In Vitro Assessment of Ozone-Treated Deoxynivalenol by Measuring Cytotoxicity and Wheat Quality



Abstract
1. Introduction
2. Results
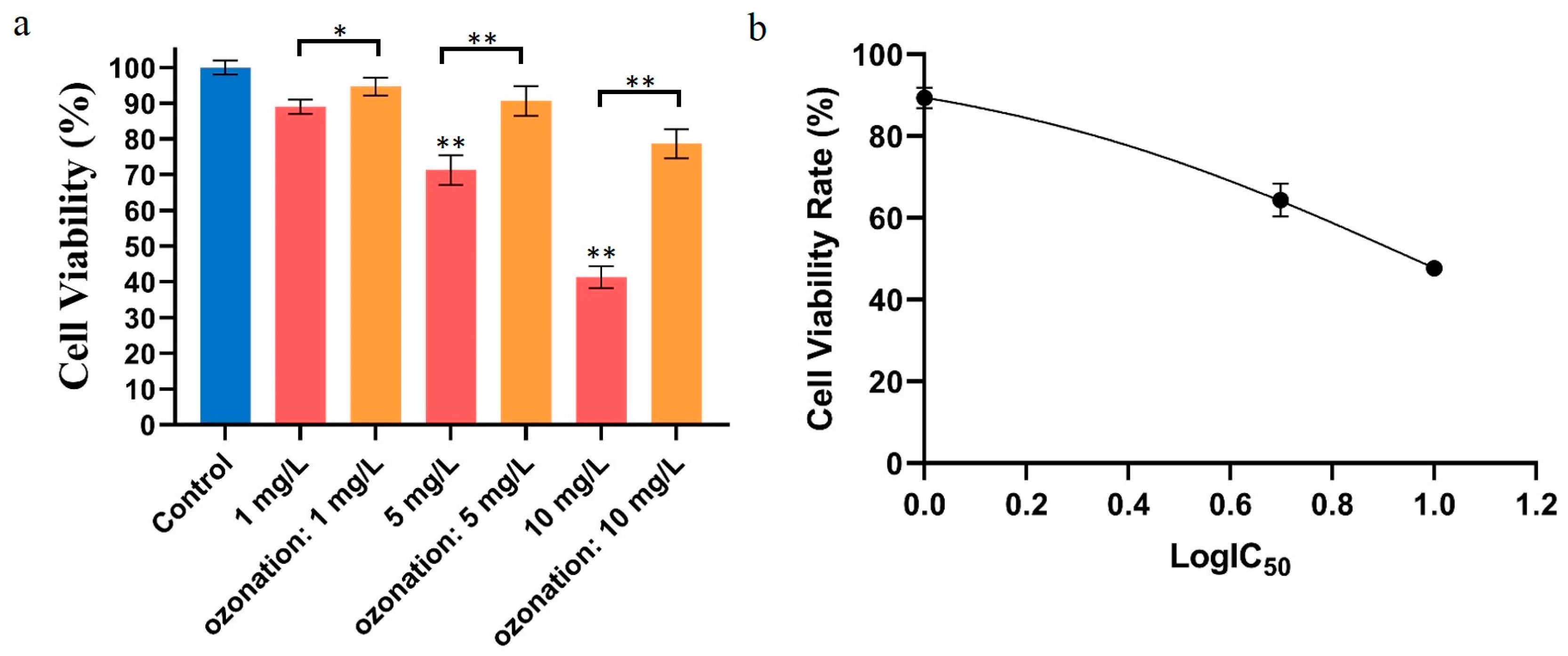
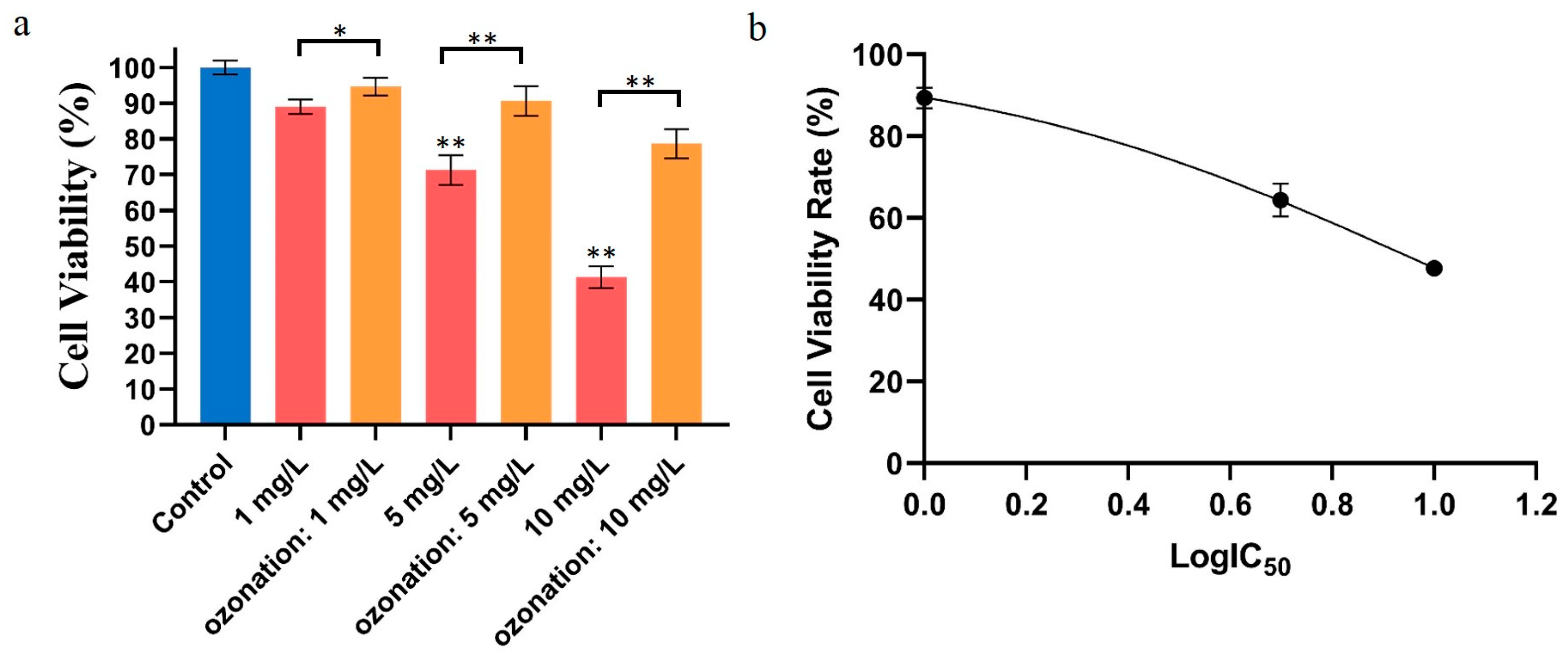
2.1. Effects of DON Degradation on HepG2 Activity
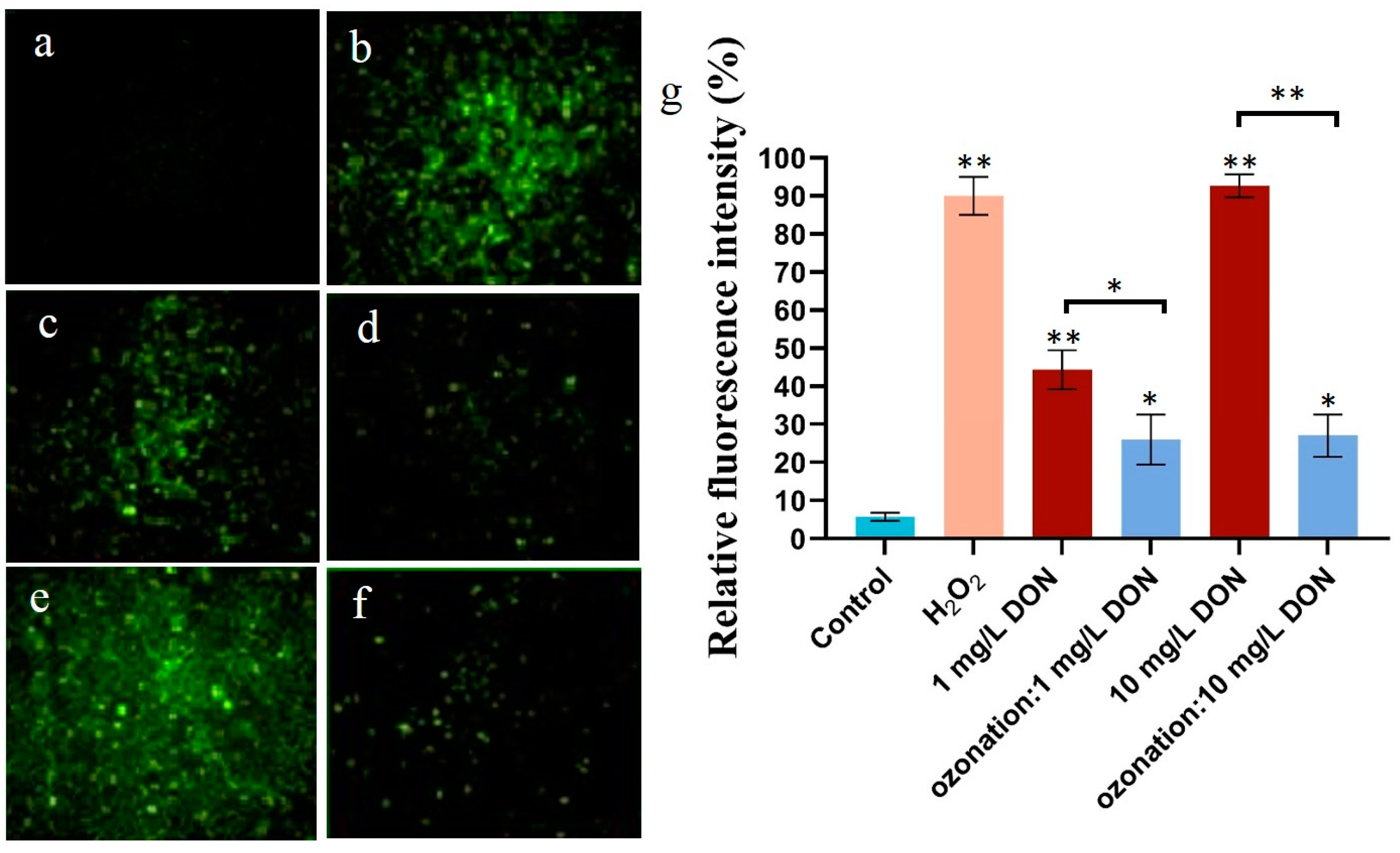
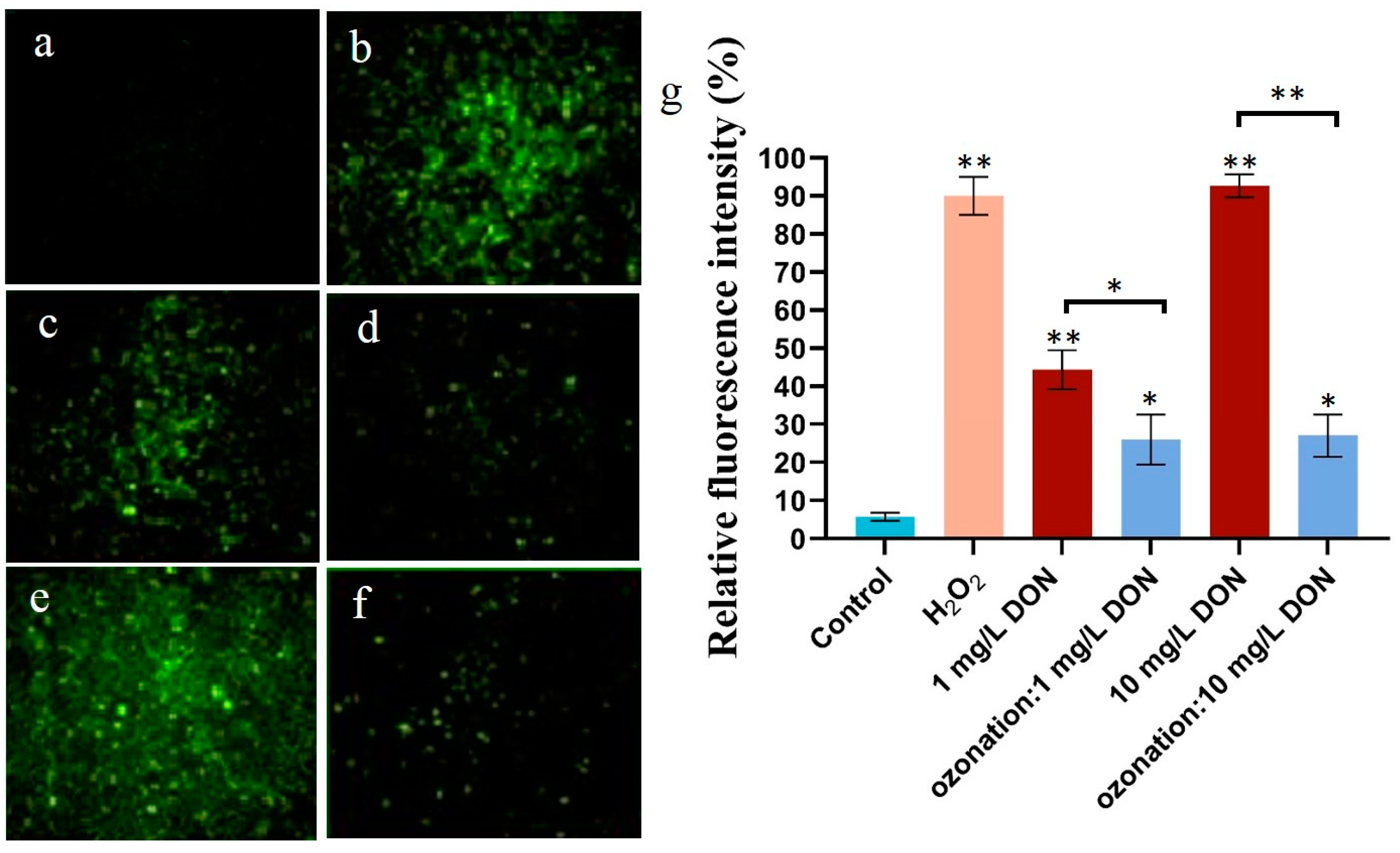
2.2. Effects of DON Ozonation on Reactive Oxygen Species in HepG2
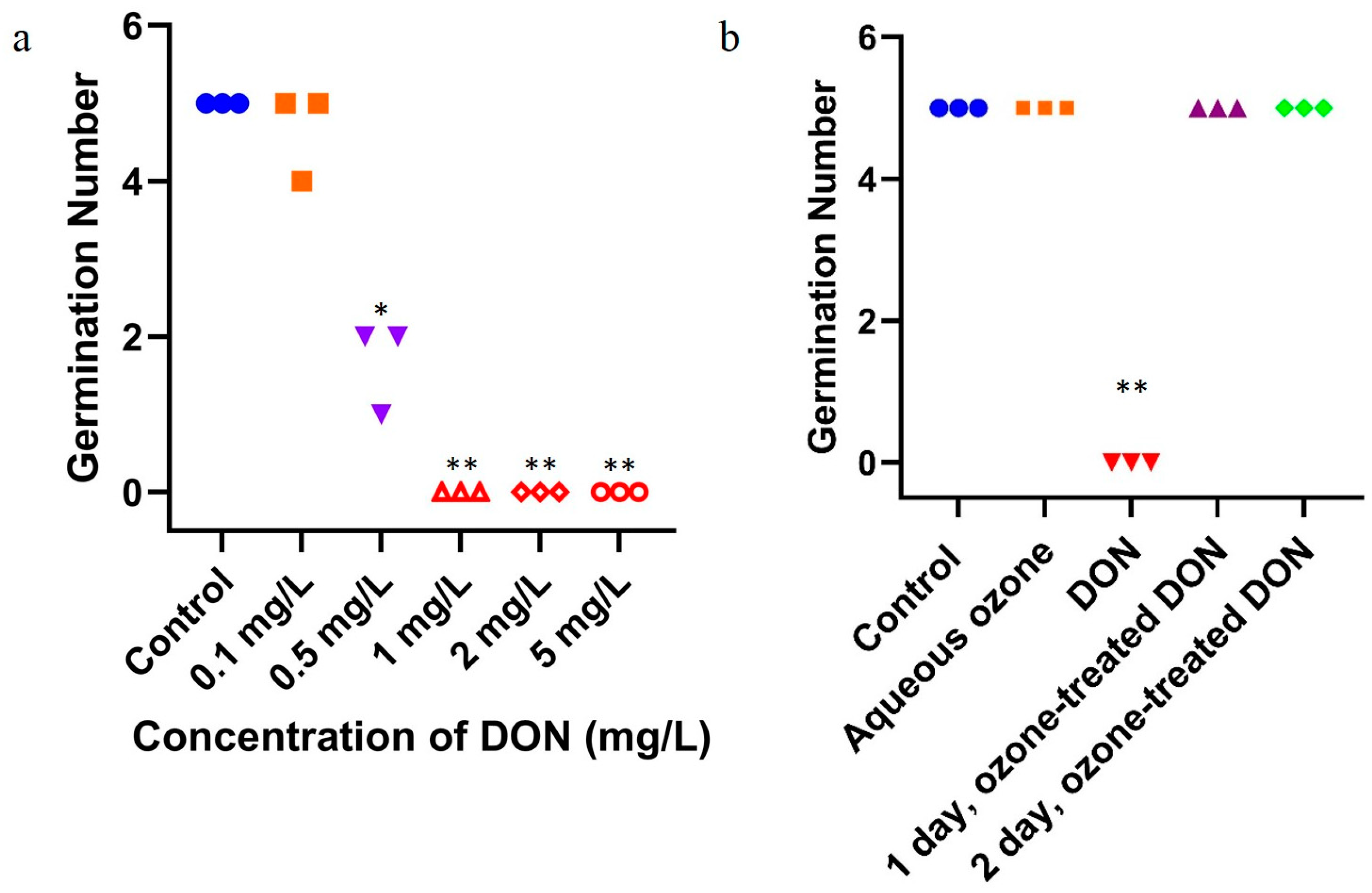
2.3. Effect of Ozone Degradation of DON on Mung Bean Germination
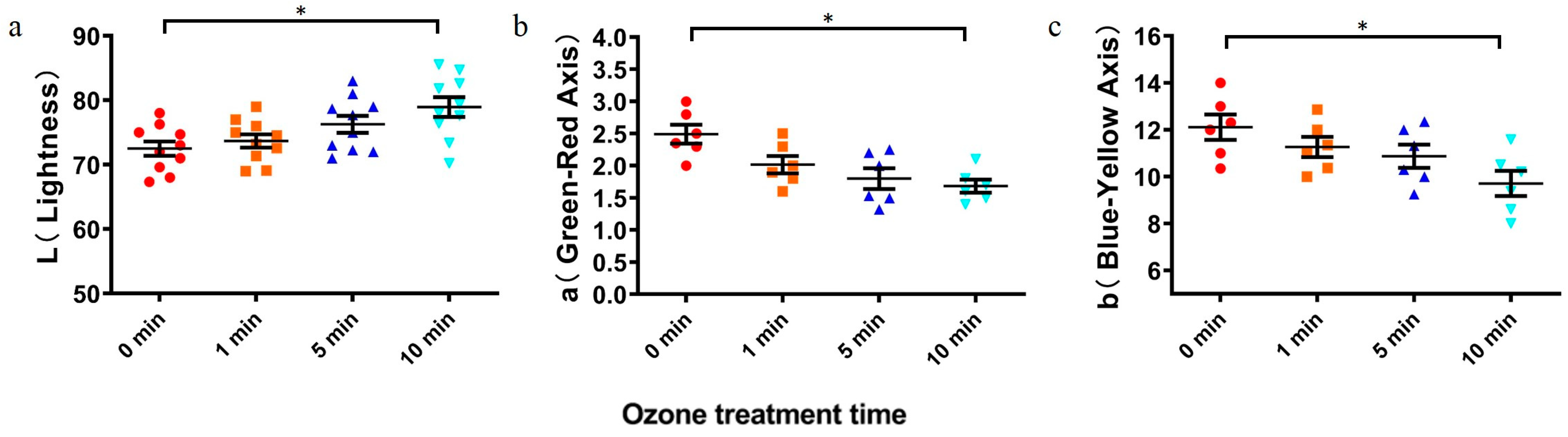
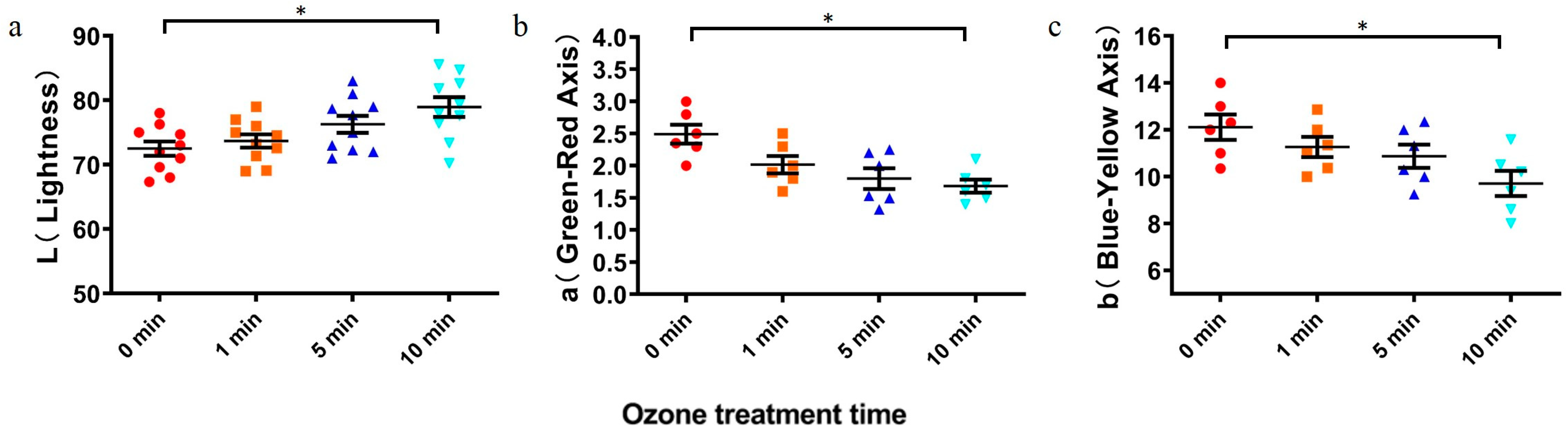
2.4. Effect of Ozone Degradation of DON on Wheat Quality
3. Discussion
4. Conclusions
5. Materials and Methods
5.1. Chemicals and Materials
5.2. Extraction of DON from Wheat
5.3. Culture of HepG2
5.4. MTT Testing of HepG2 Activity
5.5. Determination of Intracellular Reactive Oxygen Species Levels in HepG2
5.6. Inhibition Experiment of Ozone-Degrading DON on Mung Bean Germination
5.7. Determination of Ozone Degradation of DON and Its Effect on Wheat Quality
5.8. Statistical Analysis
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Sun, C.; Yang, F.; Xiao, J.; Zhou, W.; Li, J.; Gu, X. Simulating ozone degradation of deoxynivalenol and its bio-safety assessment by mouse model. Front. Microbiol. 2023, 14, 1286503. [Google Scholar] [CrossRef] [PubMed]
- Sun, C.; Zhu, P.; Ji, J.; Sun, J.; Tang, L.; Pi, F.; Sun, X. Role of aqueous chlorine dioxide in controlling the growth of Fusarium graminearum and its application on contaminated wheat. LWT-Food Sci. Technol. 2017, 84, 555–561. [Google Scholar] [CrossRef]
- European Food Safety Authority. Deoxynivalenol in food and feed: Occurrence and exposure. EFSA J. 2013, 11, 3379. [Google Scholar]
- Beisl, J.; Pahlke, G.; Ehling-Schulz, M.; Del Favero, G.; Marko, D. Cereulide and Deoxynivalenol Increase LC3 Protein Levels in HepG2 Liver Cells. Toxins 2022, 14, 151. [Google Scholar] [CrossRef] [PubMed]
- Juan-Garcia, A.; Taroncher, M.; Font, G.; Ruiz, M.J. Micronucleus induction and cell cycle alterations produced by deoxynivalenol and its acetylated derivatives in individual and combined exposure on HepG2 cells. Food Chem. Toxicol. 2018, 118, 719–725. [Google Scholar] [CrossRef] [PubMed]
- Li, H.; Su, J.; Jiang, J.; Li, Y.; Gan, Z.; Ding, Y.; Li, Y.; Liu, J.; Wang, S.; Ke, Y. Characterization of polysaccharide from Scutellaria barbata and its antagonistic effect on the migration and invasion of HT-29 colorectal cancer cells induced by TGF-beta1. Int. J. Biol. Macromol. 2019, 131, 886–895. [Google Scholar] [CrossRef]
- Ansari, K.I.; Doyle, S.M.; Kacprzyk, J.; Khan, M.R.; Walter, S.; Brennan, J.M.; Arunachalam, C.S.; McCabe, P.F.; Doohan, F.M. Light influences how the fungal toxin deoxynivalenol affects plant cell death and defense responses. Toxins 2014, 6, 679–692. [Google Scholar] [CrossRef]
- Tripathi, S.; Hussain, T. Water and wastewater treatment through ozone-based technologies. In Development in Wastewater Treatment Research and Processes; Elsevier: Amsterdam, The Netherlands, 2022; pp. 139–172. [Google Scholar]
- Savi, G.D.; Gomes, T.; Canever, S.B.; Feltrin, A.C.; Piacentini, K.C.; Scussel, R.; Oliveira, D.; Machado-de-Ávila, R.A.; Cargnin, M.; Angioletto, E. Application of ozone on rice storage: A mathematical modeling of the ozone spread, effects in the decontamination of filamentous fungi and quality attributes. J. Stored Prod. Res. 2020, 87, 101605. [Google Scholar] [CrossRef]
- Conte, G.; Fontanelli, M.; Galli, F.; Cotrozzi, L.; Pagni, L.; Pellegrini, E. Mycotoxins in Feed and Food and the Role of Ozone in Their Detoxification and Degradation: An Update. Toxins 2020, 12, 486. [Google Scholar] [CrossRef]
- Sun, C.; Ji, J.; Wu, S.; Sun, C.; Pi, F.; Zhang, Y.; Tang, L.; Sun, X. Saturated aqueous ozone degradation of deoxynivalenol and its application in contaminated grains. Food Control 2016, 69, 185–190. [Google Scholar] [CrossRef]
- Shewry, P.R. Wheat. J. Exp. Bot. 2009, 60, 1537–1553. [Google Scholar] [CrossRef]
- Li, M.; Guan, E.; Bian, K. Effect of ozone treatment on deoxynivalenol and quality evaluation of ozonised wheat. Food Addit. Contam. A 2015, 32, 544–553. [Google Scholar] [CrossRef] [PubMed]
- Ismaiel, A.; Papenbrock, J. Mycotoxins: Producing fungi and mechanisms of phytotoxicity. Agriculture 2015, 5, 492–537. [Google Scholar] [CrossRef]
- Hooshmand, H.; Klopfenstein, C. Effects of gamma irradiation on mycotoxin disappearance and amino acid con-tents of corn, wheat, and soybeans with different moisture contents. Plant Food Hum. Nutr. 1995, 47, 227–238. [Google Scholar] [CrossRef]
- Xu, R.; Karrow, N.A.; Shandilya, U.K.; Sun, L.H.; Kitazawa, H. In-Vitro Cell Culture for Efficient Assessment of Mycotoxin Exposure, Toxicity and Risk Mitigation. Toxins 2020, 12, 146. [Google Scholar] [CrossRef] [PubMed]
- Morimura, H.; Ito, M.; Yoshida, S.; Koitabashi, M.; Tsushima, S.; Camagna, M.; Chiba, S.; Takemoto, D.; Kawakita, K.; Sato, I. In Vitro Assessment of Biocontrol Effects on Fusarium Head Blight and Deoxynivalenol (DON) Accumulation by DON-Degrading Bacteria. Toxins 2020, 12, 399. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Wang, Y.; Deng, Y.; Wang, X.; Wu, W.; Nepovimova, E.; Wu, Q.; Kuca, K. Toxic mechanisms of the trichothecenes T-2 toxin and deoxynivalenol on protein synthesis. Food Chem. Toxicol. 2022, 164, 113044. [Google Scholar] [CrossRef]
- Guo, H.; Ji, J.; Wei, K.; Sun, J.; Zhang, Y.; Sun, X. MAPK/AP-1 and ROS participated in ratio- and time-dependent interaction effects of deoxynivalenol and cadmium on HT-29 cells. Food Chem. Toxicol. 2021, 148, 111921. [Google Scholar] [CrossRef]
- Fernandez-Blanco, C.; Elmo, L.; Waldner, T.; Ruiz, M.J. Cytotoxic effects induced by patulin, deoxynivalenol and toxin T2 individually and in combination in hepatic cells (HepG2). Food Chem. Toxicol. 2018, 120, 12–23. [Google Scholar] [CrossRef]
- Kang, R.F.; Li, R.N.; Dai, P.Y.; Li, Z.J.; Li, Y.S.; Li, C.M. Deoxynivalenol induced apoptosis and inflammation of IPEC-J2 cells by promoting ROS production. Environ. Pollut. 2019, 251, 689–698. [Google Scholar] [CrossRef]
- Sun, L.; Dai, J.; Xu, J.; Yang, J.; Zhang, D. Comparative Cytotoxic Effects and Possible Mechanisms of Deoxynivalenol, Zearalenone and T-2 Toxin Exposure to Porcine Leydig Cells In Vitro. Toxins 2022, 14, 113. [Google Scholar] [CrossRef] [PubMed]
- Martinez-Alonso, C.; Taroncher, M.; Rodriguez-Carrasco, Y.; Ruiz, M.J. Evaluation of the Bioaccessible Fraction of T-2 Toxin from Cereals and Its Effect on the Viability of Caco-2 Cells Exposed to Tyrosol. Toxins 2023, 15, 493. [Google Scholar] [CrossRef] [PubMed]
- Zhou, Y.; Wu, S.; Wang, F.; Li, Q.; He, C.; Duan, N.; Wang, Z. Assessing the toxicity in vitro of degradation products from deoxynivalenol photocatalytic degradation by using upconversion nanoparticles@TiO2 composite. Chemosphere 2020, 238, 124648. [Google Scholar] [CrossRef] [PubMed]
- Qiu, Y.; Chen, X.; Zhang, J.; Ding, Y.; Lyu, F. Effects of tempering with plasma activated water on the degradation of deoxynivalenol and quality properties of wheat. Food Res. Int. 2022, 162, 112070. [Google Scholar] [CrossRef]
- Chen, R.; Ma, F.; Li, P.W.; Zhang, W.; Ding, X.X.; Zhang, Q.; Li, M.; Wang, Y.R.; Xu, B.C. Effect of ozone on aflatoxins detoxification and nutritional quality of peanuts. Food Chem. 2014, 146, 284–288. [Google Scholar] [CrossRef]
- Li, J.; Xie, L.; Qian, S.; Tang, Y.; Shen, M.; Li, S.; Wang, J.; Xiong, L.; Lu, J.; Zhong, W. A Type VI Secretion System Facilitates Fitness, Homeostasis, and Competitive Advantages for Environmental Adaptability and Efficient Nicotine Biodegradation. Appl. Environ. Microbiol. 2021, 87, e03113-20. [Google Scholar] [CrossRef]
- Liu, R.; Zhang, Y.; Hu, H.; Gan, T.; Huang, Z. Retrogradation behavior of starch dough prepared from damaged cassava starch and its application in functional gluten-free noodles. Int. J. Biol. Macromol. 2023, 236, 123996. [Google Scholar] [CrossRef]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Ozone Treatment Time (min) | Protein Content (g/100 g·Wheat) | Starch Content (g/100 g·Wheat) | Fatty Acid Content (mg/100 g·Wheat) |
|---|---|---|---|
| 0 | 11.6 ± 0.1 | 62.5 ± 0.6 | 52.7 ± 0.4 |
| 1 | 12.1 ± 0.3 | 62.2 ± 0.3 | 53.2 ± 0.2 |
| 5 | 12.3 ± 0.5 | 61.5 ± 0.5 | 53.5 ± 0.7 |
| 10 | 12.6 ± 0.2 | 61.1 ± 0.3 | 53.8 ± 0.3 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sun, C.; Mao, C.; Zhou, Z.; Xiao, J.; Zhou, W.; Du, J.; Li, J. In Vitro Assessment of Ozone-Treated Deoxynivalenol by Measuring Cytotoxicity and Wheat Quality. Toxins 2024, 16, 64. https://doi.org/10.3390/toxins16020064
Sun C, Mao C, Zhou Z, Xiao J, Zhou W, Du J, Li J. In Vitro Assessment of Ozone-Treated Deoxynivalenol by Measuring Cytotoxicity and Wheat Quality. Toxins. 2024; 16(2):64. https://doi.org/10.3390/toxins16020064
Chicago/Turabian StyleSun, Chao, Chuncai Mao, Zhie Zhou, Jianhui Xiao, Wenwen Zhou, Juan Du, and Jun Li. 2024. "In Vitro Assessment of Ozone-Treated Deoxynivalenol by Measuring Cytotoxicity and Wheat Quality" Toxins 16, no. 2: 64. https://doi.org/10.3390/toxins16020064
APA StyleSun, C., Mao, C., Zhou, Z., Xiao, J., Zhou, W., Du, J., & Li, J. (2024). In Vitro Assessment of Ozone-Treated Deoxynivalenol by Measuring Cytotoxicity and Wheat Quality. Toxins, 16(2), 64. https://doi.org/10.3390/toxins16020064