Integration of Multi-Omics, Histological, and Biochemical Analysis Reveals the Toxic Responses of Nile Tilapia Liver to Chronic Microcystin-LR Exposure

 , ,
, ,  ,
, 
Abstract
1. Introduction
2. Results
2.1. Biochemical Parameters of the Serum and Liver
2.2. Hepatic Histological Characteristics
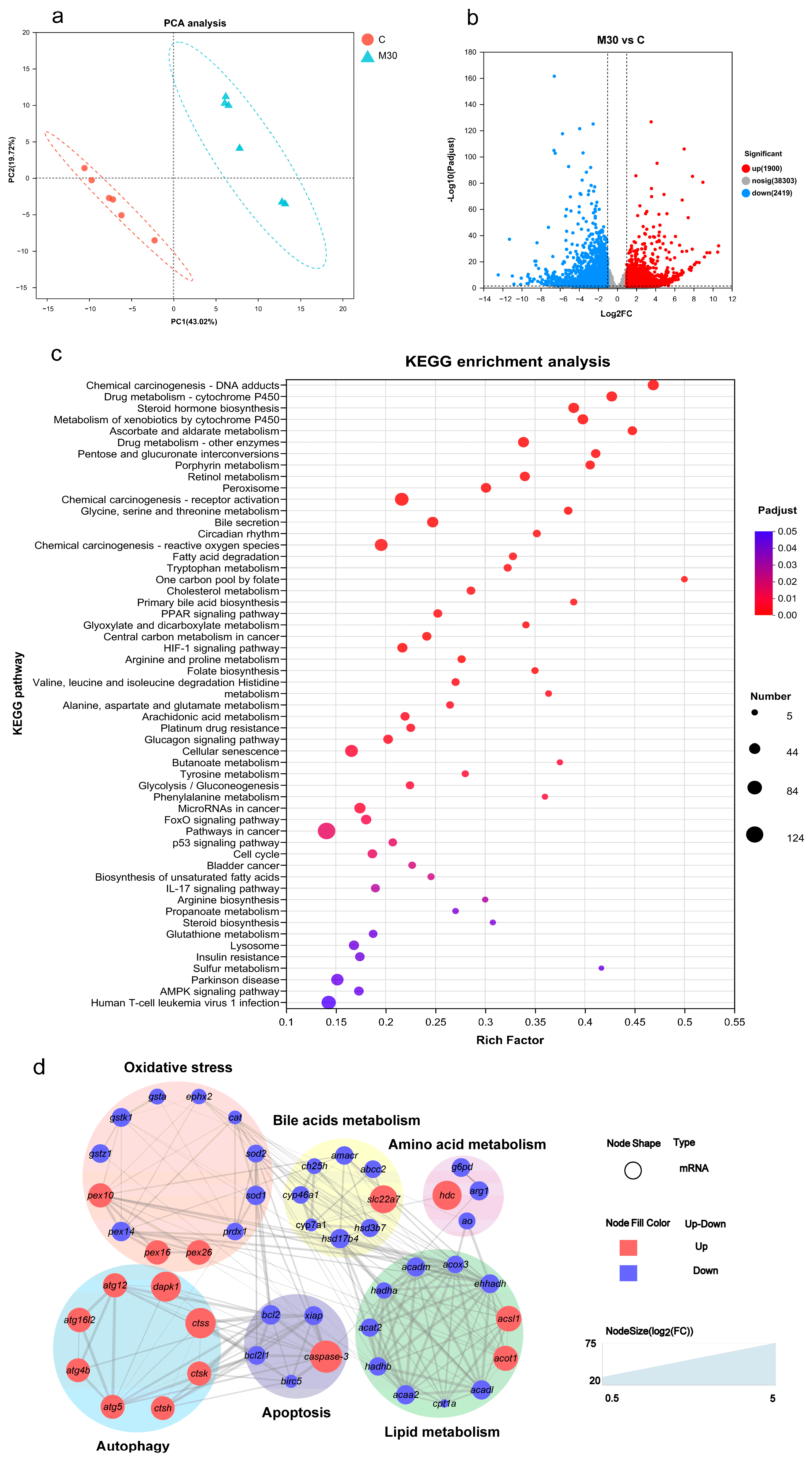
2.3. Liver Transcriptomics
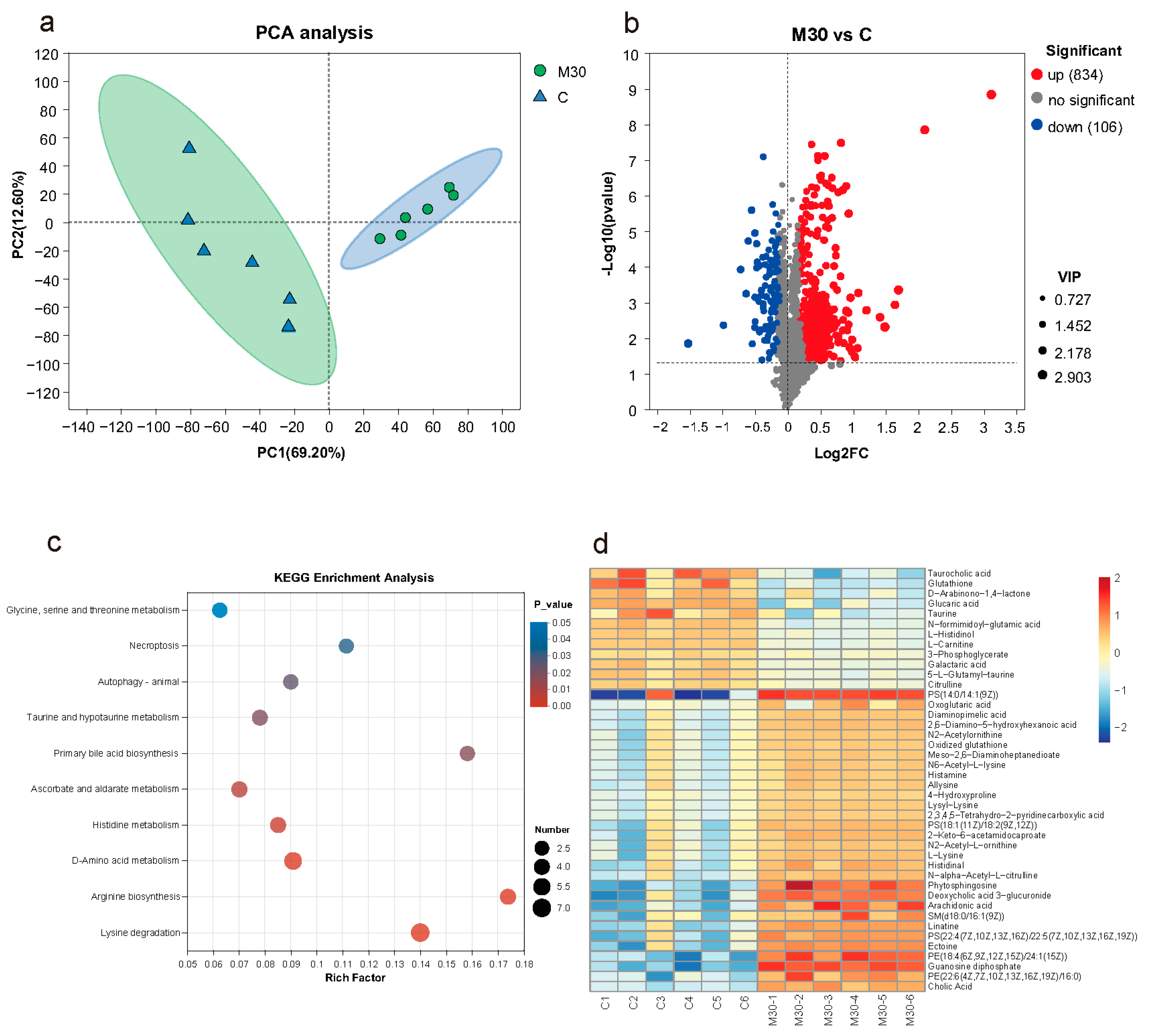
2.4. Liver Metabolomic Profiles
2.5. The Expression Levels of Genes in the Liver
2.6. Liver Bile Acid Contents
2.7. Integrated Metabolomic, Transcriptomic, and Biochemical Analysis Revealed the Mechanism of Hepatotoxicity Induced by Chronic MC-LR Exposure
3. Discussion
3.1. MC-LR Exposure Induced Hepatotoxicity
3.2. MC-LR Exposure Caused Oxidative Stress by Reducing Antioxidant Status and Triggering Apoptosis and Autophagy
3.3. MC-LR Exposure Disturbed Lipid Metabolism
3.4. MC-LR Exposure Disturbed Amino Acid Metabolism
3.5. MC-LR Exposure Inhibited BA Synthesis
4. Conclusions
5. Materials and Methods
5.1. Fish Culture and Experimental Design
5.2. Fish Sampling
5.3. Histological Examination, DHE Staining, and TUNEL Staining
5.3.1. H&E Staining
5.3.2. Oil Red O Staining
5.3.3. Dihydroethidium (DHE) Staining
5.3.4. TUNEL Staining
5.4. Biochemical Parameter Determination
5.5. Transcriptomic Analysis
5.5.1. Total Extraction of RNA and Sequencing
5.5.2. Transcript Assembly and Annotation
5.5.3. Analysis of Differentially Expressed Genes
5.6. Metabolomic Analysis
5.7. qPCR Analysis
5.8. Statistic Analysis
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Chen, L.; Hu, Y.; He, J.; Chen, J.; Giesy, J.P.; Xie, P. Responses of the Proteome and Metabolome in Livers of Zebrafish Exposed Chronically to Environmentally Relevant Concentrations of Microcystin-LR. Environ. Sci. Technol. 2017, 51, 596–607. [Google Scholar] [CrossRef] [PubMed]
- Du, C.; Zheng, S.; Yang, Y.; Feng, X.; Chen, J.; Tang, Y.; Wang, H.; Yang, F. Chronic Exposure to Low Concentration of MC-LR Caused Hepatic Lipid Metabolism Disorder. Ecotoxicol. Environ. Saf. 2022, 239, 113649. [Google Scholar] [CrossRef] [PubMed]
- Jiang, J.; Gu, X.; Song, R.; Zhang, Q.; Geng, J.; Wang, X.; Yang, L. Time-Dependent Oxidative Stress and Histopathological Changes in Cyprinus carpio L. Exposed to Microcystin-LR. Ecotoxicology 2011, 20, 1000–1009. [Google Scholar] [CrossRef] [PubMed]
- Zhang, H.; Cai, C.; Fang, W.; Wang, J.; Zhang, Y.; Liu, J.; Jia, X. Oxidative Damage and Apoptosis Induced by Microcystin-LR in the Liver of Rana Nigromaculata in Vivo. Aquat. Toxicol. 2013, 140–141, 11–18. [Google Scholar] [CrossRef] [PubMed]
- Zhang, D.; Lin, W.; Liu, Y.; Guo, H.; Wang, L.; Yang, L.; Li, L.; Li, D.; Tang, R. Chronic Microcystin-LR Exposure Induces Abnormal Lipid Metabolism via Endoplasmic Reticulum Stress in Male Zebrafish. Toxins 2020, 12, 107. [Google Scholar] [CrossRef] [PubMed]
- Xiao, L.; Pan, G. An Important Intestinal Transporter That Regulates the Enterohepatic Circulation of Bile Acids and Cholesterol Homeostasis: The Apical Sodium-Dependent Bile Acid Transporter (SLC10A2/ASBT). Clin. Res. Hepatol. Gastroenterol. 2017, 41, 509–515. [Google Scholar] [CrossRef] [PubMed]
- Arab, J.P.; Karpen, S.J.; Dawson, P.A.; Arrese, M.; Trauner, M. Bile Acids and Nonalcoholic Fatty Liver Disease: Molecular Insights and Therapeutic Perspectives. Hepatology 2017, 65, 350–362. [Google Scholar] [CrossRef] [PubMed]
- Su, X.; Steinman, A.D.; Xue, Q.; Zhao, Y.; Xie, L. Evaluating the Contamination of Microcystins in Lake Taihu, China: The Application of Equivalent Total MC-LR Concentration. Ecol. Indic. 2018, 89, 445–454. [Google Scholar] [CrossRef]
- Jayaraj, R.; Anand, T.; Rao, P.V.L. Activity and Gene Expression Profile of Certain Antioxidant Enzymes to Microcystin-LR Induced Oxidative Stress in Mice. Toxicology 2006, 220, 136–146. [Google Scholar] [CrossRef]
- Lin, W.; Hou, J.; Guo, H.; Li, L.; Wang, L.; Zhang, D.; Li, D.; Tang, R. The Synergistic Effects of Waterborne Microcystin-LR and Nitrite on Hepatic Pathological Damage, Lipid Peroxidation and Antioxidant Responses of Male Zebrafish. Environ. Pollut. 2018, 235, 197–206. [Google Scholar] [CrossRef]
- Cheong, A.W.Y.; Lee, Y.-L.; Liu, W.-M.; Yeung, W.S.B.; Lee, K.-F. Oviductal Microsomal Epoxide Hydrolase (EPHX1) Reduces Reactive Oxygen Species (ROS) Level and Enhances Preimplantation Mouse Embryo Development1. Biol. Reprod. 2009, 81, 126–132. [Google Scholar] [CrossRef] [PubMed]
- Nebert, D.W.; Vasiliou, V. Analysis of the Glutathione S-Transferase (GST) Gene Family. Human. Genom. 2004, 1, 460. [Google Scholar] [CrossRef] [PubMed]
- Jiang, J.; Shi, Y.; Shan, Z.; Yang, L.; Wang, X.; Shi, L. Bioaccumulation, Oxidative Stress and HSP70 Expression in Cyprinus Carpio L. Exposed to Microcystin-LR under Laboratory Conditions. Comp. Biochem. Phys. C 2012, 155, 483–490. [Google Scholar] [CrossRef] [PubMed]
- Scherz-Shouval, R.; Elazar, Z. Regulation of Autophagy by ROS: Physiology and Pathology. Trends Biochem. Sci. 2011, 36, 30–38. [Google Scholar] [CrossRef] [PubMed]
- Zhan, C.; Liu, W.; Zhang, F.; Zhang, X. Microcystin-LR Triggers Different Endoplasmic Reticulum Stress Pathways in the Liver, Ovary, and Offspring of Zebrafish (Danio Rerio). J. Hazard. Mater. 2020, 386, 121939. [Google Scholar] [CrossRef] [PubMed]
- Wang, K. Autophagy and Apoptosis in Liver Injury. Cell Cycle 2015, 14, 1631–1642. [Google Scholar] [CrossRef] [PubMed]
- Holcik, M.; Korneluk, R.G. XIAP, the Guardian Angel. Nat. Rev. Mol. Cell Biol. 2001, 2, 550–556. [Google Scholar] [CrossRef] [PubMed]
- Peña, L.A.; Fuks, Z.; Koksnick, R. Stress-Induced Apoptosis and the Sphingomyelin Pathway. Biochem. Pharmacol. 1997, 53, 615–621. [Google Scholar] [CrossRef] [PubMed]
- Baliou, S.; Adamaki, M.; Ioannou, P.; Pappa, A.; Panayiotidis, M.I.; Spandidos, D.A.; Christodoulou, I.; Kyriakopoulos, A.M.; Zoumpourlis, V. Protective Role of Taurine against Oxidative Stress (Review). Mol. Med. Rep. 2021, 24, 605. [Google Scholar] [CrossRef]
- Wei, L.; Fu, J.; He, L.; Wang, H.; Ruan, J.; Li, F.; Wu, H. Microcystin-LR-Induced Autophagy Regulates Oxidative Stress, Inflammation, and Apoptosis in Grass Carp Ovary Cells in Vitro. Toxicol. In Vitro 2023, 87, 105520. [Google Scholar] [CrossRef]
- Di, Y.-Q.; Han, X.-L.; Kang, X.-L.; Wang, D.; Chen, C.-H.; Wang, J.-X.; Zhao, X.-F. Autophagy Triggers CTSD (Cathepsin D) Maturation and Localization inside Cells to Promote Apoptosis. Autophagy 2021, 17, 1170–1192. [Google Scholar] [CrossRef] [PubMed]
- Singh, P.; Ravanan, P.; Talwar, P. Death Associated Protein Kinase 1 (DAPK1): A Regulator of Apoptosis and Autophagy. Front. Mol. Neurosci. 2016, 9, 46. [Google Scholar] [CrossRef] [PubMed]
- Hsu, P.; Shi, Y. Regulation of Autophagy by Mitochondrial Phospholipids in Health and Diseases. Biochim. Biophys. Acta Mol. Cell Biol. Lipids 2017, 1862, 114–129. [Google Scholar] [CrossRef]
- Tessari, P.; Coracina, A.; Cosma, A.; Tiengo, A. Hepatic Lipid Metabolism and Non-Alcoholic Fatty Liver Disease. Nutr. Metab. Cardiovasc. Dis. 2009, 19, 291–302. [Google Scholar] [CrossRef]
- Chambers, K.F.; Day, P.E.; Aboufarrag, H.T.; Kroon, P.A. Polyphenol Effects on Cholesterol Metabolism via Bile Acid Biosynthesis, CYP7A1: A Review. Nutrients 2019, 11, 2588. [Google Scholar] [CrossRef]
- Franklin, M.P.; Sathyanarayan, A.; Mashek, D.G. Acyl-CoA Thioesterase 1 (ACOT1) Regulates PPARα to Couple Fatty Acid Flux with Oxidative Capacity During Fasting. Diabetes 2017, 66, 2112–2123. [Google Scholar] [CrossRef] [PubMed]
- Ming, J.; Ye, J.; Zhang, Y.; Yang, X.; Shao, X.; Qiang, J.; Xu, P. Dietary Optimal Reduced Glutathione Improves Innate Immunity, Oxidative Stress Resistance and Detoxification Function of Grass Carp (Ctenopharyngodon Idella) against Microcystin-LR. Aquaculture 2019, 498, 594–605. [Google Scholar] [CrossRef]
- Botta, D.; White, C.C.; Vliet-Gregg, P.; Mohar, I.; Shi, S.; McGrath, M.B.; McConnachie, L.A.; Kavanagh, T.J. Modulating GSH Synthesis Using Glutamate Cysteine Ligase Transgenic and Gene-Targeted Mice. Drug Metab. Rev. 2008, 40, 465–477. [Google Scholar] [CrossRef]
- MacGlashan, D. Histamine: A Mediator of Inflammation. J. Allergy Clin. 2003, 112, S53–S59. [Google Scholar] [CrossRef]
- Ai, W.; Takaishi, S.; Wang, T.C.; Fleming, J.V. Regulation of l-Histidine Decarboxylase and Its Role in Carcinogenesis. Prog. Nucleic Acid. Res. Mol. Biol. 2006, 81, 231–270. [Google Scholar]
- Dalvie, D.; Di, L. Aldehyde Oxidase and Its Role as a Drug Metabolizing Enzyme. Pharmacol. Ther. 2019, 201, 137–180. [Google Scholar] [CrossRef]
- Chow, M.D.; Lee, Y.-H.; Guo, G.L. The Role of Bile Acids in Nonalcoholic Fatty Liver Disease and Nonalcoholic Steatohepatitis. Mol. Aspects Med. 2017, 56, 34–44. [Google Scholar] [CrossRef] [PubMed]
- Rizzolo, D.; Buckley, K.; Kong, B.; Zhan, L.; Shen, J.; Stofan, M.; Brinker, A.; Goedken, M.; Buckley, B.; Guo, G.L. Bile Acid Homeostasis in a Cholesterol 7α-Hydroxylase and Sterol 27-Hydroxylase Double Knockout Mouse Model. Hepatology 2019, 70, 389–402. [Google Scholar] [CrossRef] [PubMed]
- Nies, A.T.; Keppler, D. The Apical Conjugate Efflux Pump ABCC2 (MRP2). Pflugers Arch. 2007, 453, 643–659. [Google Scholar] [CrossRef] [PubMed]
- Martin, G.G.; Landrock, D.; Landrock, K.K.; Howles, P.N.; Atshaves, B.P.; Kier, A.B.; Schroeder, F. Relative Contributions of L-FABP, SCP-2/SCP-x, or Both to Hepatic Biliary Phenotype of Female Mice. Arch. Biochem. Biophys. 2015, 588, 25–32. [Google Scholar] [CrossRef][Green Version]
- Chi, C.; Giri, S.S.; Yu, X.W.; Liu, Y.; Chen, K.K.; Liu, W.B.; Zhang, D.D.; Jiang, G.Z.; Li, X.F.; Gao, X.; et al. Lipid Metabolism, Immune and Apoptosis Transcriptomic Responses of the Hepatopancreas of Chinese Mitten Crab to the Exposure to Microcystin-LR. Ecotoxicol. Environ. Saf. 2022, 236, 113439. [Google Scholar] [CrossRef] [PubMed]
- Duan, Y.; Xing, Y.; Zeng, S.; Dan, X.; Mo, Z.; Zhang, J.; Li, Y. Integration of Metagenomic and Metabolomic Insights into the Effects of Microcystin-LR on Intestinal Microbiota of Litopenaeus vannamei. Front. Microbiol. 2022, 13, 994188. [Google Scholar] [CrossRef]
- Hou, J.; Li, L.; Wu, N.; Su, Y.; Lin, W.; Li, G.; Gu, Z. Reproduction Impairment and Endocrine Disruption in Female Zebrafish after Long-Term Exposure to MC-LR: A Life Cycle Assessment. Environ. Pollut. 2016, 208, 477–485. [Google Scholar] [CrossRef]
- Sun, C.; Li, A.; Wang, H.; Ma, J.; Hou, J. Positive Regulation of Acetate in Adipocyte Differentiation and Lipid Deposition in Obese Mice. Nutrients 2023, 15, 3736. [Google Scholar] [CrossRef]
- Tian, J.; Jin, Y.; Yu, E.; Sun, J.; Xia, Y.; Zhang, K.; Li, Z.; Gong, W.; Wang, G.; Xie, J. Farnesoid X Receptor Is an Effective Target for Modulating Lipid Accumulation in Grass Carp, Ctenopharyngodon Idella. Aquaculture 2021, 534, 736248. [Google Scholar] [CrossRef]
- Li, Y.; Fu, B.; Zhang, J.; Wang, G.; Gong, W.; Tian, J.; Li, H.; Zhang, K.; Xia, Y.; Li, Z.; et al. Effects of Heat Stress on the Chemical Composition, Oxidative Stability, Muscle Metabolism, and Meat Quality of Nile Tilapia (Oreochromis niloticus). Food Chem. 2023, 426, 136590. [Google Scholar] [CrossRef] [PubMed]
- Wang, G.; Xin, Y.; Ping, K.; Xu, X.; Sun, Y.; Li, X.; Guan, X.; Dong, J. Silybin Mitigates Chronic Avermectin Exposure-Induced Intestinal Damage and Growth Inhibition in Carp. Environ. Toxicol. Pharmacol. 2023, 106, 104351. [Google Scholar] [CrossRef] [PubMed]
- Li, H.; Xu, W.; Wu, L.; Dong, B.; Jin, J.; Han, D.; Zhu, X.; Yang, Y.; Liu, H.; Xie, S. Differential Regulation of Endoplasmic Reticulum Stress-Induced Autophagy and Apoptosis in Two Strains of Gibel Carp (Carassius Gibelio) Exposed to Acute Waterborne Cadmium. Aquat. Toxicol. 2021, 231, 105721. [Google Scholar] [CrossRef] [PubMed]
- Li, B.; Dewey, C.N. RSEM: Accurate Transcript Quantification from RNA-Seq Data with or without a Reference Genome. BMC Bioinf. 2011, 12, 323. [Google Scholar] [CrossRef] [PubMed]
- Fu, B.; Xie, J.; Kaneko, G.; Wang, G.; Yang, H.; Tian, J.; Xia, Y.; Li, Z.; Gong, W.; Zhang, K.; et al. MicroRNA-Dependent Regulation of Targeted mRNAs for Improved Muscle Texture in Crisp Grass Carp Fed with Broad Bean. Food Res. Int. 2022, 155, 111071. [Google Scholar] [CrossRef] [PubMed]
- Bhargava, P.; Smith, M.D.; Mische, L.; Harrington, E.; Fitzgerald, K.C.; Martin, K.; Kim, S.; Reyes, A.A.; Gonzalez-Cardona, J.; Volsko, C.; et al. Bile Acid Metabolism Is Altered in Multiple Sclerosis and Supplementation Ameliorates Neuroinflammation. J. Clin. Investig. 2020, 130, 3467–3482. [Google Scholar] [CrossRef] [PubMed]
- Hu, Z.; Qi, C.; Lin, C.; Tang, R. Nitrite Stress Induces Oxidative Stress and Leads to Muscle Quality Decreased in Wuchang Bream (Megalobrama amblycephala Yih) Juveniles. Water 2022, 14, 160. [Google Scholar] [CrossRef]
- Yang, T.; Shu, T.; Liu, G.; Mei, H.; Zhu, X.; Huang, X.; Zhang, L.; Jiang, Z. Quantitative Profiling of 19 Bile Acids in Rat Plasma, Liver, Bile and Different Intestinal Section Contents to Investigate Bile Acid Homeostasis and the Application of Temporal Variation of Endogenous Bile Acids. J. Steroid Biochem. Mol. Biol. 2017, 172, 69–78. [Google Scholar] [CrossRef]
- Hu, T.; An, Z.; Shi, C.; Li, P.; Liu, L. A Sensitive and Efficient Method for Simultaneous Profiling of Bile Acids and Fatty Acids by UPLC-MS/MS. J. Pharm. Biomed. Anal. 2020, 178, 112815. [Google Scholar] [CrossRef]
- Yu, L.; Tian, J.; Zhang, C.; Xue, Z.; Lu, X.; Wen, H.; Jiang, M.; Wu, F. Acetylferulic Paeonol Ester: A New Feed Additive Reduces Lipid Accumulation in the Liver of Nile Tilapia (Oreochromis Niloticus) by Modulating Lipid and Glucose Metabolism. Aquaculture 2022, 561, 738671. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| C | M1 | M3 | M10 | M30 | |
|---|---|---|---|---|---|
| TG (mmol/L) | 5.2 ± 0.40 c | 5.62 ± 0.95 bc | 5.59 ± 0.75 bc | 6.82 ± 1.48 ab | 7.15 ± 1.40 a |
| TC (mmol/L) | 3.86 ± 0.44 b | 4.12 ± 0.83 ab | 4.31 ± 0.77 ab | 5.03 ± 1.22 ab | 5.43 ± 1.16 a |
| LDL-C (mmol/L) | 3.32 ± 0.38 b | 3.45 ± 0.48 b | 3.91 ± 0.78 ab | 4.11 ± 0.82 ab | 4.73 ± 0.81 a |
| HDL-C (mmol/L) | 2.91 ± 0.43 c | 3.15 ± 0.72 c | 3.52 ± 0.65 bc | 4.10 ± 0.42 ab | 4.59 ± 0.92 a |
| ALT (U/L) | 21.38 ± 2.86 c | 24.39 ± 4.12 bc | 27.33 ± 6.03 bc | 28.42 ± 5.19 ab | 33.15 ± 4.81 a |
| AST (U/L) | 29.90 ± 4.70 b | 31.07 ± 4.64 b | 34.51 ± 4.74 ab | 36.02 ± 4.85 ab | 39.57 ± 5.17 a |
| AKP (U/L) | 133.98 ± 11.41 b | 143.00 ± 17.01 ab | 140.90 ± 13.15 ab | 151.90 ± 15.21 ab | 160.40 ± 13.79 a |
| C | M1 | M3 | M10 | M30 | |
|---|---|---|---|---|---|
| Lipid metabolism parameters | |||||
| TG (mmol/L) | 3.02 ± 0.37 c | 3.44 ± 0.62 c | 3.76 ± 0.62 c | 4.94 ± 0.47 b | 5.77 ± 0.68 a |
| TC (mmol/L) | 1.46 ± 0.19 b | 1.55 ± 0.14 b | 1.65 ± 0.28 b | 1.71 ± 0.21 ab | 1.97 ± 0.13 a |
| LDL-C (mmol/L) | 2.71 ± 0.45 c | 2.88 ± 0.43 c | 2.80 ± 0.32 c | 3.58 ± 0.41 b | 4.53 ± 0.55 a |
| HDL-C (mmol/L) | 1.68 ± 0.22 b | 1.77 ± 0.22 b | 1.79 ± 0.23 b | 1.92 ± 0.36 b | 2.44 ± 0.55 a |
| Oxidative stress parameters | |||||
| MDA (nmol/mgprot) | 25.15 ± 2.61 b | 25.27 ± 3.93 b | 28.08 ± 4.78 ab | 29.73 ± 3.51 ab | 31.69 ± 4.37 a |
| ROS (U/g) | 2533.54 ± 373.85 b | 2760.41 ± 599.13 ab | 2759.42 ± 374.84 ab | 3350.73 ± 440.91 a | 3201.53 ± 406.28 a |
| H2O2 (μg/g) | 16.71 ± 1.71 c | 17.42 ± 3.62 bc | 18.32 ± 3.15 bc | 21.51 ± 3.70 ab | 23.46 ± 3.24 a |
| O2- (μg/g) | 84.67 ± 7.73 c | 91.56 ± 16.30 bc | 95.06 ± 10.83 bc | 106.44 ± 10.45 ab | 115.29 ± 13.03 a |
| OH- (μg/g) | 53.47 ± 5.73 | 57.83 ± 9.71 | 54.97 ± 9.45 | 64.83 ± 9.81 | 56.76 ± 7.12 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Li, Y.; Yang, H.; Fu, B.; Kaneko, G.; Li, H.; Tian, J.; Wang, G.; Wei, M.; Xie, J.; Yu, E. Integration of Multi-Omics, Histological, and Biochemical Analysis Reveals the Toxic Responses of Nile Tilapia Liver to Chronic Microcystin-LR Exposure. Toxins 2024, 16, 149. https://doi.org/10.3390/toxins16030149
Li Y, Yang H, Fu B, Kaneko G, Li H, Tian J, Wang G, Wei M, Xie J, Yu E. Integration of Multi-Omics, Histological, and Biochemical Analysis Reveals the Toxic Responses of Nile Tilapia Liver to Chronic Microcystin-LR Exposure. Toxins. 2024; 16(3):149. https://doi.org/10.3390/toxins16030149
Chicago/Turabian StyleLi, Yichao, Huici Yang, Bing Fu, Gen Kaneko, Hongyan Li, Jingjing Tian, Guangjun Wang, Mingken Wei, Jun Xie, and Ermeng Yu. 2024. "Integration of Multi-Omics, Histological, and Biochemical Analysis Reveals the Toxic Responses of Nile Tilapia Liver to Chronic Microcystin-LR Exposure" Toxins 16, no. 3: 149. https://doi.org/10.3390/toxins16030149
APA StyleLi, Y., Yang, H., Fu, B., Kaneko, G., Li, H., Tian, J., Wang, G., Wei, M., Xie, J., & Yu, E. (2024). Integration of Multi-Omics, Histological, and Biochemical Analysis Reveals the Toxic Responses of Nile Tilapia Liver to Chronic Microcystin-LR Exposure. Toxins, 16(3), 149. https://doi.org/10.3390/toxins16030149