Diversity of Mycotoxins and Other Secondary Metabolites Recovered from Blood Oranges Infected by Colletotrichum, Alternaria, and Penicillium Species

 ,
,  and
and 
Abstract
1. Introduction
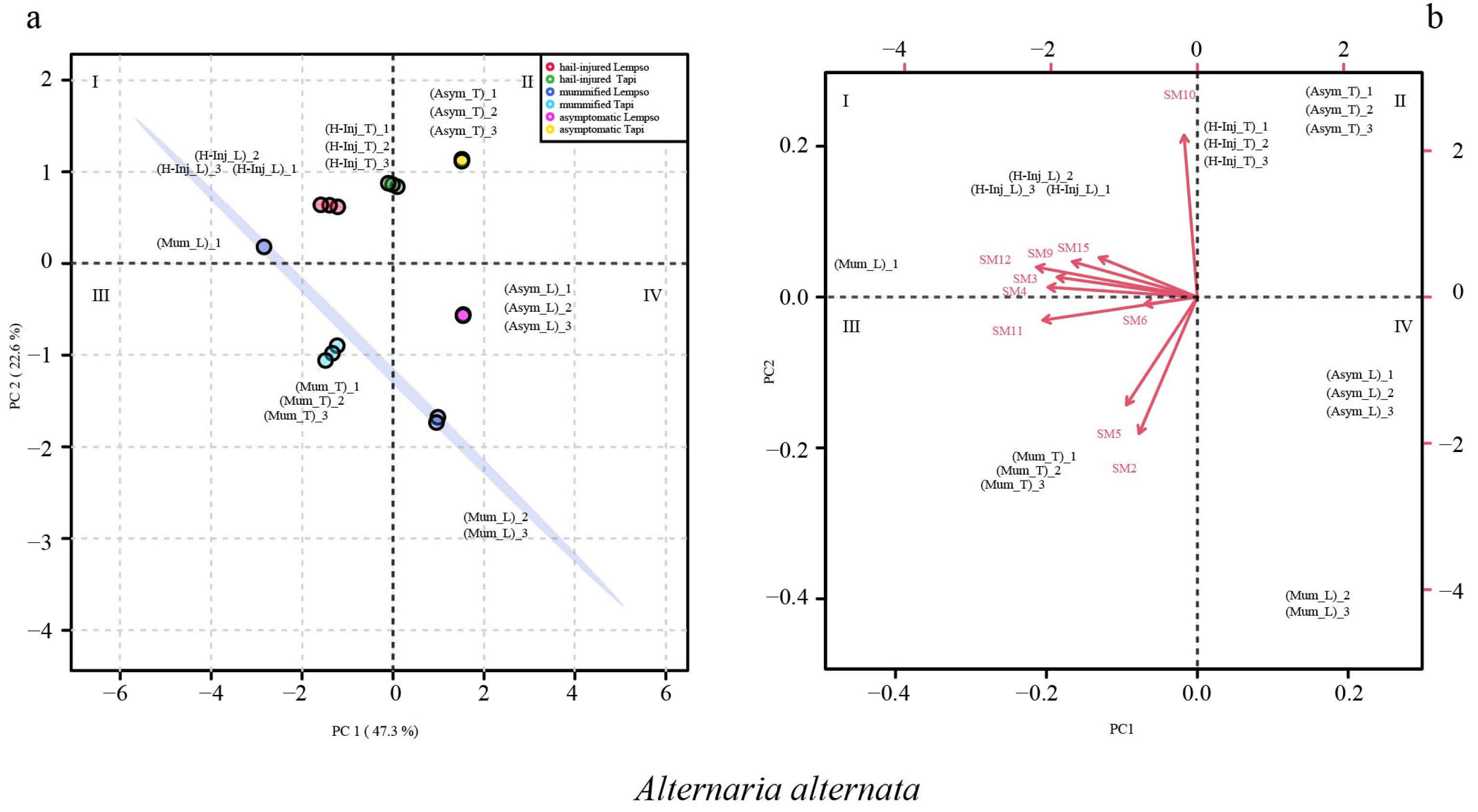
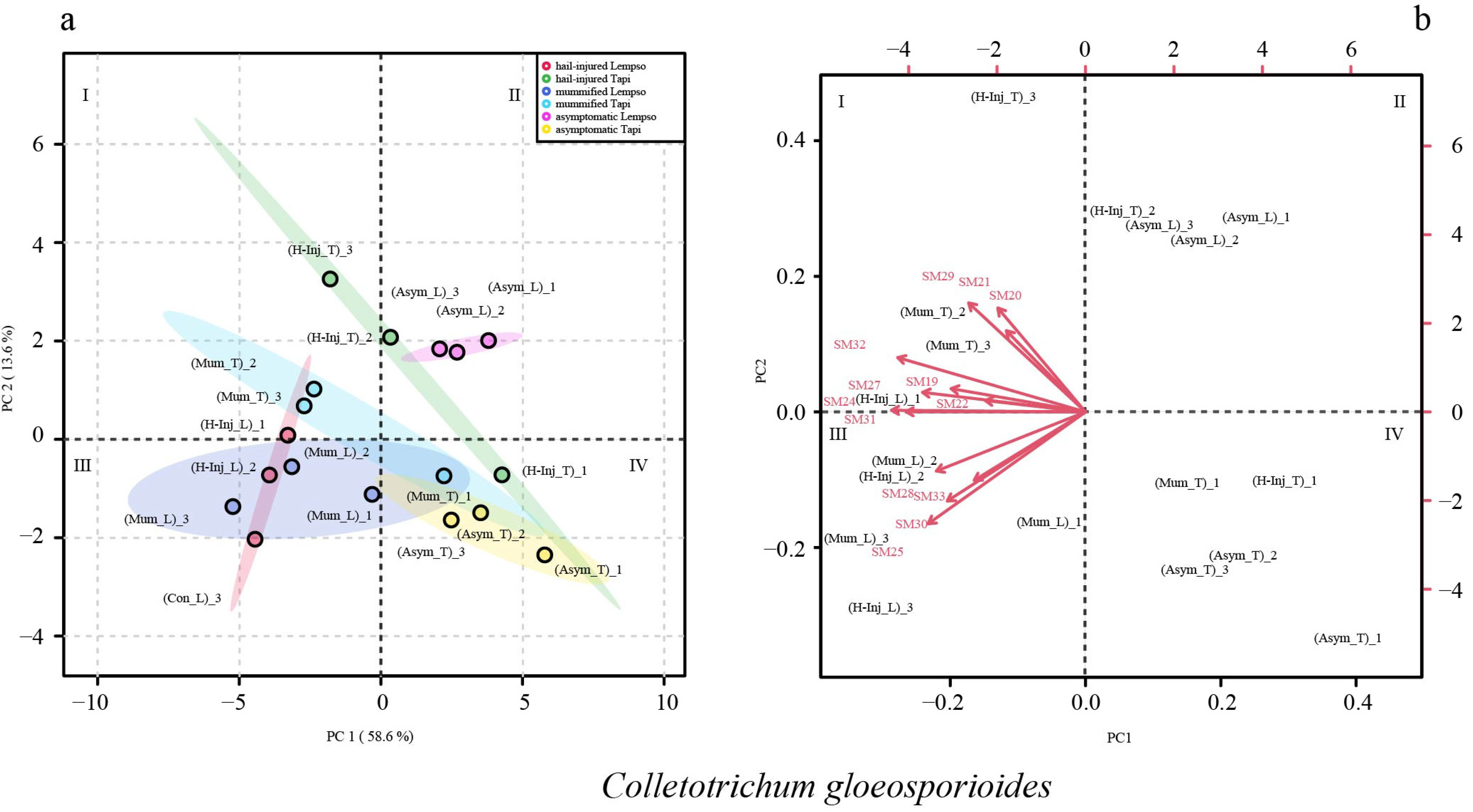
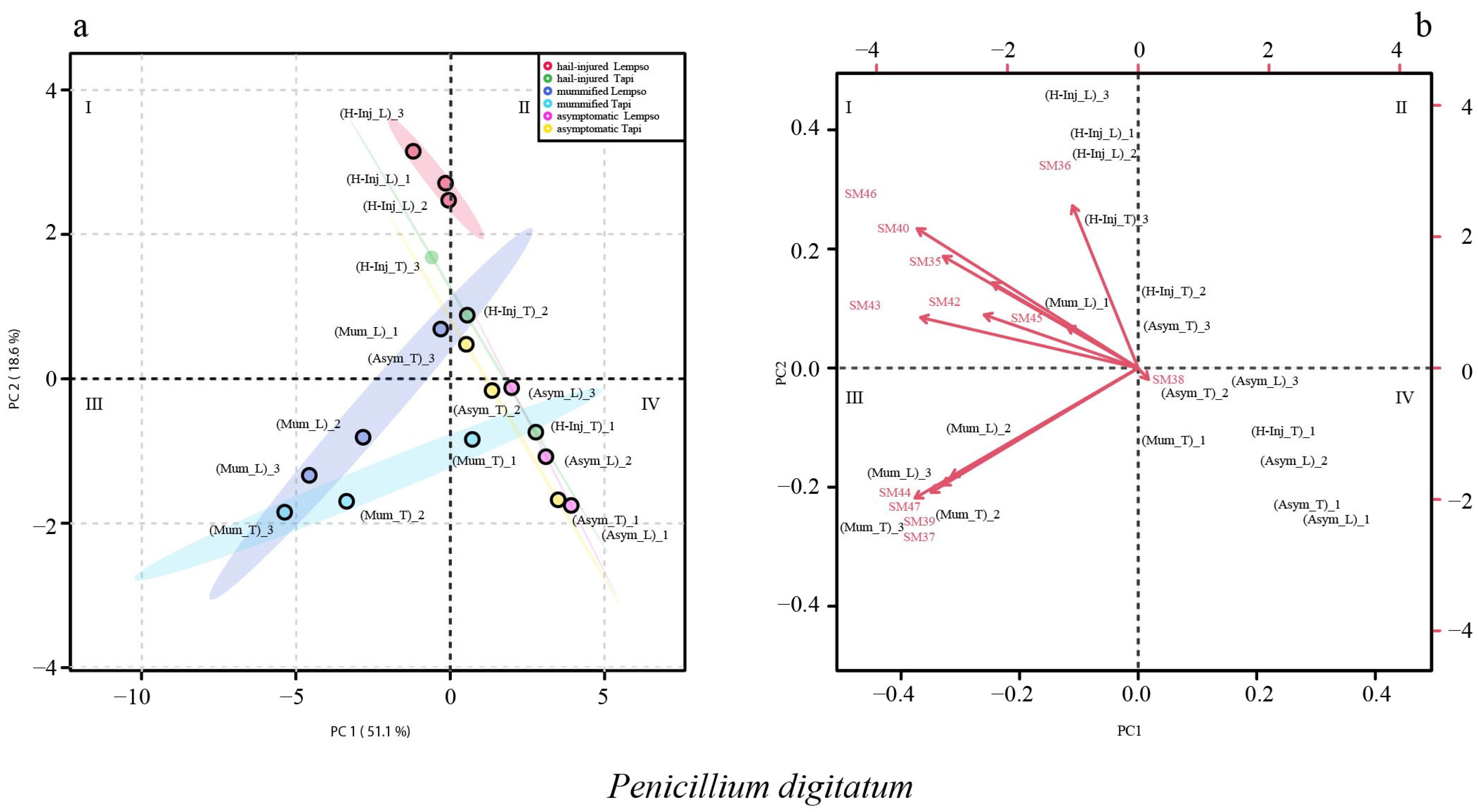
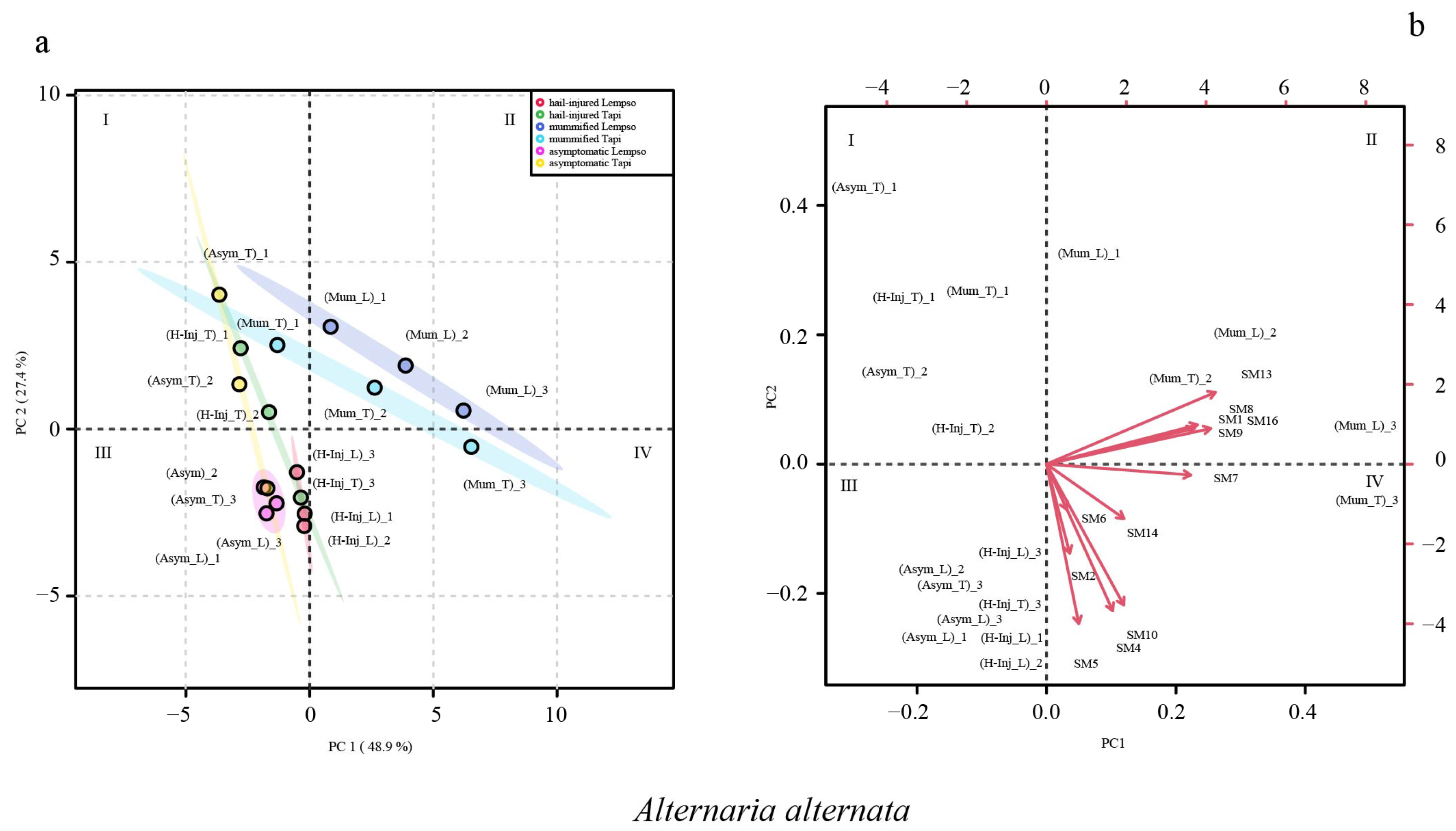
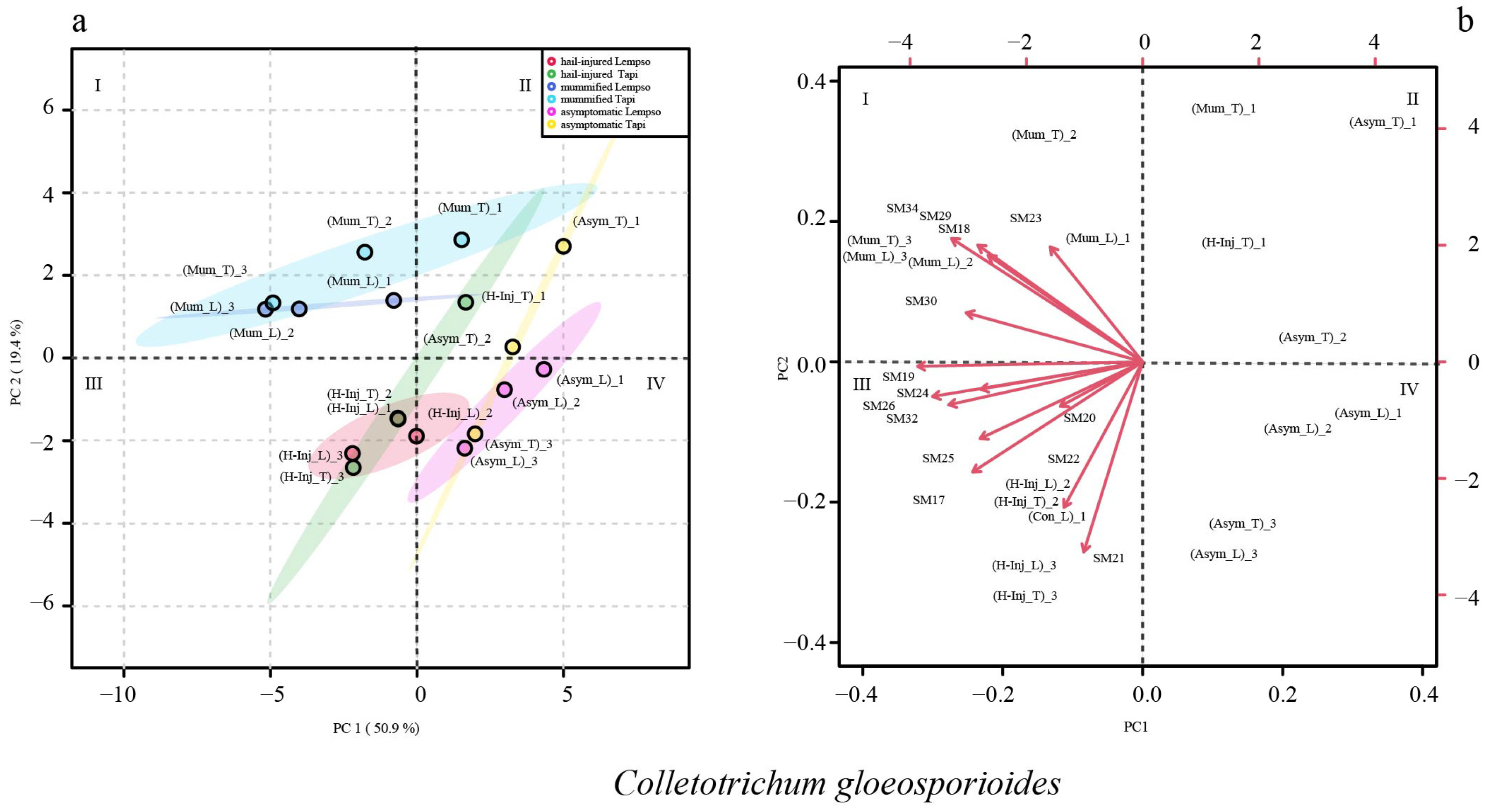
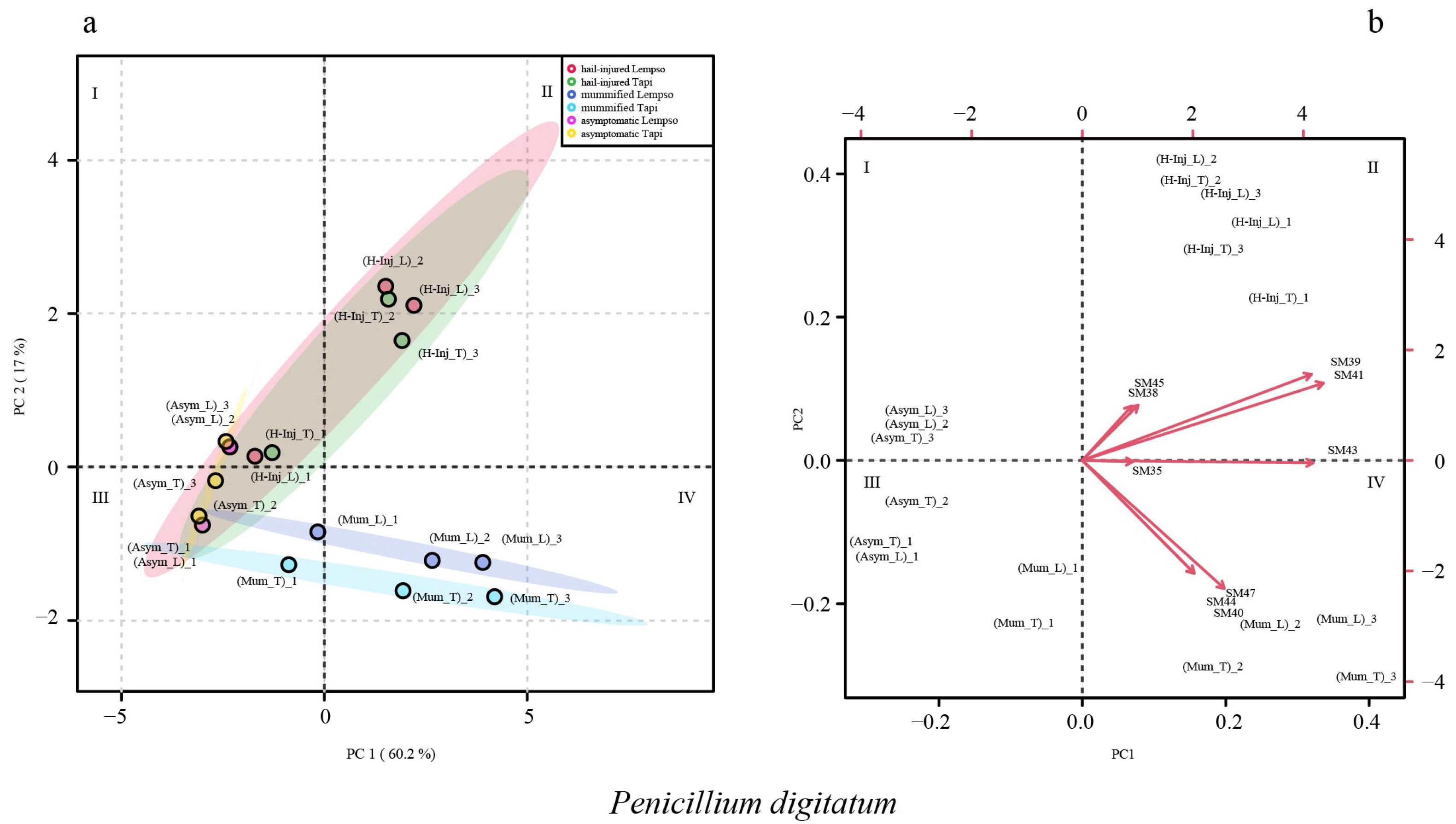
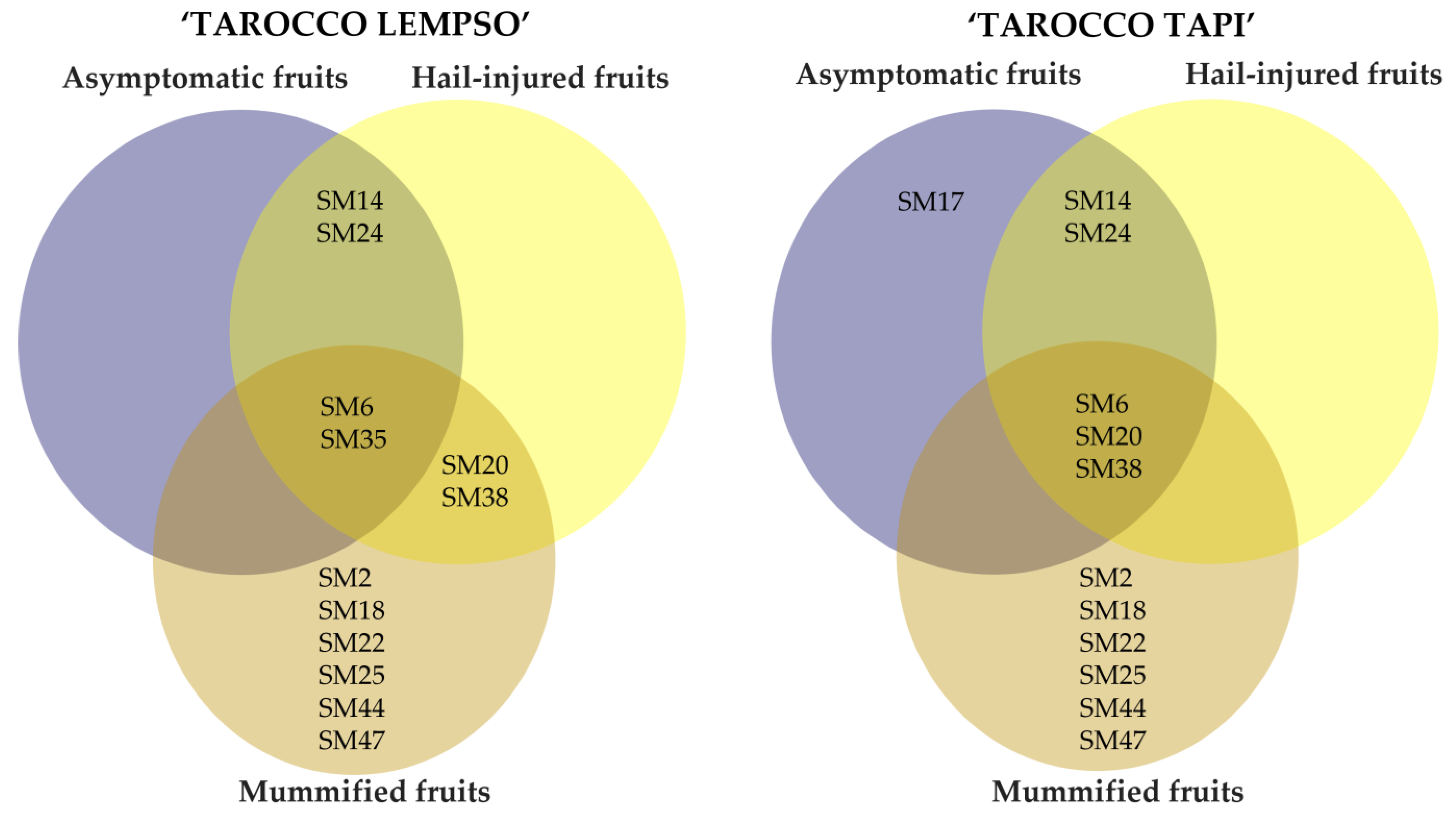
2. Results
3. Discussion
4. Conclusions
5. Materials and Methods
5.1. Orange Fruit Sampling
5.2. Isolation of Fungi
5.3. Morphological Characterization of Isolates
5.4. Molecular Characterization of Isolates
5.5. Preparation of Samples for the Analyses and Extraction of Orange Peel and Juice
5.6. Extraction of Fungal Secondary Metabolites from Orange Peel and Juice
5.7. Analysis of Fungal Secondary Metabolites by UHPLC–Q-TOF-MS
5.8. Statistical Analysis
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Ismail, M.; Zhang, J. Post-harvest citrus diseases and their control. Outlooks Pest Manag. 2004, 15, 29–35. [Google Scholar] [CrossRef]
- Wu, G.A.; Terol, J.; Ibanez, V.; López-García, A.; Pérez-Román, E.; Borredá, C.; Domingo, D.; Tadeo, F.R.; Carbonell-Caballero, J.; Alonso, R.; et al. Genomics of the origin and evolution of citrus. Nature 2018, 554, 311–316. [Google Scholar] [CrossRef] [PubMed]
- Aslam, K.; Iqbal, S.Z.; Razis, A.F.A.; Usman, S.; Ali, N.B. Patulin contamination of citrus fruits from Punjab and Northern Pakistan and estimation of associated dietary intake. Int. J. Environ. Res. Public Health 2021, 18, 2270. [Google Scholar] [CrossRef]
- FAO. Citrus Fruit Statistical Compendium Rome 2021. Available online: https://www.fao.org/publications/card/fr/c/CB6492EN/ (accessed on 8 June 2022).
- Li, L.; Lyall, G.K.; Martinez-Blazquez, J.A.; Vallejo, F.A.; Tomas-Barberan, F.; Birch, K.M.; Boesch, C. Blood orange iuice consumption increases flow-mediated dilation in adults with overweight and obesity: A randomized controlled trial. J. Nutr. 2020, 150, 2287–2294. [Google Scholar] [CrossRef]
- Abobatta, W.F. Nutritional benefits of citrus fruits. Am. J. Biomed. Sci. Res. 2019, 3, 303–306. [Google Scholar] [CrossRef]
- Kahramanoğlu, İ.; Nisar, M.F.; Chen, C.; Usanmaz, S.; Chen, J.; Wan, C. Light: An alternative method for physical control of postharvest rotting caused by fungi of citrus fruit. J. Food Qual. 2020, 2020, 8821346. [Google Scholar] [CrossRef]
- Grosso, G.; Galvano, F.; Mistretta, A.; Marventano, S.; Nolfo, F.; Calabrese, G.; Buscemi, S.; Drago, F.; Veronesi, U.; Scuderi, A. Red orange: Experimental models and epidemiological evidence of its benefits on human health. Oxid. Med. Cell Longev. 2013, 2013, 157240. [Google Scholar] [CrossRef]
- Giménez-Sanchis, A.; Zhong, K.; Pintor, A.; Farina, V.; Besada, C. Understanding blood versus blond orange consumption: A cross-cultural study in four countries. Foods 2022, 11, 2686. [Google Scholar] [CrossRef] [PubMed]
- Akimitsu, K.; Peever, T.L.; Timmer, L.W. Molecular, ecological and evolutionary approaches to understanding Alternaria diseases of citrus. Mol. Plant Pathol. 2003, 4, 435–446. [Google Scholar] [CrossRef] [PubMed]
- Ramos, A.P.; Talhinhas, P.; Sreenivasaprasad, S.; Oliveira, H. Characterization of Colletotrichum gloeosporioides, as the main causal agent of citrus anthracnose, and C. karstii as species preferentially associated with lemon twig dieback in Portugal. Phytoparasitica 2016, 44, 549–561. [Google Scholar] [CrossRef]
- El boumlasy, S.; La Spada, F.; Tuccitto, N.; Marletta, G.; Mínguez, C.L.; Meca, G.; Rovetto, E.I.; Pane, A.; Debdoubi, A.; Cacciola, S.O. Inhibitory activity of shrimp waste extracts on fungal and oomycete plant pathogens. Plants 2021, 10, 2452. [Google Scholar] [CrossRef] [PubMed]
- Strano, M.C.; Altieri, G.; Allegra, M.; Di Renzo, G.C.; Paterna, G.; Matera, A.; Genovese, F. Postharvest technologies of fresh citrus fruit: Advances and recent developments for the loss reduction during handling and storage. Horticulturae 2022, 8, 612. [Google Scholar] [CrossRef]
- Palou, L.; Usall, J.; Smilanick, J.L.; Aguilar, M.-J.; Viñas, I. Evaluation of food additives and low-toxicity compounds as alternative chemicals for the control of Penicillium digitatum and Penicillium italicum on citrus fruit. Pest Manag. Sci. 2002, 58, 459–466. [Google Scholar] [CrossRef]
- Qi, J.; Pang, Y.; An, P.; Jiang, G.; Kong, Q.; Ren, X. Determination of metabolites of Geotrichum citri-aurantii treated with peppermint oil using liquid chromatography-mass spectrometry and gas chromatography-mass spectrometry. J. Food Biochem. 2019, 43, e12745. [Google Scholar] [CrossRef]
- Huang, F.; Chen, G.Q.; Hou, X.; Fu, Y.S.; Cai, L.; Hyde, K.D.; Li, H.Y. Colletotrichum species associated with cultivated citrus in China. Fungal Divers. 2013, 61, 61–74. [Google Scholar] [CrossRef]
- Riolo, M.; Aloi, F.; Pane, A.; Cara, M.; Cacciola, S.O. Twig and shoot dieback of citrus, a new disease caused by Colletotrichum species. Cells 2021, 10, 449. [Google Scholar] [CrossRef]
- Wang, W.; de Silva, D.D.; Moslemi, A.; Edwards, J.; Ades, P.K.; Crous, P.W.; Taylor, P.W.J. Colletotrichum species causing anthracnose of citrus in Australia. J. Fungi 2021, 7, 47. [Google Scholar] [CrossRef]
- Lopes da Silva, L.; Alvarado Moreno, H.L.; Nunes Correia, H.L.; Ferreira Santana, M.; Vieira de Queiroz, M. Colletotrichum: Species complexes, lifestyle, and peculiarities of some sources of genetic variability. Appl. Microbiol. Biot. 2020, 104, 1891–1904. [Google Scholar] [CrossRef]
- Woudenberg, J.H.C.; Seidl, M.F.; Groenewald, J.Z.; de Vries, M.; Stielow, J.B.; Thomma, B.P.H.J.; Crous, P.W. Alternaria section Alternaria: Species, formae speciales or pathotypes? Stud. Mycol. 2015, 82, 1–21. [Google Scholar] [CrossRef] [PubMed]
- Lawrence, D.P.; Rotondo, F.; Gannibal, P.B. Biodiversity and taxonomy of the pleomorphic genus Alternaria. Mycol. Prog. 2016, 15, 3. [Google Scholar] [CrossRef]
- Patriarca, A. Alternaria in food products. Curr. Opin. Food Sci. 2016, 11, 1–9. [Google Scholar] [CrossRef]
- Timmer, L.V.; Peever, T.L.; Solel, Z.; Akimitzu, K. Alternaria diseases of citrus—Novel pathosystems. Phytopathol. Medit. 2003, 42, 99–112. [Google Scholar]
- Garganese, F.; Schena, L.; Siciliano, I.; Prigigallo, M.I.; Spadaro, D.; De Grassi, A.; Ippolito, A.; Sanzani, S.M. Characterization of citrus-associated Alternaria species in Mediterranean areas. PLoS ONE 2016, 16, e0163255. [Google Scholar] [CrossRef]
- Garganese, F.; Ippolito, A.; di Rienzo, V.; Lotti, C.; Montemurro, C.; Sanzani, S.M. A new high-resolution melting assay for genotyping Alternaria species causing citrus brown spot. J. Sci. Food Agric. 2018, 98, 4578–4583. [Google Scholar] [CrossRef]
- Peever, T.L.; Su, G.; Carpenter-Boggs, L.; Timmer, L.W. Molecular systematics of citrus-associated Alternaria species. Mycologia 2004, 96, 119–134. [Google Scholar] [CrossRef]
- Peever, T.L.; Carpenter-Boggs, L.; Timmer, L.W.; Carris, L.M.; Bhatia, A. Citrus black rot is caused by phylogenetically distinct lineages of Alternaria alternata. Phytopathology 2005, 95, 512–518. [Google Scholar] [CrossRef] [PubMed]
- Tsitsigiannis, D.I.; Dimakopoulou, M.; Antoniou, P.P.; Tjamos, E.C. Biological control strategies of mycotoxigenic fungi and associated mycotoxins in Mediterranean basin crops. Phytopathol. Medit. 2012, 51, 158–174. [Google Scholar]
- Bartholomew, H.P.; Bradshaw, M.; Jurick, W.M.; Fonseca, J.M. The good, the bad, and the ugly: Mycotoxin production during postharvest decay and their influence on tritrophic host–pathogen–microbe interactions. Front. Microbiol. 2021, 12, 611881. [Google Scholar] [CrossRef] [PubMed]
- Leslie, J.F.; Moretti, A.; Mesterházy, Á.; Ameye, M.; Audenaert, K.; Singh, P.K.; Richard-Forget, F.; Chulze, S.N.; Ponte, E.M.D.; Chala, A.; et al. Key global actions for mycotoxin management in wheat and other small grains. Toxins 2021, 13, 725. [Google Scholar] [CrossRef] [PubMed]
- Nan, M.; Xue, H.; Bi, Y. Contamination, detection and control of mycotoxins in fruits and vegetables. Toxins 2022, 14, 309. [Google Scholar] [CrossRef]
- Wang, B.; Nong, X.H.; Zeng, W.N.; Li, S.S.; Li, G.Y.; Liu, J.; Huang, G.L.; Zheng, C.J.; Zheng, C.J. Study on bioactive secondary metabolites from the mangrove-derived fungus Penicillium verruculosum TGM14. Chem. Nat. Compd. 2022, 58, 812–815. [Google Scholar] [CrossRef]
- Schiff, P.L., Jr. Ergot and its alkaloids. Am. J. Pharm. Educ. 2006, 70, 98. [Google Scholar] [CrossRef]
- Aloi, F.; Riolo, M.; Sanzani, S.M.; Mincuzzi, A.; Ippolito, A.; Siciliano, I.; Pane, A.; Gullino, M.L.; Cacciola, S.O. Characterization of Alternaria species associated with heart rot of pomegranate fruit. J. Fungi 2021, 7, 172. [Google Scholar] [CrossRef]
- Bentivenga, G.; Spina, A.; Ammar, K.; Allegra, M.; Cacciola, S.O. Screening of durum wheat (Triticum turgidum L. subsp. durum (Desf.) Husn.) Italian cultivars for susceptibility to Fusarium Head Blight incited by Fusarium graminearum. Plants 2021, 10, 68. [Google Scholar] [CrossRef] [PubMed]
- Stracquadanio, C.; Luz, C.; La Spada, F.; Meca, G.; Cacciola, S.O. Inhibition of mycotoxigenic fungi in different vegetable matrices by extracts of Trichoderma species. J. Fungi 2021, 7, 445. [Google Scholar] [CrossRef]
- Magan, N.; Olsen, M. (Eds.) Mycotoxins in Food: Detection and Control; Woodhead Publishing Ltd.: Cambridge, UK, 2004; p. 471. [Google Scholar]
- Fernández-Cruz, M.L.; Mansilla, M.L.; Tadeo, J.L. Mycotoxins in fruits and their processed products: Analysis, occurrence and health implications. J. Adv. Res. 2010, 1, 113–122. [Google Scholar] [CrossRef]
- Barkai-Golan, R.; Paster, N. (Eds.) Mycotoxins in Fruits and Vegetables, 1st ed.; Academic Press: San Diego, CA, USA, 2008; p. 408. [Google Scholar]
- Pallarés, N.; Carballo, D.; Ferrer, E.; Fernández-Franzón, M.; Berrada, H. Mycotoxin dietary exposure assessment through fruit juices consumption in children and adult population. Toxins 2019, 11, 684. [Google Scholar] [CrossRef] [PubMed]
- Carballo, D.; Pinheiro-Fernandes-Vieira, P.; Font, G.; Berrada, H.; Ferrer, E. Dietary exposure to mycotoxins through fruits juice consumption. Rev. Toxicol. 2018, 35, 2–6. [Google Scholar]
- Spadaro, D.; Garibaldi, A.; Gullino, M.L. Occurrence of patulin and its dietary intake through pear, peach, and apricot juices in Italy. Food Addit. Contam. 2008, 1, 134–139. [Google Scholar] [CrossRef]
- Marino, A.; Nostro, A.; Fiorentino, C. Ochratoxin A production by Aspergillus westerdijkiae in orange fruit and juice. Int. J. Food Microbiol. 2009, 132, 185–189. [Google Scholar] [CrossRef] [PubMed]
- Zouaoui, N.; Sbaii, N.; Bacha, H.; Abid-Essefi, S. Occurrence of patulin in various fruit juice marketed in Tunisia. Food Control 2015, 51, 356–360. [Google Scholar] [CrossRef]
- Juan, C.; Mañes, J.; Font, G.; Juan-García, A. Determination of mycotoxins in fruit berry by-products using QuEChERS extraction method. LWT-Food Sci. Technol. 2017, 86, 344–351. [Google Scholar] [CrossRef]
- Li, X.; Li, H.; Ma, W.; Guo, Z.; Li, X.; Li, X.; Zhang, Q. Determination of patulin in apple juice by single-drop liquid-liquid-liquid microextraction coupled with liquid chromatography-mass spectrometry. Food Chem. 2018, 257, 1–6. [Google Scholar] [CrossRef] [PubMed]
- Groopman, J.D.; Kensler, T.W. Food safety: Mycotoxins. In Encyclopedia of Human Nutrition, 2nd ed.; Caballero, B., Allen, L., Prentice, A., Eds.; Academic Press: Cambridge, MA, USA, 2005; pp. 317–323. [Google Scholar]
- Groopman, J.D.; Kensler, T.W.; Wild, C.P. Carcinogenesis in developing countries. Annu. Rev. Publ. Health 2008, 29, 187–203. [Google Scholar] [CrossRef]
- Wu, F.; Groopman, J.D.; Pestka, J.J. Public health impacts of foodborne mycotoxins. Annu. Rev. Food Sci. Technol. 2014, 5, 351–372. [Google Scholar] [CrossRef]
- EUR-Lex. Commission Regulation (EC) No 669/2009 of 24 July 2009 implementing Regulation (EC) No 882/2004 of the European Parliament and of the Council as Regards the Increased Level of Official Controls on Imports of Certain Feed and Food of Non-Animal Origin and Amending Decision 2006/504/EC (Text with EEA Relevance). Available online: http://data.europa.eu/eli/reg/2009/669/oj (accessed on 7 May 2023).
- Van de Perre, E.; Jacxsens, L.; Lachat, C.; El Tahan, F.; De Meulenaer, B. Impact of maximum levels in European legislation on exposure of mycotoxins in dried products: Case of aflatoxin B1 and ochratoxin A in nuts and dried fruits. Food Chem. Toxicol. 2015, 75, 112–117. [Google Scholar] [CrossRef] [PubMed]
- Wang, M.; Jiang, N.; Xian, H.; Wei, D.; Shi, L.; Feng, X. A single-step solid phase extraction for the simultaneous determination of 8 mycotoxins in fruits by ultra-high performance liquid chromatography tandem mass spectrometry. J. Chromat. A 2016, 1429, 22–29. [Google Scholar] [CrossRef]
- EUR-Lex. Commission Regulation (EC) No 1881/2006 of 19 December 2006 Setting Maximum Levels for Certain Contaminants in Foodstuffs (Text with EEA Relevance). Available online: http://data.europa.eu/eli/reg/2006/1881/oj (accessed on 7 May 2023).
- Hussain, S.; Asi, M.R.; Iqbal, M.; Khalid, N.; Wajih-ul-Hassan, S.; Ariño, A. Patulin mycotoxin in mango and orange fruits, juices, pulps, and jams marketed in Pakistan. Toxins 2020, 12, 52. [Google Scholar] [CrossRef] [PubMed]
- EUR-Lex. Commission Regulation (EU) No 165/2010 of 26 February 2010 Amending Regulation (EC) No 1881/2006 Setting Maximum Levels for Certain Contaminants in Foodstuffs as Regards aflatoxins (Text with EEA Relevance). Available online: http://data.europa.eu/eli/reg/2010/165/oj (accessed on 7 May 2023).
- Boudergue, C.; Burel, C.; Dragacci, S.; Favrot, M.C.; Fremy, J.M.; Massimi, C.; Prigent, P.; Debongnie, P.; Pussemier, L.; Boudra, H.; et al. Review of mycotoxin-detoxifying agents used as feed additives: Mode of action, efficacy and feed/food safety. EFSA Support. Publ. 2009, 6, 22E. [Google Scholar] [CrossRef]
- EFSA Panel on Contaminants in the Food Chain (CONTAM); Schrenk, D.; Bodin, L.; Chipman, J.K.; del Mazo, J.; Grasl-Kraupp, B.; Hogstrand, C.; Hoogenboom, L.; Leblanc, J.-C.; Nielsen, E.; et al. Risk assessment of ochratoxin A in food. EFSA J. 2020, 18, e06113. [Google Scholar]
- Escrivá, L.; Oueslati, S.; Font, G.; Manyes, L. Alternaria mycotoxins in food and feed: An overview. J. Food Qual. 2017, 2017, 1569748. [Google Scholar] [CrossRef]
- Vaclavikova, M.; Dzuman, Z.; Lacina, O.; Fenclova, M.; Veprikova, Z.; Zachariasova, M.; Hajslova, J. Monitoring survey of patulin in a variety of fruit-based products using a sensitive UHPLC–MS/MS analytical procedure. Food Control 2015, 47, 577–584. [Google Scholar] [CrossRef]
- Murillo-Arbizu, M.; Amézqueta, S.; González-Peñas, E.; de Cerain, A.L. Occurrence of patulin and its dietary intake through apple juice consumption by the Spanish population. Food Chem. 2009, 113, 420–423. [Google Scholar] [CrossRef]
- Ruan, C.; Diao, X.; Zhang, H.; Zhang, L.; Liu, C. Development of a dispersive liquid–liquid microextraction technique for the analysis of citrinin, alternariol and alternariol monomethyl ether in fruit juices. Anal. Methods 2016, 8, 7944–7950. [Google Scholar] [CrossRef]
- Víctor-Ortega, M.D.; Lara, F.J.; García-Campaña, A.M.; del Olmo-Iruela, M. Evaluation of dispersive liquid–liquid microextraction for the determination of patulin in apple juices using micellar electrokinetic capillary chromatography. Food Control 2013, 31, 353–358. [Google Scholar] [CrossRef]
- Zgoła-Grześkowiak, A.; Grześkowiak, T. Dispersive liquid-liquid microextraction. TrAC Trends Anal. Chem. 2011, 30, 1382–1399. [Google Scholar] [CrossRef]
- Pallares, N.; Font, G.; Manes, J.; Ferrer, E. Multimycotoxin LC–MS/MS analysis in tea beverages after dispersive liquid–liquid microextraction (DLLME). J. Agric. Food Chem. 2017, 65, 10282–10289. [Google Scholar] [CrossRef] [PubMed]
- Antep, H.M.; Merdivan, M. Development of new dispersive liquid−liquid microextraction technique for the identification of zearalenone in beer. Anal. Methods 2012, 4, 4129–4134. [Google Scholar] [CrossRef]
- Tolosa, J.; Font, G.; Mañes, J.; Ferrer, E. Multimycotoxin analysis in water and fish plasma by liquid chromatography-tandem mass spectrometry. Chemosphere 2016, 145, 402–408. [Google Scholar] [CrossRef] [PubMed]
- Rodríguez-Carrasco, Y.; Mañes, J.; Berrada, H.; Juan, C. Development and validation of a LC-ESI-MS/MS method for the determination of Alternaria toxins alternariol, alternariol methylether and tentoxin in tomato and tomato-based products. Toxins 2016, 8, 328. [Google Scholar] [CrossRef] [PubMed]
- Serrano, A.B.; Font, G.; Mañes, J.; Ferrer, E. Development a mitigation strategy of enniatins in pasta under home-cooking conditions. LWT-Food Sci. Technol. 2016, 65, 1017–1024. [Google Scholar] [CrossRef]
- Gilbert, J.; Anklam, E. Validation of analytical methods for determining mycotoxins in foodstuffs. TrAC Trends Anal. Chem. 2002, 21, 468–486. [Google Scholar] [CrossRef]
- Shi, H.; Li, S.; Bai, Y.; Prates, L.L.; Lei, Y.; Yu, P. Mycotoxin contamination of food and feed in China: Occurrence, detection techniques, toxicological effects and advances in mitigation technologies. Food Control 2018, 91, 202–215. [Google Scholar] [CrossRef]
- Alshannaq, A.; Yu, J.H. Occurrence, toxicity, and analysis of major mycotoxins in food. Int. J. Environ. Res. Public Health 2017, 14, 632. [Google Scholar] [CrossRef] [PubMed]
- Biselli, S.; Hummert, C. Development of a multicomponent method for Fusarium toxins using LC–MS/MS and its application during a survey for the content of T-2 toxin and deoxynivalenol in various feed and food samples. Food Addit. Contam. 2005, 22, 752–760. [Google Scholar] [CrossRef]
- Krska, R.; Welzig, E.; Boudra, H. Analysis of Fusarium toxins in feed. Anim. Feed Sci. Technol. 2007, 137, 241–264. [Google Scholar] [CrossRef]
- Martins, H.M.; Mendes Guerra, M.M.; d’Almeida Bernardo, F.M. Occurrence of aflatoxin B1 in dairy cow feed over 10 years in Portugal (1995–2004). Rev. Iberoam. Micol. 2007, 24, 69–71. [Google Scholar] [CrossRef] [PubMed]
- Vrabcheva, T.; Usleber, E.; Dietrich, R.; Martlbauer, E. Co-occurrence of ochratoxin A and citrinin in cereals from Bulgarian villages with a history of Balkan endemic nephropathy. J. Agric. Food Chem. 2000, 48, 2483–2488. [Google Scholar] [CrossRef]
- Delmulle, B.; De Saeger, S.; Adams, A.; De Kimpe, N.; Van Peteghem, C. Development of a liquid chromatography/tandem mass spectrometry method for the simultaneous determination of 16 mycotoxins on cellulose filters and in fungal cultures. Rapid Commun. Mass Spectrom. 2006, 20, 771–776. [Google Scholar] [CrossRef] [PubMed]
- Kokkonen, M.; Jestoi, M.; Rizzo, A. Determination of selected mycotoxins in mould cheeses with liquid chromatography coupled to tandem with mass spectrometry. Food Addit. Contam. 2005, 22, 449–456. [Google Scholar] [CrossRef]
- Sorensen, L.K.; Elbaek, T.H. Determination of mycotoxins in bovine milk by liquid chromatography tandem mass spectrometry. J. Chromatogr. B Analyt. Technol. Biomed. Life Sci. 2005, 820, 183–196. [Google Scholar] [CrossRef] [PubMed]
- Garcia-Villanova, R.J.; Cordon, C.; Gonzalez Paramas, A.M.; Aparicio, P.; Garcia Rosales, M.E. Simultaneous immunoaffinity column clean-up and HPLC analysis of aflatoxins and ochratoxin A in Spanish bee pollen. J. Agric. Food. Chem. 2004, 52, 7235–7239. [Google Scholar] [CrossRef]
- Aresta, A.; Cioffi, N.; Palmisano, F.; Zambonin, C.G. Simultaneous determination of ochratoxin A and cyclopiazonic, mycophenolic, and tenuazonic acids in cornflakes by solidphase microextraction coupled to high-performance liquid chromatography. J. Agric. Food Chem. 2003, 51, 5232–5237. [Google Scholar] [CrossRef] [PubMed]
- Chan, D.; MacDonald, S.J.; Boughtflower, V.; Brereton, P. Simultaneous determination of aflatoxins and ochratoxin A in food using a fully automated immunoaffinity column cleanup and liquid chromatography-fluorescence detection. J. Chromatogr. A 2004, 1059, 13–16. [Google Scholar] [CrossRef]
- Rahmani, A.; Jinap, S.; Soleimany, F. Qualitative and quantitative analysis of mycotoxins. Compr. Rev. Food Sci. Food Saf. 2009, 8, 202–251. [Google Scholar] [CrossRef] [PubMed]
- Soleas, G.J.; Yan, J.; Goldberg, D.M. Assay of ochratoxin A in wine and beer by highpressure liquid chromatography photodiode array and gas chromatography mass selective detection. J. Agric. Food Chem. 2001, 49, 2733–2740. [Google Scholar] [CrossRef]
- He, Y.; Li, Z.; Wang, W.; Sooranna, S.R.; Shi, Y.; Chen, Y.; Wu, C.; Zeng, J.; Tang, Q.; Xie, H. Chemical profiles and simultaneous quantification of Aurantii fructus by use of HPLC-Q-TOF-MS combined with GC-MS and HPLC methods. Molecules 2018, 23, 2189. [Google Scholar] [CrossRef] [PubMed]
- Dopazo, V.; Luz, C.; Mañes, J.; Quiles, J.M.; Carbonell, R.; Calpe, J.; Meca, G. Bio-preservative potential of microorganisms isolated from red grape against food contaminant fungi. Toxins 2021, 13, 412. [Google Scholar] [CrossRef] [PubMed]
- Thurman, E.M.; Ferrer, I.; Fernández-Alba, A.R. Matching unknown empirical formulas to chemical structure using LC/MS TOF accurate mass and database searching: Example of unknown pesticides on tomato skins. J. Chromat. A 2005, 1067, 127–134. [Google Scholar] [CrossRef]
- Xing, T.T.; Zhao, X.J.; Zhang, Y.D.; Li, Y.F. Fast separation and sensitive quantitation of polymethoxylated flavonoids in the peels of citrus using UPLC-Q-TOF-MS. J. Agric. Food Chem. 2017, 65, 2615–2627. [Google Scholar] [CrossRef]
- Wang, H.; Guo, Y.; Luo, Z.; Gao, L.; Li, R.; Zhang, Y.; Kalaji, H.M.; Qiang, S.; Chen, S. Recent advances in Alternaria phytotoxins: A review of their occurrence, structure, bioactivity, and biosynthesis. J. Fungi 2022, 8, 168. [Google Scholar] [CrossRef]
- Nakatsuka, S.I.; Ueda, K.; Goto, T.; Yamamoto, M.; Nishimura, S.; Kohmoto, K. Structure of AF-toxin II, one of the host-specific toxins produced by Alternaria alternata strawberry pathotype. Tetrahedron Lett. 1986, 27, 2753–2756. [Google Scholar] [CrossRef]
- Bensassi, F.; Gallerne, C.; Hajlaoui, M.R.; Bacha, H.; Lemaire, C. Mechanism of Alternariol monomethyl ether-induced mitochondrial apoptosis in human colon carcinoma cells. Toxicology 2011, 290, 230–240. [Google Scholar] [CrossRef]
- Berestetskiy, A.O.; Dalinova, A.A.; Volosatova, N.S. Metabolite profiles and biological activity of extracts from Alternaria sonchi S-102 culture grown by different fermentation methods. Appl. Biochem. Microbiol. 2019, 55, 284–293. [Google Scholar] [CrossRef]
- Debbab, A.; Aly, A.H.; Edrada-Ebel, R.; Wray, V.; Müller, W.E.; Totzke, F.; Zirrgiebel, U.; Schäctele, C.; Kubbutatt, M.H.G.; Lin, W.H.; et al. Bioactive metabolites from the endophytic fungus Stemphylium globuliferum isolated from Mentha pulegium. J. Nat. Prod. 2009, 72, 626–631. [Google Scholar] [CrossRef]
- Shaaban, M.; Shaaban, K.A.; Abdel-Aziz, M.S. Seven naphtho-γ-pyrones from the marine-derived fungus Alternaria alternata: Structure elucidation and biological properties. Org. Med. Chem. Lett. 2012, 2, 6. [Google Scholar] [CrossRef] [PubMed]
- Meena, M.; Samal, S. Alternaria host-specific (HSTs) toxins: An overview of chemical characterization, target sites, regulation and their toxic effects. Toxicol. Rep. 2019, 6, 745–758. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Palaniveloo, K.; Alias, S.A.; Sathiya Seelan, J.S. Species diversity and secondary metabolites of Sarcophyton-associated marine fungi. Molecules 2021, 26, 3227. [Google Scholar] [CrossRef]
- Yang, Y.-S.; Johnson, D.R.; Dowler, W.M. Pathogenicity of Alternaria angustiovoidea on leafy spurge. Plant Dis. 1990, 74, 601–604. [Google Scholar] [CrossRef]
- Elkhateeb, W.A.; Kolaibe, A.G.A.; Elkhateeb, A.; Daba, G.M. Allergen, pathogen, or biotechnological tool? The dematiaceous fungi Alternaria what’s for it and what’s on it. J. Pharm. Pharmacol. Res. 2021, 4, 1–6. [Google Scholar] [CrossRef] [PubMed]
- Suemitsu, R.; Ohnishi, K.; Morikawa, Y.; Ideguchi, I.; Uno, H. Porritoxinol, a phytotoxin of Alternaria porri. Phytochemistry 1994, 35, 603–605. [Google Scholar] [CrossRef]
- Jiménez-Teja, D.; Daoubi, M.; Collado, I.G.; Hernández-Galán, R. Lipase-catalyzed resolution of 5-acetoxy-1, 2-dihydroxy-1, 2, 3, 4-tetrahydronaphthalene. Application to the synthesis of (+)-(3R, 4S)-cis-4-hydroxy-6-deoxyscytalone, a metabolite isolated from Colletotrichum acutatum. Tetrahedron 2009, 65, 3392–3396. [Google Scholar] [CrossRef]
- Yang, Z.D.; Li, Z.J.; Zhao, J.W.; Sun, J.H.; Yang, L.J.; Shu, Z.M. Secondary metabolites and PI3K inhibitory activity of Colletotrichum gloeosporioides, a fungal endophyte of Uncaria rhynchophylla. Curr. Microbiol. 2019, 76, 904–908. [Google Scholar] [CrossRef]
- Chakraborty, A.; Ray, P. Mycoherbicides for the noxious meddlesome: Can Colletotrichum be a budding candidate? Front. Microbiol. 2021, 12, 754048. [Google Scholar] [CrossRef] [PubMed]
- Moraga, J.; Gomes, W.; Pinedo, C.; Cantoral, J.M.; Hanson, J.R.; Carbú, M.; Garrido, C.; Duran-Patron, R.; Collado, I.G. The current status on secondary metabolites produced by plant pathogenic Colletotrichum species. Phytochem. Rev. 2019, 18, 215–239. [Google Scholar] [CrossRef]
- Kim, J.W.; Shim, S.H. The fungus Colletotrichum as a source for bioactive secondary metabolites. Arch. Pharm. Res. 2019, 42, 735–753. [Google Scholar] [CrossRef] [PubMed]
- Gohbara, M.; Kosuge, Y.; Yamasaki, S.; Kimura, Y.; Suzuki, A.; Tamura, S. Isolation, structures and biological activities of colletotrichins, phytotoxic substances from Colletotrichum nicotianae. Agric. Biol. Chem. Tokyo 1978, 42, 1037–1043. [Google Scholar] [CrossRef]
- Inoue, M.; Takenaka, H.; Tsurushima, T.; Miyagawa, H.; Ueno, T. Colletofragarones A1 and A2, novel germination self-inhibitors from the fungus Colletotrichum fragariae. Tetrahedron Lett. 1996, 37, 5731–5734. [Google Scholar] [CrossRef]
- Aoyagi, A.; Ito-Kobayashi, M.; Ono, Y.; Furukawa, Y.; Takahashi, M.; Muramatsu, Y.; Umetani, M.; Takatsu, T. Colletoic acid, a novel 11β-hydroxysteroid dehydrogenase type 1 inhibitor from Colletotrichum gloeosporioides SANK 21404. J. Antibiot. 2008, 61, 136–141. [Google Scholar] [CrossRef]
- Hsiao, Y.; Cheng, M.J.; Chang, H.S.; Wu, M.D.; Hsieh, S.Y.; Liu, T.W.; Lin, C.H.; Yuan, G.F.; Chen, I.S. Six new metabolites produced by Colletotrichum aotearoa 09F0161, an endophytic fungus isolated from Bredia oldhamii. Nat. Prod. Res. 2016, 30, 251–258. [Google Scholar] [CrossRef]
- Hussain, H.; Root, N.; Jabeen, F.; Al-Harrasi, A.; Al-Rawahi, A.; Ahmad, M.; Hassan, Z.; Abbas, G.; Mabood, F.; Shah, A.; et al. Seimatoric acid and colletonoic acid: Two new compounds from the endophytic fungi, Seimatosporium sp. and Colletotrichum sp. Chin. Chem. Lett. 2014, 25, 1577–1579. [Google Scholar] [CrossRef]
- Wei, B.; Yang, Z.D.; Chen, X.W.; Zhou, S.Y.; Yu, H.T.; Sun, J.Y.; Yao, X.J.; Wang, Y.G.; Xue, H.Y. Colletotrilactam A–D, novel lactams from Colletotrichum gloeosporioides GT-7, a fungal endophyte of Uncaria rhynchophylla. Fitoterapia 2016, 113, 158–163. [Google Scholar] [CrossRef] [PubMed]
- Tianpanich, K.; Prachya, S.; Wiyakrutta, S.; Mahidol, C.; Ruchirawat, S.; Kittakoop, P. Radical scavenging and antioxidant activities of isocoumarins and a phthalide from the endophytic fungus Colletotrichum sp. J. Nat. Prod. 2011, 74, 79–81. [Google Scholar] [CrossRef] [PubMed]
- Yang, Z.; Bao, L.; Yin, Y.; Ding, G.; Ge, M.; Chen, D.; Qian, X. Pyrenocines N–O: Two novel pyrones from Colletotrichum sp. HCCB03289. J. Antibiot. 2014, 67, 791–793. [Google Scholar] [CrossRef]
- Ichihara, A.; Murakami, K.; Sakamura, S. Synthesis of pyrenocines A, B and pyrenochaetic acid A. Tetrahedron 1987, 43, 5245–5250. [Google Scholar] [CrossRef]
- Nunez, F.; Westphal, C.D.; Bermudez, E.; Asensio, M.A. Production of secondary metabolites by some terverticillate penicillia on carbohydrate-rich and meat substrates. J. Food Prot. 2007, 70, 2829–2836. [Google Scholar] [CrossRef] [PubMed]
- Frisvad, J.C.; Smedsgaard, J.; Larsen, T.O.; Samson, R.A. Mycotoxins, drugs and other extrolites produced by species in Penicillium subgenus Penicillium. Stud. Mycol. 2004, 49, e41. [Google Scholar]
- Ali, H.; Ries, M.I.; Lankhorst, P.P.; van der Hoeven, R.A.; Schouten, O.L.; Noga, M.; Hankemeier, T.; van Peij, N.N.M.E.; Bovenberg, R.A.L.; Vreeken, R.B.; et al. A non-canonical NRPS is involved in the synthesis of fungisporin and related hydrophobic cyclic tetrapeptides in Penicillium chrysogenum. PLoS ONE 2014, 9, e98212. [Google Scholar] [CrossRef]
- Wang, L.; Zhou, H.B.; Frisvad, J.C.; Samson, R.A. Penicillium persicinum, a new griseofulvin, chrysogine and roquefortine C producing species from Qinghai province, China. Antonie van Leeuwenhoek 2004, 86, 173–179. [Google Scholar] [CrossRef]
- Frisvad, J.C.; Samson, R.A.; Rassing, B.R.; van der Horst, M.I.; Van Rijn, F.T.J.; Stark, J. Penicillium discolor, a new species from cheese, nuts and vegetables. Antonie van Leeuwenhoek 1997, 72, 119–126. [Google Scholar] [CrossRef]
- Das, T.; Ray, P.; Nandy, S.; Al-Tawaha, A.R.; Pandey, D.K.; Kumar, V.; Dey, A. Piezophilic fungi: Sources of novel natural products with preclinical and clinical significance. In Extremophilic Fungi: Ecology, Physiology and Applications; Sahay, S., Ed.; Springer: Singapore, 2022; pp. 523–545. [Google Scholar]
- Ortega, H.E.; Torres-Mendoza, D.; Caballero, E.Z.; Cubilla-Rios, L. Structurally uncommon secondary metabolites derived from endophytic fungi. J. Fungi 2021, 7, 570. [Google Scholar] [CrossRef]
- Zain, M.E. Effect of olive oil on secondary metabolite and fatty acid profiles of Penicillium expansum, Aspergillus flavus, A. parasiticus and A. ochraceus. Aust. J. Basic Appl. Sci. 2009, 3, 4274–4280. [Google Scholar]
- Ariza, M.R.; Larsen, T.O.; Petersen, B.O.; Duus, J.Ø.; Christophersen, C.; Barrero, A.F. A novel alkaloid serantrypinone and the spiro azaphilone daldinin D from Penicillium thymicola. J. Nat. Prod. 2001, 64, 1590–1592. [Google Scholar] [CrossRef]
- Larsen, T.O.; Gareis, M.; Frisvad, J.C. Cell cytotoxicity and mycotoxin and secondary metabolite production by common penicillia on cheese agar. J. Agric. Food Chem. 2002, 50, 6148–6152. [Google Scholar] [CrossRef]
- Evidente, A.; Kornienko, A.; Cimmino, A.; Andolfi, A.; Lefranc, F.; Mathieu, V.; Kiss, R. Fungal metabolites with anticancer activity. Nat. Prod. Rep. 2014, 31, 617–627. [Google Scholar] [CrossRef] [PubMed]
- Perrone, G.; Susca, A. Penicillium species and their associated mycotoxins. In Mycotoxigenic Fungi; Springer: New York, NY, USA, 2017; pp. 107–119. [Google Scholar]
- Riolo, M.; Pane, A.; Santilli, E.; Moricca, S.; Cacciola, S.O. Susceptibility of Italian olive cultivars to various Colletotrichum species associated with fruit anthracnose. Plant Pathol. 2023, 72, 255–267. [Google Scholar] [CrossRef]
- Awuchi, C.G.; Ondari, E.N.; Ogbonna, C.U.; Upadhyay, A.K.; Baran, K.; Okpala, C.O.R.; Korzeniowska, M.; Guiné, R.P.F. Mycotoxins affecting animals, foods, humans, and plants: Types, occurrence, toxicities, action mechanisms, prevention, and detoxification strategies—A revisit. Foods 2021, 10, 1279. [Google Scholar] [CrossRef]
- Müller, M.E.; Steinhaus, M.; Böhm, J.; Müller, A.; Langer, E.; Andratsch, M.; Mack, B.; Novohradská, E.; Klempier, N. In vitro anti-inflammatory and anti-oxidative effects of altenariol and altertoxin II isolated from Alternaria species. Toxins 2018, 10, 228. [Google Scholar]
- Mitchell, C.G.; Slight, J.; Donaldson, K. Diffusible component from the spore surface of the fungus Aspergillus fumigatus which inhibits the macrophage oxidative burst is distinct from gliotoxin and other hyphal toxins. Thorax 1997, 52, 796–801. [Google Scholar] [CrossRef]
- Tolosa, J.; Barba, F.J.; Pallarés, N.; Ferrer, E. Mycotoxin identification and in silico toxicity assessment prediction in Atlantic Salmon. Mar. Drugs 2020, 18, 629. [Google Scholar] [CrossRef] [PubMed]
- da Cruz Cabral, L.; Rodriguero, M.; Stenglein, S.; Nielsen, K.F.; Patriarca, A. Characterization of small-spored Alternaria from Argentinean crops through a polyphasic approach. Int. J. Food Microbiol. 2017, 257, 206–215. [Google Scholar] [CrossRef] [PubMed]
- Man, Y.; Liang, G.; Li, A.; Pan, L. Analytical methods for the determination of Alternaria mycotoxins. Chromatographia 2017, 80, 9–22. [Google Scholar] [CrossRef]
- Mirocha, C.J.; Gilchrist, D.G.; Shier, W.T.; Abbas, H.K.; Wen, Y.; Vesonder, R.F. AAL Toxins, funionisms (biology and chemistry) and host-specificity concepts. Mycopathologia 1992, 117, 47–56. [Google Scholar] [CrossRef]
- Yamagishi, D.; Akamatsu, H.; Otani, H.; Kodama, M. Pathological evaluation of host-specific AAL-toxins and fumonisin mycotoxins produced by Alternaria and Fusarium species. J. Gen. Plant Pathol. 2006, 72, 323–326. [Google Scholar] [CrossRef]
- Tsuge, T.; Harimoto, Y.; Akimitsu, K.; Ohtani, K.; Kodama, M.; Akagi, Y.; Egusa, M.; Yamamoto, M.; Otani, H. Host-selective toxins produced by the plant pathogenic fungus Alternaria alternata. FEMS Microbiol. Rev. 2013, 37, 44–66. [Google Scholar] [CrossRef]
- Xu, J.; Yang, X.; Lin, Q. Chemistry and biology of Pestalotiopsis-derived natural products. Fungal Divers. 2014, 66, 37–68. [Google Scholar] [CrossRef]
- Evidente, A.; Punzo, B.; Andolfi, A.; Berestetskiy, A.; Motta, A. Alternethanoxin A and B, polycyclic ethanones produced by Alternaria sonchi, potential mycoherbicides for Sonchus arvenis Biocontrol. J. Agric. Food Chem. 2009, 57, 6656–6660. [Google Scholar] [CrossRef] [PubMed]
- Riolo, M.; Luz, C.; Santilli, E.; Meca, G.; Cacciola, S.O. Secondary metabolites produced by Colletotrichum spp. on different olive cultivars. Fungal Biol. 2023, in press. [CrossRef]
- Sadahiro, Y.; Hitora, Y.; Kimura, I.; Hitora-Imamura, N.; Onodera, R.; Motoyama, K.; Tsukamoto, S. Colletofragarone A2 Inhibits Cancer Cell Growth In Vivo and Leads to the Degradation and Aggregation of Mutant p53. Chem. Res. Toxicol. 2022, 35, 1598–1603. [Google Scholar] [CrossRef]
- Yan, X.; Qi, M.; Li, P.; Zhan, Y.; Shao, H. Apigenin in cancer therapy: Anti-cancer effects and mechanisms of action. Cell Biosci. 2017, 7, 50. [Google Scholar] [CrossRef]
- El Hajj Assaf, C.; Zetina-Serrano, C.; Tahtah, N.; Khoury, A.E.; Atoui, A.; Oswald, I.P.; Puel, O.; Lorber, S. Regulation of Secondary Metabolism in the Penicillium Genus. Int. J. Mol. Sci. 2020, 21, 9462. [Google Scholar] [CrossRef]
- Moake, M.M.; Zakour, O.I.P.; Worobo, R.W. Comprehensive review of patulin control methods in food. Compr. Rev. Food Sci. Food Saf. 2005, 4, 8–21. [Google Scholar] [CrossRef] [PubMed]
- Wogan, G.N.; Edwards, G.S.; Newberne, P.M. Acute and chronic toxicity of rubratoxin B. Toxicol. Appl. Pharmacol. 1971, 19, 712–720. [Google Scholar] [CrossRef]
- Yabe, K.; Nakamura, S.; Nakajima, M.; Fujimoto, H. Isolation and characterization of rubratoxin B biosynthetic intermediates from Penicillium sp. TP-F0595. J. Antibiot. 2003, 56, 534–540. [Google Scholar]
- Ding, Z.; Tao, T.; Wang, L.; Zhao, Y.; Huang, H.; Zhang, D.; Liu, M.; Wang, Z.; Han, J. Bioprospecting of novel and bioactive metabolites from endphytic fungi isolated from rubber tree Ficus elastica leaves. Microbiol. Biotechnol. 2019, 29, 731–738. [Google Scholar] [CrossRef]
- Ren-Yi, G.; Lei, X.; Yi, K.; Iii-Ming, C.; Jian-Chun, Q.; Li, L.; Sheng-Xiang, Y.; Li-Chun, Z. Chaetominine,(+)-alantrypinone, questin, isorhodoptilometrin, and 4-hydroxybenzaldehyde produced by the endophytic fungus Aspergillus sp. YL-6 inhibit wheat (Triticum aestivum) and radish (Raphanus sativus) germination. J. Plant Interact. 2015, 10, 87–92. [Google Scholar] [CrossRef]
- Kuriyama, T.; Kakemoto, E.; Takahashi, N.; Imamura, K.I.; Oyama, K.; Suzuki, E.; Harimaya, K.; Yaguchi, T.; Ozoe, Y. Receptor assay-guided isolation of anti-GABAergic insecticidal alkaloids from a fungal culture. J. Agric. Food Chem. 2004, 52, 3884–3887. [Google Scholar] [CrossRef] [PubMed]
- Zhu, X.; Zhou, D.; Liang, F.; Wu, Z.; She, Z.; Li, C. enochalasin K, a new unusual chaetoglobosin from the mangrove endophytic fungus Penicillium chrysogenum V11 and its effective semi-synthesis. Fitoterapia 2017, 123, 23–28. [Google Scholar] [CrossRef]
- Masi, M.; Aloi, F.; Nocera, P.; Cacciola, S.O.; Surico, G.; Evidente, A. Phytotoxic metabolites isolated from Neufusicoccum batangarum, the causal agent of the scabby canker of cactus pear (Opuntia ficus-indica L.). Toxins 2020, 12, 126. [Google Scholar] [CrossRef]
- Pryor, B.M.; Michailides, T.J. Morphological, pathogenic, and molecular characterization of Alternaria isolates associated with Alternaria late blight of pistachio. Phytopathology 2002, 92, 406–416. [Google Scholar] [CrossRef]
- Simmons, E.G. Alternaria: An Identification Manual: Fully Illustrated and with Catalogue Raisonné 1796–2007; CBS Fungal Biodiversity Centre: Utrecht, The Netherlands, 2007; p. 775. [Google Scholar]
- Agrios, G. Plant Pathology, 5th ed.; Elsevier: Amsterdam, The Netherlands; Academic Press: Cambridge, MA, USA, 2005; pp. 26–27, 398–401. [Google Scholar]
- White, T.J.; Bruns, T.; Lee, S.; Taylor, J.W. Amplification and direct sequencing of fungal ribosomal RNA genes for phylogenetics. In PCR Protocols: A Guide to Methods and Applications; Innis, M.A., Gelfand, D.H., Sninsky, J.J., White, T.J., Eds.; Academic Press, Inc.: San Diego, CA, USA, 1990; Volume 18, pp. 315–322. [Google Scholar]
- Carbone, I.; Kohn, L.M. A method for designing primer sets for speciation studies in filamentous ascomycetes. Mycologia 1999, 91, 553–556. [Google Scholar] [CrossRef]
- Glass, N.L.; Donaldson, G.C. Development of primer sets designed for use with the PCR to amplify conserved genes from filamentous ascomycetes. Appl. Environ. Microbiol. 1995, 61, 1323–1330. [Google Scholar] [CrossRef] [PubMed]
- Hong, S.G.; Cramer, R.A.; Lawrence, C.B.; Pryor, B.M. Alt a 1 allergen homologs from Alternaria and related taxa: Analysis of phylogenetic content and secondary structure. Fungal Genet. Biol. 2005, 42, 119–129. [Google Scholar] [CrossRef] [PubMed]
- Widiantini, F.; Yulia, E.; Nasahi, C. Antifungal activity of methanol-extracted secondary metabolites of rhizobacteria isolated from rhizosphere of oil palm trees against Ganoderma boninense Pat. IOP Conf. Ser. Earth Environ. Sci. 2019, 334, 012037. [Google Scholar] [CrossRef]
- El Jai, A.; Juan, C.; Juan-Garcia, A.; Manes, J.; Zinedine, A. Multi-mycotoxin contamination of green tea infusion and dietary exposure assessment in Moroccan population. Food Res. Intern. 2021, 140, 109958. [Google Scholar] [CrossRef]
- Pang, Z.; Chong, J.; Zhou, G.; De Lima Morais, D.A.; Chang, L.; Barrette, M.; Gauthier, C.; Jacques, P.É.; Li, S.; Xia, J. MetaboAnalyst 5.0: Narrowing the gap between raw spectra and functional insights. Nucleic Acids Res. 2021, 49, W388–W396. [Google Scholar] [CrossRef]














{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Pathogen | Types of Fruit | Cluster | Rate of Pathogens Isolated (%) | Total Rate (%) |
|---|---|---|---|---|
| Alternaria alternata | hail-injured | hail-injured ‘Lempso’ | 5% | 12% |
| hail-injured ‘Tapi’ | 4% | |||
| mummified | mummified ‘Lempso’ | 0% | ||
| mummified ‘Tapi’ | 0% | |||
| asymptomatic | asymptomatic ‘Lempso’ | 1% | ||
| asymptomatic ‘Tapi’ | 2% | |||
| Colletotrichum gloeosporioides | hail-injured | hail-injured ‘Lempso’ | 5% | 28% |
| hail-injured ‘Tapi’ | 7% | |||
| mummified | mummified ‘Lempso’ | 2% | ||
| mummified ‘Tapi’ | 2% | |||
| asymptomatic | asymptomatic ‘Lempso’ | 7% | ||
| asymptomatic ‘Tapi’ | 5% | |||
| Penicillium digitatum | hail-injured | hail-injured ‘Lempso’ | 10.5% | 60% |
| hail-injured ‘Tapi’ | 10% | |||
| mummified | mummified ‘Lempso’ | 15.5% | ||
| mummified ‘Tapi’ | 15% | |||
| asymptomatic | asymptomatic ‘Lempso’ | 4% | ||
| asymptomatic ‘Tapi’ | 5% |
| Isolate Code | Species | Accession Number | |||
|---|---|---|---|---|---|
| ITS-rDNA | ß-tubulin 2 | TEF-1a | Alt-1a | ||
| ID8O4 | Alternaria alternata | OQ929058 | - | OQ939919 | OQ938706 |
| ID8O5 | Alternaria alternata | OQ929059 | - | OQ939920 | OQ938707 |
| ID8O6 | Alternaria alternata | OQ929060 | - | OQ939921 | OQ938708 |
| ID8O7 | Alternaria alternata | OQ929061 | - | OQ939922 | OQ938709 |
| ID8O8 | Alternaria alternata | OQ929062 | - | OQ939923 | OQ938710 |
| ID8O9 | Alternaria alternata | OQ929063 | - | OQ939924 | OQ938711 |
| ID8O10 | Alternaria alternata | OQ929064 | - | OQ939925 | OQ938712 |
| ID8O11 | Alternaria alternata | OQ929065 | - | OQ939926 | OQ938713 |
| ID8O12 | Alternaria alternata | OQ929066 | - | OQ939927 | OQ938714 |
| ID3O13 | Alternaria alternata | OQ929067 | - | OQ939928 | OQ938715 |
| ID3O14 | Alternaria alternata | OQ929068 | - | OQ939929 | OQ938716 |
| ID3O15 | Alternaria alternata | OQ929069 | - | OQ939930 | OQ938717 |
| ID3O16 | Alternaria alternata | OQ929070 | - | OQ939931 | OQ938718 |
| ID3O17 | Alternaria alternata | OQ929071 | - | OQ939932 | OQ938719 |
| ID3O18 | Alternaria alternata | OQ929072 | - | OQ939933 | OQ938720 |
| ID3O19 | Alternaria alternata | OQ929073 | - | OQ939934 | OQ938721 |
| ID3O20 | Alternaria alternata | OQ929074 | - | OQ939935 | OQ938722 |
| ID3O21 | Alternaria alternata | OQ929075 | - | OQ939936 | OQ938723 |
| ID3O22 | Alternaria alternata | OQ929076 | - | OQ939937 | OQ938724 |
| ID3O23 | Alternaria alternata | OQ929077 | - | OQ939938 | OQ938725 |
| ID5O12 | Colletotrichum gloeosporioides | OQ929078 | OQ938686 | - | - |
| ID5O13 | Colletotrichum gloeosporioides | OQ929079 | OQ938687 | - | - |
| ID5O14 | Colletotrichum gloeosporioides | OQ929080 | OQ938688 | - | - |
| ID5O15 | Colletotrichum gloeosporioides | OQ929081 | OQ938689 | - | - |
| ID5O16 | Colletotrichum gloeosporioides | OQ929082 | OQ938690 | - | - |
| ID5O17 | Colletotrichum gloeosporioides | OQ929083 | OQ938691 | - | - |
| ID5O18 | Colletotrichum gloeosporioides | OQ929084 | OQ938692 | - | - |
| ID5O19 | Colletotrichum gloeosporioides | OQ929085 | OQ938693 | - | - |
| ID1O20 | Colletotrichum gloeosporioides | OQ929086 | OQ938694 | - | - |
| ID1O21 | Colletotrichum gloeosporioides | OQ929087 | OQ938695 | - | - |
| ID1O22 | Colletotrichum gloeosporioides | OQ929088 | OQ938696 | - | - |
| ID1O23 | Colletotrichum gloeosporioides | OQ929089 | OQ938697 | - | - |
| ID1O24 | Colletotrichum gloeosporioides | OQ929090 | OQ938698 | - | - |
| ID1O25 | Colletotrichum gloeosporioides | OQ929091 | OQ938699 | - | - |
| ID1O26 | Colletotrichum gloeosporioides | OQ929092 | OQ938700 | - | - |
| ID1O27 | Colletotrichum gloeosporioides | OQ929093 | OQ938701 | - | - |
| ID1O28 | Colletotrichum gloeosporioides | OQ929094 | OQ938702 | - | - |
| ID1O29 | Colletotrichum gloeosporioides | OQ929095 | OQ938703 | - | - |
| ID1O30 | Colletotrichum gloeosporioides | OQ929096 | OQ938704 | - | - |
| ID1O31 | Colletotrichum gloeosporioides | OQ929097 | OQ938705 | - | - |
| ID4O5 | Penicillium digitatum | OQ929098 | OQ938666 | - | - |
| ID4O6 | Penicillium digitatum | OQ929099 | OQ938667 | - | - |
| ID4O7 | Penicillium digitatum | OQ929100 | OQ938668 | - | - |
| ID4O8 | Penicillium digitatum | OQ929101 | OQ938669 | - | - |
| ID4O9 | Penicillium digitatum | OQ929102 | OQ938670 | - | - |
| ID4O10 | Penicillium digitatum | OQ929103 | OQ938671 | - | - |
| ID4O11 | Penicillium digitatum | OQ929104 | OQ938672 | - | - |
| ID4O12 | Penicillium digitatum | OQ929105 | OQ938673 | - | - |
| ID4O13 | Penicillium digitatum | OQ929106 | OQ938674 | - | - |
| ID4O14 | Penicillium digitatum | OQ929107 | OQ938675 | - | - |
| ID4O15 | Penicillium digitatum | OQ929108 | OQ938676 | - | - |
| ID4O16 | Penicillium digitatum | OQ929109 | OQ938677 | - | - |
| ID4O17 | Penicillium digitatum | OQ929110 | OQ938678 | - | - |
| ID4O18 | Penicillium digitatum | OQ929111 | OQ938679 | - | - |
| ID10O4 | Penicillium digitatum | OQ929112 | OQ938680 | - | - |
| ID10O5 | Penicillium digitatum | OQ929113 | OQ938681 | - | - |
| ID10O6 | Penicillium digitatum | OQ929114 | OQ938682 | - | - |
| ID10O7 | Penicillium digitatum | OQ929115 | OQ938683 | - | - |
| ID10O8 | Penicillium digitatum | OQ929116 | OQ938684 | - | - |
| ID10O9 | Penicillium digitatum | OQ929117 | OQ938685 | - | - |
| Secondary Metabolites | ID Code a | Matrices b | Chemical Class | Reference c |
|---|---|---|---|---|
| AAL-toxin TB2 | SM1 | juice | Mycotoxins (Cyclic peptides) | [88] |
| AAL-toxin TE2 | SM2 | peel and juice | Mycotoxins | [88] |
| AF-toxin II | SM3 | peel | Mycotoxins (Fumonisins) | [89] |
| Altenusin (ALN) | SM4 | peel and juice | Benzoxazole | [58] |
| Alternariol monomethyl ether (AME) | SM5 | peel and juice | Mycotoxins | [90] |
| Alternethanoxin A | SM6 | peel and juice | Ketone | [91] |
| Altersolanol L | SM7 | juice | Derivatives of anthraquinone | [92] |
| Aurasperone C | SM8 | juice | Prenylated benzophenones | [93] |
| Curvularin | SM9 | peel and juice | Macrocyclic lactones | [94] |
| Dihydroaltersolanol | SM10 | peel and juice | Cyclohexenones | [95] |
| Erythroglaucin | SM11 | peel | Anthraquinones | [88] |
| Macrosporin A | SM12 | peel | Macrolides | [58] |
| Maculosin | SM13 | juice | Tetramic acids | [96] |
| Porriolide | SM14 | juice | Furanones | [97] |
| Porritoxinol | SM15 | peel | Phthalides | [98] |
| Tentoxin (TEN) | SM16 | juice | Cyclic tetrapeptides | [58] |
| (+)-(3R,4S)-cis-4-hydroxy-6-deoxyscytalone | SM17 | juice | Tetramic acids | [99] |
| Ergosterol peroxide | SM18 | juice | Steroids | [100] |
| 5,4-dihydroxy-3,6,7-trimethoxy-8C-methylflavone | SM19 | peel and juice | Flavones | [101] |
| 5,4-dihydroxy-3,7,8-trimethoxy-6C-methylflavone | SM20 | peel and juice | Flavones | [101] |
| Alternariol-5-Me ether | SM21 | peel and juice | Coumarins | [102] |
| Apigenin-8-C-β-D-glucopyranoside | SM22 | peel and juice | Flavonoid glycosides | [103] |
| Collectotrichin A | SM23 | juice | Cyclopeptides | [104] |
| Colletofragarone A1 | SM24 | peel and juice | Sesquiterpenoids | [105] |
| Colletoic acid | SM25 | peel and juice | Diketopiperazines | [106] |
| Colletoketol | SM26 | juice | Macrolides | [101] |
| Colletomelleins B | SM27 | peel | Quinones | [107] |
| Colletonoic acid | SM28 | peel | Carboxylic acids | [108] |
| Colletotrilactam A | SM29 | peel and juice | Lactams | [109] |
| Colletotrilactam D | SM30 | peel and juice | Lactams | [109] |
| Fusarentin 6,7-dimethyl ether | SM31 | peel | Coumarins | [110] |
| Novae-zelandin A | SM32 | peel and juice | Phenazines | [111] |
| Phthalide | SM33 | peel | Cyclic anhydrides | [110] |
| Pyrenocine A | SM34 | juice | Pyrenocines | [112] |
| Alantrypinone | SM35 | peel and juice | Quinazolinones | [88] |
| Anacine | SM36 | peel | Quinazolinones | [113] |
| Asteltoxin | SM37 | peel | Anthraquinones | [114] |
| Atrovenetins | SM38 | peel and juice | Xanthones | [114] |
| Fungisporin | SM39 | peel and juice | Cyclohexenones | [115] |
| Lichexanthone | SM40 | peel and juice | Xanthones | [116] |
| Palitantin | SM41 | juice | Flavones | [117] |
| Penipacid B | SM42 | peel | Diketopiperazines | [118] |
| Penochalasin K | SM43 | peel and juice | Macrolides | [119] |
| Rubratoxin B | SM44 | juice | Mycotoxins | [120] |
| Serantrypinone | SM45 | peel and juice | Quinones | [121] |
| Solistatin | SM46 | peel | Lactones | [122] |
| Patulin | SM47 | peel and juice | Mycotoxins (Furanocoumarins) | [46] |
| Cluster | Number of Fruits | Cultivar |
|---|---|---|
| Asymptomatic fruits (Asym_L) | 60 a | Tarocco Lempso |
| Hail-injured fruits (H-Inj_L) | 60 | |
| Mummified fruits (Mum_L) | 60 | |
| Asymptomatic fruits (Asym_T) | 60 | Tarocco Tapi |
| Hail-injured fruits (H-Inj_T) | 60 | |
| Mummified fruits (Mum_T) | 60 |
| Barcode Region/Gene | Target Organism/s | Primers | Expected Amplicon Size (bp) | References |
|---|---|---|---|---|
| Internal transcribed spacer (ITS) region of the ribosomal DNA | Alternaria, Colletotrichum, and Penicillium | ITS-1 5′-TCCGTAGGTGAACCTGCGG-3′ | ~700 | [152] |
| ITS-4 5′-TCCTCCGCTTA TTGATATGC-3′ | ||||
| Intron of translation elongation factor 1 alpha gene (tef1) | Alternaria | EF1-728F 5′-CAT CGA GAA GTT CGA GAA GG-3′ | ~350 | [153] |
| EF1-986R 5′-TAC TTG AAG GAA CCC TTA CC-3′ | ||||
| β-tubulin gene (tub2) | Colletotrichum and Penicillium | Bt2a 5′-GGTAACCAAATCGGTGCTGCTTTC-3′ | ~400 (Penicillium) ~773 (Colletotrichum) | [154] |
| Bt2b 5′-ACCCTCAGTGTAGTGACCCTTGGC-3′ | ||||
| Alt-1a allergen protein | Alternaria | Alt-for 5′-TATGCAGTTCACCACCATCGC-3′ | ~400 | [155] |
| Alt-rev 5′-ACGAGGGTGAYGTAGGCGTC-3′ |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Rovetto, E.I.; Luz, C.; La Spada, F.; Meca, G.; Riolo, M.; Cacciola, S.O. Diversity of Mycotoxins and Other Secondary Metabolites Recovered from Blood Oranges Infected by Colletotrichum, Alternaria, and Penicillium Species. Toxins 2023, 15, 407. https://doi.org/10.3390/toxins15070407
Rovetto EI, Luz C, La Spada F, Meca G, Riolo M, Cacciola SO. Diversity of Mycotoxins and Other Secondary Metabolites Recovered from Blood Oranges Infected by Colletotrichum, Alternaria, and Penicillium Species. Toxins. 2023; 15(7):407. https://doi.org/10.3390/toxins15070407
Chicago/Turabian StyleRovetto, Ermes Ivan, Carlos Luz, Federico La Spada, Giuseppe Meca, Mario Riolo, and Santa Olga Cacciola. 2023. "Diversity of Mycotoxins and Other Secondary Metabolites Recovered from Blood Oranges Infected by Colletotrichum, Alternaria, and Penicillium Species" Toxins 15, no. 7: 407. https://doi.org/10.3390/toxins15070407
APA StyleRovetto, E. I., Luz, C., La Spada, F., Meca, G., Riolo, M., & Cacciola, S. O. (2023). Diversity of Mycotoxins and Other Secondary Metabolites Recovered from Blood Oranges Infected by Colletotrichum, Alternaria, and Penicillium Species. Toxins, 15(7), 407. https://doi.org/10.3390/toxins15070407