The Effect of Mycotoxins and Their Mixtures on Bovine Spermatozoa Characteristics


Abstract
:1. Introduction
2. Results
2.1. Solvent Toxicity Examination
2.2. Dose-Response Study—The Cytotoxicity of Single Mycotoxins
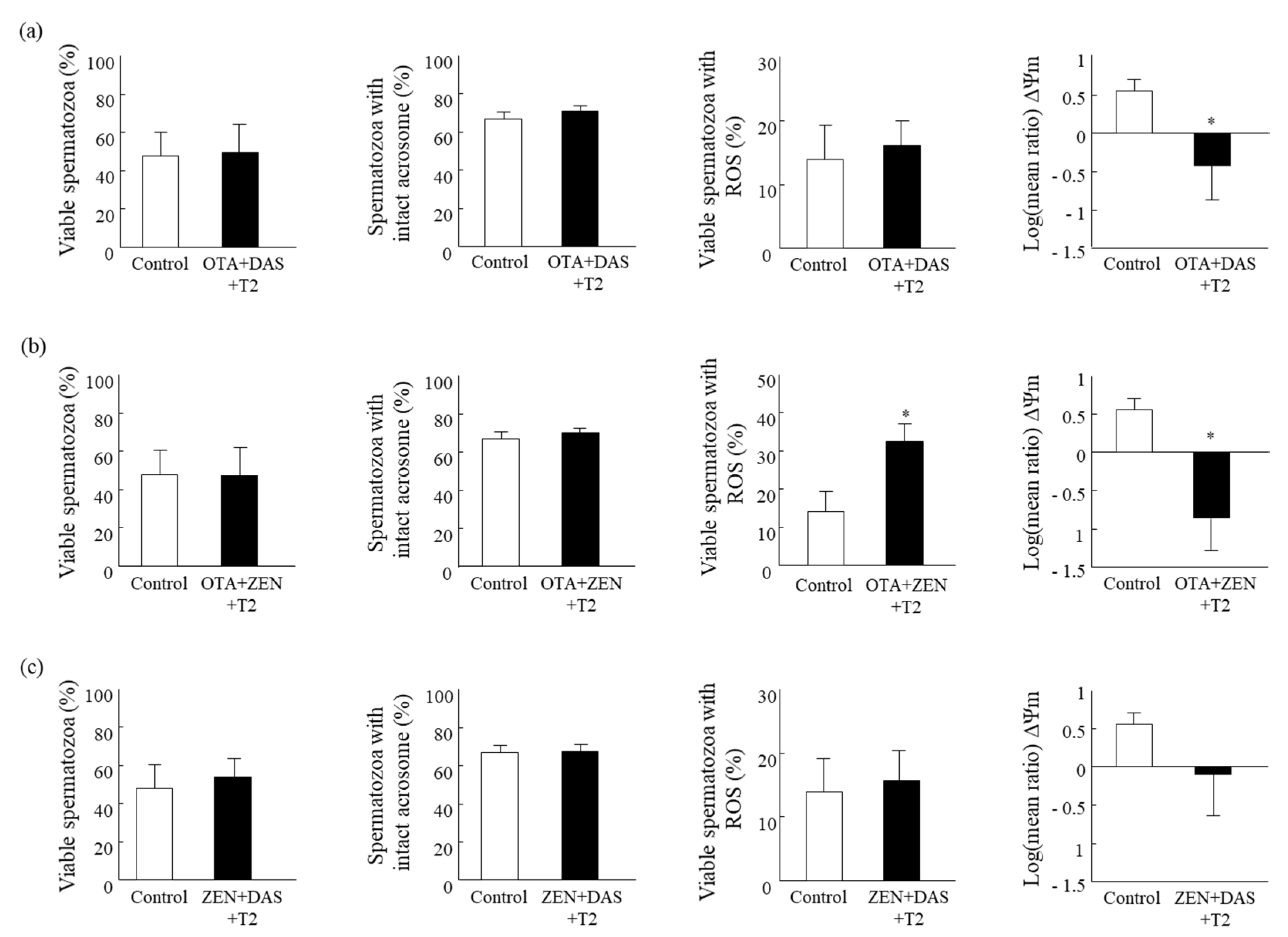
2.3. Effect of Mixture-II—Cytotoxicity of the Binary Mixtures of Mycotoxins
Estimated Interactions between Two Mycotoxins
2.4. Effect of Mixture-III—The Cytotoxicity of Ternary Mixtures of Mycotoxins
Estimated Interactions between Three Mycotoxins
3. Discussion
3.1. Evaluation of the Spermatozoa Quality
3.2. Cytotoxicity of Single Mycotoxins
3.3. Cytotoxicity of Binary Mycotoxin Mixtures
3.4. Cytotoxicity of Ternary Mycotoxin Mixtures
4. Conclusions
5. Materials and Methods
5.1. Mycotoxin Concentration
5.2. Sample Preparation
Mycotoxin Exposure
5.3. Cell-Based Assessment of Spermatozoa
5.3.1. Plasma Membrane Integrity
5.3.2. Acrosome Membrane Integrity
5.3.3. Mitochondrial Features
Mitochondrial Membrane Potential
Oxidation Status
5.4. Estimation of the Interactive Effects of Combined Mycotoxins
- (1)
- For the binary mixtures:
- a.
- Mean expected adjusted to control (mycotoxin 1+ mycotoxin 2) = mean adjusted to control (mycotoxin 1) + mean adjusted to control (mycotoxin 2)
- b.
- SEM expected (mycotoxin 1 + mycotoxin 2) = [(SEM for mycotoxin 1)2 + (SEM for mycotoxin 2)2]1/2
- (2)
- For the ternary mixtures:
- a.
- Mean expected adjusted to control (mycotoxin 1+ mycotoxin 2+ mycotoxin 3) = mean adjusted to control (mycotoxin 1) + mean adjusted to control (mycotoxin 2) + mean adjusted to control (mycotoxin 3)
- b.
- SEM (expected for mycotoxin 1 + mycotoxin 2 + mycotoxin 3) = [(SEM for mycotoxin 1)2 + (SEM for mycotoxin 2)2 + (SEM for mycotoxin 3)2]1/2
5.5. Statistical Analysis
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Ismaiel, A.A.; Papenbrock, J. Mycotoxins: Producing Fungi and Mechanisms of Phytotoxicity. Agriculture 2015, 5, 492–537. [Google Scholar] [CrossRef]
- Bennett, J.W. Mycotoxins, Mycotoxicoses, Mycotoxicology and Mycopathologia. Mycopathologia 1987, 100, 3–5. [Google Scholar] [CrossRef]
- Gallo, A.; Giuberti, G.; Frisvad, J.C.; Bertuzzi, T.; Nielsen, K.F. Review on Mycotoxin Issues in Ruminants: Occurrence in Forages, Effects of Mycotoxin Ingestion on Health Status and Animal Performance and Practical Strategies to Counteract Their Negative Effects. Toxins 2015, 7, 3057–3111. [Google Scholar] [CrossRef]
- Pleadin, J.; Frece, J.; Markov, K. Mycotoxins in Food and Feed. Adv. Food Nutr. Res. 2019, 89, 297–345. [Google Scholar] [CrossRef]
- Coffey, R.; Cummins, E.; Ward, S. Exposure Assessment of Mycotoxins in Dairy Milk. Food Control 2009, 20, 239–249. [Google Scholar] [CrossRef]
- Eskola, M.; Kos, G.; Elliott, C.T.; Hajšlová, J.; Mayar, S.; Krska, R. Worldwide Contamination of Food-Crops with Mycotoxins: Validity of the Widely Cited “FAO Estimate” of 25. Crit. Rev. Food Sci. Nutr. 2020, 60, 2773–2789. [Google Scholar] [CrossRef]
- Bennett, J.W.; Klich, M. Mycotoxins. Clin. Microbiol. Rev. 2003, 16, 497–516. [Google Scholar] [CrossRef]
- Turner, N.W.; Subrahmanyam, S.; Piletsky, S.A. Analytical Methods for Determination of Mycotoxins: A Review. Anal. Chim. Acta 2009, 632, 168–180. [Google Scholar] [CrossRef] [PubMed]
- Van Egmond, H.P.; Jonker, M.A. Current Situation on Regulations for Mycotoxins. Mycotoxins 2003, 2003, 1–15. [Google Scholar] [CrossRef] [PubMed]
- Knutsen, H.K.; Alexander, J.; Barregård, L.; Bignami, M.; Brüschweiler, B.; Ceccatelli, S.; Cottrill, B.; Dinovi, M.; Grasl-Kraupp, B.; Hogstrand, C.; et al. Risk to Human and Animal Health Related to the Presence of 4,15-Diacetoxyscirpenol in Food and Feed. EFSA J. 2018, 16, e05367. [Google Scholar] [CrossRef]
- Commision Recommendation (EU) 2016/1319 Amending Recommendation 2006/576/EC as Regards Deoxynivalenol, Zearalenone and Ochratoxin A in Pet Food. Off. J. Eur. Union L208/58 2016, 73, 58–60.
- Eze, U.A.; Huntriss, J.; Routledge, M.N.; Gong, Y.Y. Toxicological Effects of Regulated Mycotoxins and Persistent Organochloride Pesticides: In Vitro Cytotoxic Assessment of Single and Defined Mixtures on MA-10 Murine Leydig Cell Line. Toxicol. Vitr. 2018, 48, 93–103. [Google Scholar] [CrossRef] [PubMed]
- Eze, U.A.; Lewis, S.E.M.; Connolly, L.; Gong, Y.Y. Mycotoxins as Potential Cause of Human Infertility—A Review of Evidence from Animal and Cellular Models. Acta Hortic. 2018, 1225, 513–525. [Google Scholar] [CrossRef]
- Driehuis, F.; Spanjer, M.C.; Scholten, J.M.; Te Giffel, M.C. Occurrence of Mycotoxins in Feedstuffs of Dairy Cows and Estimation of Total Dietary Intakes. J. Dairy Sci. 2008, 91, 4261–4271. [Google Scholar] [CrossRef] [PubMed]
- Cinar, A.; Onbaşı, E. Mycotoxins: The Hidden Danger in Foods. In Mycotoxins and Food Safety; IntechOpen: London, UK, 2020. [Google Scholar] [CrossRef]
- Li, L.; Zhang, T.; Ren, X.; Li, B.; Wang, S. Male Reproductive Toxicity of Zearalenone-Meta-Analysis with Mechanism Review. Ecotoxicol. Environ. Saf. 2021, 221, 112457. [Google Scholar] [CrossRef]
- Deng, H.; Chen, W.; Zhang, B.; Zhang, Y.; Han, L.; Zhang, Q.; Yao, S.; Wang, H.; Shen, X.L. Excessive ER-Phagy Contributes to Ochratoxin A-Induced Apoptosis. Food Chem. Toxicol. 2023, 176, 113793. [Google Scholar] [CrossRef]
- Pfohl-Leszkowicz, A.; Manderville, R.A. Ochratoxin A: An Overview on Toxicity and Carcinogenicity in Animals and Humans. Mol. Nutr. Food Res. 2007, 51, 61–99. [Google Scholar] [CrossRef]
- Zhao, T.; Shen, X.L.; Chen, W.; Liao, X.; Yang, J.; Wang, Y.; Zou, Y.; Fang, C. Advances in Research of Nephrotoxicity and Toxic Antagonism of Ochratoxin A. Toxin Rev. 2016, 36, 39–44. [Google Scholar] [CrossRef]
- Ringot, D.; Chango, A.; Schneider, Y.J.; Larondelle, Y. Toxicokinetics and Toxicodynamics of Ochratoxin A, an Update. Chem. Biol. Interact. 2006, 159, 18–46. [Google Scholar] [CrossRef]
- Wu, Q.; Kuca, K.; Nepovimova, E.; Wu, W. Type A Trichothecene Diacetoxyscirpenol-Induced Emesis Corresponds to Secretion of Peptide YY and Serotonin in Mink. Toxins 2020, 12, 419. [Google Scholar] [CrossRef] [PubMed]
- Yang, X.; Liu, P.; Cui, Y.; Xiao, B.; Liu, M.; Song, M.; Huang, W.; Li, Y. Review of the Reproductive Toxicity of T-2 Toxin. J. Agric. Food Chem. 2020, 68, 727–734. [Google Scholar] [CrossRef] [PubMed]
- Cimbalo, A.; Frangiamone, M.; Font, G.; Manyes, L. The Importance of Transcriptomics and Proteomics for Studying Molecular Mechanisms of Mycotoxin Exposure: A Review. Food Chem. Toxicol. 2022, 169, 113396. [Google Scholar] [CrossRef]
- Rocha, O.; Ansari, K.; Doohan, F.M. Effects of Trichothecene Mycotoxins on Eukaryotic Cells: A Review. Food Addit. Contam. 2005, 22, 369–378. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Wang, Y.; Deng, Y.; Wang, X.; Wu, W.; Nepovimova, E.; Wu, Q.; Kuca, K. Toxic Mechanisms of the Trichothecenes T-2 Toxin and Deoxynivalenol on Protein Synthesis. Food Chem. Toxicol. 2022, 164, 113044. [Google Scholar] [CrossRef]
- Ogunade, I.M.; Martinez-Tuppia, C.; Queiroz, O.C.M.; Jiang, Y.; Drouin, P.; Wu, F.; Vyas, D.; Adesogan, A.T. Silage Review: Mycotoxins in Silage: Occurrence, Effects, Prevention, and Mitigation. J. Dairy Sci. 2018, 101, 4034–4059. [Google Scholar] [CrossRef]
- Malir, F.; Ostry, V.; Pfohl-Leszkowicz, A.; Novotna, E. Ochratoxin A: Developmental and Reproductive Toxicity-an Overview. Birth Defects Res. B Dev. Reprod. Toxicol. 2013, 98, 493–502. [Google Scholar] [CrossRef]
- Smith, M.C.; Madec, S.; Coton, E.; Hymery, N. Natural Co-Occurrence of Mycotoxins in Foods and Feeds and Their in Vitro Combined Toxicological Effects. Toxins 2016, 8, 94. [Google Scholar] [CrossRef]
- Roell, K.R.; Reif, D.M.; Motsinger-Reif, A.A. An Introduction to Terminology and Methodology of Chemical Synergy—Perspectives from Across Disciplines. Front. Pharmacol. 2017, 8, 158. [Google Scholar] [CrossRef]
- Speijers, G.J.A.; Speijers, M.H.M. Combined Toxic Effects of Mycotoxins. Toxicol. Lett. 2004, 153, 91–98. [Google Scholar] [CrossRef]
- Kifer, D.; Jakšić, D.; Šegvić Klarić, M. Assessing the Effect of Mycotoxin Combinations: Which Mathematical Model Is (the Most) Appropriate? Toxins 2020, 12, 153. [Google Scholar] [CrossRef] [PubMed]
- Bonde, J.P.E.; Ernst, E.; Jensen, T.K.; Hjollund, I.N.H.; Kolstad, H.; Henriksen, T.B.; Scheike, T.; Giwercman, A.; Olsen, J.; Skakkebæk, N.E. Relation between Semen Quality and Fertility: A Population-Based Study of 430 First-Pregnancy Planners. Lancet 1998, 352, 1172–1177. [Google Scholar] [CrossRef] [PubMed]
- Komsky-Elbaz, A.; Kalo, D.; Roth, Z. New Evidence for Deleterious Effects of Environmental Contaminants on the Male Gamete. Anim. Reprod. Sci. 2021, 246, 106886. [Google Scholar] [CrossRef] [PubMed]
- Leisegang, K.; Dutta, S. Do Lifestyle Practices Impede Male Fertility? Andrologia 2021, 53, e13595. [Google Scholar] [CrossRef] [PubMed]
- Mann, U.; Shiff, B.; Patel, P. Reasons for Worldwide Decline in Male Fertility. Curr. Opin. Urol. 2020, 30, 296–301. [Google Scholar] [CrossRef]
- Eze, U.A.; Okonofua, F.E. High Prevalence of Male Infertility in Africa: Are Mycotoxins to Blame? Afr. J. Reprod. Health 2015, 19, 9–17. [Google Scholar]
- Eze, U.A.; Routledge, M.N.; Okonofua, F.E.; Huntriss, J.; Gong, Y.Y. Mycotoxin Exposure and Adverse Reproductive Health Outcomes in Africa: A Review. World Mycotoxin J. 2018, 11, 321–339. [Google Scholar] [CrossRef]
- El Khoury, D.; Fayjaloun, S.; Nassar, M.; Sahakian, J.; Aad, P.Y. Updates on the Effect of Mycotoxins on Male Reproductive Efficiency in Mammals. Toxins 2019, 11, 515. [Google Scholar] [CrossRef]
- Komsky-Elbaz, A.; Saktsier, M.; Roth, Z. Aflatoxin B1 Impairs Sperm Quality and Fertilization Competence. Toxicology 2018, 393, 42–50. [Google Scholar] [CrossRef]
- Komsky-Elbaz, A.; Kalo, D.; Roth, Z. Effect of Aflatoxin B1 on Bovine Spermatozoa’s Proteome and Embryo’s Transcriptome. Reproduction 2020, 160, 709–723. [Google Scholar] [CrossRef]
- Winkler, J.; Kersten, S.; Meyer, U.; Engelhardt, U.; Dänicke, S. Residues of Zearalenone (ZEN), Deoxynivalenol (DON) and Their Metabolites in Plasma of Dairy Cows Fed Fusarium Contaminated Maize and Their Relationships to Performance Parameters. Food Chem. Toxicol. 2014, 65, 196–204. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Z.; Fan, Z.; Nie, D.; Zhao, Z.; Han, Z. Analysis of the Carry-Over of Ochratoxin A from Feed to Milk, Blood, Urine, and Different Tissues of Dairy Cows Based on the Establishment of a Reliable LC-MS/MS Method. Molecules 2019, 24, 2823. [Google Scholar] [CrossRef] [PubMed]
- Gallo, A.; Mosconi, M.; Trevisi, E.; Santos, R.R. Adverse Effects of Fusarium Toxins in Ruminants: A Review of In Vivo and In Vitro Studies. Dairy 2022, 3, 474–499. [Google Scholar] [CrossRef]
- Thomas, C.A.; Garner, D.L.; DeJarnette, J.M.; Marshall, C.E. Fluorometric Assessments of Acrosomal Integrity and Viability in Cryopreserved Bovine Spermatozoa. Biol. Reprod. 1997, 56, 991–998. [Google Scholar] [CrossRef] [PubMed]
- Sellem, E.; Broekhuijse, M.L.W.J.; Chevrier, L.; Camugli, S.; Schmitt, E.; Schibler, L.; Koenen, E.P.C. Use of Combinations of In Vitro Quality Assessments to Predict Fertility of Bovine Semen. Theriogenology 2015, 84, 1447–1454. [Google Scholar] [CrossRef] [PubMed]
- Rodríguez-Martínez, H. Sperm Function in Cattle and Pigs: Morphological and Functional Aspects|Request PDF. Arch. fur Tierzucht 2001, 44, 103–113. [Google Scholar]
- Rodríguez-Martínez, H. Laboratory Semen Assessment and Prediction of Fertility: Still Utopia? Reprod. Domest. Anim. 2003, 38, 312–318. [Google Scholar] [CrossRef]
- Garner, D.L.; Pinkel, D.; Johnson, L.A.; Pace, M.M. Assessment of Spermatozoal Function Using Dual Fluorescent Staining and Flow Cytometric Analyses. Biol. Reprod. 1986, 34, 127–138. [Google Scholar] [CrossRef]
- Hossain, M.S.; Johannisson, A.; Wallgren, M.; Nagy, S.; Siqueira, A.P.; Rodriguez-Martinez, H. Flow Cytometry for the Assessment of Animal Sperm Integrity and Functionality: State of the Art. Asian J. Androl. 2011, 13, 406–419. [Google Scholar] [CrossRef]
- Sambuu, R.; Takagi, M.; Namula, Z.; Nii, M.; Taniguchi, M.; Uno, S.; Kokushi, E.; Tshering, C.; Dos Santos, R.R.; Fink-Gremmels, J.; et al. Effects of Long-Term in Vitro Exposure of Ejaculated Boar Sperm to Zearalenone and α-Zearalenol in Sperm Liquid Storage Medium. Anim. Sci. J. 2013, 84, 28–34. [Google Scholar] [CrossRef]
- Zhang, T.Y.; Wu, R.Y.; Zhao, Y.; Xu, C.S.; Zhang, W.D.; Ge, W.; Liu, J.; Sun, Z.Y.; Zou, S.H.; Shen, W. Ochratoxin A Exposure Decreased Sperm Motility via the AMPK and PTEN Signaling Pathways. Toxicol. Appl. Pharmacol. 2018, 340, 49–57. [Google Scholar] [CrossRef] [PubMed]
- Tkachev, A.V.; Tkacheva, O.L.; Zubova, T.V.; Pleshkov, V.A.; Smolovskaya, O.V. Effect of Mycotoxins on the Spermatozoa and Embryos of Animals. Adv. Anim. Vet. Sci. 2020, 8, 47–55. [Google Scholar] [CrossRef]
- Yousef, M.S.; Rezk, W.R.; El-Naby, A.; Shimaa, A.H.H.; Mahmoud, K.G.M.; Takagi, M.; Miyamoto, A.; Megahed, G.A. In Vitro Effect of Zearalenone on Sperm Parameters, Oocyte Maturation and Embryonic Development in Buffalo. Reprod. Biol. 2023, 23, 100732. [Google Scholar] [CrossRef]
- Tassis, P.D.; Tsakmakidis, I.A.; Nagl, V.; Reisinger, N.; Tzika, E.; Gruber-Dorninger, C.; Michos, I.; Mittas, N.; Basioura, A.; Schatzmayr, D. Individual and Combined In Vitro Effects of Deoxynivalenol and Zearalenone on Boar Semen. Toxins 2020, 12, 495. [Google Scholar] [CrossRef]
- Benzoni, E.; Minervini, F.; Giannoccaro, A.; Fornelli, F.; Vigo, D.; Visconti, A. Influence of in Vitro Exposure to Mycotoxin Zearalenone and Its Derivatives on Swine Sperm Quality. Reprod. Toxicol. 2008, 25, 461–467. [Google Scholar] [CrossRef]
- Buffone, M.G.; Foster, J.A.; Gerton, G.L. The Role of the Acrosomal Matrix in Fertilization. Int. J. Dev. Biol. 2008, 52, 511–522. [Google Scholar] [CrossRef] [PubMed]
- Tsakmakidis, I.A.; Lymberopoulos, A.G.; Alexopoulos, C.; Boscos, C.M.; Kyriakis, S.C. In Vitro Effect of Zearalenone and α-Zearalenol on Boar Sperm Characteristics and Acrosome Reaction. Reprod. Domest. Anim. 2006, 41, 394–401. [Google Scholar] [CrossRef]
- Filannino, A.; Stout, T.A.E.; Gadella, B.M.; Sostaric, E.; Pizzi, F.; Colenbrander, B.; Dell’Aquila, M.E.; Minervini, F. Dose-Response Effects of Estrogenic Mycotoxins (Zearalenone, Alpha- and Beta-Zearalenol) on Motility, Hyperactivation and the Acrosome Reaction of Stallion Sperm. Reprod. Biol. Endocrinol. 2011, 9, 134. [Google Scholar] [CrossRef]
- Kasai, T.; Ogawa, K.; Mizuno, K.; Nagai, S.; Uchida, Y.; Ohta, S.; Fujie, M.; Suzuki, K.; Hirata, S.; Hoshi, K. Relationship between Sperm Mitochondrial Membrane Potential, Sperm Motility, and Fertility Potential. Asian J. Androl. 2002, 4, 97–103. [Google Scholar]
- Agnihotri, S.K.; Agrawal, A.K.; Hakim, B.A.; Vishwakarma, A.L.; Narender, T.; Sachan, R.; Sachdev, M. Mitochondrial Membrane Potential (MMP) Regulates Sperm Motility. Vitr. Cell. Dev. Biol. Anim. 2016, 52, 953–960. [Google Scholar] [CrossRef]
- Amaral, A.; Lourenço, B.; Marques, M.; Ramalho-Santos, J. Mitochondria Functionality and Sperm Quality. Reproduction 2013, 146, R163–R174. [Google Scholar] [CrossRef] [PubMed]
- Rivlin, J.; Mendel, J.; Rubinstein, S.; Etkovitz, N.; Breitbart, H. Role of Hydrogen Peroxide in Sperm Capacitation and Acrosome Reaction. Biol. Reprod. 2004, 70, 518–522. [Google Scholar] [CrossRef] [PubMed]
- Du Plessis, S.S.; Agarwal, A.; Halabi, J.; Tvrda, E. Contemporary Evidence on the Physiological Role of Reactive Oxygen Species in Human Sperm Function. J. Assist. Reprod. Genet. 2015, 32, 509–520. [Google Scholar] [CrossRef] [PubMed]
- Agarwal, A.; Saleh, R.A.; Bedaiwy, M.A. Role of Reactive Oxygen Species in the Pathophysiology of Human Reproduction. Fertil. Steril. 2003, 79, 829–843. [Google Scholar] [CrossRef]
- Wang, X.; Sharma, R.K.; Gupta, A.; George, V.; Thomas, A.J.; Falcone, T.; Agarwal, A. Alterations in Mitochondria Membrane Potential and Oxidative Stress in Infertile Men: A Prospective Observational Study. Fertil. Steril. 2003, 80, 844–850. [Google Scholar] [CrossRef]
- Tvrdá, E.; Greifová, H.; Ďuračka, M.; Ondruška, Ľ.; Halenár, M.; Kolesárová, A. Comparative Analysis of the Detrimental in Vitro Effects of Three Fusariotoxins on the Selected Structural and Functional Characteristics of Rabbit Spermatozoa. Drug Chem. Toxicol. 2021, 45, 2519–2527. [Google Scholar] [CrossRef]
- Park, H.; Park, H.S.; Lim, W.; Song, G. Ochratoxin A Suppresses Proliferation of Sertoli and Leydig Cells in Mice. Med. Mycol. 2020, 58, 71–82. [Google Scholar] [CrossRef]
- Bose, S.; Sinha, S.P. Modulation of Ochratoxin-Produced Genotoxicity in Mice by Vitamin C. Food Chem. Toxicol. 1994, 32, 533–537. [Google Scholar] [CrossRef]
- Chakraborty, D.; Verma, R. Ameliorative Effect of Emblica Officinalis Aqueous Extract on Ochratoxin-Induced Lipid Peroxidation in the Kidney and Liver of Mice. Int. J. Occup. Med. Environ. Health 2010, 23, 63–73. [Google Scholar] [CrossRef]
- Ribelin, W.E.; Fukushima, K.; Still, P.E. The Toxicity of Ochratoxin to Ruminants. Can. J. Comp. Med. 1978, 42, 172. [Google Scholar]
- Tkachev, A.V.; Tkacheva, O.L. Comparison of the Cytotoxic Effects of Zearalenon and T-2 Toxin on the Horses and Oxen Germ Cell in Vitro before and after Cryopreservation. Tsitologiia 2017, 59, 45–52. [Google Scholar] [PubMed]
- Elweza, A.E.; Marey, M.A.; Elesh, I.F.; Zinnah, M.A.; Akthar, I.; Kanno, C.; Takagi, M.; Miyamoto, A. Zearalenone Interferes with the Sperm-Triggered Inflammation in the Bovine Uterus in Vitro: Negative Impact on Sperm Motility and Survival. Reprod. Toxicol. 2022, 107, 81–89. [Google Scholar] [CrossRef]
- Tsakmakidis, I.A.; Lymberopoulos, A.G.; Khalifa, T.A.A.; Boscos, C.M.; Saratsi, A.; Alexopoulos, C. Evaluation of Zearalenone and α-Zearalenol Toxicity on Boar Sperm DNA Integrity. J. Appl. Toxicol. 2008, 28, 681–688. [Google Scholar] [CrossRef] [PubMed]
- Richardson, K.; Hagler, W.; Mirocha, C. Production of Zearalenone, Alpha- and Beta-Zearalenol, and Alpha- and Beta-Zearalanol by Fusarium Spp. in Rice Culture. J. Agric. Food Chem. 1985, 33, 862–866. [Google Scholar] [CrossRef]
- Ying Yang, J.; Fa Zhang, Y.; Liang, A.M.; Feng Kong, X.; Xiao Li, Y.; Wang Ma, K.; Jing, A.H.; Ying Feng, S.; Lan Qiao, X. Toxic Effects of T-2 Toxin on Reproductive System in Male Mice. Toxicol. Ind. Health 2010, 26, 25–31. [Google Scholar] [CrossRef]
- Yang, X.; Zhang, X.; Zhang, J.; Ji, Q.; Huang, W.; Zhang, X.; Li, Y. Spermatogenesis Disorder Caused by T-2 Toxin Is Associated with Germ Cell Apoptosis Mediated by Oxidative Stress. Environ. Pollut. 2019, 251, 372–379. [Google Scholar] [CrossRef]
- Kovács, M.; Tornyos, G.; Matics, Z.; Kametler, L.; Rajli, V.; Bodnár, Z.; Kulcsár, M.; Huszenicza, G.; Keresztes, Z.; Cseh, S. Subsequent Effect of Subacute T-2 Toxicosis on Spermatozoa, Seminal Plasma and Testosterone Production in Rabbits. Animal 2011, 5, 1563–1569. [Google Scholar] [CrossRef]
- Alm, K.; Dahlbom, M.; Säynäjärvi, M.; Andersson, M.A.; Salkinoja-Salonen, M.S.; Andersson, M.C. Impaired Semen Quality of AI Bulls Fed with Moldy Hay: A Case Report. Theriogenology 2002, 58, 1497–1502. [Google Scholar] [CrossRef] [PubMed]
- Brake, J.; Hamilton, P.B.; Kittrell, R.S. Effects of the Tricothecene Mycotoxin Diacetoxyscirpenol on Fertility and Hatchability of Broiler Breeders. Poult. Sci. 1999, 78, 1690–1694. [Google Scholar] [CrossRef]
- Hassanane, M.S.; Abdalla, E.S.A.; EI-Fiky, S.; Amer, M.A.; Hamdy, A. Mutagenicity of the Mycotoxin Diacetoxyscirpenol on Somatic and Germ Cells of Mice. Mycotoxin Res. 2000, 16, 53–64. [Google Scholar] [CrossRef]
- Rodrigues, I.; Naehrer, K. A Three-Year Survey on the Worldwide Occurrence of Mycotoxins in Feedstuffs and Feed. Toxins 2012, 4, 663–675. [Google Scholar] [CrossRef] [PubMed]
- Kemboi, D.C.; Antonissen, G.; Ochieng, P.E.; Croubels, S.; Okoth, S.; Kangethe, E.K.; Faas, J.; Lindahl, J.F.; Gathumbi, J.K. A Review of the Impact of Mycotoxins on Dairy Cattle Health: Challenges for Food Safety and Dairy Production in Sub-Saharan Africa. Toxins 2020, 12, 222. [Google Scholar] [CrossRef] [PubMed]
- Prabu, P.C.; Dwivedi, P.; Sharma, A.K. Toxicopathological Studies on the Effects of Aflatoxin B(1), Ochratoxin A and Their Interaction in New Zealand White Rabbits. Exp. Toxicol. Pathol. 2013, 65, 277–286. [Google Scholar] [CrossRef]
- Ficheux, A.S.; Sibiril, Y.; Parent-Massin, D. Co-Exposure of Fusarium Mycotoxins: InVitro Myelotoxicity Assessment on Human Hematopoietic Progenitors. Toxicon 2012, 60, 1171–1179. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Zhang, B.; He, X.; Cheng, W.H.; Xu, W.; Luo, Y.; Liang, R.; Luo, H.; Huang, K. Analysis of Individual and Combined Effects of Ochratoxin A and Zearalenone on HepG2 and KK-1 Cells with Mathematical Models. Toxins 2014, 6, 1177. [Google Scholar] [CrossRef] [PubMed]
- Harvey, R.B.; Kubena, L.F.; Elissalde, M.H.; Rottinghaus, G.E.; Corrier, D.E. Administration of Ochratoxin A and T-2 Toxin to Growing Swine. Am. J. Vet. Res. 1994, 55, 1757–1761. [Google Scholar] [PubMed]
- Kubena, L.F.; Harvey, R.B.; Edrington, T.S.; Rottinghaus, G.E. Influence of Ochratoxin A and Diacetoxyscirpenol Singly and in Combination on Broiler Chickens. Poult. Sci. 1994, 73, 408–415. [Google Scholar] [CrossRef]
- Hood, R.D.; Kuczuk, M.H.; Szczech, G.M. Effects in Mice of Simultaneous Prenatal Exposure to Ochratoxin A and T-2 Toxin. Teratology 1978, 17, 25–29. [Google Scholar] [CrossRef]
- Bouaziz, C.; Sharaf el Dein, O.; El Golli, E.; Abid-Essefi, S.; Brenner, C.; Lemaire, C.; Bacha, H. Different Apoptotic Pathways Induced by Zearalenone, T-2 Toxin and Ochratoxin A in Human Hepatoma Cells. Toxicology 2008, 254, 19–28. [Google Scholar] [CrossRef]
- Barbagallo, F.; La Vignera, S.; Cannarella, R.; Aversa, A.; Calogero, A.E.; Condorelli, R.A. Evaluation of Sperm Mitochondrial Function: A Key Organelle for Sperm Motility. J. Clin. Med. 2020, 9, 363. [Google Scholar] [CrossRef]
- Szabo Fodor, J.; Kachlek, M.; Cseh, S.; Somoskői, B.; Szabó, A.; Bodnár, Z.; Tornyos, G.; Mezes, M.; Balogh, K.; Glávits, R.; et al. Individual and Combined Effects of Subchronic Exposure of Three Fusarium Toxins (Fumonisin B, Deoxynivalenol and Zearalenone) in Rabbit Bucks. J. Clin. Toxicol. 2015, 5, 264. [Google Scholar] [CrossRef]
- Klarić, M.Š.; Rumora, L.; Ljubanović, D.; Pepeljnjak, S. Cytotoxicity and Apoptosis Induced by Fumonisin B1, Beauvericin and Ochratoxin a in Porcine Kidney PK15 Cells: Effects of Individual and Combined Treatment. Arch. Toxicol. 2008, 82, 247–255. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.W.; Wang, J.Q.; Zheng, B.Q.; Li, S.L.; Zhang, Y.D.; Li, F.D.; Zheng, N. Cytotoxicity Induced by Ochratoxin A, Zearalenone, and α-Zearalenol: Effects of Individual and Combined Treatment. Food Chem. Toxicol. 2014, 71, 217–224. [Google Scholar] [CrossRef] [PubMed]
- Orgal, S.; Zeron, Y.; Elior, N.; Biran, D.; Friedman, E.; Druker, S.; Roth, Z. Season-Induced Changes in Bovine Sperm Motility Following a Freeze-Thaw Procedure. J. Reprod. Dev. 2012, 58, 212–218. [Google Scholar] [CrossRef] [PubMed]
- Komsky-Elbaz, A.; Roth, Z. Fluorimetric Techniques for the Assessment of Sperm Membranes. Jove 2018, 114, e58622. [Google Scholar]
- Kalo, D.; Reches, D.; Netta, N.; Komsky-Elbaz, A.; Zeron, Y.; Moallem, U.; Roth, Z. Carryover Effects of Feeding Bulls with an Omega-3-Enriched-Diet—From Spermatozoa to Developed Embryos. PLoS ONE 2022, 17, e0265650. [Google Scholar] [CrossRef]
- Weber, F.; Freudinger, R.; Schwerdt, G.; Gekle, M. A Rapid Screening Method to Test Apoptotic Synergisms of Ochratoxin A with Other Nephrotoxic Substances. Toxicol. In Vitr. 2005, 19, 135–143. [Google Scholar] [CrossRef]

{kind=link}
| Group | * Viability (%) | p-Value | ** Mitochondrial Membrane Potential Log(Ratio) | p-Value | * Intact Acrosome Membrane (%) | p-Value | * Viable Spermatozoa with ROS (%) | p-Value |
|---|---|---|---|---|---|---|---|---|
| Untreated | 43.71 ± 2.7 | 0.98 | 0.17 ± 0.2 | 0.25 | 56.35 ± 1.9 | 0.89 | 6.94 ± 1.2 | 0.32 |
| Control | 43.64 ± 3.5 | 0.53 ± 0.2 | 56.73 ± 2.0 | 8.67 ± 1.2 |
| Mycotoxin | Concentration (ppm) | * Viability (%) | ** Mitochondrial Membrane Potential | * Acrosome Membrane Integrity (%) | * Oxidation Status of Viable Spermatozoa (%) | ||
|---|---|---|---|---|---|---|---|
| Damage | Intact | ROS- | ROS+ | ||||
| DAS | 0 (control) | 37.9 ± 8.5 | 0.23 ± 0.42 | 6.9 ± 0.5 | 59.9 ± 1.9 | 53.4 ± 10.1 | 9.4 ± 2.4 |
| 0.01 | 44.3 ± 4.9 | −0.07 ± 0.42 | 6.5 ± 2.4 | 59.2 ± 2.4 | 53.5 ± 6.1 | 8.9 ± 1.6 | |
| 0.05 | 44.9 ± 4.1 | −0.25 ± 0.34 | 5.0 ± 1.4 | 62.0 ± 1.2 | 55.4 ± 10.0 | 7.7 ± 2.1 | |
| 0.1 | 44.8 ± 4.3 | 0.36 ± 0.48 | 4.9 ± 1.4 | 61.0 ± 1.6 | 45.3 ± 7.2 | 8.8 ± 1.5 | |
| 0.5 | 44.0 ± 5.2 | −0.02 ± 0.39 | 7.9 ± 2.1 | 56.7 ± 2.6 | 47.3 ± 7.1 | 7.2 ± 1.9 | |
| OTA | 0 (control) | 47.9 ± 5.1 | 0.32 ± 0.34 | 5.9 ± 1.4 | 52.4 ± 5.1 | 66.3 ± 5.1 | 3.9 ± 0.9 |
| 0.05 | 47.2 ± 5.1 | 0.07 ± 0.33 | 6.6 ± 2.6 | 54.9 ± 5.0 | 65.9 ± 5.2 | 4.1 ± 1.1 | |
| 0.1 | 44.3 ± 4.7 | 0.24 ± 0.38 | 5.1 ± 1.0 | 57.1 ± 5.3 | 59.9 ± 7.2 | 3.8 ± 1.2 | |
| 0.25 | 43.1 ± 3.5 | 0.33 ± 0.49 | 6.9 ± 2.6 | 53.6 ± 5.4 | 63.0 ± 5.5 | 4.8 ± 1.0 | |
| 1 | 44.5 ± 3.2 | 0.46 ± 0.36 | 4.4 ± 1.7 | 55.1 ± 5.8 | 62.3 ± 2.5 | 5.5 ± 1.6 | |
| 10 | 44.8 ± 5.2 | 0.18 ± 029 | 6.4 ± 1.1 | 55.4 ± 5.9 | 62.3 ± 3.9 | 3.4 ± 0.8 | |
| T2 | 0 (control) | 38.5 ± 9.1 | −0.04 ± 0.35 | 9.2 ± 2.3 | 59.2 ± 6.5 | 61.7 ± 8.4 | 7.4 ± 1.5 |
| 0.01 | 41.5 ± 7.1 | 0.15 ± 0.28 | 7.9 ± 2.0 | 58.3 ± 5.3 | 63.2 ± 8.1 | 5.4 ± 1.2 | |
| 0.05 | 41.2 ± 7.7 | 0.31 ± 0.56 | 9.0 ± 2.5 | 58.2 ± 6.1 | 60.6 ± 6.2 | 6.4 ± 1.1 | |
| 0.1 | 38.9 ± 6.4 | 0.42 ± 0.44 | 8.5 ± 2.2 | 58.7 ± 5.7 | 61.6 ± 7.7 | 6.6 ± 2.1 | |
| 0.5 | 45.3 ± 7.0 | 0.19 ± 0.32 | 10.2 ± 2.0 | 52.9 ± 5.9 | 56.2 ± 7.7 | 11.4 ± 4.2 | |
| ZEN | 0 (control) | 49.2 ± 5.4 | 1.53 ± 0.53 | 3.0 ± 0.5 | 55.7 ± 2.3 | 29.0 ± 1.1 | 7.1 ± 1.4 |
| 0.1 | 47.1 ± 4.7 | 0.87 ± 0.26 | 3.1 ± 0.4 | 57.3 ± 2.1 | 24.8 ± 3.4 | 8.7 ± 1.6 | |
| 0.5 | 43.6 ± 4.9 | 1.05 ± 0.52 | 4.6 ± 1.3 | 54.6 ± 2.2 | 24.8 ± 1.3 | 8.4 ± 0.6 | |
| 1 | 48.2 ± 5.2 | 1.36 ± 0.53 | 3.3 ± 0.4 | 55.8 ± 2.8 | 25.2 ± 2.5 | 8.2 ± 2.1 | |
| 5 | 46.5 ± 5.3 | 0.37 ± 0.26 | 4.1 ± 1.0 | 55.4 ± 2.5 | 26.6 ± 1.6 | 10.4 ± 0.6 | |
| 10 | 48.2 ± 3.5 | 0.71 ± 0.32 | 5.1 ± 1.3 | 53.6 ± 2.2 | 33.5 ± 1.5 | 3.3 ± 0.6 | |
| Mixture | * Mixture | ** Viability (%) | *** Mitochondrial Membrane Potential | ** Acrosomal Membrane Integrity (%) | ** Oxidation Status of Viable Spermatozoa (%) | ||
|---|---|---|---|---|---|---|---|
| Damage | Intact | ROS- | ROS+ | ||||
| DAS + T2 | Control | 46.0 ± 6.8 | 0.42 ± 0.5 | 3.2 ± 1.5 | 64.4 ± 6.9 | 35.2 ± 2.4 | 8.0 ± 2.8 |
| Low | 53.8 ± 4.5 | 0.29 ± 0.3 | 3.0 ± 0.8 | 64.3 ± 0.9 | 32.9 ± 1.5 | 10.1 ± 2.3 | |
| Intermediate | 43.1 ± 9.2 | −0.29 ± 0.3 | 3.5 ± 2.2 | 67.0 ± 3.7 | 34.4 ± 5.5 | 11.1 ± 2.6 | |
| High | 43.9 ± 9.3 | 0.25 ± 0.3 | 5.1 ± 1.7 | 62.9 ± 3.4 | 30.7 ± 3.2 | 10.4 ± 3.4 | |
| OTA + DAS | Control | 56.8 ± 8.4 | 0.58 ± 0.2 | 4.5 ± 1.0 | 63.8 ± 6.7 | 38.6 ± 10.8 | 18.9 ± 1.5 |
| Low | 60.3 ± 7.9 | 0.51 ± 0.1 | 3.5 ± 1.4 | 66.5 ± 8.5 | 36.2 ± 10.2 | 16.9 ± 2.4 | |
| Intermediate | 56.3 ± 7.3 | 0.85 ± 0.2 | 4.1 ± 1.4 | 64.9 ± 7.7 | 29.5 ± 7.6 | 20.03 ± 1.2 | |
| High | 56.3 ± 7.6 | 0.49 ± 0.4 | 3.2 ± 0.8 | 66.1 ± 7.4 | 38.7 ± 9.9 | 14.9 ± 4.0 | |
| OTA + T2 | Control | 37.2 ± 10.8 | 0.89 ± 0.2 | 11.2 ± 3.8 | 54.8 ± 2.6 | 37.8 ± 4.7 | 17.1 ± 2.4 |
| Low | 37.8 ± 11.8 | 1.26 ± 0.1 | 6.4 ± 1.2 | 59.7 ± 3.8 | 39.7 ± 4.8 | 16.7 ± 3.0 | |
| Intermediate | 37.2 ± 10.8 | 1.21 ± 0.1 | 7.6 ± 3.8 | 58.5 ± 4.5 | 31.7 ± 5.8 | 16.6 ± 3.4 | |
| High | 39.1 ± 11.5 | 1.05 ± 0.1 | 9.9 ± 5.6 | 58.5 ± 7.5 | 35.01 ± 4.5 | 20.5 ± 2.9 | |
| OTA + ZEN | Control | 42.3 ± 13.7 | 0.69 ± 0.3 | 9.4 ± 3.5 | 69.2 ± 0.8 | 14.8 ± 4.7 | 44.1 ± 8.0 |
| Low | 38.7 ± 12.2 | 0.93 ± 0.9 | 8.2 ± 4.1 | 70.1 ± 4.2 | 15.6 ± 5.0 | 42.1 ± 8.2 | |
| Intermediate | 37.9 ± 12.9 | 0.69 ± 0.4 | 8.6 ± 2.0 | 72.4 ± 2.1 | 13.3 ± 5.1 | 41.9 ± 9.7 | |
| High | 38.8 ± 8.6 | 0.66 ± 0.3 | 11.6 ± 2.6 | 67.1 ± 3.5 | 11.8 ± 3.9 | 45.3 ± 9.8 | |
| ZEN + T2 | Control | 57.1 ± 11.7 | 2.09 ± 0.6 | 4.8 ± 0.9 | 69.8 ± 1.8 | 30.6 ± 8.6 | 19.0 ± 0.3 |
| Low | 55.3 ± 7.0 | 0.87 ± 0.1 | 3.4 ± 0.5 | 70.9 ± 2.3 | 35.2 ± 6.9 | 15.2 ± 1.6 | |
| Intermediate | 57.3 ± 8.8 | 1.08 ± 0.1 | 4.2 ± 1.8 | 73.5 ± 2.1 | 36.7 ± 11.1 | 12.7 ± 4.2 | |
| High | 56.3 ± 7.6 | 0.79 ± 0.2 | 2.8 ± 0.5 | 74.0 ± 3.5 | 34.2 ± 7.8 | 17.4 ± 0.3 | |
| ZEN + DAS | Control | 51.3 ± 11.0 | 0.81 ± 0.9 | 3.0 ± 1.7 | 62.7 ± 4.0 | 65.6 ± 4.3 | 7.8 ± 2.4 |
| Low | 57.3 ± 8.4 | 0.36 ± 0.9 | 3.1 ± 0.8 | 60.4 ± 6.7 | 53.8 ± 5.4 | 8.6 ± 0.9 | |
| Intermediate | 49.7 ± 9.7 | 1.04 ± 0.3 | 2.3 ± 0.6 | 61.5 ± 5.6 | 47.6 ± 8.9 | 10.0 ± 0.9 | |
| High | 54.1 ± 7.2 | 0.06 ± 0.8 | 3.4 ± 0.5 | 60.9 ± 4.5 | 50.0 ± 12.5 | 9.7 ± 2.4 | |
| (a) | ||||||
| Mixture | Examined Parameter | Concentration Combination | Expected Value | Measured Value | p-Value | Interaction |
| DAS + T2 | Viable | Low | −5.7 ± 4.68 | 7.80 ± 5.28 | 0.1 | Additive |
| Intermediate | −6.81 ± 5.57 | −2.84 ± 3.63 | 0.57 | Additive | ||
| High | −6.67 ± 6.05 | −2.14 ± 3.42 | 0.54 | Additive | ||
| ΔΨm | Low | −1.48 ± 1.36 | −0.13 ± 0.12 | 0.36 | Additive | |
| Intermediate | −1.24 ± 1.08 | −0.71 ± 0.22 | 0.65 | Additive | ||
| High | −1.45 ± 1.19 | −0.17 ± 0.58 | 0.37 | Additive | ||
| Damaged | Low | −2.88 ± 1.68 | −0.21 ± 0.86 | 0.21 | Additive | |
| acrosome | Intermediate | −1.73 ± 1.22 | 0.29 ± 0.95 | 0.24 | Additive | |
| High | −0.96 ± 3.26 | 1.61 ± 1.37 | 0.45 | Additive | ||
| Intact | Low | 2.47 ± 1.08 | −0.08 ± 6.58 | 0.71 | Additive | |
| acrosome | Intermediate | 0.83 ± 4.09 | 2.51 ± 4.00 | 0.71 | Additive | |
| High | −4.16 ± 6.62 | −1.56 ± 5.42 | 0.72 | Additive | ||
| Viable | Low | −8.69 ± 7.04 | −2.29 ± 1.55 | 0.41 | Additive | |
| ROS- | Intermediate | −7.44 ± 10.76 | −0.83 ± 3.14 | 0.58 | Additive | |
| High | −4.55 ± 10.22 | −4.49 ± 1.13 | 0.99 | Additive | ||
| Viable | Low | 0.49 ± 2.49 | 2.14 ± 1.32 | 0.58 | Additive | |
| ROS+ | Intermediate | −3.72 ± 3.54 | 3.13 ± 0.39 | 0.1 | Additive | |
| High | −4.96 ± 2.29 | 2.47 ± 2.11 | 0.1 | Additive | ||
| (b) | ||||||
| Mixture | Examined Parameter | Concentration Combination | Expected Value | Measured Value | p-Value | Interaction |
| OTA + DAS | Viable | Low | −3.61 ± 5.03 | −0.50 ± 1.55 | 0.51 | Additive |
| Intermediate | −11.77 ± 3.39 | −0.48 ± 1.51 * | 0.02 | Synergistic | ||
| High | −4.60 ± 6.79 | 3.49 ± 0.81 | 0.21 | Additive | ||
| ΔΨm | Low | −1.22 ± 0.96 | −0.07 ± 0.23 | 0.31 | Additive | |
| Intermediate | −0.22 ± 0.67 | 0.27 ± 0.46 | 0.64 | Additive | ||
| High | −1.06 ± 0.90 | −0.09 ± 0.67 | 0.51 | Additive | ||
| Damaged | Low | 0.74 ± 1.09 | −0.40 ± 0.85 | 0.32 | Additive | |
| acrosome | Intermediate | 2.92 ± 2.99 | −1.25 ± 0.87 | 0.22 | Additive | |
| High | 2.93 ± 0.84 | −0.96 ± 0.51 * | 0.007 | Antagonist | ||
| Intact | Low | 0.26 ± 1.17 | 1.08 ± 2.14 | 0.78 | Additive | |
| acrosome | Intermediate | −2.32 ± 4.01 | 2.25 ± 1.42 | 0.3 | Additive | |
| High | −7.19 ± 3.51 | 2.71 ± 2.00 * | 0.05 | Synergistic | ||
| Viable | Low | −1.76 ± 3.47 | −2.37 ± 1.76 | 0.77 | Additive | |
| ROS- | Intermediate | −5.16 ± 9.29 | −9.11 ± 4.45 | 0.7 | Additive | |
| High | −6.64 ± 10.81 | 0.15 ± 1.15 | 0.47 | Additive | ||
| Viable | Low | 0.22 ± 2.40 | −2.00 ± 1.41 | 0.44 | Additive | |
| ROS+ | Intermediate | −2.06 ± 3.36 | 1.11 ± 1.48 | 0.44 | Additive | |
| High | −3.66 ± 3.19 | −4.01 ± 3.25 | 0.97 | Additive | ||
| (c) | ||||||
| Mixture | Examined Parameter | Concentration Combination | Expected Value | Measured Value | p-Value | Interaction |
| OTA + T2 | Viable | Low | −1.04 ± 3.04 | 0.58 ± 1.26 | 0.65 | Additive |
| Intermediate | −10.98 ± 4.59 | −0.08 ± 0.69 * | 0.05 | Synergistic | ||
| High | −4.55 ± 4.34 | 1.86 ± 1.10 | 0.2 | Additive | ||
| ΔΨm | Low | −0.69 ± 0.99 | 0.37 ± 0.44 | 0.36 | Additive | |
| Intermediate | −0.21 ± 0.91 | 0.32 ± 0.37 | 0.61 | Additive | ||
| High | −0.63 ± 1.07 | 0.16 ± 0.31 | 0.51 | Additive | ||
| Damaged | Low | −2.65 ± 1.97 | −4.87 ± 3.66 | 0.61 | Additive | |
| acrosome | Intermediate | 0.98 ± 2.99 | −3.64 ± 6.31 | 0.54 | Additive | |
| High | −1.78 ± 3.25 | −1.38 ± 8.70 | 0.97 | Additive | ||
| Intact | Low | 3.78 ± 1.57 | 4.87 ± 3.84 | 0.8 | Additive | |
| acrosome | Intermediate | 1.16 ± 1.42 | 3.71 ± 6.86 | 0.75 | Additive | |
| High | 1.11 ± 2.98 | 3.63 ± 9.77 | 0.81 | Additive | ||
| Viable | Low | −5.19 ± 6.66 | 1.93 ± 3.26 | 0.37 | Additive | |
| ROS- | Intermediate | −5.39 ± 7.24 | −6.08 ± 2.00 | 0.93 | Additive | |
| High | −8.91 ± 5.85 | −2.81 ± 0.31 | 0.33 | Additive | ||
| Viable | Low | −1.17 ± 1.05 | −0.44 ± 3.38 | 0.84 | Additive | |
| ROS+ | Intermediate | −2.91 ± 1.61 | −0.55 ± 1.47 | 0.32 | Additive | |
| High | −1.54 ± 1.09 | 3.36 ± 2.07 | 0.08 | Additive | ||
| (d) | ||||||
| Mixture | Examined Parameter | Concentration Combination | Expected Value | Measured Value | p-Value | Interaction |
| OTA + ZEN | Viable | Low | −1.93 ± 2.66 | −3.64 ± 2.78 | 0.74 | Additive |
| Intermediate | −8.74 ± 3.83 | −4.46 ± 1.72 | 0.38 | Additive | ||
| High | −2.69 ± 7.44 | −3.56 ± 5.21 | 0.86 | Additive | ||
| ΔΨm | Low | −0.74 ± 0.74 | 0.24 ± 0.77 | 0.38 | Additive | |
| Intermediate | 0.67 ± 0.25 | 0.001 ± 0.47 | 0.3 | Additive | ||
| High | −0.85 ± 0.84 | −0.02 ± 0.26 | 0.37 | Additive | ||
| Damaged | Low | 0.63 ± 1.79 | −1.16 ± 1.96 | 0.47 | Additive | |
| acrosome | Intermediate | 4.33 ± 1.44 | −0.77 ± 3.19 | 0.39 | Additive | |
| High | 2.13 ± 3.65 | 2.21 ± 4.08 | 0.99 | Additive | ||
| Intact | Low | 3.26 ± 1.15 | 0.96 ± 3.68 | 0.65 | Additive | |
| acrosome | Intermediate | −4.07 ± 1.42 | 3.23 ± 2.64 | 0.25 | Additive | |
| High | −1.98 ± 3.04 | −2.11 ± 3.67 | 0.98 | Additive | ||
| Viable | Low | −2.48 ± 9.06 | 0.82 ± 0.49 | 0.58 | Additive | |
| ROS- | Intermediate | −6.27 ± 8.17 | −1.55 ± 1.08 | 0.58 | Additive | |
| High | −1.93 ± 5.22 | −2.97 ± 1.67 | 0.86 | Additive | ||
| Viable | Low | 1.06 ± 2.99 | −2.01 ± 0.36 | 0.37 | Additive | |
| ROS+ | Intermediate | 0.84 ± 4.66 | −1.55 ± 2.57 | 0.62 | Additive | |
| High | −3.95 ± 1.80 | 1.18 ± 2.96 | 0.18 | Additive | ||
| (e) | ||||||
| Mixture | Examined Parameter | Concentration Combination | Expected Value | Measured Value | p-Value | Interaction |
| T2 + ZEN | Viable | Low | −3.86 ± 1.92 | 0.19 ± 2.98 | 0.29 | Additive |
| Intermediate | −3.19 ± 5.85 | −0.83 ± 4.3 | 0.76 | Additive | ||
| High | −2.54 ± 6.76 | −1.82 ± 6.48 | 0.94 | Additive | ||
| ΔΨm | Low | −1.08 ± 1.21 | −1.23 ± 0.78 | 0.92 | Additive | |
| Intermediate | −0.49 ± 0.88 | −1.01 ± 0.49 | 0.62 | Additive | ||
| High | −1.45 ± 1.14 | −1.31 ± 0.78 | 0.92 | Additive | ||
| Damaged | Low | −3.37 ± 1.79 | −0.54 ± 0.94 | 0.21 | Additive | |
| acrosome | Intermediate | −1.43 ± 1.44 | −1.97 ± 1.48 | 0.8 | Additive | |
| High | −1.67 ± 3.65 | −1.39 ± 1.42 | 0.94 | Additive | ||
| Intact | Low | 4.93 ± 1.15 | 3.69 ± 1.26 | 0.49 | Additive | |
| acrosome | Intermediate | 0.63 ± 1.42 | 4.19 ± 2.43 | 0.25 | Additive | |
| High | 0.58 ± 3.04 | 1.03 ± 1.24 | 0.89 | Additive | ||
| Viable | Low | −10.56 ± 9.06 | −11.76 ± 6.62 | 0.92 | Additive | |
| ROS- | Intermediate | −7.66 ± 8.17 | −18.01 ± 8.61 | 0.42 | Additive | |
| High | 1.93 ± 5.22 | −15.60 ± 13.06 | 0.26 | Additive | ||
| Viable | Low | 1.08 ± 2.98 | 0.81 ± 2.91 | 0.5 | Additive | |
| ROS+ | Intermediate | −1.28 ± 4.66 | 2.14 ± 1.73 | 0.52 | Additive | |
| High | −5.18 ± 1.80 | 1.87 ± 1.86 * | 0.03 | Synergistic | ||
| (f) | ||||||
| Mixture | Examined Parameter | Concentration Combination | Expected Value | Measured Value | p-Value | Interaction |
| ZEN + DAS | Viable | Low | −6.99 ± 4.43 | −1.60 ± 2.78 | 0.34 | Additive |
| Intermediate | −4.28 ± 4.96 | 2.82 ± 4.00 | 0.31 | Additive | ||
| High | −4.04 ± 8.55 | 6.03 ± 5.73 | 0.36 | Additive | ||
| ΔΨm | Low | −1.57 ± 1.19 | −0.45 ± 0.12 | 0.39 | Additive | |
| Intermediate | −0.41 ± 0.63 | 0.22 ± 0.64 | 0.5 | Additive | ||
| High | −1.70 ± 0.98 | −0.75 ± 0.49 | 0.42 | Additive | ||
| Damaged | Low | 0.38 ± 0.73 | −0.72 ± 1.09 | 0.43 | Additive | |
| acrosome | Intermediate | 0.58 ± 0.71 | 0.37 ± 1.76 | 0.92 | Additive | |
| High | 3.10 ± 1.86 | 0.09 ± 1.49 | 0.25 | Additive | ||
| Intact | Low | 1.52 ± 0.47 | −1.18 ± 1.62 | 0.16 | Additive | |
| acrosome | Intermediate | −3.07 ± 1.95 | −1.77 ± 2.25 | 0.68 | Additive | |
| High | −7.47 ± 3.55 | −2.27 ± 3.55 | 0.34 | Additive | ||
| Viable | Low | −6.61 ± 7.06 | 4.62 ± 1.79 | 0.17 | Additive | |
| ROS- | Intermediate | −7.28 ± 10.03 | 6.17 ± 4.26 | 0.26 | Additive | |
| High | 2.54 ± 10.48 | 3.58 ± 4.72 | 0.93 | Additive | ||
| Viable | Low | 2.52 ± 3.69 | −3.86 ± 1.31 | 0.15 | Additive | |
| ROS+ | Intermediate | −0.28 ± 5.51 | −6.33 ± 4.48 | 0.43 | Additive | |
| High | −7.45 ± 3.51 | −1.63 ± 3.24 | 0.27 | Additive | ||
| Mixture | Examined Parameter | Expected Value | Measured Value | p-Value | Interaction |
|---|---|---|---|---|---|
| OTA + T2 + ZEN | Viable | −11.31 ± 5.92 | −0.83 ± 2.01 | 0.13 | Additive |
| ΔΨm | −0.05 ± 0.91 | −1.42 ± −3.54 | 0.22 | Additive | |
| Damaged acrosome | 1.43 ± 3.32 | −2.86 ± 0.87 | 0.25 | Additive | |
| Intact acrosome | −0.48 ± 3.96 | 3.17 ± 1.98 | 0.43 | Additive | |
| Viable ROS- | −9.14 ± 8.84 | −4.40 ± 8.11 | 0.7 | Additive | |
| Viable ROS+ | −1.83 ± 4.73 | 18.35 ± 9.01 | 0.08 | Additive | |
| DAS + T2 + OTA | Viable | −14.93 ± 5.64 | 1.87 ± 2.47 * | 0.02 | Synergistic |
| ΔΨm | −0.79 ± 1.10 | −0.97 ± 0.35 | 0.88 | Additive | |
| Damaged acrosome | 1.12 ± 3.23 | −3.47 ± 0.90 | 0.21 | Additive | |
| Intact acrosome | −0.27 ± 4.08 | 4.09 ± 0.81 | 0.33 | Additive | |
| Viable ROS- | −8.92 ± 11.28 | −2.48 ± 5.43 | 0.32 | Additive | |
| Viable ROS+ | −4.26 ± 3.63 | 2.16 ± 4.63 | 0.31 | Additive | |
| DAS + T2 + ZEN | Viable | −7.14 ± 6.70 | 5.97 ± 2.92 | 0.11 | Additive |
| ΔΨm | −1.07 ± 1.08 | −0.63 ± 0.69 | 0.74 | Additive | |
| Damaged acrosome | −1.29 ± 1.46 | 0.44 ± 0.80 | 0.97 | Additive | |
| Intact acrosome | −0.81 ± 2.09 | 0.61 ± 0.28 | 0.52 | Additive | |
| Viable ROS- | −11.19 ± 11.89 | −3.17 ± 9.75 | 0.62 | Additive | |
| Viable ROS+ | −2.64 ± 5.68 | 1.07 ± 4.45 | 0.69 | Additive |
| Mycotoxin | Concentration (ppm) | Concentration (µM) | Range of Admitted Limits in Animal Feed (ppm) |
|---|---|---|---|
| ZEN | 0.1 | 0.34 | 0.1–3 |
| 0.5 | 1.7 | ||
| 1 | 3.4 | ||
| 5 | 17 | ||
| 10 | 34 | ||
| OTA | 0.05 | 0.124 | 0.05–0.25 |
| 0.1 | 0.248 | ||
| 0.25 | 0.62 | ||
| 1 | 2.48 | ||
| 10 | 24.8 | ||
| T2 | 0.01 | 0.021 | 0.05–2 |
| 0.05 | 0.107 | ||
| 0.1 | 0.214 | ||
| 0.5 | 1.07 | ||
| DAS | 0.01 | 0.027 | 0.2 |
| 0.05 | 0.136 | ||
| 0.1 | 0.272 | ||
| 0.5 | 1.364 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kalo, D.; Mendelson, P.; Komsky-Elbaz, A.; Voet, H.; Roth, Z. The Effect of Mycotoxins and Their Mixtures on Bovine Spermatozoa Characteristics. Toxins 2023, 15, 556. https://doi.org/10.3390/toxins15090556
Kalo D, Mendelson P, Komsky-Elbaz A, Voet H, Roth Z. The Effect of Mycotoxins and Their Mixtures on Bovine Spermatozoa Characteristics. Toxins. 2023; 15(9):556. https://doi.org/10.3390/toxins15090556
Chicago/Turabian StyleKalo, Dorit, Paz Mendelson, Alisa Komsky-Elbaz, Hillary Voet, and Zvi Roth. 2023. "The Effect of Mycotoxins and Their Mixtures on Bovine Spermatozoa Characteristics" Toxins 15, no. 9: 556. https://doi.org/10.3390/toxins15090556
APA StyleKalo, D., Mendelson, P., Komsky-Elbaz, A., Voet, H., & Roth, Z. (2023). The Effect of Mycotoxins and Their Mixtures on Bovine Spermatozoa Characteristics. Toxins, 15(9), 556. https://doi.org/10.3390/toxins15090556