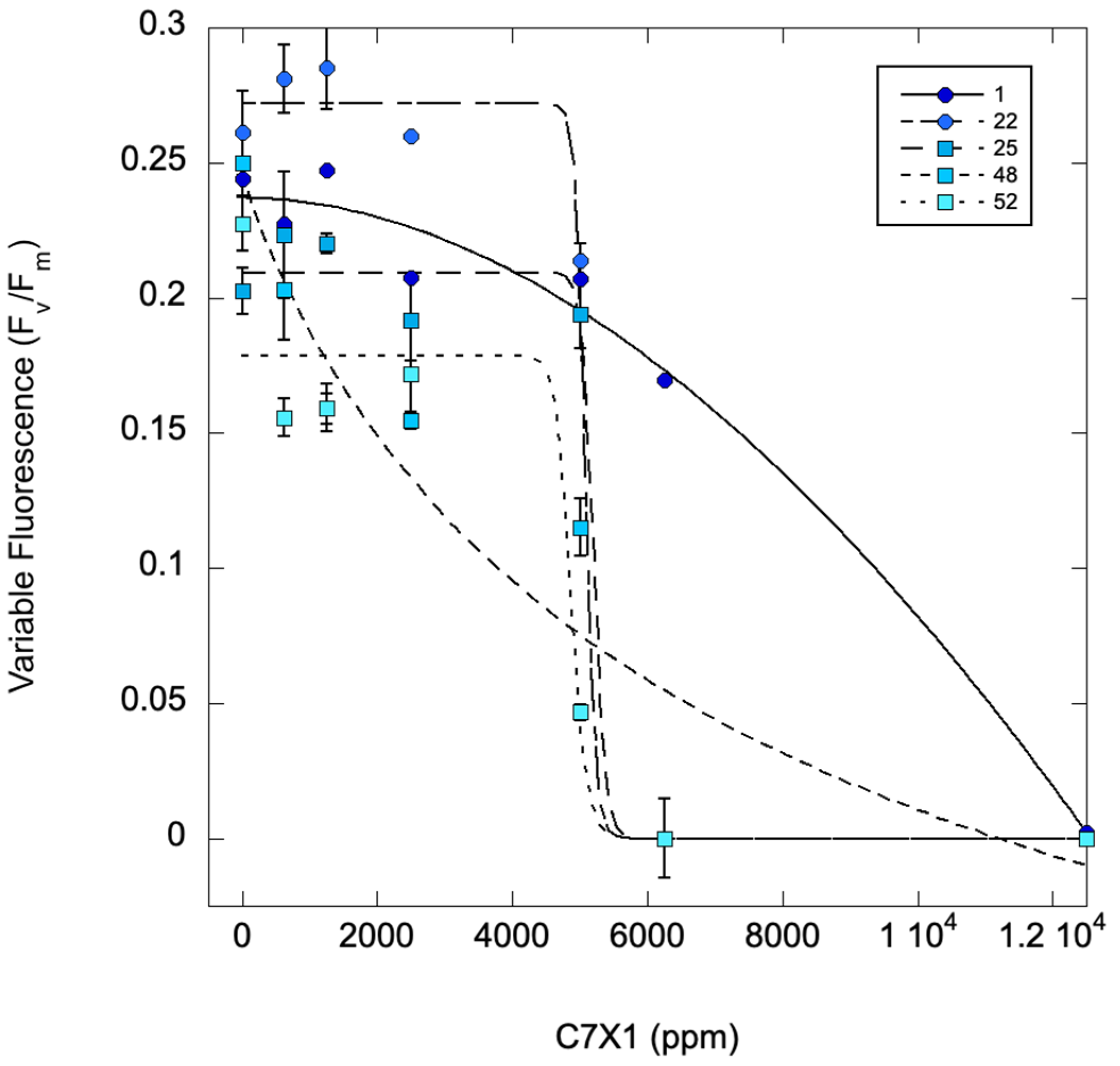
2.3. Effect of Algal Biomass on C71X Efficacy
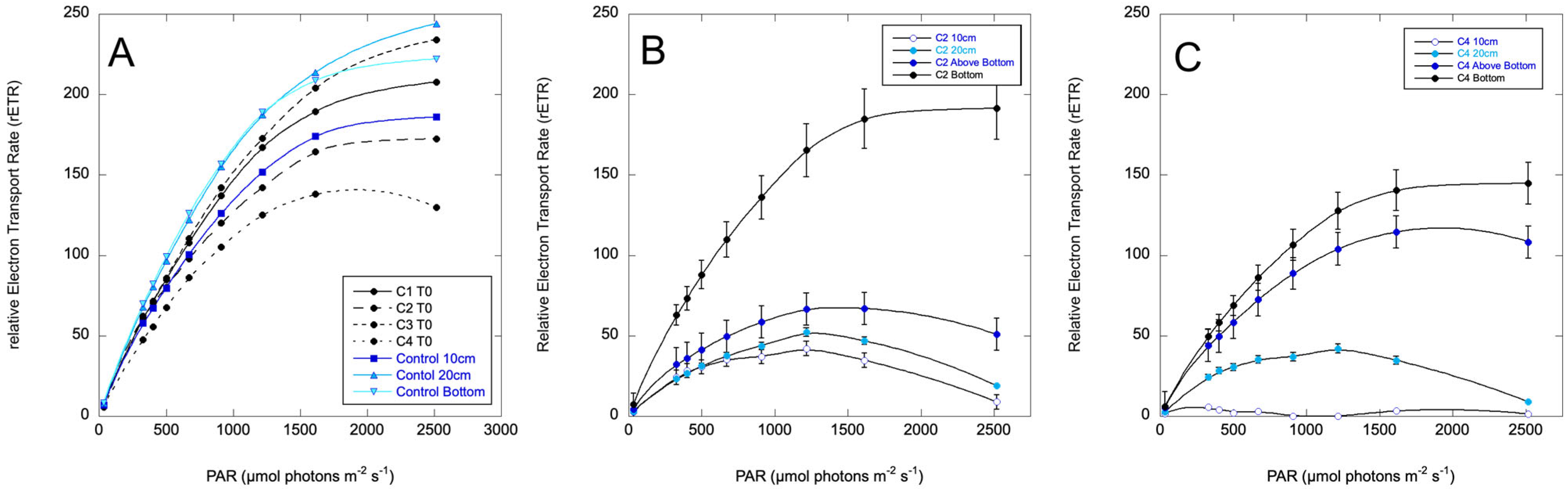
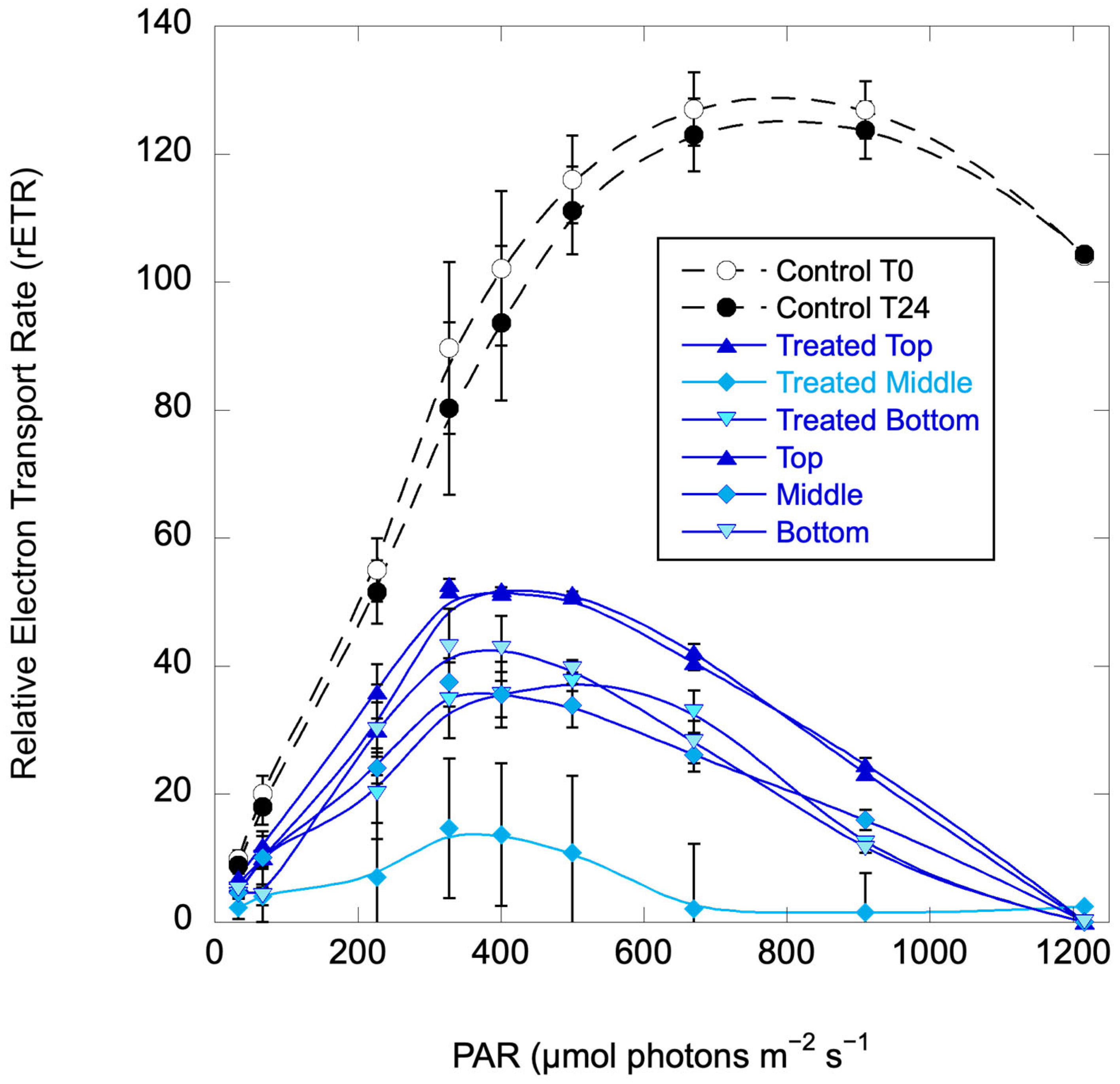
2.4. Vertical Column Experiments
2.4.1. Pinto Lake, 18 August 2024
2.4.2. Pinto Lake, 28 September 2024
2.4.3. Heterosigma akashiwo Cultures
2.4.4. Pinto Lake Large Volume Columns
2.5. Toxin Release
2.6. Stability of the Algaecide
3. Discussion
3.1. Exposure Concentrations for Inhibition of Photosynthesis
3.2. Mode of Action for Inhibition
3.3. Extent of Toxin Release After Exposure to C7X1
3.4. From Bench Scale to Environmental Testing
3.5. Future Directions
4. Conclusions
5. Materials and Methods
5.1. Algaecide Source, Storage, and Degradation
5.2. Algal Strains
5.3. Effects of C7X1 on Chlorophyll and Cell Density
5.4. Field Water Collection
5.5. Experimental Treatments for IC50
5.6. Inhibitory Concentration Curves and Statistics
5.7. Assessment of Variable Fluorescence and Electron Transport Rates
5.8. Pinto Lake Column Experiments
5.9. Hetereosigma akashiwo Column Experiments
5.10. Toxin Testing
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Appendix A
Field Sampling

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Date | Temperature (°C) | Chlorophyll (µg L−1) | Community Composition |
|---|---|---|---|
| 7/31/24 | 24.3 | 900.6 | A—Dolichospermum, R—Microcystis, Aphanizomenon, Straustrum, Cymbella |
| 8/6/24 | 24.7 | 546.9 | A—Microcystis, R—Aphanizomenon, Dolichospermum, Straustrum, diatom (Leptocylindrus-like) |
| 8/13/24 | 24 | 55.8 | A—Microcystis, R—Aphanizomenon, Dolichospermum, Straustrum, Ceratium |
| 8/20/24 | 24.3 | 59.6 | A—Microcystis, R—Aphanizomenon, Dolichospermum, Oscillatoria, Ceratium |
| 8/27/24 | 23.7 | 45.4 | A—Microcystis, R—Aphanizomenon, Dolichospermum, Straustrum |
| 9/3/24 | 23.7 | 78.3 | A—Microcystis, P—Dolichospermum, R—Aphanizomenon, Ceratium |
| 9/17/24 | 22.1 | 77.7 | A—Microcystis, C—Dolichospermum, R—Aphanizomenon, Ceratium |
| 9/24/24 | 22.3 | 85.9 | A—Microcystis, P—Dolichospermum, R—Aphanizomenon, Pediastrum |
| 10/1/24 | 21.9 | 74.9 | A—Microcystis, R—Dolichospermum, Pediastrum |
| 10/15/24 | 21.4 | 120.4 | A—Microcystis, R—Dolichospermum, Oscillatoria, Ceratium, diatom (Leptocylindrus-like) |
| 10/22/24 | 18.9 | 957.1 | A—Microcystis, R—Dolichospermum, Aphanizomenon, Oscillatoria, flagellate |
| 11/5/24 | 16.9 | 2446.3 | A—Microcystis, R—Aphanizomenon, Ceratium |
| 11/12/24 | 15.9 | 188.2 | A—Microcystis |
| 11/25/24 | 14.4 | 174.0 | A—Microcystis, R—Ceratium |
| 12/3/24 | 12.8 | 1340.3 | A—Microcystis, R—Aphanizomenon |
| 12/10/24 | 11.7 | 372.4 | A—Microcystis, R—Aphanizomenon |
| 12/17/24 | 12.2 | 1360.4 | A—Microcystis, R—Aphanizomenon |
| 1/7/25 | 12 | 12.6 | C—Aphanizomenon, Microcystis, R—Ceratium, flagellate |
| 1/14/25 | 10.6 | 6.6 | C—Aphanizomenon, Microcystis, R—Ceratium, flagellate, tons of sparkles/bacteria |
| 1/21/25 | 10.3 | 5.5 | C—Aphanizomenon, Microcystis, R—Ceratium |
| 2/5/25 | 12.9 | 3.6 | A—Microcystis, P—Aphanizomenon, R—flagellate |
| 2/11/25 | 11.3 | 2.1 | A—Microcystis, R—flagellate |
| 2/18/25 | 12.5 | 1.7 | A—Microcystis, R—Ceratium |
| 2/25/25 | 14.8 | 2.1 | C—Microcystis, Ceratium, Coelastrum, tons of sparkles/bacteria |
| 3/4/25 | 14.3 | 2.8 | C—Microcystis, Ceratium, diatom (Leptocylindrus-like), tons of sparkles/bacteria, |
| 3/18/25 | 14.5 | 6.7 | A—Microcystis, R—Ceratium |
| 3/25/25 | 18.5 | 6.9 | C—Microcystis, Aphanizomenon, flagellate, R—Ceratium, Coelastrum, diatom (Leptocylindrus-like) |
References
- Feng, L.; Wang, Y.; Hou, X.; Qin, B.; Kutser, T.; Qu, F.; Chen, N.; Paerl, H.W.; Zheng, C. Harmful Algal Blooms in Inland Waters. Nat. Rev. Earth Environ. 2024, 5, 631–644. [Google Scholar] [CrossRef] [PubMed]
- Anderson, D.M.; Fensin, E.; Gobler, C.J.; Hoeglund, A.E.; Hubbard, K.A.; Kulis, D.M.; Landsberg, J.H.; Lefebvre, K.A.; Provoost, P.; Richlen, M.L.; et al. Marine Harmful Algal Blooms (HABs) in the United States: History, Current Status and Future Trends. Harmful Algae 2021, 102, 101975. [Google Scholar] [CrossRef]
- Enevoldsen, H.O.; Isensee, K.; Lee, Y.J. State of the Ocean Report 2024; UNESCO-IOC: Paris, France, 2024. [Google Scholar]
- Woods Hole Oceanographic Institution; Anderson, D.M.; Backer, L.C.; Bouma-Gregson, K.; Bowers, H.A.; Bricelj, V.M.; D’Anglada, L.; Deeds, J.; Dortch, Q.; Doucette, G.J.; et al. Harmful Algal Research & Response: A National Environmental Science Strategy (HARRNESS), 2024–2034; Woods Hole Oceanographic Institution: Woods Hole, MA, USA, 2024. [Google Scholar]
- Trainer, V.L.; Bates, S.S.; Lundholm, N.; Thessen, A.E.; Cochlan, W.P.; Adams, N.G.; Trick, C.G. Pseudo-Nitzschia Physiological Ecology, Phylogeny, Toxicity, Monitoring and Impacts on Ecosystem Health. Harmful Algae 2012, 14, 271–300. [Google Scholar] [CrossRef]
- Harke, M.J.; Steffen, M.M.; Gobler, C.J.; Otten, T.G.; Wilhelm, S.W.; Wood, S.A.; Paerl, H.W. A Review of the Global Ecology, Genomics, and Biogeography of the Toxic Cyanobacterium, Microcystis spp. Harmful Algae 2016, 54, 4–20. [Google Scholar] [CrossRef] [PubMed]
- Mehdizadeh Allaf, M. Heterosigma akashiwo, a Fish-Killing Flagellate. Microbiol. Res. 2023, 14, 132–147. [Google Scholar] [CrossRef]
- Shikata, T.; Yuasa, K.; Kitatsuji, S.; Sakamoto, S.; Akita, K.; Fujinami, Y.; Nishiyama, Y.; Kotake, T.; Tanaka, R.; Yamasaki, Y. Superoxide Production by the Red Tide-Producing Chattonella marina Complex (Raphidophyceae) Correlates with Toxicity to Aquacultured Fishes. Antioxidants 2021, 10, 1635. [Google Scholar] [CrossRef]
- Agnihotri, V.K. Anabaena Flos-Aquae. Crit. Rev. Environ. Sci. Technol. 2014, 44, 1995–2037. [Google Scholar] [CrossRef]
- Ibelings, B.W.; Chorus, I. Accumulation of Cyanobacterial Toxins in Freshwater “Seafood” and Its Consequences for Public Health: A Review. Environ. Pollut. 2007, 150, 177–192. [Google Scholar] [CrossRef]
- Peacock, M.B.; Gibble, C.M.; Senn, D.B.; Cloern, J.E.; Kudela, R.M. Blurred Lines: Multiple Freshwater and Marine Algal Toxins at the Land-Sea Interface of San Francisco Bay, California. Harmful Algae 2018, 73, 138–147. [Google Scholar] [CrossRef]
- Miller, M.A.; Kudela, R.M.; Mekebri, A.; Crane, D.; Oates, S.C.; Tinker, M.T.; Staedler, M.; Miller, W.A.; Toy-Choutka, S.; Dominik, C.; et al. Evidence for a Novel Marine Harmful Algal Bloom: Cyanotoxin (Microcystin) Transfer from Land to Sea Otters. PLoS ONE 2010, 5, e12576. [Google Scholar] [CrossRef]
- Gibble, C.M.; Peacock, M.B.; Kudela, R.M. Evidence of Freshwater Algal Toxins in Marine Shellfish: Implications for Human and Aquatic Health. Harmful Algae 2016, 59, 59–66. [Google Scholar] [CrossRef] [PubMed]
- Shahmohamadloo, R.S.; Frenken, T.; Rudman, S.M.; Ibelings, B.W.; Trainer, V.L. Diseases and disorders in fish due to harmful algal blooms. In Climate Change on Diseases and Disorders of Finfish in Cage Culture, 3rd ed.; Woo, P.T.K., Subasinghe, R.P., Eds.; CABI: Dorset, UK, 2023; pp. 387–429. ISBN 978-1-80062-162-6. [Google Scholar]
- Glibert, P.M.; Al-Azri, A.; Icarus Allen, J.; Bouwman, A.F.; Beusen, A.H.W.; Burford, M.A.; Harrison, P.J.; Zhou, M. Key Questions and Recent Research Advances on Harmful Algal Blooms in Relation to Nutrients and Eutrophication. In Global Ecology and Oceanography of Harmful Algal Blooms; Glibert, P.M., Berdalet, E., Burford, M.A., Pitcher, G.C., Zhou, M., Eds.; Ecological Studies; Springer International Publishing: Cham, Switzerland, 2018; Volume 232, pp. 229–259. ISBN 978-3-319-70068-7. [Google Scholar]
- Paerl, H.W.; Otten, T.G.; Kudela, R. Mitigating the Expansion of Harmful Algal Blooms Across the Freshwater-to-Marine Continuum. Environ. Sci. Technol. 2018, 52, 5519–5529. [Google Scholar] [CrossRef]
- Kinley-Baird, C.; Calomeni, A.; Berthold, D.E.; Lefler, F.W.; Barbosa, M.; Rodgers, J.H.; Laughinghouse, H.D. Laboratory-Scale Evaluation of Algaecide Effectiveness for Control of Microcystin-Producing Cyanobacteria from Lake Okeechobee, Florida (USA). Ecotoxicol. Environ. Saf. 2021, 207, 111233. [Google Scholar] [CrossRef] [PubMed]
- Hennessey, A.V.; McDonald, M.B.; Johnson, P.P.; Gladfelter, M.F.; Merrill, K.L.; Tenison, S.E.; Ganegoda, S.S.; Hoang, T.C.; Torbert, H.A.; Beck, B.H.; et al. Evaluating the Tolerance of Harmful Algal Bloom Communities to Copper. Environ. Pollut. 2025, 368, 125691. [Google Scholar] [CrossRef]
- Moreno-Andrés, J.; Romero-Martínez, L.; Seoane, S.; Acevedo-Merino, A.; Moreno-Garrido, I.; Nebot, E. Evaluation of Algaecide Effectiveness of Five Different Oxidants Applied on Harmful Phytoplankton. J. Hazard. Mater. 2023, 452, 131279. [Google Scholar] [CrossRef] [PubMed]
- Kiaune, L.; Singhasemanon, N. Pesticidal Copper (I) Oxide: Environmental Fate and Aquatic Toxicity. In Reviews of Environmental Contamination and Toxicology Volume 213; Whitacre, D.M., Ed.; Reviews of Environmental Contamination and Toxicology; Springer New York: New York, NY, USA, 2011; Volume 213, pp. 1–26. ISBN 978-1-4419-9859-0. [Google Scholar]
- Geer, T.D.; Kinley, C.M.; Iwinski, K.J.; Calomeni, A.J.; Rodgers, J.H. Comparative Toxicity of Sodium Carbonate Peroxyhydrate to Freshwater Organisms. Ecotoxicol. Environ. Saf. 2016, 132, 202–211. [Google Scholar] [CrossRef]
- Calomeni, A.J.; Iwinski, K.J.; Kinley, C.M.; McQueen, A.; Rodgers, J.H. Responses of Lyngbya wollei to Algaecide Exposures and a Risk Characterization Associated with Their Use. Ecotoxicol. Environ. Saf. 2015, 116, 90–98. [Google Scholar] [CrossRef]
- Li, H.; Xing, R.; Ji, X.; Liu, Y.; Chu, X.; Gu, J.; Wang, S.; Wang, G.; Zhao, S.; Cao, X. Natural Algicidal Compounds: Strategies for Controlling Harmful Algae and Application. Plant Physiol. Biochem. 2024, 215, 108981. [Google Scholar] [CrossRef]
- Coyne, K.J.; Wang, Y.; Johnson, G. Algicidal Bacteria: A Review of Current Knowledge and Applications to Control Harmful Algal Blooms. Front. Microbiol. 2022, 13, 871177. [Google Scholar] [CrossRef]
- Yang, S.; Williams, S.J.; Courtney, M.; Burchill, L. Warfare under the Waves: A Review of Bacteria-Derived Algaecidal Natural Products. Nat. Prod. Rep. 2025, 4, 641–756. [Google Scholar] [CrossRef]
- Raveau, R.; Fontaine, J.; Lounès-Hadj Sahraoui, A. Essential Oils as Potential Alternative Biocontrol Products against Plant Pathogens and Weeds: A Review. Foods 2020, 9, 365. [Google Scholar] [CrossRef] [PubMed]
- Chen, S.; Zheng, T.; Ye, C.; Huannixi, W.; Yakefu, Z.; Meng, Y.; Peng, X.; Tian, Z.; Wang, J.; Ma, Y.; et al. Algicidal Properties of Extracts from Cinnamomum camphora Fresh Leaves and Their Main Compounds. Ecotoxicol. Environ. Saf. 2018, 163, 594–603. [Google Scholar] [CrossRef]
- Liu, H.; Huang, J.; Yang, S.; Li, J.; Zhou, L. Chemical Composition, Algicidal, Antimicrobial, and Antioxidant Activities of the Essential Oils of Taiwania flousiana Gaussen. Molecules 2020, 25, 967. [Google Scholar] [CrossRef] [PubMed]
- Bácsi, I.; Gonda, S.; Nemes-Kókai, Z.; B-Béres, V.; Vasas, G. Horseradish Essential Oil as a Promising Anti-Algal Product for Prevention of Phytoplankton Proliferation and Biofouling. Plants 2021, 10, 1550. [Google Scholar] [CrossRef]
- Wang, Y.; Wang, D.; Zhao, W.; Liu, H.; Li, L.; Bai, J. Inhibitory Effect and Mechanism of a Compound Essential Oils on Cladophora Glomerata. Mar. Pollut. Bull. 2023, 188, 114668. [Google Scholar] [CrossRef]
- Liu, F.; He, Z.-B.; Li, H.-Y.; Liu, J.-S.; Yang, W.-D. Inhibition of Five Natural Products from Chinese Herbs on the Growth of Chattonella Marina. Environ. Sci. Pollut. Res. 2016, 23, 17793–17800. [Google Scholar] [CrossRef] [PubMed]
- Zhou, S.; Shao, Y.; Gao, N.; Deng, Y.; Qiao, J.; Ou, H.; Deng, J. Effects of Different Algaecides on the Photosynthetic Capacity, Cell Integrity and Microcystin-LR Release of Microcystis aeruginosa. Sci. Total Environ. 2013, 463–464, 111–119. [Google Scholar] [CrossRef]
- Zhao, G.; Hong, Y.; Li, L.; Zhang, H.; Xu, R.; Hao, Y. Selection and Characterization of Plant-Derived Alkaloids with Strong Antialgal Inhibition: Growth Inhibition Selectivity and Inhibitory Mechanism. Harmful Algae 2022, 117, 102272. [Google Scholar] [CrossRef]
- Zhu, X.; Dao, G.; Tao, Y.; Zhan, X.; Hu, H. A Review on Control of Harmful Algal Blooms by Plant-Derived Allelochemicals. J. Hazard. Mater. 2021, 401, 123403. [Google Scholar] [CrossRef]
- Lan, Y.; Chen, Q.; Gou, T.; Sun, K.; Zhang, J.; Sun, D.; Duan, S. Algicidal Activity of Cyperus rotundus Aqueous Extracts Reflected by Photosynthetic Efficiency and Cell Integrity of Harmful Algae Phaeocystis Globosa. Water 2020, 12, 3256. [Google Scholar] [CrossRef]
- Hu, J.; Kokoette, E.; Xu, C.; Huang, S.; Tang, T.; Zhang, Y.; Liu, M.; Huang, Y.; Yu, S.; Zhu, J.; et al. Natural Algaecide Sphingosines Identified in Hybrid Straw Decomposition Driven by White-Rot Fungi. Adv. Sci. 2023, 10, 2300569. [Google Scholar] [CrossRef] [PubMed]
- Zhou, L.H.; Zheng, T.L.; Wang, X.; Ye, J.L.; Tian, Y.; Hong, H.S. Effect of Five Chinese Traditional Medicines on the Biological Activity of a Red-Tide Causing Alga—Alexandrium tamarense. Harmful Algae 2007, 6, 354–360. [Google Scholar] [CrossRef]
- Yang, W.-D.; Liu, J.-S.; Li, H.-Y.; Zhang, X.-L.; Qi, Y.-Z. Inhibition of the Growth of Alexandrium tamarense by Algicidal Substances in Chinese Fir (Cunninghamia lanceolata). Bull. Environ. Contam. Toxicol. 2009, 83, 537–541. [Google Scholar] [CrossRef] [PubMed]
- Zhou, L.H.; Zheng, T.L.; Chen, X.H.; Wang, X.; Chen, S.B.; Tian, Y.; Hong, H.S. The Inhibitory Effects of Garlic (Allium sativum) and Diallyl Trisulfide on Alexandrium tamarense and Other Harmful Algal Species. J. Appl. Phycol. 2008, 20, 349–358. [Google Scholar] [CrossRef]
- Barani, M.; Yousefzadi, M.; Moezi, M. Essential Oils, New Source of Algicidal Compounds. J. Appl. Phycol. 2015, 27, 267–273. [Google Scholar] [CrossRef]
- Wang, H.Q.; Liang, F.; Qiao, N.; Dong, J.-X.; Zhang, L.-Y.; Guo, Y.-F. Chemical Composition of Volatile Oil from Two Emergent Plants and Their Algae Inhibition Activity. Pol. J. Environ. Stud. 2014, 23, 2371–2374. [Google Scholar]
- Anantapantula, S.S.; Wilson, A.E. Most Treatments to Control Freshwater Algal Blooms Are Not Effective: Meta-Analysis of Field Experiments. Water Res. 2023, 243, 120342. [Google Scholar] [CrossRef] [PubMed]
- Kudela, R.M.; Palacios, S.L.; Austerberry, D.C.; Accorsi, E.K.; Guild, L.S.; Torres-Perez, J. Application of Hyperspectral Remote Sensing to Cyanobacterial Blooms in Inland Waters. Remote Sens. Environ. 2015, 167, 196–205. [Google Scholar] [CrossRef]
- Kurobe, T.; Baxa, D.V.; Mioni, C.E.; Kudela, R.M.; Smythe, T.R.; Waller, S.; Chapman, A.D.; Teh, S.J. Identification of Harmful Cyanobacteria in the Sacramento-San Joaquin Delta and Clear Lake, California by DNA Barcoding. SpringerPlus 2013, 2, 491. [Google Scholar] [CrossRef]
- Kudela, R.; Howard, M.; Monismith, S.; Paerl, H. Status, Trends, and Drivers of Harmful Algal Blooms Along the Freshwater-to-Marine Gradient in the San Francisco Bay–Delta System. SFEWS 2023, 20, 1–51. [Google Scholar] [CrossRef]
- Kudela, R.M.; Senn, D.B.; Richardson, E.T.; Bouma-Gregson, K.; Bergamaschi, B.A.; Sim, L. Evaluation and Refinement of Chlorophyll-a Algorithms for High-Biomass Blooms in San Francisco Bay (USA). Remote Sens. 2024, 16, 1103. [Google Scholar] [CrossRef]
- Jeong, S.; Yang, D.; Joo, S.; Park, S. Allelopathic Inhibition Effects of Myriophyllum spicatum on Growths of Bloom-Forming Cyanobacteria and Other Phytoplankton Species in Coexistence Experiments. J. Plant Biol. 2021, 64, 501–510. [Google Scholar] [CrossRef]
- Wagner, N.D.; Osburn, F.S.; Wang, J.; Taylor, R.B.; Boedecker, A.R.; Chambliss, C.K.; Brooks, B.W.; Scott, J.T. Biological Stoichiometry Regulates Toxin Production in Microcystis aeruginosa (UTEX 2385). Toxins 2019, 11, 601. [Google Scholar] [CrossRef]
- Perez, J.L.; Chu, T. Effect of Zinc on Microcystis aeruginosa UTEX LB 2385 and Its Toxin Production. Toxins 2020, 12, 92. [Google Scholar] [CrossRef]
- Fredrickson, K.A.; Strom, S.L.; Crim, R.; Coyne, K.J. Interstrain Variability in Physiology and Genetics of Heterosigma Akashiwo (Raphidophyceae) from the West Coast of North America. J. Phycol. 2011, 47, 25–35. [Google Scholar] [CrossRef] [PubMed]
- Dick, G.J.; Duhaime, M.B.; Evans, J.T.; Errera, R.M.; Godwin, C.M.; Kharbush, J.J.; Nitschky, H.S.; Powers, M.A.; Vanderploeg, H.A.; Schmidt, K.C.; et al. The Genetic and Ecophysiological Diversity of Microcystis. Environ. Microbiol. 2021, 23, 7278–7313. [Google Scholar] [CrossRef]
- Xiao, M.; Li, M.; Reynolds, C.S. Colony Formation in the Cyanobacterium Microcystis. Biol. Rev. 2018, 93, 1399–1420. [Google Scholar] [CrossRef]
- Burkholder, J.; Glibert, P. Intraspecific Variability: An Important Consideration in Forming Generalisations about Toxigenic Algal Species. Afr. J. Mar. Sci. 2006, 28, 177–180. [Google Scholar] [CrossRef]
- Kudela, R.M. Characterization and Deployment of Solid Phase Adsorption Toxin Tracking (SPATT) Resin for Monitoring of Microcystins in Fresh and Saltwater. Harmful Algae 2011, 11, 117–125. [Google Scholar] [CrossRef]
- Hossain, I.; Mays, B.; Hanhart, S.L.; Hubble, J.; Azizihariri, P.; McLean, T.I.; Pierce, R.; Lovko, V.; John, V.T. An Effective Algaecide for the Targeted Destruction of Karenia brevis. Harmful Algae 2024, 138, 102707. [Google Scholar] [CrossRef]
- Guo, P.; Liu, Y.; Liu, C. Effects of Chitosan, Gallic Acid, and Algicide on the Physiological and Biochemical Properties of Microcystis Flos-Aquae. Environ. Sci. Pollut. Res. 2015, 22, 13514–13521. [Google Scholar] [CrossRef] [PubMed]
- Mehdizadeh Allaf, M.; Erratt, K.J.; Peerhossaini, H. Comparative Assessment of Algaecide Performance on Freshwater Phytoplankton: Understanding Differential Sensitivities to Frame Cyanobacteria Management. Water Res. 2023, 234, 119811. [Google Scholar] [CrossRef] [PubMed]
- Lürling, M.; Meng, D.; Faassen, E. Effects of Hydrogen Peroxide and Ultrasound on Biomass Reduction and Toxin Release in the Cyanobacterium, Microcystis aeruginosa. Toxins 2014, 6, 3260–3280. [Google Scholar] [CrossRef] [PubMed]
- Ralph, P.J.; Gademann, R. Rapid Light Curves: A Powerful Tool to Assess Photosynthetic Activity. Aquat. Bot. 2005, 82, 222–237. [Google Scholar] [CrossRef]
- Nishiyama, Y. Oxidative Stress Inhibits the Repair of Photodamage to the Photosynthetic Machinery. EMBO J. 2001, 20, 5587–5594. [Google Scholar] [CrossRef]
- Bittencourt-Oliveira, M.D.C.; Cordeiro-Araújo, M.K.; Chia, M.A.; Arruda-Neto, J.D.D.T.; Oliveira, Ê.T.D.; Santos, F.D. Lettuce Irrigated with Contaminated Water: Photosynthetic Effects, Antioxidative Response and Bioaccumulation of Microcystin Congeners. Ecotoxicol. Environ. Saf. 2016, 128, 83–90. [Google Scholar] [CrossRef]
- Cordeiro-Araújo, M.K.; Chia, M.A.; Arruda-Neto, J.D.D.T.; Tornisielo, V.L.; Vilca, F.Z.; Bittencourt-Oliveira, M.D.C. Microcystin-LR Bioaccumulation and Depuration Kinetics in Lettuce and Arugula: Human Health Risk Assessment. Sci. Total Environ. 2016, 566–567, 1379–1386. [Google Scholar] [CrossRef]
- Melaram, R.; Newton, A.R.; Chafin, J. Microcystin Contamination and Toxicity: Implications for Agriculture and Public Health. Toxins 2022, 14, 350. [Google Scholar] [CrossRef]
- Chorus, I.; Welker, M. Toxic Cyanobacteria in Water: A Guide to Their Public Health Consequences, Monitoring and Management, 2nd ed.; CRC Press: London, UK, 2021; ISBN 978-1-00-308144-9. [Google Scholar]
- Trapp, A.; Hayashi, K.; Fiechter, J.; Kudela, R.M. What Happens in the Shadows—Influence of Seasonal and Non-Seasonal Dynamics on Domoic Acid Monitoring in the Monterey Bay Upwelling Shadow. Harmful Algae 2023, 129, 102522. [Google Scholar] [CrossRef]
- Greenfield, D.I.; Duquette, A.; Goodson, A.; Keppler, C.J.; Williams, S.H.; Brock, L.M.; Stackley, K.D.; White, D.; Wilde, S.B. The Effects of Three Chemical Algaecides on Cell Numbers and Toxin Content of the Cyanobacteria Microcystis aeruginosa and Anabaenopsis sp. Environ. Manag. 2014, 54, 1110–1120. [Google Scholar] [CrossRef]
- Shahmohamadloo, R.S.; Rudman, S.M.; Clare, C.I.; Westrick, J.A.; Wang, X.; De Meester, L.; Fryxell, J.M. Intraspecific Diversity Is Critical to Population-Level Risk Assessments. Sci. Rep. 2024, 14, 25883. [Google Scholar] [CrossRef] [PubMed]
- Wei, D.; Kisuno, A.; Kameya, T.; Urano, K. A New Method for Evaluating Biological Safety of Environmental Water with Algae, Daphnia and Fish Toxicity Ranks. Sci. Total Environ. 2006, 371, 383–390. [Google Scholar] [CrossRef]
- Sengco, M.R. Prevention and Control of Karenia brevis Blooms. Harmful Algae 2009, 8, 623–628. [Google Scholar] [CrossRef]
- Cloern, J.E.; Schraga, T.S.; Lopez, C.B.; Knowles, N.; Grover Labiosa, R.; Dugdale, R. Climate Anomalies Generate an Exceptional Dinoflagellate Bloom in San Francisco Bay. Geophys. Res. Lett. 2005, 32, 2005GL023321. [Google Scholar] [CrossRef]
- Welschmeyer, N.A. Fluorometric Analysis of Chlorophyll a in the Presence of Chlorophyll b and Pheopigments. Limnol. Oceanogr. 1994, 39, 1985–1992. [Google Scholar] [CrossRef]
- Santabarbara, S.; Villafiorita Monteleone, F.; Remelli, W.; Rizzo, F.; Menin, B.; Casazza, A.P. Comparative Excitation-emission Dependence of the FV / FM Ratio in Model Green Algae and Cyanobacterial Strains. Physiol. Plant. 2019, 166, 351–364. [Google Scholar] [CrossRef]
- Schreiber, U. Pulse-Amplitude-Modulation (PAM) Fluorometry and Saturation Pulse Method: An Overview. In Chlorophyll a Fluorescence; Papageorgiou, G.C., Govindjee, Eds.; Advances in Photosynthesis and Respiration; Springer: Dordrecht, The Netherlands, 2004; Volume 19, pp. 279–319. ISBN 978-1-4020-3217-2. [Google Scholar]
- Platt, T.T.; Gallegos, C.; Harrison, W. Photoinhibition of Photosynthesis in Natural Assemblages of Marine Phytoplankton. J. Mar. Res. 1980, 38, 687–701. [Google Scholar]






| Ingredient Name | CAS | Percent (%) |
|---|---|---|
| Peppermint (A) | N/A | 0.1 |
| Lemongrass Oil (A) | 8007-02-1 | 1.0 |
| Cinnamon (A) | N/A | 2.0 |
| Cottonseed Oil (A) | 8001-29-4 | 2.0 |
| Geranium Oil (A) | 8000-46-2 | 2.0 |
| Thyme (A) | N/A | 2.0 |
| Rosemary (A) | N/A | 3.0 |
| Clove (A) | N/A | 4.0 |
| Garlic (A) | N/A | 4.0 |
| Total Active Ingredients: | 20.1 | |
| Citrus Pectin (I) | 9000-69-5 | |
| Citrus Peel Extract (I) | 94226-47-4 | |
| Guar Gum (I) | 9000-30-0 | |
| Isopropyl Alcohol (I) | 67-63-0 | |
| Water (I) | N/A | |
| Xanthan Gum (I) | 11138-66-2 | |
| Total Inactive Ingredients: | 79.1 |
| Algal Strain | IC50 (SE) | Fv/Fm < 0.05 |
|---|---|---|
| Anabaena flos-aquae | 13,247 (6962) | 20,000 |
| Gloeocapsa alpicola | 2015 (292) | 10,000 |
| Microcystis aeruginosa | 976 (96) | 6000 |
| Chattonella marina | 393 (135) | 10,000 |
| Chattonella subsalsa | 713 (140) | 10,000 |
| Heterosigma akashiwo | 1618 (835) | 5000 |
| Isochrysis galbana | 3456 (571) | 10,000 |
| Pseudo-nitzschia multiseries | 4073 (748) | 10,000 |
| Algal Strain (Class) | Strain/ID | Source | Temperature (°C) | Salinity (PSU) | Growth Rate (d−1) |
|---|---|---|---|---|---|
| Anabaena flos-aquae (Cyanophycae) | B 1444 | UTEX | ~20 | 0 | 0.173 (0.084) |
| Gloeocapsa alpicola (Cyanophycae) | 589 | UTEX | ~20 | 0 | 0.172 (0.02) |
| Microcystis aeruginosa (Cyanophycae) | LB 2385 | UTEX | ~20 | 0 | 0.072 (0.002) |
| Chattonella marina (Raphidophyceae) | CCMP 2049 | NCMA | 20 | 32 | 0.229 (0.025) |
| Chattonella subsalsa (Raphidophyceae) | CCMP 2821 | NCMA | 20 | 32 | 0.433 (0.029) |
| Heterosigma akashiwo (Raphidophycae) | EB L71 | EBL | 15 | 24 | 0.192 (0.051) |
| Isochrysis galbana (Coccolithophyceae) | CCMP 1323 | NCMA | 15 | 32 | 0.314 (0.003) |
| Pseudo-nitzschia multiseries (Bacillariophyceae) | EBL 123 | EBL | 15 | 32 | 0.285 (0.028) |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the author. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kudela, R.M. Laboratory-Scale Evaluation of a Plant-Based Algaecide for Harmful and Non-Harmful Algae. Toxins 2025, 17, 270. https://doi.org/10.3390/toxins17060270
Kudela RM. Laboratory-Scale Evaluation of a Plant-Based Algaecide for Harmful and Non-Harmful Algae. Toxins. 2025; 17(6):270. https://doi.org/10.3390/toxins17060270
Chicago/Turabian StyleKudela, Raphael M. 2025. "Laboratory-Scale Evaluation of a Plant-Based Algaecide for Harmful and Non-Harmful Algae" Toxins 17, no. 6: 270. https://doi.org/10.3390/toxins17060270
APA StyleKudela, R. M. (2025). Laboratory-Scale Evaluation of a Plant-Based Algaecide for Harmful and Non-Harmful Algae. Toxins, 17(6), 270. https://doi.org/10.3390/toxins17060270