Modulation of Broiler Intestinal Changes Induced by Clostridium perfringens and Deoxynivalenol through Probiotic, Paraprobiotic, and Postbiotic Supplementation

 ,
, 
 , , , ,
, , , ,
Abstract
1. Introduction
2. Results
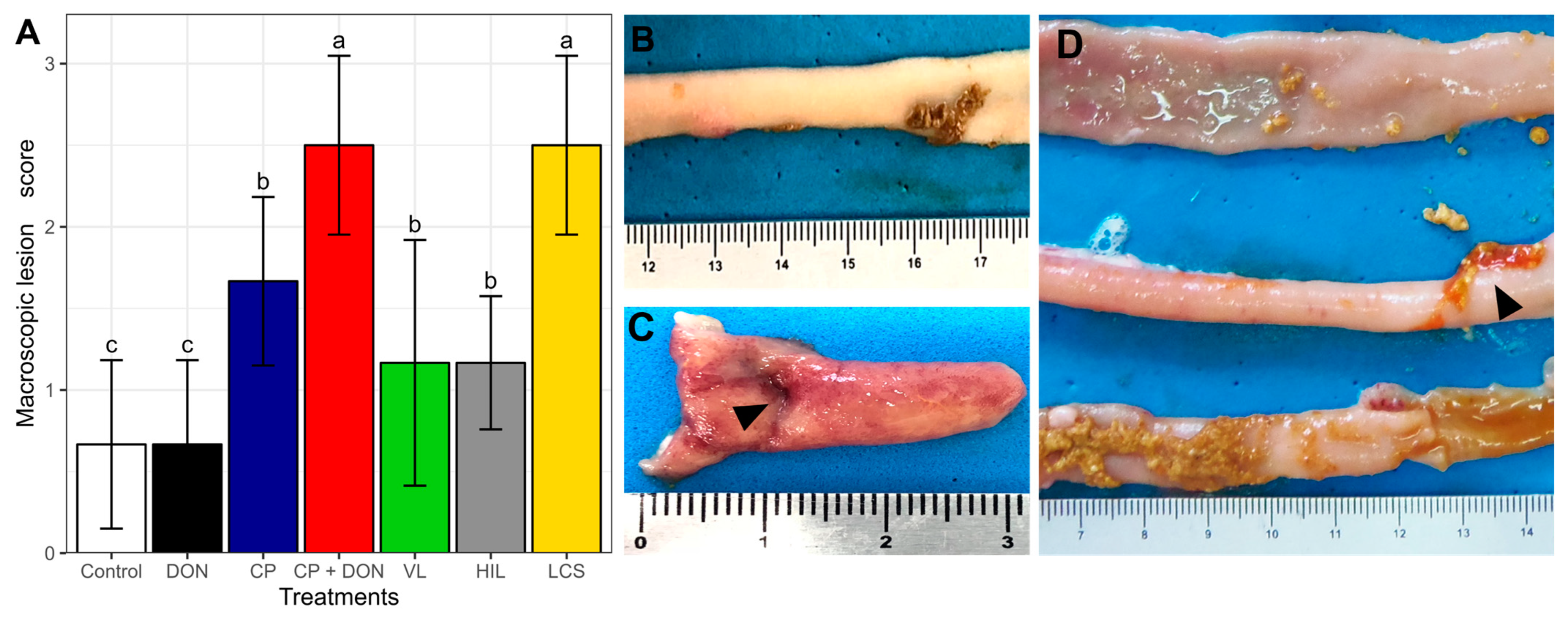
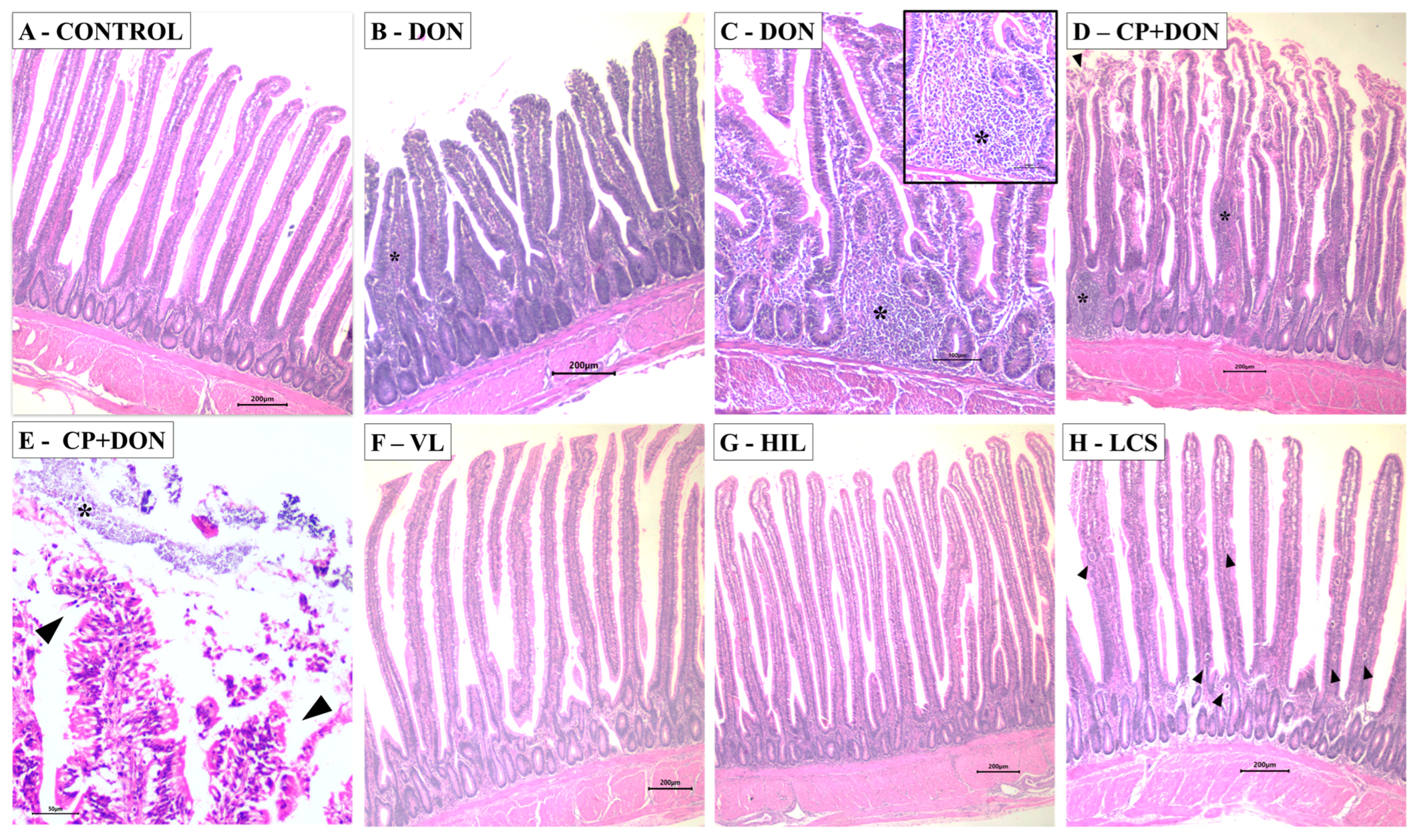
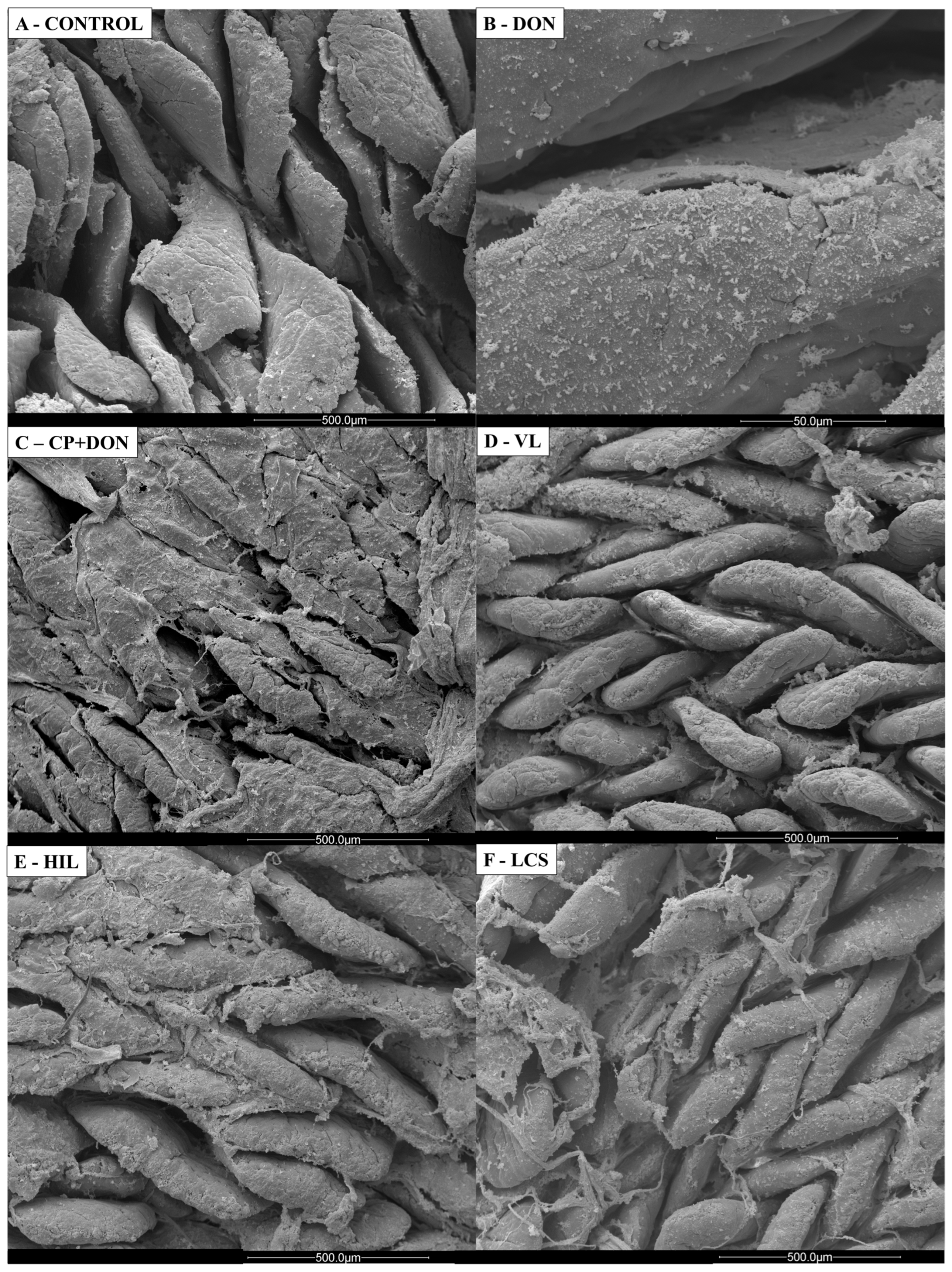
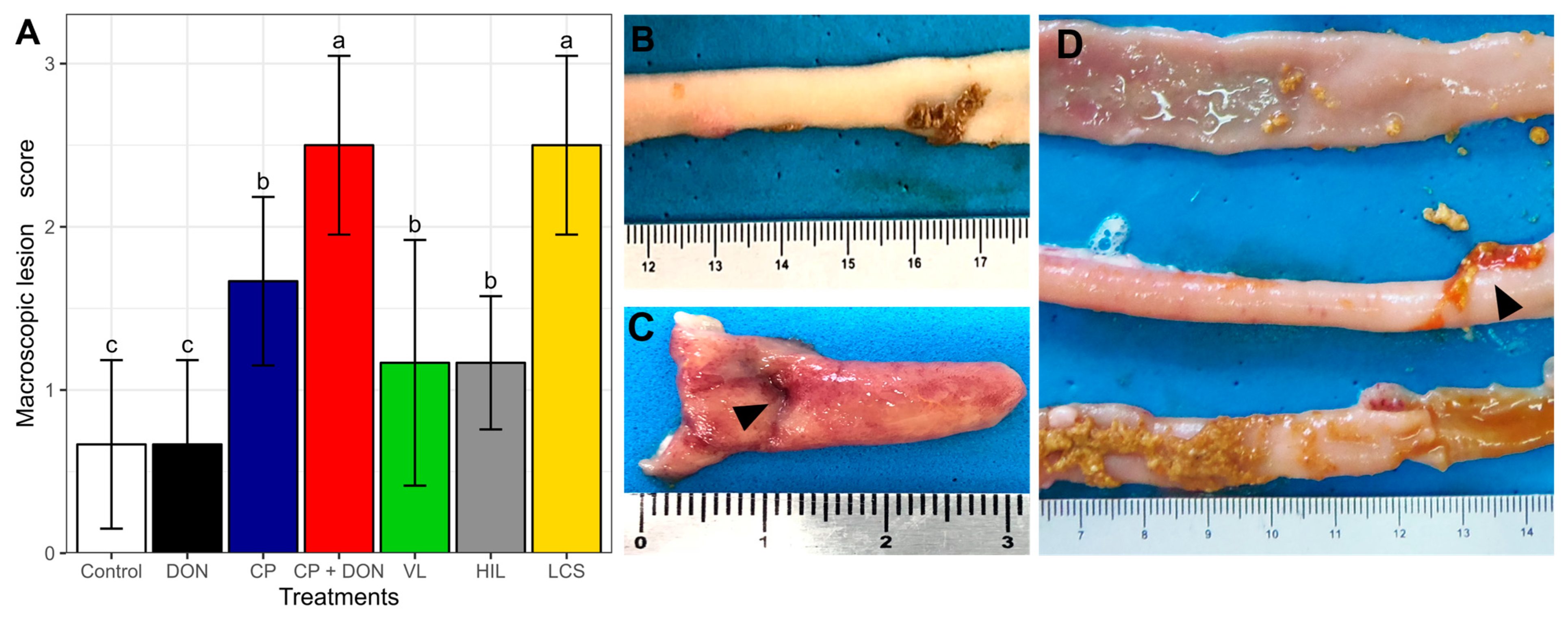
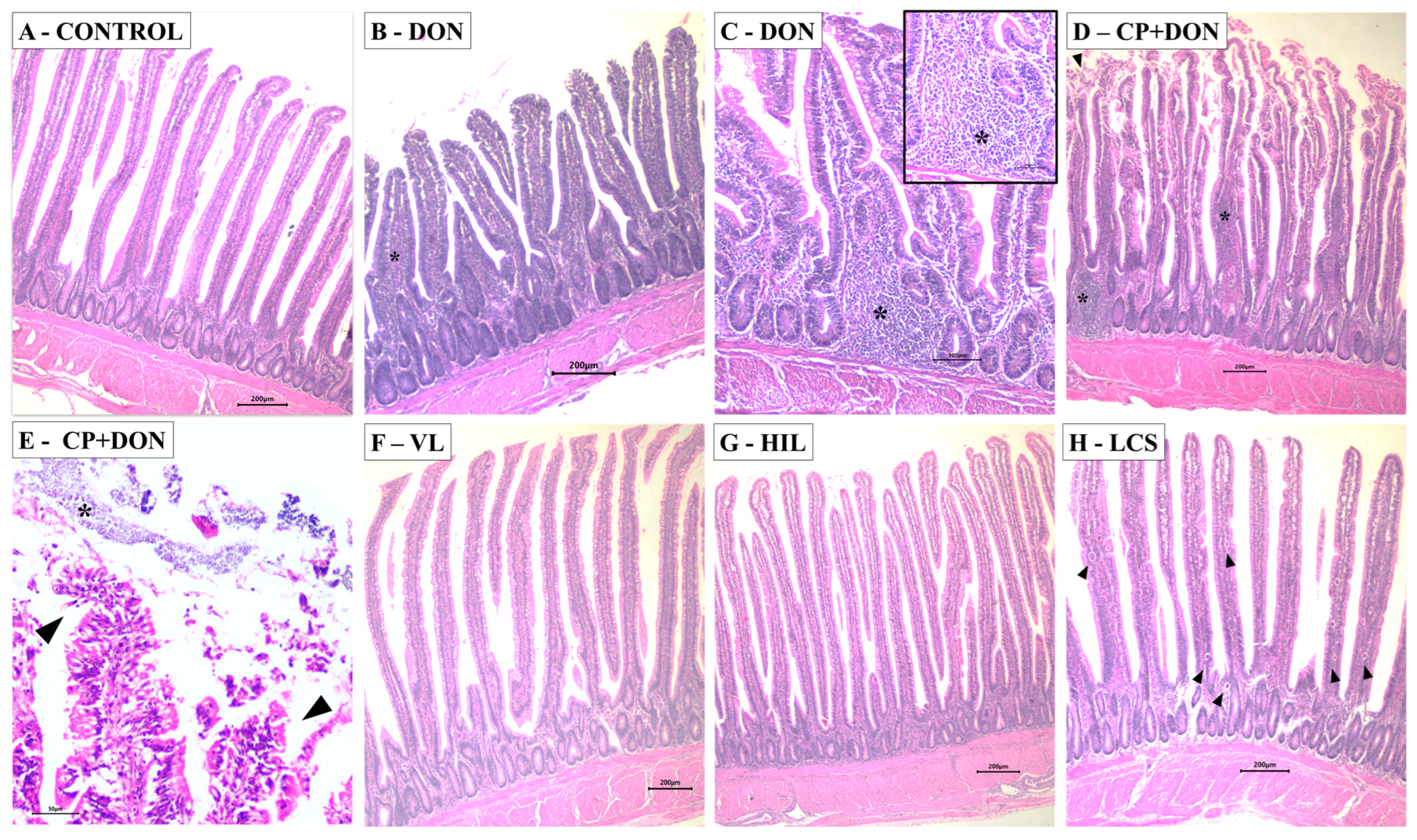
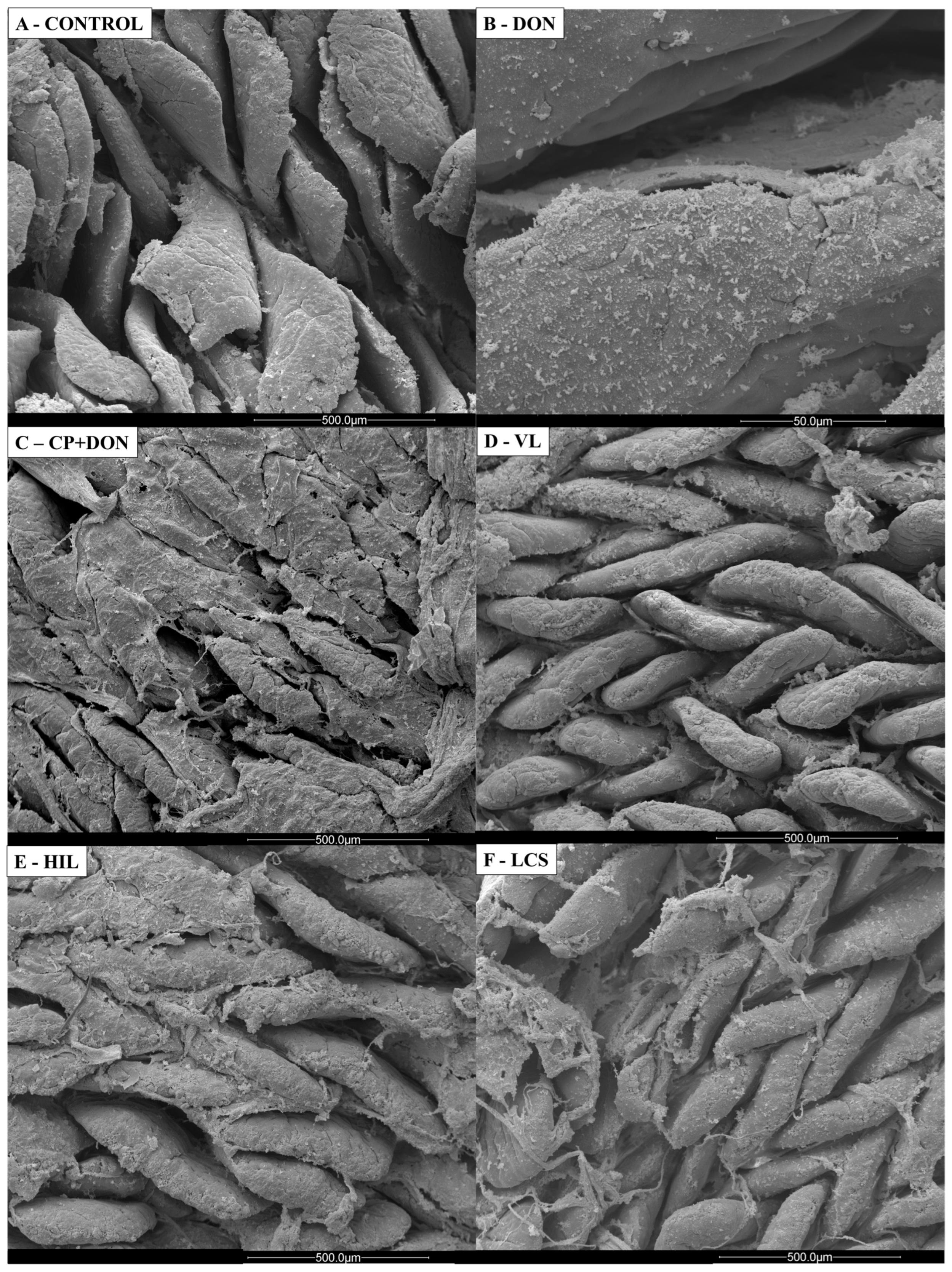
2.1. Effects of DON, C. perfringens, and Lactobacillus spp. Mixture on the Intestines
2.2. Effects of DON, C. perfringens, and Lactobacillus spp. Mixture on Redox Status
2.3. Effects of DON, C. perfringens, and Lactobacillus spp. on Intestinal Secretory IgA Levels
3. Discussion
4. Conclusions
5. Materials and Methods
5.1. Study Location and Ethical Approval
5.2. Animals and Treatments
5.3. Diets
5.4. Necrotic Enteritis Induction
5.5. Lactobacillus spp. Mixture Administration
5.6. Sample Collection
- Macroscopic intestinal lesion score
- ELISA
- Histology and scanning electron microscopy
- Oxidative stress response
5.7. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Yegani, M.; Korver, D.R. Factors Affecting Intestinal Health in Poultry. Poult. Sci. 2008, 87, 2052–2063. [Google Scholar] [CrossRef]
- No 1831/2003; Regulation (EC) no 1831/2003 of the European Parliament and of the Council of 22 September 2003 on Additives for Use in Animal Nutrition. FAO: Rome, Italy, 2003; L 268/30, p. 15.
- MAPA. Instrução Normativa Nº 1, de 13 de Janeiro de 2020; Ministério da Agricultura Pecuária e Abastecimento: Brasília, Brazil, 2020. [Google Scholar]
- Van Immerseel, F.; Lyhs, U.; Pedersen, K.; Prescott, J.F. Recent breakthroughs have unveiled the many knowledge gaps in Clostridium perfringens-associated necrotic enteritis in chickens: The first International Conference on Necrotic Enteritis in Poultry. Avian Pathol. 2016, 45, 269–270. [Google Scholar] [CrossRef] [PubMed]
- Kulkarni, R.R.; Gaghan, C.; Gorrell, K.; Sharif, S.; Taha-Abdelaziz, K. Probiotics as Alternatives to Antibiotics for the Prevention and Control of Necrotic Enteritis in Chickens. Pathogens 2022, 11, 692. [Google Scholar] [CrossRef] [PubMed]
- Abd El-Hack, M.E.; El-Saadony, M.T.; Elbestawy, A.R.; El-Shall, N.A.; Saad, A.M.; Salem, H.M.; El-Tahan, A.M.; Khafaga, A.F.; Taha, A.E.; AbuQamar, S.F.; et al. Necrotic enteritis in broiler chickens: Disease characteristics and prevention using organic antibiotic alternatives—A comprehensive review. Poult. Sci. 2022, 101, 101590. [Google Scholar] [CrossRef] [PubMed]
- Mwangi, S.; Timmons, J.; Fitz-coy, S.; Parveen, S. Characterization of Clostridium perfringens recovered from broiler chicken affected by necrotic enteritis. Poult. Sci. 2019, 98, 128–135. [Google Scholar] [CrossRef] [PubMed]
- Dierick, E.; Hirvonen, O.P.; Haesebrouck, F.; Ducatelle, R.; Van Immerseel, F.; Goossens, E. Rapid growth predisposes broilers to necrotic enteritis. Avian Pathol. 2019, 48, 416–422. [Google Scholar] [CrossRef]
- Antonissen, G.; Van Immerseel, F.; Pasmans, F.; Ducatelle, R.; Haesebrouck, F.; Timbermont, L.; Verlinden, M.; Janssens, G.P.J.; Eeckhaut, V.; Eeckhout, M.; et al. The Mycotoxin Deoxynivalenol Predisposes for the Development of Clostridium perfringens-Induced Necrotic Enteritis in Broiler Chickens. PLoS ONE 2014, 9, 1–8. [Google Scholar] [CrossRef]
- Wade, B.; Keyburn, A. The True Cost of Necrotic Enteritis. Available online: https://www.poultryworld.net/poultry/the-true-cost-of-necrotic-enteritis/ (accessed on 30 March 2023).
- Paterson, R.R.M.; Lima, N. How will climate change affect mycotoxins in food? Food Res. Int. 2010, 43, 1902–1914. [Google Scholar] [CrossRef]
- Shanmugasundaram, R.; Adams, D.; Ramirez, S.; Murugesan, G.R.; Applegate, T.J.; Cunningham, S.; Pokoo-Aikins, A.; Glenn, A.E. Subclinical Doses of Combined Fumonisins and Deoxynivalenol Predispose Clostridium perfringens–Inoculated Broilers to Necrotic Enteritis. Front. Physiol. 2022, 13, 934660. [Google Scholar] [CrossRef]
- Guo, F.; Wang, F.; Ma, H.; Ren, Z.; Yang, X.; Yang, X. Study on the interactive effect of deoxynivalenol and Clostridium perfringens on the jejunal health of broiler chickens. Poult. Sci. 2021, 100, 100807. [Google Scholar] [CrossRef] [PubMed]
- BIOMIN. BIOMIN Mycotoxin Survey Q3 2021 Results. Available online: https://www.biomin.net/science-hub/biomin-mycotoxin-survey-q3-2021-results/ (accessed on 30 March 2023).
- Von Buchholz, J.S.; Ruhnau, D.; Hess, C.; Aschenbach, J.R.; Hess, M.; Awad, W.A. Paracellular intestinal permeability of chickens induced by DON and/or C. jejuni is associated with alterations in tight junction mRNA expression. Microb. Pathog. 2022, 168, 105509. [Google Scholar] [CrossRef]
- Souza, M.; Baptista, A.A.S.; Valdiviezo, M.J.J.; Justino, L.; Menck-Costa, M.F.; Ferraz, C.R.; da Gloria, E.M.; Verri, W.A.; Bracarense, A.P.F.R.L. Lactobacillus spp. reduces morphological changes and oxidative stress induced by deoxynivalenol on the intestine and liver of broilers. Toxicon 2020, 185, 203–212. [Google Scholar] [CrossRef] [PubMed]
- Osselaere, A.; Santos, R.; Hautekiet, V.; De Backer, P.; Chiers, K.; Ducatelle, R.; Croubels, S. Deoxynivalenol Impairs Hepatic and Intestinal Gene Expression of Selected Oxidative Stress, Tight Junction and Inflammation Proteins in Broiler Chickens, but Addition of an Adsorbing Agent Shifts the Effects to the Distal Parts of the Small Intestine. PLoS ONE 2013, 8, e69014. [Google Scholar] [CrossRef] [PubMed]
- Jia, B.; Lin, H.; Yu, S.; Liu, N.; Yu, D.; Wu, A. Mycotoxin deoxynivalenol-induced intestinal flora disorders, dysfunction and organ damage in broilers and pigs. J. Hazard. Mater. 2023, 451, 131172. [Google Scholar] [CrossRef]
- Awad, W.A.; Zentek, J. The feed contaminant deoxynivalenol affects the intestinal barrier permeability through inhibition of protein synthesis. Arch. Toxicol. 2015, 89, 961–965. [Google Scholar] [CrossRef]
- Grenier, B.; Dohnal, I.; Shanmugasundaram, R.; Eicher, S.D.; Selvaraj, R.K.; Schatzmayr, G.; Applegate, T.J. Susceptibility of broiler chickens to coccidiosis when fed subclinical doses of deoxynivalenol and fumonisins—Special emphasis on the immunological response and the mycotoxin interaction. Toxins 2016, 8, 231. [Google Scholar] [CrossRef]
- Cremon, C.; Barbaro, M.R.; Ventura, M.; Barbara, G. Pre- and probiotic overview. Curr. Opin. Pharm. 2018, 43, 87–92. [Google Scholar] [CrossRef]
- Martyniak, A.; Medyńska-Przęczek, A.; Wędrychowicz, A.; Skoczeń, S.; Tomasik, P.J. Prebiotics, Probiotics, Synbiotics, Paraprobiotics and Postbiotic Compounds in IBD. Biomolecules 2021, 11, 1903. [Google Scholar] [CrossRef]
- Aguilar-Toalá, J.E.; Garcia-Varela, R.; Garcia, H.S.; Mata-Haro, V.; González-Córdova, A.F.; Vallejo-Cordoba, B.; Hernández-Mendoza, A. Postbiotics: An evolving term within the functional foods field. Trends Food Sci. Technol. 2018, 75, 105–114. [Google Scholar] [CrossRef]
- Maidana, L.; Souza, M.; Bracarense, A.P.F.R.L. Lactobacillus plantarum and Deoxynivalenol Detoxification: A Concise Review. J. Food Prot. 2022, 85, 1815–1823. [Google Scholar] [CrossRef] [PubMed]
- Gong, L.; Wang, B.; Zhou, Y.; Tang, L.; Zeng, Z.; Zhang, H.; Li, W. Protective Effects of Lactobacillus plantarum 16 and Paenibacillus polymyxa 10 Against Clostridium perfringens Infection in Broilers. Front. Immunol. 2020, 11, 628374. [Google Scholar] [CrossRef]
- Cao, L.; Wu, X.H.; Bai, Y.L.; Wu, X.Y.; Gu, S.B. Anti-inflammatory and antioxidant activities of probiotic powder containing Lactobacillus plantarum 1.2567 in necrotic enteritis model of broiler chickens. Livest. Sci. 2019, 223, 157–163. [Google Scholar] [CrossRef]
- Abd El-Ghany, W.A.; Abdel-Latif, M.A.; Hosny, F.; Alatfeehy, N.M.; Noreldin, A.E.; Quesnell, R.R.; Chapman, R.; Sakai, L.; Elbestawy, A.R. Comparative efficacy of postbiotic, probiotic, and antibiotic against necrotic enteritis in broiler chickens. Poult. Sci. 2022, 101, 101988. [Google Scholar] [CrossRef] [PubMed]
- Schedle, K.; Pfaffl, M.W.; Plitzner, C.; Meyer, H.H.D.; Windisch, W. Effect of insoluble fibre on intestinal morphology and mRNA expression pattern of inflammatory, cell cycle and growth marker genes in a piglet model. Arch. Anim. Nutr. 2008, 62, 427–438. [Google Scholar] [CrossRef]
- Kaer, L.V.; Olivares-Villagomez, D. Development, homeostasis, and functions of intestinal intraepithelial lymphocytes. J. Immunol. 2018, 200, 2235–2244. [Google Scholar] [CrossRef] [PubMed]
- Halliwell, B. Biochemistry of oxidative stress. Biochem. Soc. Trans. 2007, 35, 1147–1150. [Google Scholar] [CrossRef]
- Van Loo, J. How Chicory Fructans Contribute to Zootechnical Performance and Well-Being in Livestock and Companion Animals. J. Nutr. 2007, 137, 2594S–2597S. [Google Scholar] [CrossRef]
- Swatson, H.K.; Gous, R.; Iji, P.A.; Zarrinkalam, R. Effect of dietary protein level, amino acid balance and feeding level on growth, gastrointestinal tract, and mucosal structure of the small intestine in broiler chickens. Anim. Res. 2002, 51, 501–515. [Google Scholar] [CrossRef]
- Ghareeb, K.; Awad, W.A.; Bohm, J.; Zebeli, Q. Impacts of the feed contaminant deoxynivalenol on the intestine of monogastric animals: Poultry and swine. J. Appl. Toxicol. 2015, 35, 327–337. [Google Scholar] [CrossRef]
- Allaart, J.G.; de Bruijn, N.D.; van Asten, A.J.A.M.; Fabri, T.H.F.; Gröne, A. NetB-producing and beta2-producing Clostridium perfringens associated with subclinical necrotic enteritis in laying hens in the Netherlands. Avian Pathol. 2012, 41, 541–546. [Google Scholar] [CrossRef] [PubMed]
- Wang, A.; Hogan, N.S. Performance effects of feed-borne Fusarium mycotoxins on broiler chickens: Influences of timing and duration of exposure. Anim. Nutr. 2019, 5, 32–40. [Google Scholar] [CrossRef]
- Nii, T.; Shinkoda, T.; Isobe, N.; Yoshimura, Y. Intravaginal injection of Lactobacillus johnsonii may modulates oviductal microbiota and mucosal barrier function of laying hens. Poult. Sci. 2023, 102, 102699. [Google Scholar] [CrossRef]
- Ni, Y.; Zhang, Y.; Zheng, L.; Rong, N.; Yang, Y.; Gong, P.; Yang, Y.; Siwu, X.; Zhang, C.; Zhu, L.; et al. Bifidobacterium and Lactobacillus improve inflammatory bowel disease in zebrafish of different ages by regulating the intestinal mucosal barrier and microbiota. Life Sci. 2023, 324, 121699. [Google Scholar] [CrossRef]
- Moore, R.J. Necrotic enteritis predisposing factors in broiler chickens. Avian Pathol. 2016, 45, 275–281. [Google Scholar] [CrossRef] [PubMed]
- Luis, A.S.; Hansson, G.C. Intestinal mucus and their glycans: A habitat for thriving microbiota. Cell Host Microbe 2023, 31, 1087–1100. [Google Scholar] [CrossRef] [PubMed]
- Yang, X.; Li, L.; Duan, Y.; Yang, X. Antioxidant activity of Lactobacillus plantarum JM113 in vitro and its protective effect on broiler chickens challenged with deoxynivalenol. J. Anim. Sci. 2017, 95, 837–846. [Google Scholar]
- Awad, W.A.; Ghareeb, K.; Dadak, A.; Hess, M.; Böhm, J. Single and combined effects of deoxynivalenol mycotoxin and a microbial feed additive on lymphocyte DNA damage and oxidative stress in broiler chickens. PLoS ONE 2014, 9, 1–6. [Google Scholar] [CrossRef]
- Silva, E.O.; Bracarense, A.P.F.L.; Oswald, I.P. Mycotoxins and oxidative stress: Where are we? World Mycotoxin J. 2018, 11, 113–134. [Google Scholar] [CrossRef]
- Baehner, R.L.; Boxer, L.A.; Davis, J. The biochemical basis of nitroblue tetrazolium reduction in normal human and chronic granulomatous disease polymorphonuclear leukocytes. Blood 1976, 48, 309–313. [Google Scholar] [CrossRef]
- Zhou, M.; Zeng, D.; Ni, X.; Tu, T.; Yin, Z.; Pan, K.; Jing, B. Effects of Bacillus licheniformis on the growth performance and expression of lipid metabolism-related genes in broiler chickens challenged with Clostridium perfringens-induced necrotic enteritis. Lipids Health Dis. 2016, 15, 48. [Google Scholar] [CrossRef] [PubMed]
- El-Houseiny, W.; Khalil, A.A. The effects of toxigenic Clostridium perfringens types A and D on survival, as well as innate immune, inflammatory and oxidative stress responses in Nile tilapia. Aquaculture 2020, 529, 735694. [Google Scholar] [CrossRef]
- Benzie, I.F.F.; Strain, J.J. The Ferric Reducing Ability of Plasma (FRAP) as a Measure of “Antioxidant Power”: The FRAP Assay. Anal. Biochem. 1996, 239, 70–76. [Google Scholar] [CrossRef] [PubMed]
- Maresca, M. From the gut to the brain: Journey and pathophysiological effects of the food-associated trichothecene mycotoxin deoxynivalenol. Toxins 2013, 5, 784–820. [Google Scholar] [CrossRef]
- Yu, H.; Zhou, T.; Gong, J.; Young, C.; Su, X.; Li, X.-Z.; Zhu, H.; Tsao, R.; Yang, R. Isolation of deoxynivalenol-transforming bacteria from the chicken intestines using the approach of PCR-DGGE guided microbial selection. BMC Microbiol. 2010, 10, 182–191. [Google Scholar] [CrossRef]
- Yunus, A.; Blajet-Kosicka, A.; Kosicki, R.; Khan, M.; Rehman, H.; Böhm, J. Deoxynivalenol as a contaminant of broiler feed: Intestinal development, absorptive functionality, and metabolism of the mycotoxin. Poult. Sci. 2012, 91, 852–861. [Google Scholar] [CrossRef]
- Dänicke, S.; Brezina, U. Kinetics and metabolism of the Fusarium toxin deoxynivalenol in farm animals: Consequences for diagnosis of exposure and intoxication and carry over. Food Chem. Toxicol. 2013, 60, 58–75. [Google Scholar] [CrossRef]
- Chen, S.S.; Li, Y.-H.; Lin, M.-F. Chronic Exposure to the Fusarium Mycotoxin Deoxynivalenol: Impact on Performance, Immune Organ, and Intestinal Integrity of Slow-Growing Chickens. Toxins 2017, 9, 334. [Google Scholar] [CrossRef]
- Franco, T.S.; Garcia, S.; Hirooka, E.Y.; Ono, Y.S.; dos Santos, J.S. Lactic acid bacteria in the inhibition of Fusarium graminearum and deoxynivalenol detoxification. J. Appl. Microbiol. 2011, 111, 739–748. [Google Scholar] [CrossRef]
- Zhai, Y.; Hu, S.; Zhong, L.; Lu, Z.; Bie, X.; Zhao, H.; Zhang, C.; Lu, F. Characterization of deoxynivalenol detoxification by Lactobacillus paracasei LHZ-1 isolated from yogurt. J. Food Prot. 2019, 82, 1292–1299. [Google Scholar] [CrossRef]
- Qu, R.; Jiang, C.; Wu, W.; Pang, B.; Lei, S.; Lian, Z.; Shao, D.; Jin, M.; Shi, J. Conversion of DON to 3-epi-DON in vitro and toxicity reduction of DON in vivo by Lactobacillus rhamnosus. Food Funct. 2019, 10, 2785–2796. [Google Scholar]
- El-Nezami, H.S.; Chrevatidis, A.; Auriola, S.; Salminen, S.; Mykkanen, H. Removal of common Fusarium toxins in vitro by strains of Lactobacillus and Propionibacterium. Food Addit. Contam. 2002, 19, 680–686. [Google Scholar] [CrossRef] [PubMed]
- Shojadoost, B.; Alizadeh, M.; Boodhoo, N.; Astill, J.; Karimi, S.H.; Shoja Doost, J.; Taha-Abdelaziz, K.; Kulkarni, R.; Sharif, S. Effects of Treatment with Lactobacilli on Necrotic Enteritis in Broiler Chickens. Probiotics Antimicrob. Proteins 2022, 14, 1110–1129. [Google Scholar] [CrossRef] [PubMed]
- Qing, X.; Zeng, D.; Wang, H.; Ni, X.; Liu, L.; Lai, J.; Khalique, A.; Pan, K.; Jing, B. Preventing subclinical necrotic enteritis through Lactobacillus johnsonii BS15 by ameliorating lipid metabolism and intestinal microflora in broiler chickens. AMB Express 2017, 7, 139. [Google Scholar] [CrossRef]
- Emami, N.K.; White, M.B.; Calik, A.; Kimminau, E.A.; Dalloul, R.A. Managing broilers gut health with antibiotic-free diets during subclinical necrotic enteritis. Poult. Sci. 2021, 100, 101055. [Google Scholar] [CrossRef]
- Adhikari, P.; Kiess, A.; Adhikari, R.; Jha, R. An approach to alternative strategies to control avian coccidiosis and necrotic enteritis. J. Appl. Poult. Res. 2020, 29, 515–534. [Google Scholar] [CrossRef]
- Aviagen. Ross Manual de Manejo de Frangos de Corte 2018; Aviagen: Huntsville, AL, USA, 2018. [Google Scholar]
- Drew, M.D.; Syed, N.A.; Goldade, B.G.; Laarveld, B.; Van Kessel, A.G. Effects of Dietary Protein Source and Level on Intestinal Populations of Clostridium perfringens in Broiler Chickens. Poult. Sci. 2004, 83, 414–420. [Google Scholar] [CrossRef]
- Sivaseelan, S.; Vijayakumar, S.; Malmarugan, S.; Balachandran, P.; Balasubramaniam, G.A. Assessment of predisposing effect of coccidiosis to necrotic enteritis in broiler chickens. Vet. Arh. 2013, 83, 653–664. [Google Scholar]
- Shojadoost, B.; Vince, A.R.; Prescott, J.F. The successful experimental induction of necrotic enteritis in chickens by Clostridium perfringens: a critical review. Vet. Res. 2012, 43, 74. [Google Scholar] [CrossRef]
- Rocha, T.S.; Baptista, A.A.S.; Donato, T.C.; Milbradt, E.L.; Okamoto, A.S.; Andreatti Filho, R.L. Identification and adhesion profile of Lactobacillus spp. strains isolated from poultry. Braz. J. Microbiol. 2014, 45, 1065–1073. [Google Scholar] [CrossRef]
- Maidana, L.G.; Gerez, J.; Hohmann, M.N.S.; Verri, W.A.; Bracarense, A.P.F.L. Lactobacillus plantarum metabolites reduce deoxynivalenol toxicity on jejunal explants of piglets. Toxicon 2021, 203, 12–21. [Google Scholar] [CrossRef] [PubMed]
- Cravens, R.L.; Goss, G.R.; Chi, F.; De Boer, E.D.; Davis, S.W.; Hendrix, S.M.; Richardson, J.A.; Johnston, S.L. The effects of necrotic enteritis, aflatoxin B1, and virginiamycin on growth performance, necrotic enteritis lesion scores, and mortality in young broilers. Poult. Sci. 2013, 92, 1997–2004. [Google Scholar] [CrossRef] [PubMed]
- Souza, M.d.; Cicero, C.E.; Menck-Costa, M.F.; Justino, L.; Gerez, J.R.; Baptista, A.A.S.; Bracarense, A.P.F.R.L. Histological evaluation of the intestine of broiler chickens: Comparison of three sampling methods. Semina Ciências Agrárias 2021, 42, 3247–3258. [Google Scholar] [CrossRef]
- Terciolo, C.; Bracarense, A.P.; Souto, P.C.M.C.; Cossalter, A.-M.; Dopavogui, L.; Loiseau, N.; Oliveira, C.A.F.; Pinton, P.; Oswald, I.P. Fumonisins at Doses below EU Regulatory Limits Induce Histological Alterations in Piglets. Toxins 2019, 11, 548. [Google Scholar] [CrossRef] [PubMed]
- Sedlak, J.; Lindsay, R.H. Estimation of total, protein-bound, and nonprotein sulfhydryl groups in tissue with Ellman’s reagent. Anal. Biochem. 1968, 25, 192–205. [Google Scholar] [CrossRef] [PubMed]
- Katalinic, V.; Modun, D.; Music, I.; Boban, M. Gender differences in antioxidant capacity of rat tissues determined by 2,2′-azinobis (3-ethylbenzothiazoline 6-sulfonate; ABTS) and ferric reducing antioxidant power (FRAP) assays. Comp. Biochem. Physiol. Part C: Toxicol. Pharmacol. 2005, 140, 47–52. [Google Scholar]
- Fattori, V.; Pinho-Ribeiro, F.A.; Borghi, S.M.; Alves-Filho, J.C.; Cunha, T.M.; Cunha, F.Q.; Casagrande, R.; Verri-Jr, W.A. Curcumin inhibits superoxide anion-induced pain-like behavior and leukocyte recruitment by increasing Nrf2 expression and reducing NF-κB activation. Inflammation Res. 2015, 64, 993–1003. [Google Scholar] [CrossRef]
- Sattler, W.; Malle, E.; Kostner, G.M. Methodological approaches for assessing lipid and protein oxidation and modification in plasma and isolated lipoproteins. Methods Mol. Biol. 1998, 110, 167–191. [Google Scholar]
- Manchope, M.F.; Artero, N.A.; Fattori, V.; Mizokami, S.S.; Pitol, D.L.; Issa, J.P.M.; Fukada, S.Y.; Cunha, T.M.; Alves-Filho, J.C.; Cunha, F.Q.; et al. Naringenin mitigates titanium dioxide (TiO2)-induced chronic arthritis in mice: Role of oxidative stress, cytokines, and NFκB. Inflammation Res. 2018, 67, 997–1012. [Google Scholar] [CrossRef]

{kind=link}
{kind=link}
{kind=link}
| Treatment | Villus Height [µm] | Crypt Depth [µm] | Villi: Crypt Ratio | Microscopic Lesion Score | IEL | Goblet Cells |
|---|---|---|---|---|---|---|
| Duodenum | ||||||
| Control | 1315.02 ± 189.85 | 145.91 ± 18.95 | 9.13 ± 1.78 | 7.50 b ± 1.22 | NA | NA |
| DON | 1216.70 ± 182.97 | 158.64 ± 19.96 | 7.84 ± 1.92 | 9.16 b ± 3.31 | NA | NA |
| CP | 1159.23 ± 202.50 | 133.11 ± 15.99 | 8.71 ± 1.19 | 11.83 a ± 5.81 | NA | NA |
| CP + DON | 1296.70 ± 258.16 | 148.60 ± 17.92 | 8.80 ± 1.93 | 15.40 a ± 3.71 | NA | NA |
| VL | 1299.06 ± 135.00 | 164.57 ± 43.96 | 8.22 ± 1.84 | 11.40 b ± 1.52 | NA | NA |
| HIL | 1352.86 ± 99.07 | 140.28 ± 13.89 | 9.76 ± 1.53 | 8.40 b ± 3.05 | NA | NA |
| LCS | 1180.26 ± 98.46 | 138.52 ± 14.33 | 8.55 ± 1.19 | 9.40 b ± 2.51 | NA | NA |
| Jejunum | ||||||
| Control | 970.52 a ± 159.33 | 133.87 ± 13.01 | 7.30 a ± 1.34 | 4.16 b ± 2.86 | 20.18 b ± 2.89 | NA |
| DON | 632.43 b ± 109.73 | 129.62 ± 12.69 | 4.89 b ± 0.83 | 8.16 a ± 1.47 | 25.68 b ± 2.66 | NA |
| CP | 840.82 b ± 232.02 | 126.74 ± 11.09 | 6.59 b ± 1.43 | 11.17 a ± 3.71 | 28.84 a ± 2.79 | NA |
| CP + DON | 789.68 b ± 77.20 | 127.23 ± 9.13 | 6.22 b ± 0.69 | 12.00 a ± 2.16 | 32.70 a ± 4.94 | NA |
| VL | 1075.19 a ± 154.24 | 122.39 ± 18.15 | 8.88 a ± 1.47 | 8.60 a ± 3.36 | 24.76 b ± 3.99 | NA |
| HIL | 961.53 a ± 225.59 | 131.70 ± 16.69 | 7.48 a ± 2.35 | 7.00 b ± 3.67 | 23.41 b ± 1.90 | NA |
| LCS | 901.30 a ± 94.33 | 113.33 ± 14.87 | 8.02a ± 0.96 | 9.00 a ± 3.46 | 30.26 a ± 5.77 | NA |
| Ileum | ||||||
| Control | 750.29 a ± 49.07 | 123.43 ± 16.56 | 6.20 ± 1.21 | 4.50 c ± 1.87 | 18.98 b ± 1.07 | 66.97 c ± 5.87 |
| DON | 605.68 b ± 59.83 | 128.91 ± 25.98 | 4.86 ± 1.02 | 6.50 b ± 1.87 | 23.52 a ± 2.23 | 73.63 c ± 5.19 |
| CP | 671.15 b ± 107.38 | 118.87 ± 12.92 | 5.64 ± 0.62 | 8.67 a ± 3.93 | 26.11 a ± 5.50 | 87.34 b ± 11.32 |
| CP + DON | 713.88 a ± 86.42 | 115.98 ± 11.15 | 6.16 ± 0.58 | 9.00 a ± 1.87 | 26.75 a ± 2.47 | 88.54 b ± 9.36 |
| VL | 729.05 a ± 52.30 | 137.01 ± 13.66 | 5.34 ± 0.40 | 4.20 c ± 1.79 | 22.46 a ± 2.05 | 108.04 a ± 9.29 |
| HIL | 703.70 a ± 42.50 | 131.71 ± 10.05 | 5.35 ± 0.26 | 9.20 a ± 2.28 | 24.26 a ± 4.27 | 105.21 a ± 5.83 |
| LCS | 731.06 a ± 38.13 | 131.85 ± 16.41 | 5.59 ± 0.54 | 6.80 b ± 2.49 | 24.11 a ± 2.55 | 104.81 a ± 10.18 |
| Treatment | TBARS | NBT | GSH | ABTS | FRAP |
|---|---|---|---|---|---|
| Jejunum | |||||
| Control | 0.04 b ± 0.03 | 6.67 b ± 3.19 | 2809.33 a ± 1215.38 | 0.70 ± 0.09 | 0.59 b ± 0.17 |
| DON | 0.08 a ± 0.05 | 7.56 b ± 2.30 | 1313.11 b ± 447.71 | 1.10 ± 0.18 | 0.75 b ± 0.25 |
| CP | 0.03 b ± 0.01 | 32.46 a ± 12.51 | 447.51 c ± 110.76 | 1.13 ± 0.44 | 1.65 a ± 0.56 |
| CP + DON | 0.05 a ± 0.01 | 24.51 a ± 13.54 | 458.97 c ± 20.38 | 1.11 ± 0.39 | 1.21 a ± 0.23 |
| VL | 0.03 b ± 0.01 | 32.85 a ± 27.97 | 593.57 c ± 75.59 | 1.00 ± 0.25 | 1.40 a ± 0.15 |
| HIL | 0.02 b ± 0.01 | 48.70 a ± 25.97 | 585.41 c ± 110.41 | 0.85 ± 0.25 | 1.55 a ± 0.33 |
| LCS | 0.03 b ± 0.01 | 25.55 a ± 16.27 | 400.20 c ± 107.88 | 1.37 ± 0.40 | 1.98 a ± 0.45 |
| Ileum | |||||
| Control | 0.02 ± 0.007 | 6.81 b ± 0.64 | 1568.72 a ± 731.15 | 0.81 ± 0.17 | 1.28 ± 0.94 |
| DON | 0.04 ± 0.03 | 6.48 b ± 3.46 | 1702.98 a ± 412.50 | 0.64 ± 0.19 | 0.96 ± 0.49 |
| CP | 0.02 ± 0.008 | 30.80 a ± 8.62 | 588.39 b ± 120.34 | 0.89 ± 0.36 | 1.78 ± 0.57 |
| CP + DON | 0.03 ± 0.02 | 44.17 a ± 14.05 | 556.43 b ± 111.85 | 0.75 ± 0.35 | 0.97 ± 0.21 |
| VL | 0.02 ± 0.01 | 39.59 a ± 14.33 | 639.80 b ± 250.44 | 0.88 ± 0.22 | 1.18 ± 0.30 |
| HIL | 0.03 ± 0.005 | 36.29 a ± 13.26 | 606.61 b ± 87.30 | 0.57 ± 0.34 | 1.41 ± 0.52 |
| LCS | 0.03 ± 0.009 | 26.03 a ± 17.26 | 663.84 b ± 137.50 | 0.59 ± 0.34 | 0.93 ± 0.23 |
| Ingredients (g/kg) | Diet 1 0–6 Days | Diet 2 7–14 Days | Diet 3 15–23 Days |
|---|---|---|---|
| Corn | 481.6 | 150 | 150 |
| Soya bean meal (46% CP) | 437 | 353.14 | - |
| Soybean oil | 32 | 54 | 54 |
| Wheat | - | 404.33 | 404.33 |
| Fishmeal | - | - | 353.14 |
| Dicalcium phosphate | 25 | 9.28 | 9.29 |
| Limestone | 1.5 | 13.17 | 13.16 |
| Sodium chloride | 6 | 5.03 | 5.04 |
| Premix | 5 | 5 | 5 |
| L-lysine HCL | 0.968 | 2.1 | 2.08 |
| DL-methionine | 3.36 | 3 | 2.99 |
| L-threonine | 0.38 | 0.98 | 0.98 |
| Nutritional Levels | |||
| Energy mcal/kg | 2975 | 3050 | 3010 |
| Protein (%) | 24.27 | 23.31 | 25.61 |
| Linoleic acid (%) | 3.355 | - | - |
| Calcium (%) | 0.971 | 0.878 | 2.910 |
| Phosphorus available (%) | 0.463 | 0.310 | 1.563 |
| Lysine dig (%) | 1.307 | 1.256 | 1.418 |
| Methionine dig (%) | 0.646 | 0.600 | 0.869 |
| Methionine + cistine dig (%) | 0.967 | 0.929 | 1.176 |
| Threonine dig (%) | 0.863 | 0.829 | 0.931 |
| Tryptophan dig (%) | 0.277 | 0.271 | 0.190 |
| Sodium (%) | 0.225 | 0.218 | 0.451 |
| Contamination Level [µg kg−1] | |||
|---|---|---|---|
| Mycotoxin | 1–6 Days Uncontaminated Diet | 7–23 Days Uncontaminated Diet | 7–23 Days Contaminated Diet |
| DON | <LOQ | 200 | 19,309.4 |
| AFB1 | <LOQ | <LOQ | <LOQ |
| AFB2 | <LOQ | <LOQ | <LOQ |
| AFG1 | <LOQ | <LOQ | <LOQ |
| AFG2 | <LOQ | <LOQ | <LOQ |
| FB1 | 252.9 | <LOQ | <LOQ |
| FB2 | <LOQ | <LOQ | <LOQ |
| ZEA | 31.4 | <LOQ | 4878.7 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
de Souza, M.; Baptista, A.A.S.; Menck-Costa, M.F.; Justino, L.; da Glória, E.M.; Shimizu, G.D.; Ferraz, C.R.; Verri, W.A.; Van Immerseel, F.; Bracarense, A.P.F.R.L. Modulation of Broiler Intestinal Changes Induced by Clostridium perfringens and Deoxynivalenol through Probiotic, Paraprobiotic, and Postbiotic Supplementation. Toxins 2024, 16, 46. https://doi.org/10.3390/toxins16010046
de Souza M, Baptista AAS, Menck-Costa MF, Justino L, da Glória EM, Shimizu GD, Ferraz CR, Verri WA, Van Immerseel F, Bracarense APFRL. Modulation of Broiler Intestinal Changes Induced by Clostridium perfringens and Deoxynivalenol through Probiotic, Paraprobiotic, and Postbiotic Supplementation. Toxins. 2024; 16(1):46. https://doi.org/10.3390/toxins16010046
Chicago/Turabian Stylede Souza, Marielen, Ana Angelita Sampaio Baptista, Maísa Fabiana Menck-Costa, Larissa Justino, Eduardo Micotti da Glória, Gabriel Danilo Shimizu, Camila Rodrigues Ferraz, Waldiceu A. Verri, Filip Van Immerseel, and Ana Paula Frederico Rodrigues Loureiro Bracarense. 2024. "Modulation of Broiler Intestinal Changes Induced by Clostridium perfringens and Deoxynivalenol through Probiotic, Paraprobiotic, and Postbiotic Supplementation" Toxins 16, no. 1: 46. https://doi.org/10.3390/toxins16010046
APA Stylede Souza, M., Baptista, A. A. S., Menck-Costa, M. F., Justino, L., da Glória, E. M., Shimizu, G. D., Ferraz, C. R., Verri, W. A., Van Immerseel, F., & Bracarense, A. P. F. R. L. (2024). Modulation of Broiler Intestinal Changes Induced by Clostridium perfringens and Deoxynivalenol through Probiotic, Paraprobiotic, and Postbiotic Supplementation. Toxins, 16(1), 46. https://doi.org/10.3390/toxins16010046