Inhibition of CYP1A1 Alleviates Colchicine-Induced Hepatotoxicity

Abstract
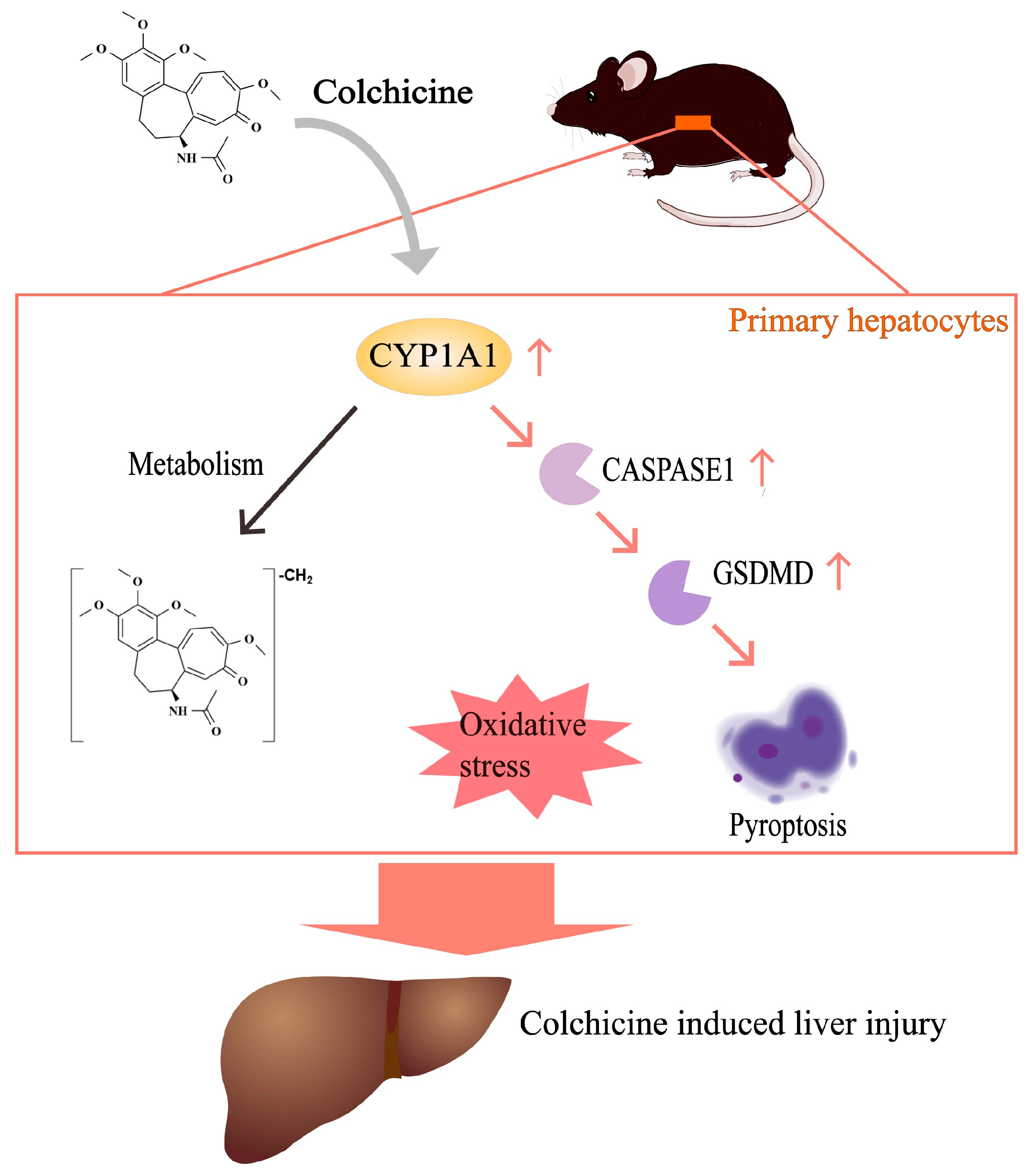
1. Introduction
2. Results
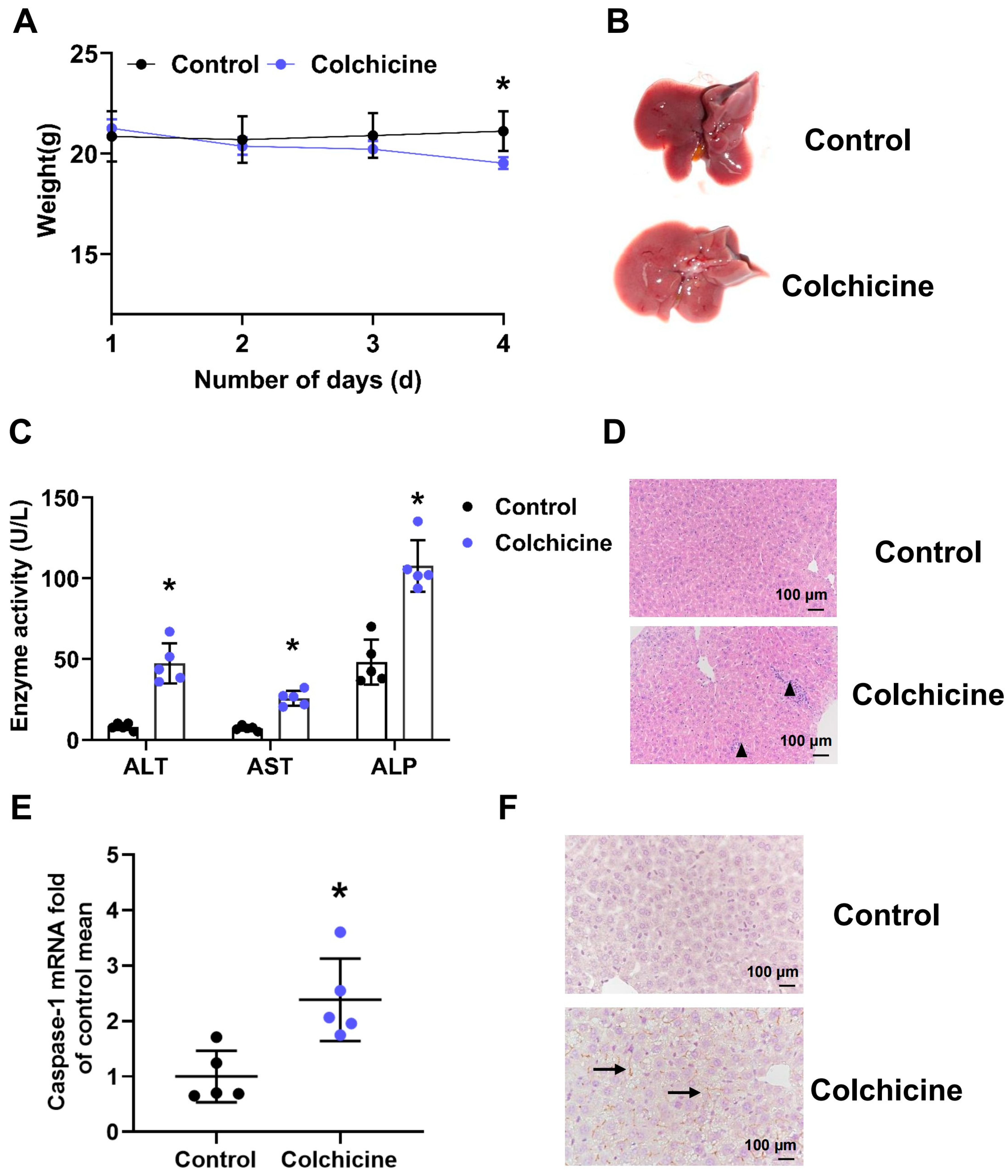
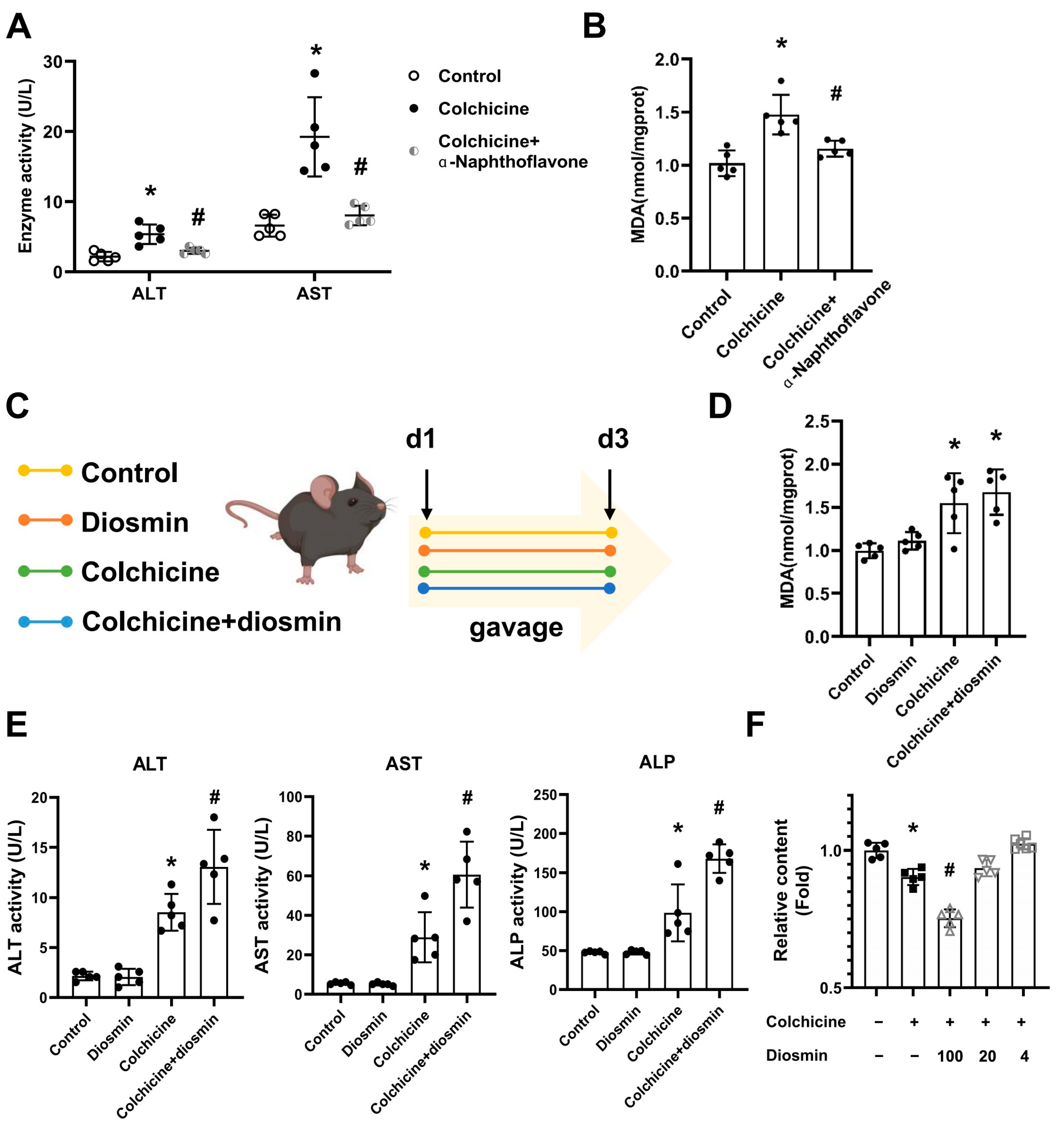
2.1. Liver Injury Induced by Colchicine in Mice
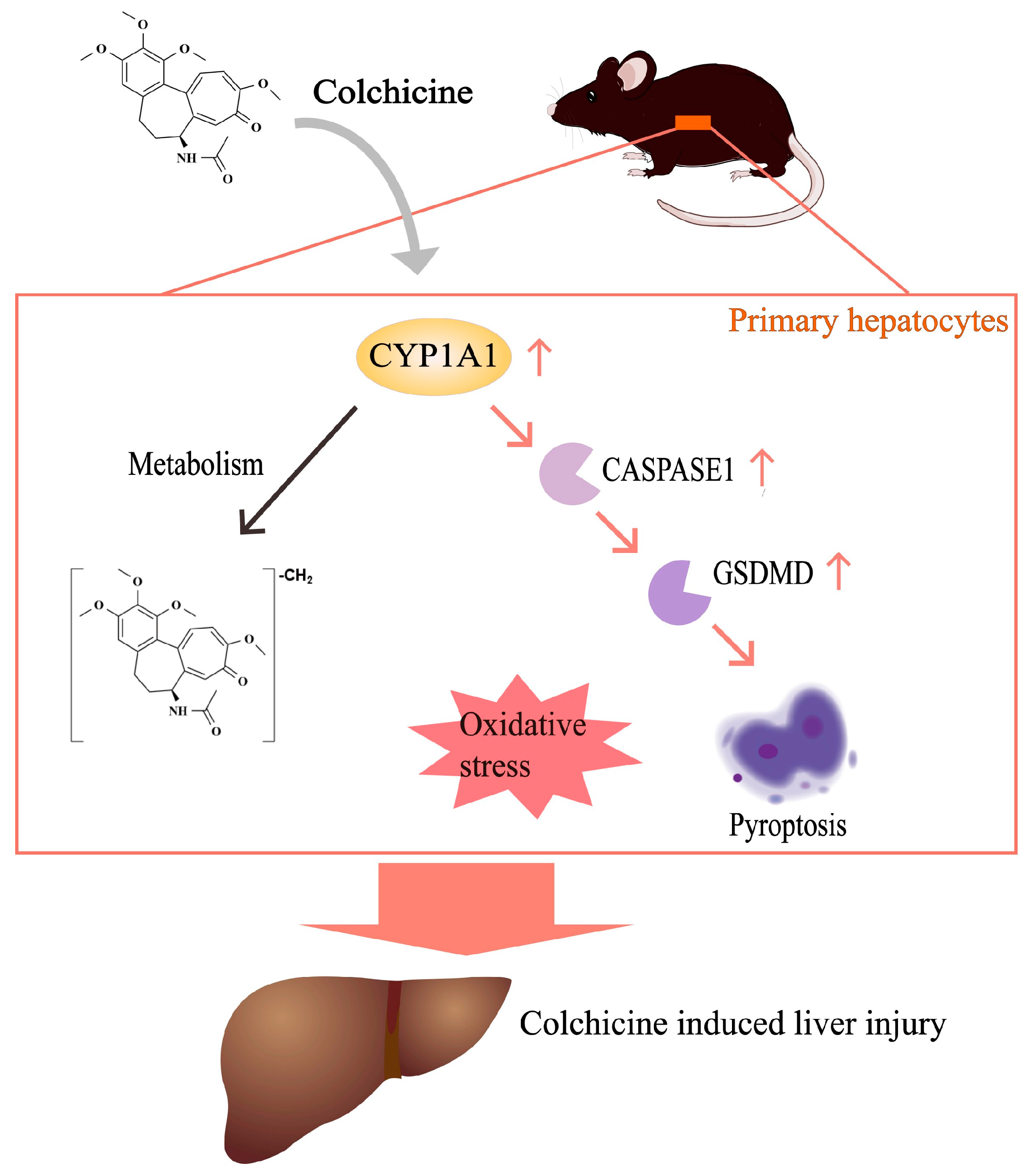
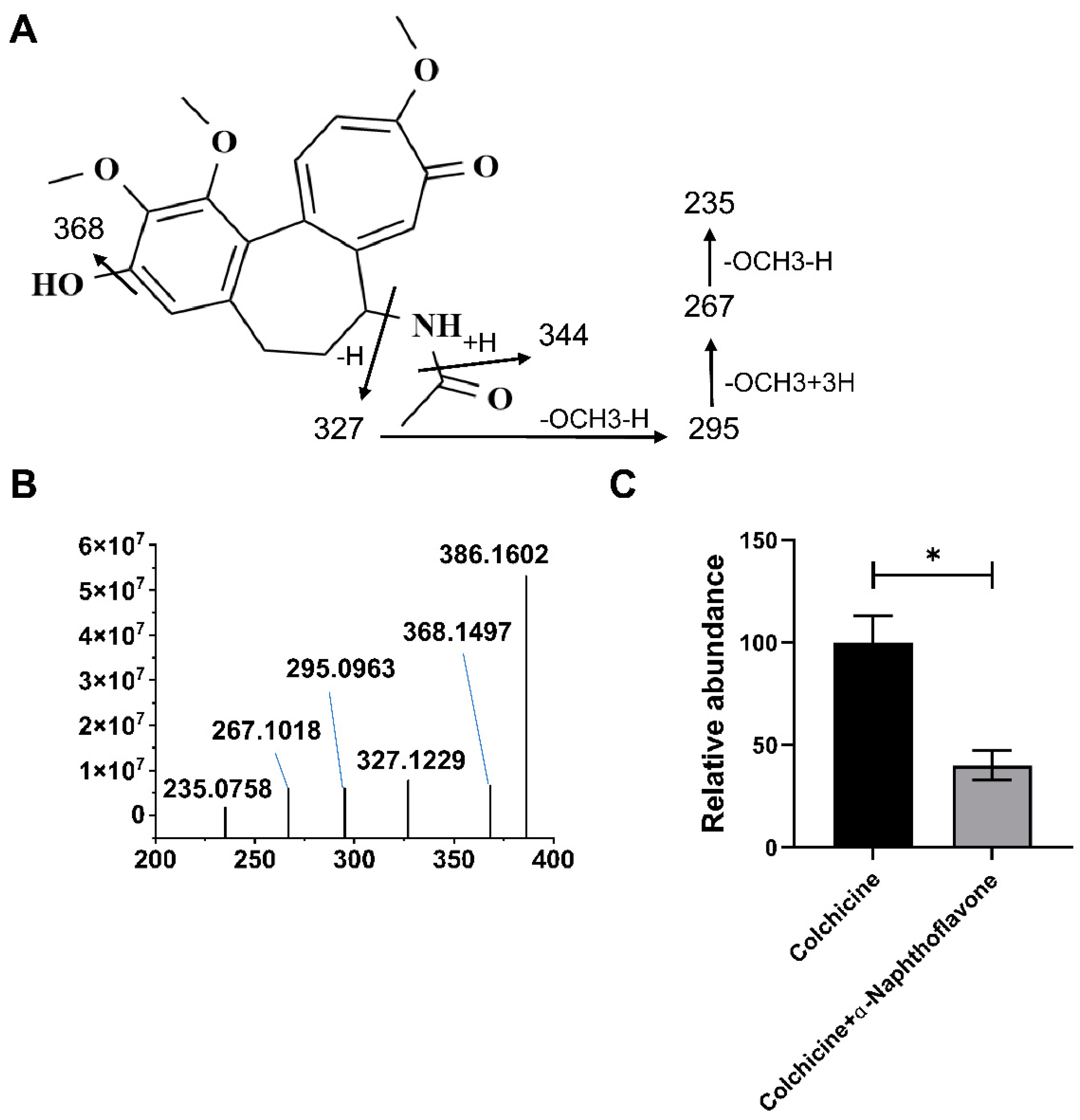
2.2. CYP1A1 Metabolizes Colchicine
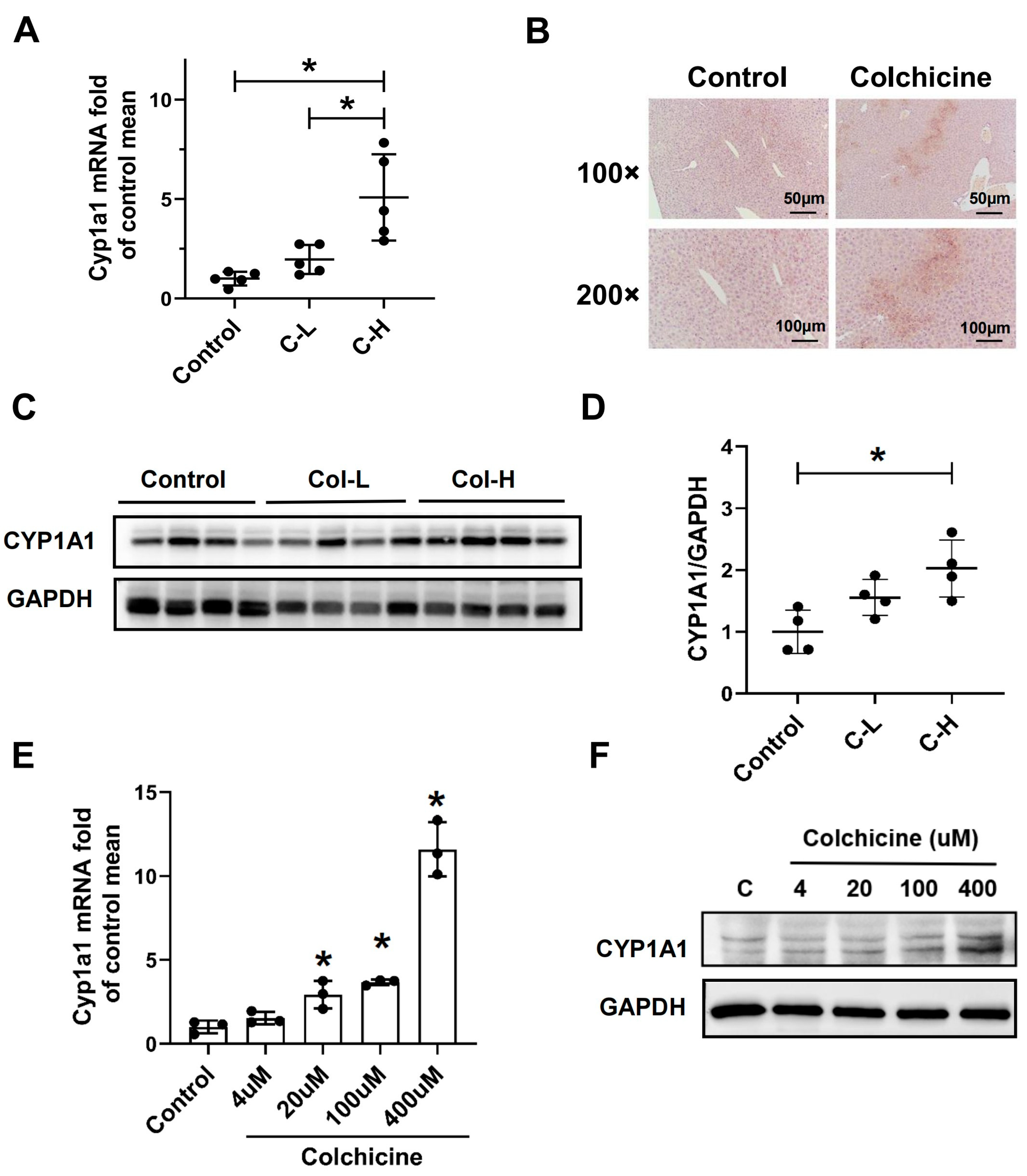
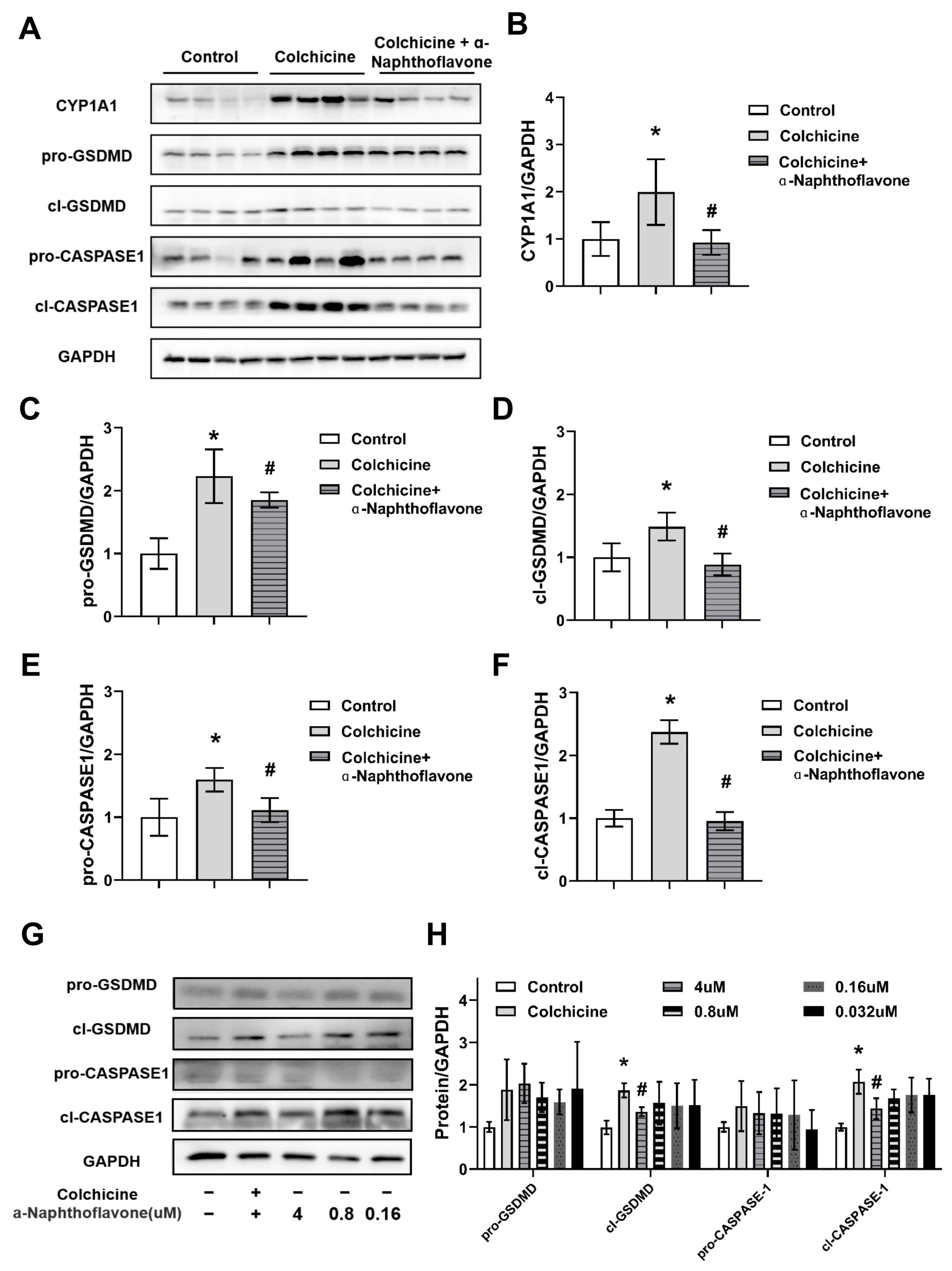
2.3. CYP1A1 Was Significantly Elevated by Colchicine
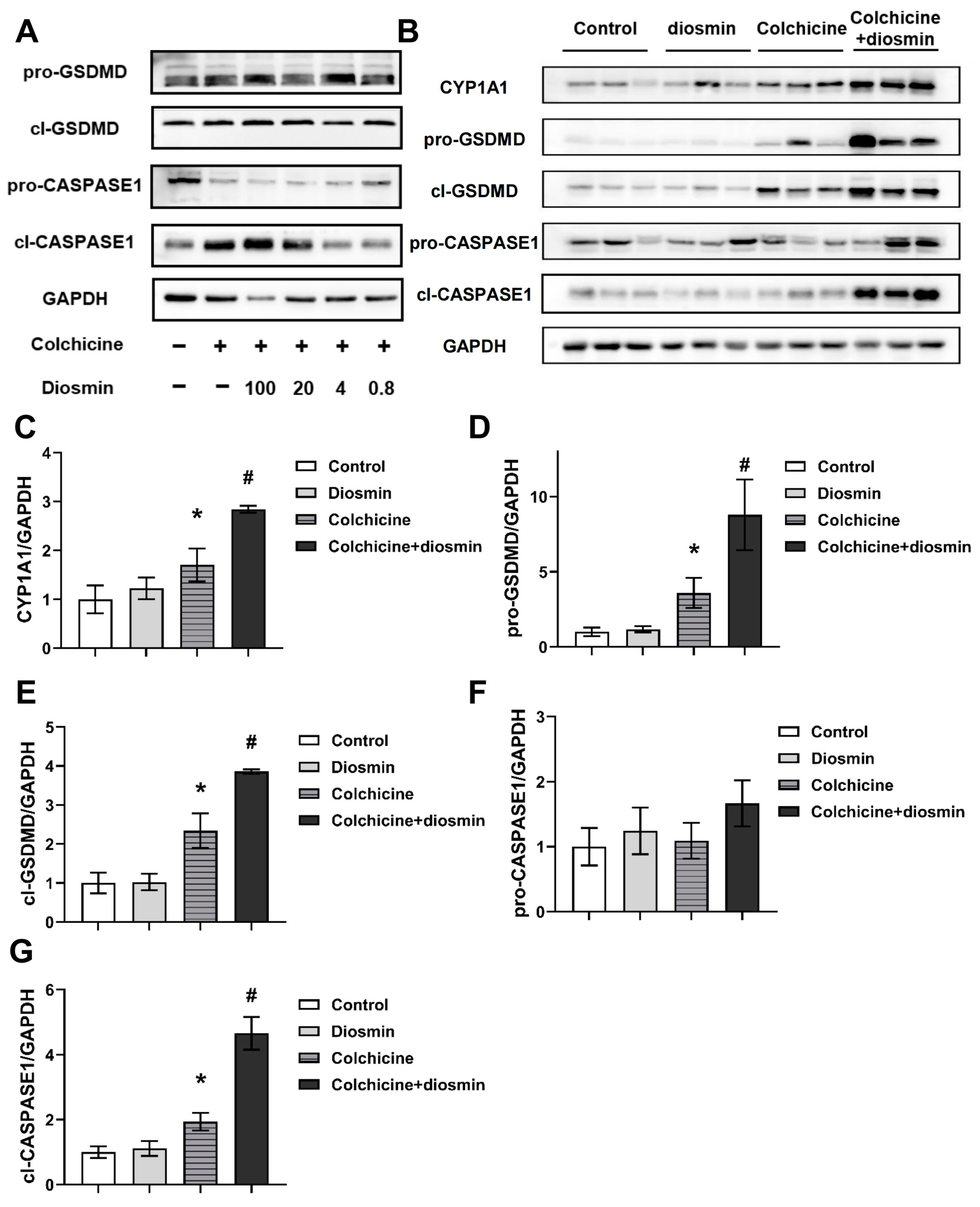
2.4. Regulation of CYP1A1 Could Regulate Colchicine-Induced Liver Injury
2.5. Inhibition of CASPASE-1-Mediated Pyroptosis Pathway by Inhibiting CYP1A1
2.6. Aggravation of CASPASE-1-Mediated Pyroptosis Pathway by Activating CYP1A1
3. Discussion
4. Conclusions
5. Materials and Methods
5.1. Chemicals and Reagents
5.2. Animals
5.3. Animal Treatment
5.4. In Vitro Metabolism of Colchicine
5.5. UHPLC-Q Exactive Plus MS Analysis
5.6. QPCR, WB, Histological, Immunohistochemistry and Biochemical Assessment
5.7. Preparation of Primary Hepatocytes and Cell Viability Assay
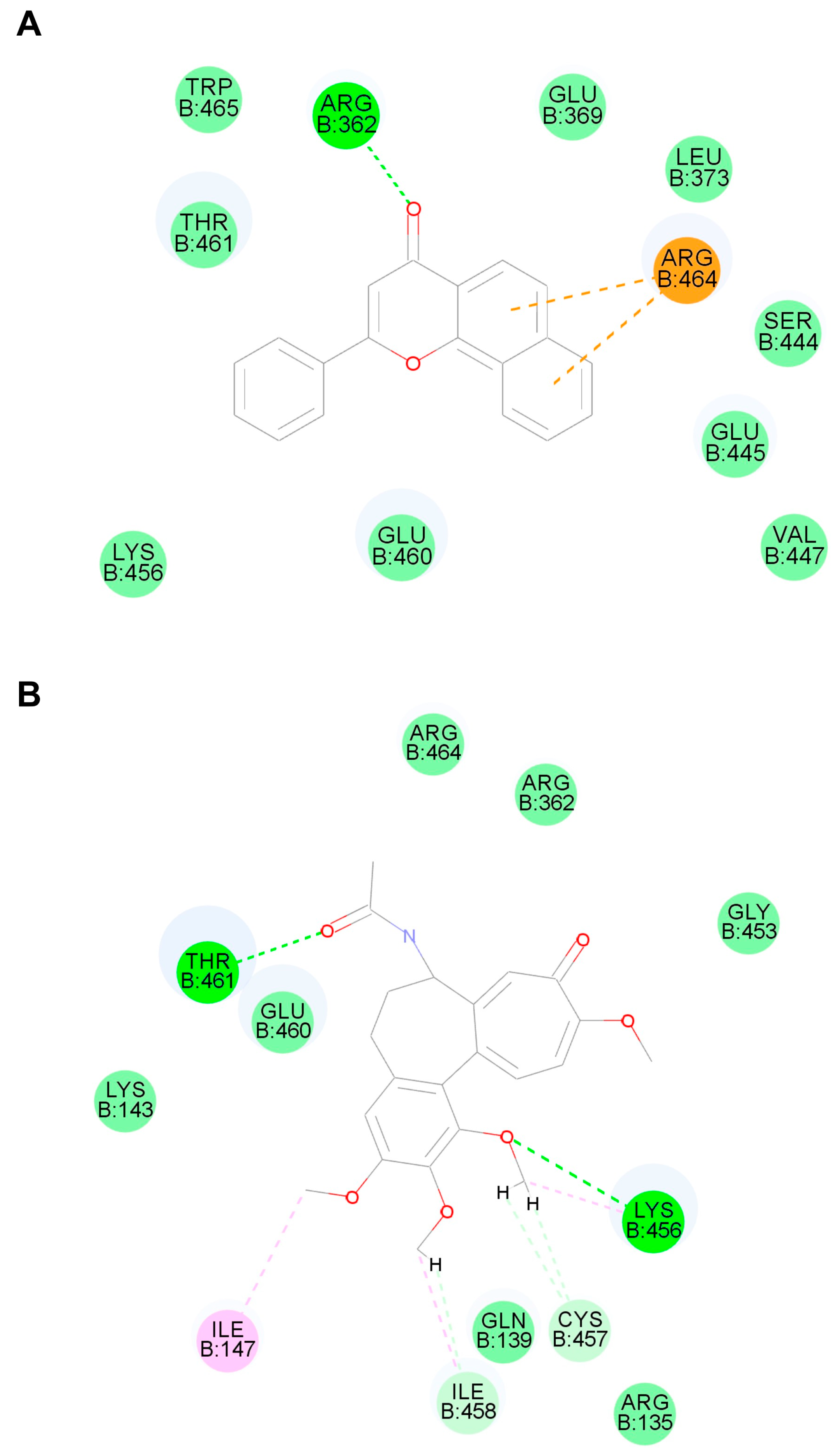
5.8. Molecular Docking
5.9. Data and Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
References
- Dasgeb, B.; Kornreich, D.; McGuinn, K.; Okon, L.; Brownell, I.; Sackett, D.L. Colchicine: An ancient drug with novel applications. Br. J. Dermatol. 2018, 178, 350–356. [Google Scholar] [CrossRef] [PubMed]
- Reyes, A.Z.; Hu, K.A.; Teperman, J.; Muskardin, T.L.W.; Tardif, J.C.; Shah, B.; Pillinger, M.H. Anti-inflammatory therapy for COVID-19 infection: The case for colchicine. Ann. Rheum. Dis. 2020, 80, 550–557. [Google Scholar] [CrossRef] [PubMed]
- Deftereos, S.G.; Beerkens, F.J.; Shah, B.; Giannopoulos, G.; Vrachatis, D.A.; Giotaki, S.G.; Siasos, G.; Nicolas, J.; Arnott, C.; Patel, S.; et al. Colchicine in Cardiovascular Disease: In-Depth Review. Circulation 2022, 145, 61–78. [Google Scholar] [PubMed]
- Bouabdallaoui, N.; Tardif, J.C.; Waters, D.D.; Pinto, F.J.; Maggioni, A.P.; Diaz, R.; Berry, C.; Koenig, W.; Sendon, J.L.; Gamra, H.; et al. Time-to-treatment initiation of colchicine and cardiovascular outcomes after myocardial infarction in the Colchicine Cardiovascular Outcomes Trial (COLCOT). Eur. Heart J. 2020, 41, 4092–4099. [Google Scholar] [CrossRef] [PubMed]
- Fu, M.; Zhao, J.; Li, Z.; Zhao, H.; Lu, A. Clinical outcomes after colchicine overdose: A case report. Medicine 2019, 98, e16580. [Google Scholar] [CrossRef] [PubMed]
- Jerschke, E.; Barkovsky, M.; Jung, N.; Neuberger, H.; Stenzel, J.; Eyer, F.; Skerra, A.; Geith, S. In vivo neutralization of colchicine toxicity by a PASylated anticalin in a rat model. Toxicology 2023, 492, 153526. [Google Scholar] [CrossRef] [PubMed]
- Finkelstein, Y.; Aks, S.E.; Hutson, J.R.; Juurlink, D.N.; Nguyen, P.; Dubnov-Raz, G.; Pollak, U.; Koren, G.; Bentur, Y. Colchicine poisoning: The dark side of an ancient drug. Clin. Toxicol. 2010, 48, 407–414. [Google Scholar] [CrossRef]
- Mescher, M.; Haarmann-Stemmann, T. Modulation of CYP1A1 metabolism: From adverse health effects to chemoprevention and therapeutic options. Pharmacol. Ther. 2018, 187, 71–87. [Google Scholar] [CrossRef]
- Xia, H.; Zhu, X.; Zhang, X.; Jiang, H.; Li, B.; Wang, Z.; Li, D.; Jin, Y. Alpha-naphthoflavone attenuates non-alcoholic fatty liver disease in oleic acid-treated HepG2 hepatocytes and in high fat diet-fed mice. Biomed. Pharmacother. 2019, 118, 109287. [Google Scholar] [CrossRef]
- Vogel, C.F.A.; Winkle, L.S.V.; Esser, C.; Haarmann-Stemmann, T. The aryl hydrocarbon receptor as a target of environmental stressors—Implications for pollution mediated stress and inflammatory responses. Redox Biol. 2020, 34, 101530. [Google Scholar] [CrossRef]
- Chinn, L.W.; Kroetz, D.L. ABCB1 pharmacogenetics: Progress, pitfalls, and promise. Clin. Pharmacol. Ther. 2007, 81, 265–269. [Google Scholar] [CrossRef] [PubMed]
- Davis, M.W.; Wason, S. Effect of steady-state atorvastatin on the pharmacokinetics of a single dose of colchicine in healthy adults under fasted conditions. Clin. Drug Investig. 2014, 34, 259–267. [Google Scholar] [CrossRef] [PubMed]
- Imazio, M.; Nidorf, M. Colchicine and the heart. Eur. Heart J. 2021, 42, 2745–2760. [Google Scholar] [CrossRef] [PubMed]
- Guo, L.; Liu, J.; Hu, Y.; Wang, D.; Li, Z.; Zhang, J.; Qin, T.; Liu, X.; Liu, C.; Zhao, X.; et al. Astragalus polysaccharide and sulfated epimedium polysaccharide synergistically resist the immunosuppression. Carbohydr. Polym. 2012, 90, 1055–1060. [Google Scholar] [CrossRef] [PubMed]
- Lv, W.J.; Ma, Y.M.; Huang, J.Y.; He, S.Q.; Li, S.P.; Lin, J.; Chen, R.; Lun, J.C.; Liu, J.; Guo, S.N. Polysaccharides derived from Shenling Baizhu San improve colitis via modulating tryptophan metabolism in mice. Int. J. Biol. Macromol. 2022, 222, 1127–1136. [Google Scholar] [CrossRef]
- Jin, J.; Yuan, P.; Yu, W.; Lin, J.; Xu, A.; Xu, X.; Lou, J.; Yu, T.; Qian, C.; Liu, B.; et al. Mitochondria-Targeting Polymer Micelle of Dichloroacetate Induced Pyroptosis to Enhance Osteosarcoma Immunotherapy. ACS Nano 2022, 16, 10327–10340. [Google Scholar] [CrossRef]
- Zhao, Q.; Dai, M.Y.; Huang, R.Y.; Duan, J.Y.; Zhang, T.; Bao, W.M.; Zhang, J.Y.; Gui, S.Q.; Xia, S.M.; Dai, C.T.; et al. Parabacteroides distasonis ameliorates hepatic fibrosis potentially via modulating intestinal bile acid metabolism and hepatocyte pyroptosis in male mice. Nat. Commun. 2023, 14, 1829. [Google Scholar] [CrossRef]
- Xu, W.; Che, Y.; Zhang, Q.; Huang, H.; Ding, C.; Wang, Y.; Wang, G.; Cao, L.; Hao, H. Apaf-1 Pyroptosome Senses Mitochondrial Permeability Transition. Cell Metab. 2020, 33, 424–436. [Google Scholar] [CrossRef]
- McKenzie, B.J.; Wechalekar, M.D.; Johnston, R.V.; Schlesinger, N.; Buchbinder, R. Colchicine for acute gout. Cochrane Database Syst. Rev. 2021, 8, CD006190. [Google Scholar] [CrossRef]
- Zhongying, L.; Jianping, Y.; Shixue, C.; Yuanyong, Y. Effect of Dihydromyricetin from Cany Tea on Acute Gouty Arthritis Model Rats. Chin. J. Mod. Appl. Pharm. 2015, 32, 396–399. [Google Scholar] [CrossRef]
- Fu, D.J.; Liu, S.M.; Yang, J.J.; Li, J. Novel piperidine derivatives as colchicine binding site inhibitors induce apoptosis and inhibit epithelial-mesenchymal transition against prostate cancer PC3 cells. J. Enzym. Inhib. Med. Chem. 2020, 35, 1403–1413. [Google Scholar] [CrossRef]
- Denk, H.; Eckerstorfer, R. Colchicine-induced Mallory body formation in the mouse. Lab Investig. 1997, 36, 563–565. [Google Scholar]
- Liu, Y.; Su, B.; Pei, R.; Yeh, C.; Yeh, K.; Lee, K.Y.; Hsu, Y.; Ho, C.; Lai, Y. The stability of cytokeratin 18 in human liver cells during colchicine-induced microtubule disruption. Food Chem. Toxicol. 2001, 39, 85–89. [Google Scholar] [CrossRef] [PubMed]
- Kumar, A.; Seghal, N.; Padi, S.V.; Naidu, P.S. Differential effects of cyclooxygenase inhibitors on intracerebroventricular colchicine-induced dysfunction and oxidative stress in rats. Eur. J. Pharmacol. 2006, 551, 58–66. [Google Scholar] [CrossRef] [PubMed]
- Zhang, D.; Li, L.; Li, J.; Wei, Y.; Tang, J.; Man, X.; Liu, F. Colchicine improves severe acute pancreatitis-induced acute lung injury by suppressing inflammation, apoptosis and oxidative stress in rats. Biomed. Pharmacother. 2022, 153, 113461. [Google Scholar] [CrossRef] [PubMed]
- Zhang, B.; Huang, R.; Yang, D.; Chen, G.; Chen, Y.; Han, J.; Zhang, S.; Ma, L.; Yang, X. Combination of Colchicine and Ticagrelor Inhibits Carrageenan-Induced Thrombi in Mice. Oxid. Med. Cell. Longev. 2022, 2022, 3087198. [Google Scholar] [CrossRef] [PubMed]
- Robertson, S.; Martínez, G.J.; Payet, C.A.; Barraclough, J.Y.; Celermajer, D.S.; Bursill, C.; Patel, S. Colchicine therapy in acute coronary syndrome patients acts on caspase-1 to suppress NLRP3 inflammasome monocyte activation. Clin. Sci. 2016, 130, 1237–1246. [Google Scholar] [CrossRef]
- Guo, X.; Chen, Y.; Li, Q.; Yang, X.; Zhao, G.; Peng, Y.; Zheng, J. Studies on hepatotoxicity and toxicokinetics of colchicine. J. Biochem. Mol. Toxicol. 2019, 33, e22366. [Google Scholar] [CrossRef]
- Szychowski, K.A.; Wnuk, A.; Kajta, M.; Wójtowicz, A.K. Triclosan activates aryl hydrocarbon receptor (AhR)-dependent apoptosis and affects Cyp1a1 and Cyp1b1 expression in mouse neocortical neurons. Environ. Res. 2016, 151, 106–114. [Google Scholar] [CrossRef]
- Long, Y.; Liu, X.; Tan, X.Z.; Jiang, C.X.; Chen, S.W.; Liang, G.N.; He, X.M.; Wu, J.; Chen, T.; Xu, Y. ROS-induced NLRP3 inflammasome priming and activation mediate PCB 118- induced pyroptosis in endothelial cells. Ecotoxicol. Environ. Saf. 2020, 189, 109937. [Google Scholar] [CrossRef]
- Wang, Q.; Wang, D.; Zuo, Z.; Ye, B.; Dong, Z.; Zou, J. Effects of Dietary Koumine on Growth Performance, Intestinal Morphology, Microbiota, and Intestinal Transcriptional Responses of Cyprinus carpio. Int. J. Mol. Sci. 2022, 23, 11860. [Google Scholar] [CrossRef] [PubMed]
- Li, H.; Gao, Y.; Song, L.; Chen, T.; Zhang, G.; Ye, Z.; Gao, Y.; Huo, W. Ginsenoside Rg1 protects mice against 2,3,7,8-tetrachlorodibenzo-p-dioxin-induced liver injury by inhibiting CYP1A1 through the aryl hydrocarbon receptor. J. Ethnopharmacol. 2022, 294, 115394. [Google Scholar] [CrossRef] [PubMed]
- Dvorák, Z.; Vrzal, R.; Ulrichová, J.; Pascussi, J.M.; Maurel, P.; Modriansky, M. Involvement of cytoskeleton in AhR-dependent CYP1A1 expression. Curr. Drug Metab. 2006, 7, 301–313. [Google Scholar] [CrossRef] [PubMed]
- Magnotti, F.; Lefeuvre, L.; Benezech, S.; Malsot, T.; Waeckel, L.; Martin, A.; Kerever, S.; Chirita, D.; Desjonqueres, M.; Duquesne, A.; et al. Pyrin dephosphorylation is sufficient to trigger inflammasome activation in familial Mediterranean fever patients. EMBO Mol. Med. 2019, 11, e10547. [Google Scholar] [CrossRef] [PubMed]
- Yang, M.; Lv, H.; Liu, Q.; Zhang, L.; Zhang, R.; Huang, X.; Wang, X.; Han, B.; Hou, S.; Liu, D.; et al. Colchicine Alleviates Cholesterol Crystal-Induced Endothelial Cell Pyroptosis through Activating AMPK/SIRT1 Pathway. Oxid. Med. Cell. Longev. 2020, 2020, 9173530. [Google Scholar] [CrossRef] [PubMed]
- Villa Zapata, L.; Hansten, P.D.; Horn, J.R.; Boyce, R.D.; Gephart, S.; Subbian, V.; Romero, A.; Malone, D.C. Evidence of Clinically Meaningful Drug–Drug Interaction with Concomitant Use of Colchicine and Clarithromycin. Drug Saf. 2020, 43, 661–668. [Google Scholar] [CrossRef] [PubMed]
- Haj Yahia, S.; Ben Zvi, I.; Livneh, A.A.O. Colchicine intoxication in familial Mediterranean fever patients using clarithromycin for the treatment of Helicobacter pylori: A series of six patients. Rheumatol. Int. 2018, 38, 141–147. [Google Scholar] [CrossRef] [PubMed]
- Huang, R.; Wang, C.; Wu, Z.E.; Zhao, Q.; Duan, J.; Huang, W.; Cheng, Y.; Zhu, B.; Li, F. Metabolomics reveals that sulfotransferase 1 may regulate colchicine-induced liver injury. Chem.-Biol. Interact. 2023, 386, 110776. [Google Scholar] [CrossRef]
- Hassan, R.; Myllys, M.; Brackhagen, L.; Hobloss, Z.; González, D.; Seddek, A.L.; Friebel, A.; Hoehme, S.; Marchan, R.; Trauner, M.; et al. Colchicine overdose impairs the capacity of Kupffer cells to clear foreign particles and endotoxins. Arch. Toxicol. 2022, 96, 3067–3076. [Google Scholar] [CrossRef]
- Trebach, J.; Boyd, M.; Crane, A.; DiSalvo, P.; Biary, R.; Hoffman, R.S.; Su, M.K. Confirmed Fatal Colchicine Poisoning in an Adolescent with Blood and Bile Concentrations-Implications for GI Decontamination? J. Med. Toxicol. 2023, 19, 280–283. [Google Scholar] [CrossRef]
- Liang, S.C.; Xia, Y.L.; Hou, J.; Ge, G.B.; Zhang, J.W.; He, Y.Q.; Wang, J.Y.; Qi, X.Y.; Yang, L. Methylation, Glucuronidation, and Sulfonation of Daphnetin in Human Hepatic Preparations In Vitro: Metabolic Profiling, Pathway Comparison, and Bioactivity Analysis. J. Pharm. Sci. 2016, 105, 808–816. [Google Scholar] [CrossRef] [PubMed]
- Hu, H.; Gehart, H.; Artegiani, B.; LÖpez-Iglesias, C.; Dekkers, F.; Basak, O.; Es, J.v.; Lopes, S.M.C.d.S.; Begthel, H.; Korving, J.; et al. Long-Term Expansion of Functional Mouse and Human Hepatocytes as 3D Organoids. Cell 2018, 175, 1591–1606. [Google Scholar] [CrossRef] [PubMed]
- Dowden, R.A.; McGuinness, L.R.; Wisniewski, P.J.; Campbell, S.C.; Guers, J.J.; Oydanich, M.; Vatner, S.F.; Häggblom, M.M.; Kerkhof, L.J. Host genotype and exercise exhibit species-level selection for members of the gut bacterial communities in the mouse digestive system. Sci. Rep. 2020, 10, 8984. [Google Scholar] [CrossRef] [PubMed]
- Tan, W.C.C.; Nerurkar, S.N.; Cai, H.Y.; Ng, H.H.M.; Wu, D.; Wee, Y.T.F.; Lim, J.C.T.; Yeong, J.; Lim, T.K.H. Overview of multiplex immunohistochemistry/immunofluorescence techniques in the era of cancer immunotherapy. Cancer Commun. 2020, 40, 135–153. [Google Scholar] [CrossRef]
- Jabir, N.R.; Rehman, M.T.; Alsolami, K.; Shakil, S.; Zughaibi, T.A.; Alserihi, R.F.; Khan, M.S.; AlAjmi, M.F.; Tabrez, S. Concatenation of molecular docking and molecular simulation of BACE-1, γ-secretase targeted ligands: In pursuit of Alzheimer’s treatment. Ann. Med. 2021, 53, 2332–2344. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| M1 | M2 | M3 | M4 | M5 | M6 | M7 | M8 | |
|---|---|---|---|---|---|---|---|---|
| Control | 0.00 ± 0.00 | 0.00 ± 0.00 | 0.00 ± 0.00 | 0.00 ± 0.00 | 0.00 ± 0.00 | 0.00 ± 0.00 | 0.00 ± 0.00 | 0.00 ± 0.00 |
| CYP1A1 | 3.91 ± 3.16 | 4.51 ± 2.24 | 4.92 ± 1.48 | 6.99 ± 0.30 | 6.87 ± 0.00 | 8.78 ± 1.10 | 8.53 ± 0.61 | 9.32 ± 5.12 |
| CYP1A2 | 6.36 ± 0.19 | 0.00 ± 0.00 | 7.83 ± 0.27 | 6.72 ± 0.44 | 8.33 ± 0.00 | 8.69 ± 0.17 | 5.42 ± 0.50 | 8.67 ± 3.80 |
| CYP1B1 | 4.56 ± 1.73 | 2.53 ± 0.65 | 8.08 ± 0.40 | 7.00 ± 0.10 | 7.27 ± 2.20 | 7.14 ± 0.77 | 4.80 ± 0.73 | 7.48 ± 2.17 |
| CYP2A6 | 5.39 ± 0.90 | 2.64 ± 0.16 | 8.53 ± 1.02 | 6.66 ± 2.12 | 7.26 ± 8.64 | 8.25 ± 0.69 | 4.07 ± 0.65 | 4.23 ± 0.62 |
| CYP2B6 | 6.90 ± 0.52 | 4.42 ± 0.21 | 8.37 ± 0.52 | 7.77 ± 0.57 | 7.86 ± 0.00 | 9.86 ± 0.58 | 12.35 ± 0.43 | 10.13 ± 3.38 |
| CYP2C19 | 8.35 ± 0.65 | 6.27 ± 0.30 | 7.68 ± 0.04 | 6.69 ± 1.91 | 8.63 ± 1.05 | 6.74 ± 0.36 | 4.10 ± 0.63 | 7.66 ± 2.52 |
| CYP2C8 | 6.82 ± 0.03 | 3.69 ± 0.02 | 7.94 ± 0.13 | 7.21 ± 0.40 | 7.37 ± 0.00 | 6.90 ± 1.01 | 10.77 ± 0.42 | 8.61 ± 4.55 |
| CYP2C9 | 13.68 ± 5.29 | 5.57 ± 1.45 | 6.83 ± 0.42 | 6.44 ± 0.02 | 7.59 ± 0.00 | 7.33 ± 0.35 | 9.27 ± 0.35 | 6.12 ± 3.22 |
| CYP2D6 | 7.91 ± 0.60 | 3.23 ± 0.56 | 7.20 ± 0.32 | 11.27 ± 3.15 | 7.65 ± 11.17 | 7.59 ± 0.36 | 0.00 ± 0.15 | 0.00 ± 0.00 |
| CYP2E1 | 4.77 ± 2.02 | 3.69 ± 0.51 | 8.11 ± 0.05 | 6.93 ± 0.40 | 7.32 ± 0.00 | 8.09 ± 0.37 | 7.85 ± 0.54 | 6.78 ± 0.72 |
| CYP3A4 | 14.93 ± 1.41 | 47.73 ± 5.45 | 7.46 ± 0.07 | 12.18 ± 0.70 | 7.72 ± 15.62 | 6.55 ± 0.65 | 14.09 ± 0.08 | 12.27 ± 1.54 |
| CYP3A5 | 9.93 ± 0.49 | 12.14 ± 0.15 | 8.56 ± 0.20 | 7.36 ± 0.44 | 8.12 ± 0.00 | 8.06 ± 0.09 | 14.87 ± 0.31 | 11.08 ± 1.09 |
| CYP4A11 | 6.48 ± 0.33 | 3.57 ± 0.39 | 8.49 ± 0.07 | 6.78 ± 0.33 | 8.01 ± 0.00 | 6.01 ± 0.22 | 3.87 ± 0.04 | 7.65 ± 1.95 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Huang, R.; Duan, J.; Huang, W.; Cheng, Y.; Zhu, B.; Li, F. Inhibition of CYP1A1 Alleviates Colchicine-Induced Hepatotoxicity. Toxins 2024, 16, 35. https://doi.org/10.3390/toxins16010035
Huang R, Duan J, Huang W, Cheng Y, Zhu B, Li F. Inhibition of CYP1A1 Alleviates Colchicine-Induced Hepatotoxicity. Toxins. 2024; 16(1):35. https://doi.org/10.3390/toxins16010035
Chicago/Turabian StyleHuang, Ruoyue, Jingyi Duan, Wen Huang, Yan Cheng, Beiwei Zhu, and Fei Li. 2024. "Inhibition of CYP1A1 Alleviates Colchicine-Induced Hepatotoxicity" Toxins 16, no. 1: 35. https://doi.org/10.3390/toxins16010035
APA StyleHuang, R., Duan, J., Huang, W., Cheng, Y., Zhu, B., & Li, F. (2024). Inhibition of CYP1A1 Alleviates Colchicine-Induced Hepatotoxicity. Toxins, 16(1), 35. https://doi.org/10.3390/toxins16010035