Tetrodotoxin and Its Analogues (TTXs) in the Food-Capture and Defense Organs of the Palaeonemertean Cephalothrix cf. simula


Abstract
1. Introduction
2. Results
2.1. General Results of HPLC–MS/MS
2.2. Body (Without Proboscis)
2.3. Proboscis
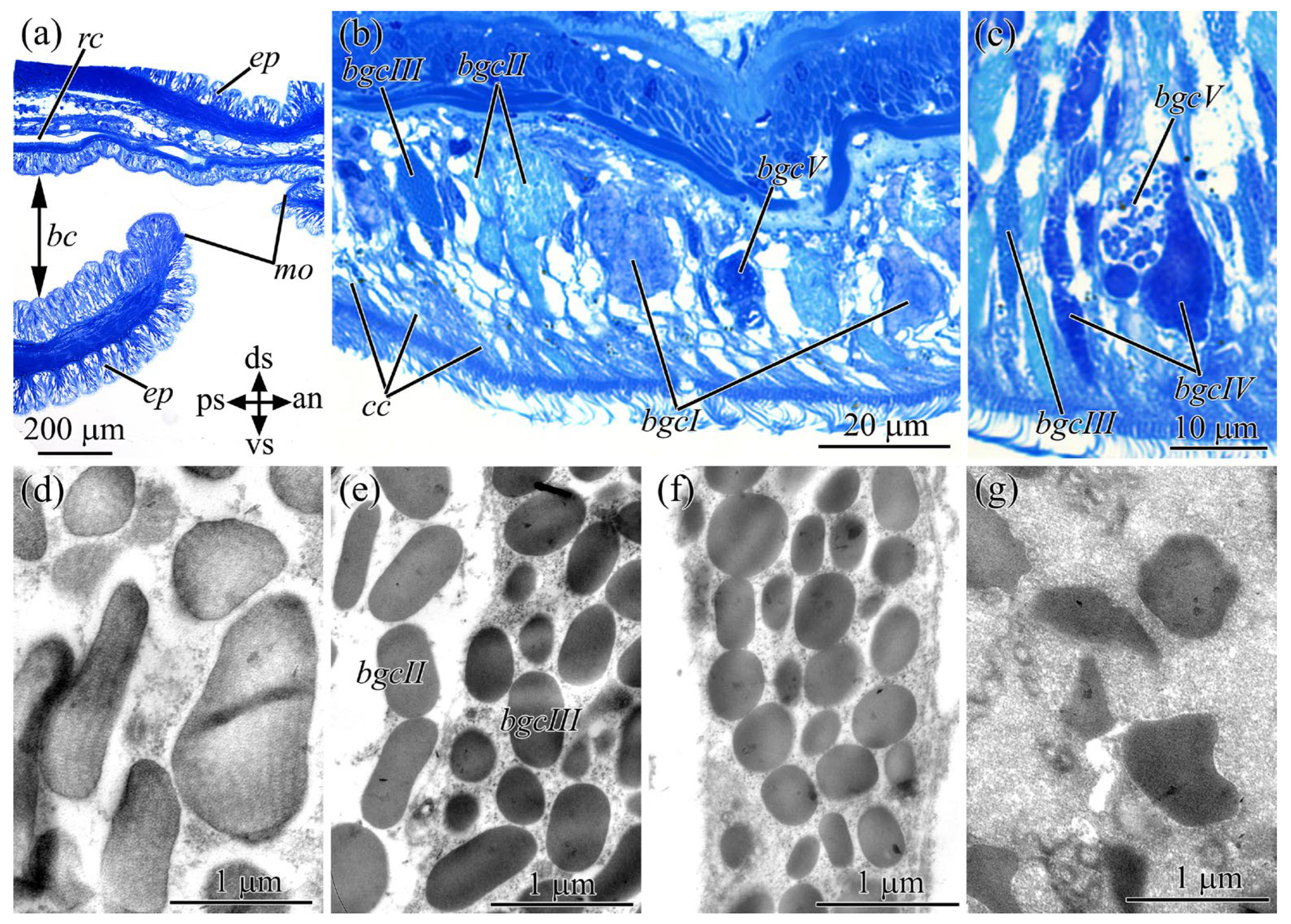
2.4. Buccal Cavity
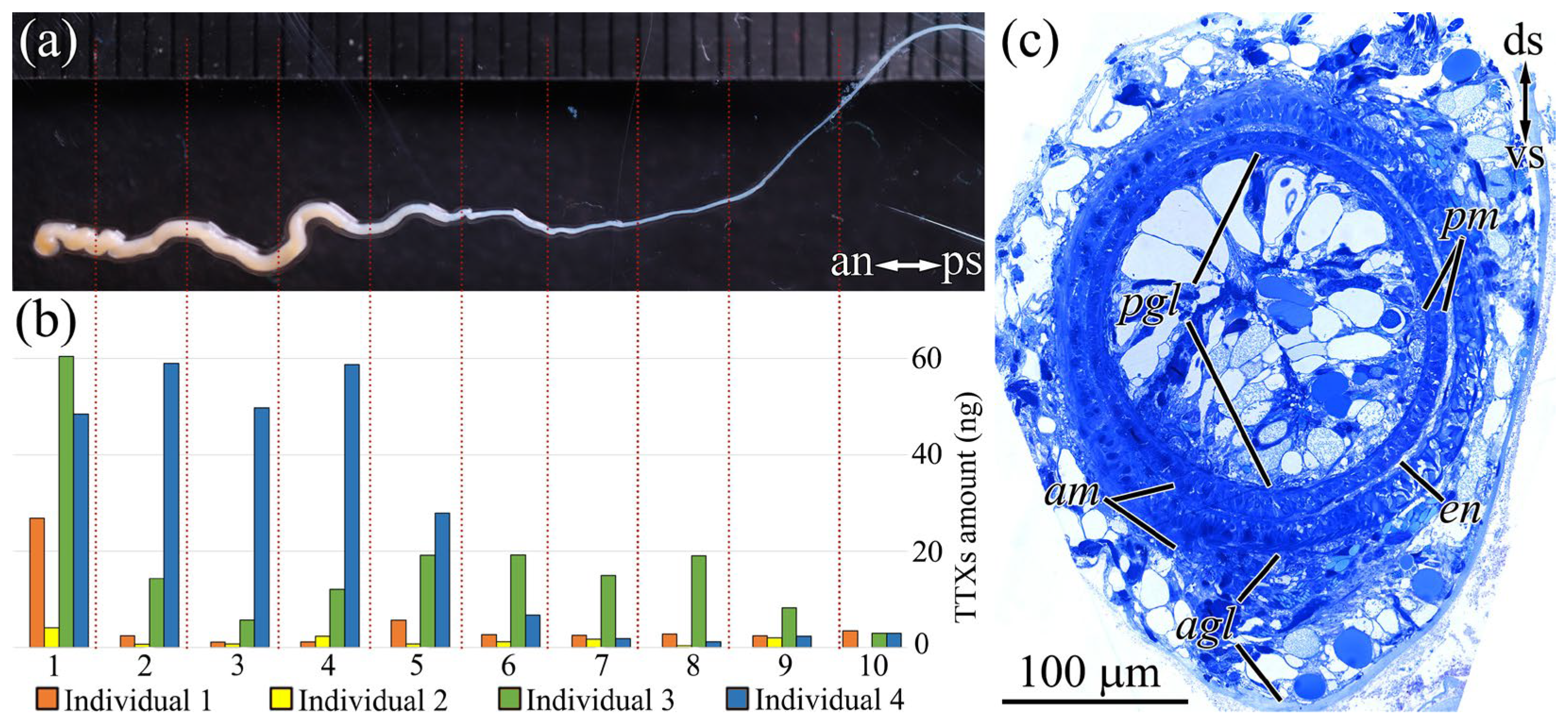
2.5. Cephalic Gland
2.6. In Vivo Feeding Behavior
3. Discussion
4. Materials and Methods
4.1. Nemertans Collection
4.2. CLSM and Morphological Studies
4.3. Extraction and Analysis of TTX and Its Analogues
4.4. Feeding Experiments
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Kajihara, H.; Sun, S.C.; Chernyshev, A.V.; Chen, H.X.; Ito, K.; Asakawa, M.; Maslakova, S.A.; Norenburg, J.L.; Strand, M.; Sundberg, P.; et al. Taxonomic identity of a tetrodotoxin-accumulating ribbon-worm Cephalothrix simula (Nemertea: Palaeonemertea): A species artificially introduced from the pacific to Europe. Zoolog. Sci. 2013, 30, 985–997. [Google Scholar] [CrossRef] [PubMed]
- Faasse, M.A.; Turbeville, J.M. The first record of the north-west Pacific nemertean Cephalothrix simula in northern Europe. Mar. Biodivers. Rec. 2015, 8, 6–11. [Google Scholar] [CrossRef]
- Turner, A.D.; Fenwick, D.; Powell, A.; Dhanji-Rapkova, M.; Ford, C.; Hatfield, R.G.; Santos, A.; Martinez-Urtaza, J.; Bean, T.P.; Baker-Austin, C.; et al. New invasive nemertean species (Cephalothrix simula) in England with high levels of tetrodotoxin and a microbiome linked to toxin metabolism. Mar. Drugs 2018, 16, 452. [Google Scholar] [CrossRef] [PubMed]
- Ali, A.E.; Arakawa, O.; Noguchi, T.; Miyazawa, K.; Shida, Y.; Hashimoto, K. Tetrodotoxin and related substances in a ribbon worm Cephalothrix linearis (Nemertean). Toxicon 1990, 28, 1083–1093. [Google Scholar] [CrossRef] [PubMed]
- Asakawa, M.; Toyoshima, T.; Shida, Y.; Noguchi, T.; Miyazawa, K. Paralytic toxins in a ribbon worm Cephalothrix species (Nemertean) adherent to cultured oysters in Hiroshima Bay, Hiroshima Prefecture, Japan. Toxicon 2000, 38, 763–773. [Google Scholar] [CrossRef]
- Asakawa, M.; Toyoshima, T.; Ito, K.; Bessho, K.; Yamaguchi, C.; Tsunetsugu, S.; Shida, Y.; Kajihara, H.; Mawatari, S.F.; Noguchi, T.; et al. Paralytic toxicity in the ribbon worm Cephalothrix species (Nemertea) in Hiroshima Bay, Hiroshima Prefecture, Japan and the isolation of tetrodotoxin as a main component of its toxins. Toxicon 2003, 41, 747–753. [Google Scholar] [CrossRef]
- Kajihara, H. Resolving a 200-year-old taxonomic conundrum: Neotype designation for Cephalothrix linearis (nemertea: Palaeonemertea) based on a topotype from Bergen, Norway. Fauna Nor. 2019, 39, 39–76. [Google Scholar] [CrossRef]
- Miyazawa, K.; Higashiyama, M.; Ito, K.; Noguchi, T.; Arakawa, O.; Shida, Y.; Hashimoto, K. Tetrodotoxin in two species of ribbon worm (Nemertini), Lineus fuscoviridis and Tubulanus punctatus. Toxicon 1988, 26, 867–874. [Google Scholar] [CrossRef]
- Tanu, M.B.; Mahmud, Y.; Arakawa, O.; Takatani, T.; Kajihara, H.; Kawatsu, K.; Hamano, Y.; Asakawa, M.; Miyazawa, K.; Noguchi, T. Immunoenzymatic visualization of tetrodotoxin (TTX) in Cephalothrix species (Nemertea: Anopla: Palaeonemertea: Cephalotrichidae) and Planocera reticulata (Platyhelminthes: Turbellaria: Polycladida: Planoceridae). Toxicon 2004, 44, 515–520. [Google Scholar] [CrossRef]
- Malykin, G.V.; Chernyshev, A.V.; Magarlamov, T.Y. Intrabody tetrodotoxin distribution and possible hypothesis for its migration in ribbon worms Cephalothrix cf. simula (Palaeonemertea, Nemertea). Mar. Drugs 2021, 19, 494. [Google Scholar] [CrossRef]
- Oaks, J.A. Ultrastructure of Lineus ruber (rhyncocoela) epidermis. Tissue Cell 1978, 10, 227–242. [Google Scholar] [CrossRef] [PubMed]
- Ferraris, J.D. Histological study of cephalic glands in selected nemertina. Trans. Am. Microsc. Soc. 1979, 93, 437–446. [Google Scholar] [CrossRef]
- Gibson, R. Nemertea. In Synopsis and Classification of Living Organisms; Parker, S.P., Ed.; McGraw-Hill: New York, NY, USA, 1982; pp. 823–846. [Google Scholar]
- Norenburg, J.L. Structure of the nemertine integument with consideration of its ecological and phylogenetic significance. Am. Zool. 1985, 25, 37–51. [Google Scholar] [CrossRef]
- Turbeville, J.M. Nemertinea. In Microscopic Anatomy of Invertebrates, Vol. 3. Platyhelminthes and Nemertinea; Harrison, W., Bogitsh, B.J., Eds.; Wiley-Liss: New York, NY, USA, 1991; pp. 285–328. [Google Scholar]
- Chernyshev, A.V. Comparative Morphology, Systematics and Phylogeny of the Nemerteans; Dalnauka: Vladivostok, Russia, 2011. [Google Scholar]
- Ling, E.A. The proboscis apparatus of the nemertine Lineus ruber. Philos. Trans. R. Soc. Lond. B 1971, 262, 1–22. [Google Scholar]
- Stricker, S.A.; Cloney, R.A. The ultarstructure of venom-producing cells in Paranemertes peregrina (Nemertea, Hoplonemertea). J. Morphol. 1983, 177, 89–107. [Google Scholar] [CrossRef]
- Vlasenko, A.E.; Magarlamov, T.Y. Tetrodotoxin and its analogues in Cephalothrix cf. simula (Nemertea: Palaeonemertea) from the Sea of Japan (Peter the Great Gulf): Intrabody distribution and secretions. Toxins 2020, 12, 745. [Google Scholar] [CrossRef]
- Chulanetra, M.; Sookrung, N.; Srimanote, P.; Indrawattana, N.; Thanongsaksrikul, J.; Sakolvaree, Y.; Chongsa-Nguan, M.; Kurazono, H.; Chaicumpa, W. Toxic marine puffer fish in Thailand seas and tetrodotoxin they contained. Toxins 2011, 3, 1249–1262. [Google Scholar] [CrossRef]
- Turner, A.D.; Powell, A.; Schofield, A.; Lees, D.N.; Baker-Austin, C. Detection of the pufferfish toxin tetrodotoxin in European bivalves, England, 2013 to 2014. Eurosurveillance 2015, 20, 21009. [Google Scholar] [CrossRef]
- Turner, A.D.; Dhanji-Rapkova, M.; Coates, L.; Bickerstaff, L.; Milligan, S.; O’Neill, A.; Faulkner, D.; McEneny, H.; Baker-Austin, C.; Lees, D.N.; et al. Detection of tetrodotoxin shellfish poisoning (TSP) toxins and causative factors in bivalve molluscs from the UK. Mar. Drugs 2017, 15, 277. [Google Scholar] [CrossRef]
- Gerssen, A.; Bovee, T.H.F.; Klijnstra, M.D.; Poelman, M.; Portier, L.; Hoogenboom, R.L.A.P. First report on the occurrence of tetrodotoxins in bivalve mollusks in the Netherlands. Toxins 2018, 10, 450. [Google Scholar] [CrossRef]
- Thuy, L.V.; Yamamoto, S.; Kawaura, R.; Takemura, N.; Yamaki, K.; Yasumoto, K.; Takada, K.; Watabe, S.; Sato, S. Tissue distribution of tetrodotoxin and its analogs in Lagocephalus pufferfish collected in Vietnam. Fish. Sci. 2020, 86, 1101–1110. [Google Scholar] [CrossRef]
- Costa, P.R.; Giráldez, J.; Rodrigues, S.M.; Leão, J.M.; Pinto, E.; Soliño, L.; Gago-Martínez, A. High Levels of tetrodotoxin (TTX) in trumpet shell Charonia lampas from the Portuguese coast. Toxins 2021, 13, 250. [Google Scholar] [CrossRef] [PubMed]
- Dhanji-Rapkova, M.; Turner, A.D.; Baker-Austin, C.; Huggett, J.F.; Ritchie, J.M. Distribution of tetrodotoxin in pacific oysters (Crassostrea gigas). Mar. Drugs 2021, 19, 84. [Google Scholar] [CrossRef] [PubMed]
- Matsui, T.; Taketsugu, S.; Sato, H.; Yamamori, K.; Kodama, K.; Ishii, A.; Hirose, H.; Shimizu, C. Toxification of cultured puffer fish by the administration of tetrodotoxin producing bacteria. Nippon Suisan Gakkaishi 1990, 56, 705. [Google Scholar] [CrossRef]
- Kono, M.; Matsui, T.; Furukawa, K.; Yotsu-Yamashita, M.; Yamamori, K. Accumulation of tetrodotoxin and 4,9-anhydrotetrodotoxin in cultured juvenile kusafugu Fugu niphobles by dietary administration of natural toxic komonfugu Fugu poecilonotus liver. Toxicon 2008, 51, 1269–1273. [Google Scholar] [CrossRef] [PubMed]
- Okita, K.; Yamazaki, H.; Sakiyama, K.; Yamane, H.; Niina, S.; Takatani, T.; Arakawa, O.; Sakakura, Y. Puffer smells tetrodotoxin. Ichthyol. Res. 2013, 60, 386–389. [Google Scholar] [CrossRef]
- Itoi, S.; Ishizuka, K.; Mitsuoka, R.; Takimoto, N.; Yokoyama, N.; Detake, A.; Takayanagi, C.; Yoshikawa, S.; Sugita, H. Seasonal changes in the tetrodotoxin content of the pufferfish Takifugu niphobles. Toxicon 2016, 114, 53–58. [Google Scholar] [CrossRef]
- Tatsuno, R.; Gao, W.; Ibi, K.; Mine, T.; Okita, K.; Nishihara, G.N.; Takatani, T.; Arakawa, O. Profile differences in tetrodotoxin transfer to skin and liver in the pufferfish Takifugu rubripes. Toxicon 2017, 130, 73–78. [Google Scholar] [CrossRef]
- Gao, W.; Kanahara, Y.; Yamada, M.; Tatsuno, R.; Yoshikawa, H.; Doi, H.; Takatani, T.; Arakawa, O. Contrasting toxin selectivity between the marine pufferfish Takifugu pardalis and the freshwater pufferfish Pao suvattii. Toxins 2019, 11, 470. [Google Scholar] [CrossRef]
- Kosker, A.R.; Özogul, F.; Ayas, D.; Durmus, M.; Ucar, Y.; Regenstein, J.M.; Özogul, Y. Tetrodotoxin levels of three pufferfish species (Lagocephalus sp.) caught in the North-Eastern Mediterranean sea. Chemosphere 2019, 219, 95–99. [Google Scholar] [CrossRef]
- Akbora, H.D.; Kunter, İ.; Erçetin, T.; Elagöz, A.M.; Çiçek, B.A. Determination of tetrodotoxin (TTX) levels in various tissues of the silver cheeked puffer fish (Lagocephalus sceleratus (Gmelin, 1789)) in Northern Cyprus Sea (Eastern Mediterranean). Toxicon 2020, 175, 1–6. [Google Scholar] [CrossRef] [PubMed]
- Ritson-Williams, R.; Yotsu-Yamashita, M.; Paul, V.J. Ecological functions of tetrodotoxin in a deadly polyclad flatworm. Proc. Natl. Acad. Sci. USA 2006, 103, 3176–3179. [Google Scholar] [CrossRef] [PubMed]
- Vlasenko, A.E.; Kuznetsov, V.G.; Malykin, G.V.; Pereverzeva, A.O.; Velansky, P.V.; Yakovlev, K.V.; Magarlamov, T.Y. Tetrodotoxins secretion and voltage-gated sodium channel adaptation in the ribbon worm Kulikovia alborostrata (Takakura, 1898) (Nemertea). Toxins 2021, 13, 606. [Google Scholar] [CrossRef] [PubMed]
- Cardall, B.L.; Brodie, E.D., Jr.; Brodie, E.D., III; Hanifin, C.T. Secretion and regeneration of tetrodotoxin in the rough-skin newt (Taricha granulosa). Toxicon 2004, 44, 933–938. [Google Scholar] [CrossRef]
- Matsumoto, T.; Nagashima, Y.; Kusuhara, H.; Sugiyama, Y.; Ishizaki, S.; Shimakura, K.; Shiomi, K. Involvement of carrier-mediated transport system in uptake of tetrodotoxin into liver tissue slices of puffer fish Takifugu rubripes. Toxicon 2007, 50, 173–179. [Google Scholar] [CrossRef]
- Hwang, D.F.; Arakawa, O.; Saito, T.; Noguchi, T.; Simidu, U.; Tsukamoto, K.; Shida, Y.; Hashimoto, K. Tetrodotoxin-producing bacteria from the blue-ringed octopus Octopus maculosus. Mar. Biol. 1989, 100, 327–332. [Google Scholar] [CrossRef]
- Asakawa, M.; Matsumoto, T.; Umezaki, K.; Kaneko, K.; Yu, X.; Gomez-Delan, G.; Tomano, S.; Noguchi, T.; Ohtsuka, S. Toxicity and toxin composition of the greater blue-ringed octopus Hapalochlaena lunulata from ishigaki island, Okinawa prefecture, Japan. Toxins 2019, 11, 245. [Google Scholar] [CrossRef]
- Malykin, G.V.; Velansky, P.V.; Melnikova, D.I.; Magarlamov, T.Y. Tetrodotoxins in larval development of ribbon worm Cephalothrix cf. simula (Palaeonemertea, Nemertea). Mar. Biotechnol. 2023, 25, 918–934. [Google Scholar] [CrossRef]
- Boie, H.-J. Die Paketdrüsenzone von Lineus rüber O. F. Müller (Nemertini). Z. Morphol. Okol. Tiere 1952, 41, 188–222. [Google Scholar] [CrossRef]
- Kem, W.R. Structure and action of nemertine toxins. Integr. Comp. Biol. 1985, 25, 99–111. [Google Scholar] [CrossRef]
- Wang, H.; Sun, S.; Li, Q. Laboratory observations on the feeding behavior and feeding rate of the nemertean Procephalothrix simulus. Biol. Bull. 2008, 214, 166–175. [Google Scholar] [CrossRef] [PubMed]
- Chen, H.; Strand, M.; Norenburg, J.L.; Sun, S.; Kajihara, H.; Chernyshev, A.V.; Maslakova, S.A.; Sundberg, P. Statistical parsimony networks and species assemblages in cephalotrichid nemerteans (Nemertea). PLoS ONE 2010, 5, e12885. [Google Scholar] [CrossRef]
- Sagorny, C.; Wesseler, C.; Krämer, D.; von Döhren, J. Assessing the diversity and distribution of Cephalothrix species (Nemertea: Palaeonemertea) in European waters by comparing different species delimitation methods. J. Zool. Syst. Evol. Res. 2019, 57, 497–519. [Google Scholar] [CrossRef]
- Bane, V.; Brosnan, B.; Barnes, P.; Lehane, M.; Furey, A. High-resolution mass spectrometry analysis of tetrodotoxin (TTX) and its analogues in puffer fish and shellfish. Food Addit. Contam. Part A 2016, 33, 1468–1489. [Google Scholar] [CrossRef] [PubMed]
- Vlasenko, A.E.; Velansky, P.V.; Chernyshev, A.V.; Kuznetsov, V.G.; Magarlamov, T.Y. Tetrodotoxin and its analogues profile in nemertean species from the Sea of Japan. Toxicon 2018, 156, 48–51. [Google Scholar] [CrossRef]
- Vale, P. Complex profiles of hydrophobic paralytic shellfish poisoning compounds in Gymnodinium catenatum identified by liquid chromatography with fluorescence detection and mass spectrometry. J. Chromatogr. A 2008, 1195, 85–93. [Google Scholar] [CrossRef] [PubMed]
- Kudo, Y.; Yasumoto, T.; Konoki, K.; Cho, Y.; Yotsu-Yamashita, M. Isolation and structural determination of the first 8-epi-type tetrodotoxin analogs from the newt, Cynops ensicauda popei, and comparison of tetrodotoxin analogs profiles of this newt and the puffer fish, Fugu poecilonotus. Mar. Drugs 2012, 10, 655–667. [Google Scholar] [CrossRef]
- Bane, V.; Lehane, M.; Dikshit, M.; O’Riordan, A.; Furey, A. Tetrodotoxin: Chemistry, toxicity, source, distribution and detection. Toxins 2014, 6, 693–755. [Google Scholar] [CrossRef]
- Puilingi, C.G.; Kudo, Y.; Cho, Y.; Konoki, K.; Yotsu-Yamashita, M. Tetrodotoxin and its analogues in the pufferfish Arothron hispidus and A. nigropunctatus from the Solomon Islands: A comparison of their toxin profiles with the same species from Okinawa, Japan. Toxins 2015, 7, 3436–3454. [Google Scholar] [CrossRef]
- Turner, A.D.; Boundy, M.J.; Rapkova, M.D. Development and single-laboratory validation of a liquid chromatography tandem mass spectrometry method for quantitation of tetrodotoxin in mussels and oysters. J. AOAC Int. 2017, 100, 1469–1482. [Google Scholar] [CrossRef]
- Chen, X.W.; Liu, H.X.; Jin, Y.B.; Li, S.F.; Bi, X.; Chung, S.; Zhang, S.S.; Jiang, Y.Y. Separation, identification and quantification of tetrodotoxin and its analogs by LC-MS without calibration of individual analogs. Toxicon 2011, 57, 938–943. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Individual No. | Weight, g | Sex | Region | Organ/Segment | Toxins | ||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| ng * | % | ||||||||||||||||||
| TTX | 4-epiTTX | 11-norTTX-6(r)-ol | 11-norTTX-6(s)-ol | 4,9-anhydroTTX | 5-deoxyTTX | 11-deoxyTTX | 5,6,11-trideoxyTTX | 11-oxoTTX | Total of TTXs | Total of TTXs in Regions | Total Amount of TTXs | % of Amount in Region | % of TTXs in Organ/Section in Total Amount | % of TTXs in Region of Total Amount | |||||
| 1 | 0.2 | Female | Body | Internal organs | 78,651.9 | 323.4 | 11.5 | <0.02 | 203.9 | 1610.2 | 1160.4 | 30,903.6 | 10.2 | 112,875.2 | 167,216.9 | 172,836.7 | 67.5 | 65.3 | 96.7 |
| Body wall | 38,724.7 | 270.2 | 3.2 | <0.02 | 124.3 | 1077.8 | 639.5 | 13,501.4 | 0.6 | 54,341.7 | 32.5 | 31.4 | |||||||
| Bc + Cg | Internal organs | 2897.5 | 24.9 | <0.02 | 1.4 | 9.8 | 148.5 | 27.1 | 515 | 8.9 | 3633 | 5533.1 | 65.7 | 2.1 | 3.2 | ||||
| Body wall | 1406.3 | 13.5 | <0.02 | <0.02 | 3.9 | 78.8 | 13.6 | 383.9 | <0.02 | 1 900 | 34.3 | 1.1 | |||||||
| Proboscis | Anterior segment | 27.2 | 0.6 | <0.02 | 0.7 | 0.1 | 6.5 | 0.3 | 13 | 6.4 | 54.8 | 86.8 | 63.1 | 0.0 | 0.1 | ||||
| Posterior segment | 10.4 | 0.4 | <0.02 | <0.06 | <0.06 | 8.2 | 0.2 | 11.9 | 0.9 | 32 | 36.9 | 0.0 | |||||||
| 2 | 0.07 | Female | Body | Internal organs | 1023.1 | 142.8 | 0.4 | 4.4 | 48.8 | 282.3 | 64.7 | 605.4 | 19.3 | 2191.1 | 16,230.8 | 19,162.0 | 13.5 | 11.4 | 84.7 |
| Body wall | 7382.3 | 604.4 | 0.2 | 6.2 | 166.8 | 256.3 | 144.5 | 5478.9 | <0.02 | 14,039.7 | 86.5 | 73.3 | |||||||
| Bc | Internal organs | 57.3 | 3.5 | <0.02 | 0.1 | 1.1 | 4.5 | 1.2 | 30.6 | 1.7 | 100 | 2040.9 | 4.9 | 0.5 | 10.7 | ||||
| Body wall | 1123.2 | 55.2 | <0.02 | 1.2 | 7.3 | 40.5 | 16.9 | 696.6 | <0.02 | 1940.9 | 95.1 | 10.1 | |||||||
| Cg | Cg | 36.7 | 2 | <0.02 | 0.2 | 0.6 | 4.5 | 0.8 | 25.5 | <0.02 | 70.4 | 870.8 | 8.1 | 0.4 | 4.5 | ||||
| Body wall | 282.2 | 88.0 | 0.1 | 1.3 | 24.6 | 47.5 | 5.7 | 351.1 | <0.02 | 800.4 | 91.9 | 4.2 | |||||||
| Proboscis | Anterior segment | 2.3 | 0.2 | 0.2 | 0.3 | 0.1 | 0.5 | <0.06 | 4.5 | 5.2 | 13.3 | 19.5 | 66.3 | 0.1 | 0.1 | ||||
| Posterior segment | 1.8 | 0.1 | <0.02 | 0.4 | <0.02 | 0.3 | <0.02 | 0.5 | 3.1 | 6.2 | 33.7 | 0.0 | |||||||
| 3 | 0.09 | Male | Body | Internal organs | 393.4 | 20.2 | 0.2 | 2.6 | 4.4 | 195.5 | 31.9 | 85.1 | 11 | 744.4 | 1659 | 2030.7 | 44.9 | 36.7 | 81.7 |
| Body wall | 238.6 | 43.2 | 0.4 | 3.3 | 11.1 | 293.2 | 79.8 | 245.1 | <0.02 | 914.7 | 55.1 | 45.0 | |||||||
| Bc | Internal organs | 23.4 | 0.9 | <0.02 | 0.1 | 0.2 | 8.9 | 1.6 | 4.7 | 0.8 | 40.6 | 130.2 | 31.2 | 2.0 | 6.4 | ||||
| Body wall | 21.5 | 4.1 | <0.06 | 0.4 | 0.6 | 23 | 7.3 | 32.6 | <0.02 | 89.6 | 68.8 | 4.4 | |||||||
| Proboscis | Anterior segment | 49.9 | 3.1 | 0.4 | 14 | 0.7 | 28.5 | 2.2 | 17.2 | 36.6 | 152.5 | 241.5 | 63.2 | 7.5 | 11.9 | ||||
| Posterior segment | 28.5 | 1.4 | 0.2 | 10.7 | 0.3 | 17.6 | 1.6 | 9.7 | 19 | 89 | 36.8 | 4.4 | |||||||
| 4 | 0.11 | Male | Body | Internal organs | 715.3 | 37.8 | 1.3 | 0.8 | 6.6 | 617 | 24.3 | 240.7 | 1.4 | 1645 | 3314.1 | 4990.8 | 49.6 | 33.0 | 63.6 |
| Body wall | 444.1 | 117.3 | 2.4 | 2 | 9 | 652.2 | 78.5 | 363.6 | <0.02 | 1669.1 | 50.4 | 33.4 | |||||||
| Bc | Internal organs | 63.1 | 3.7 | 0.1 | <0.06 | 0.6 | 48.7 | 2.4 | 44 | <0.02 | 162.5 | 331.9 | 49 | 3.3 | 6.4 | ||||
| Body wall | 38 | 14.5 | 0.4 | 0.2 | 0.7 | 57.4 | 10.2 | 48 | <0.02 | 169.3 | 51 | 3.4 | |||||||
| Cg | Cg | 199.3 | 22.4 | 0.4 | 0.7 | 8.7 | 91.1 | 3.7 | 68.2 | <0.02 | 394.6 | 953.6 | 41.4 | 7.9 | 18.3 | ||||
| Body wall | 161.3 | 48.7 | 2.5 | 1.8 | 12.0 | 205.1 | 22.7 | 104.9 | <0.02 | 559 | 58.6 | 11.2 | |||||||
| Proboscis | Anterior segment | 206.2 | 6.1 | 0.9 | 6.8 | 2.2 | 58.3 | 3.9 | 68.3 | 4.9 | 357.4 | 391.3 | 91.3 | 7.2 | 11.7 | ||||
| Posterior segment | 11.5 | 0.5 | 0.1 | 0.2 | 0.2 | 10.1 | 0.3 | 10.7 | 0.3 | 33.9 | 8.7 | 0.7 | |||||||
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Malykin, G.V.; Velansky, P.V.; Magarlamov, T.Y. Tetrodotoxin and Its Analogues (TTXs) in the Food-Capture and Defense Organs of the Palaeonemertean Cephalothrix cf. simula. Toxins 2024, 16, 43. https://doi.org/10.3390/toxins16010043
Malykin GV, Velansky PV, Magarlamov TY. Tetrodotoxin and Its Analogues (TTXs) in the Food-Capture and Defense Organs of the Palaeonemertean Cephalothrix cf. simula. Toxins. 2024; 16(1):43. https://doi.org/10.3390/toxins16010043
Chicago/Turabian StyleMalykin, Grigorii V., Peter V. Velansky, and Timur Yu. Magarlamov. 2024. "Tetrodotoxin and Its Analogues (TTXs) in the Food-Capture and Defense Organs of the Palaeonemertean Cephalothrix cf. simula" Toxins 16, no. 1: 43. https://doi.org/10.3390/toxins16010043
APA StyleMalykin, G. V., Velansky, P. V., & Magarlamov, T. Y. (2024). Tetrodotoxin and Its Analogues (TTXs) in the Food-Capture and Defense Organs of the Palaeonemertean Cephalothrix cf. simula. Toxins, 16(1), 43. https://doi.org/10.3390/toxins16010043