Zearalenone Exposure Disrupts STAT-ISG15 in Rat Colon: A Potential Linkage between Zearalenone and Inflammatory Bowel Disease


Abstract
1. Introduction
2. Results
2.1. ZEN Exposure Induces Histopathological Changes in the Colon
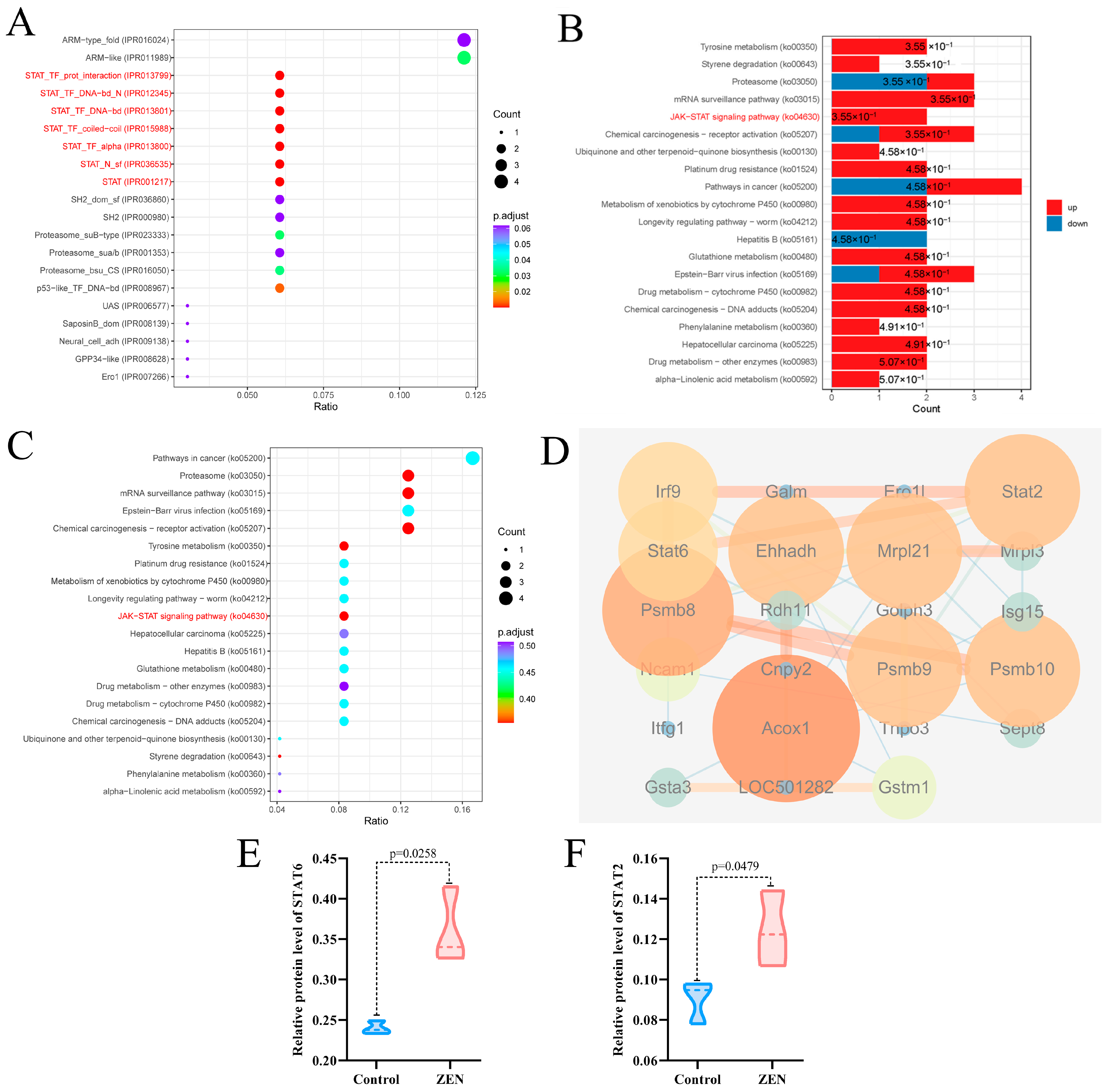
2.2. ZEN Exposure Alters the Proteomics of the Colon
2.3. ZEN Exposure Affects Proteasome Mediated Ubiquitination in the Colon
2.4. ZEN Exposure Affects the Expression Levels of STAT Proteins in the Colon
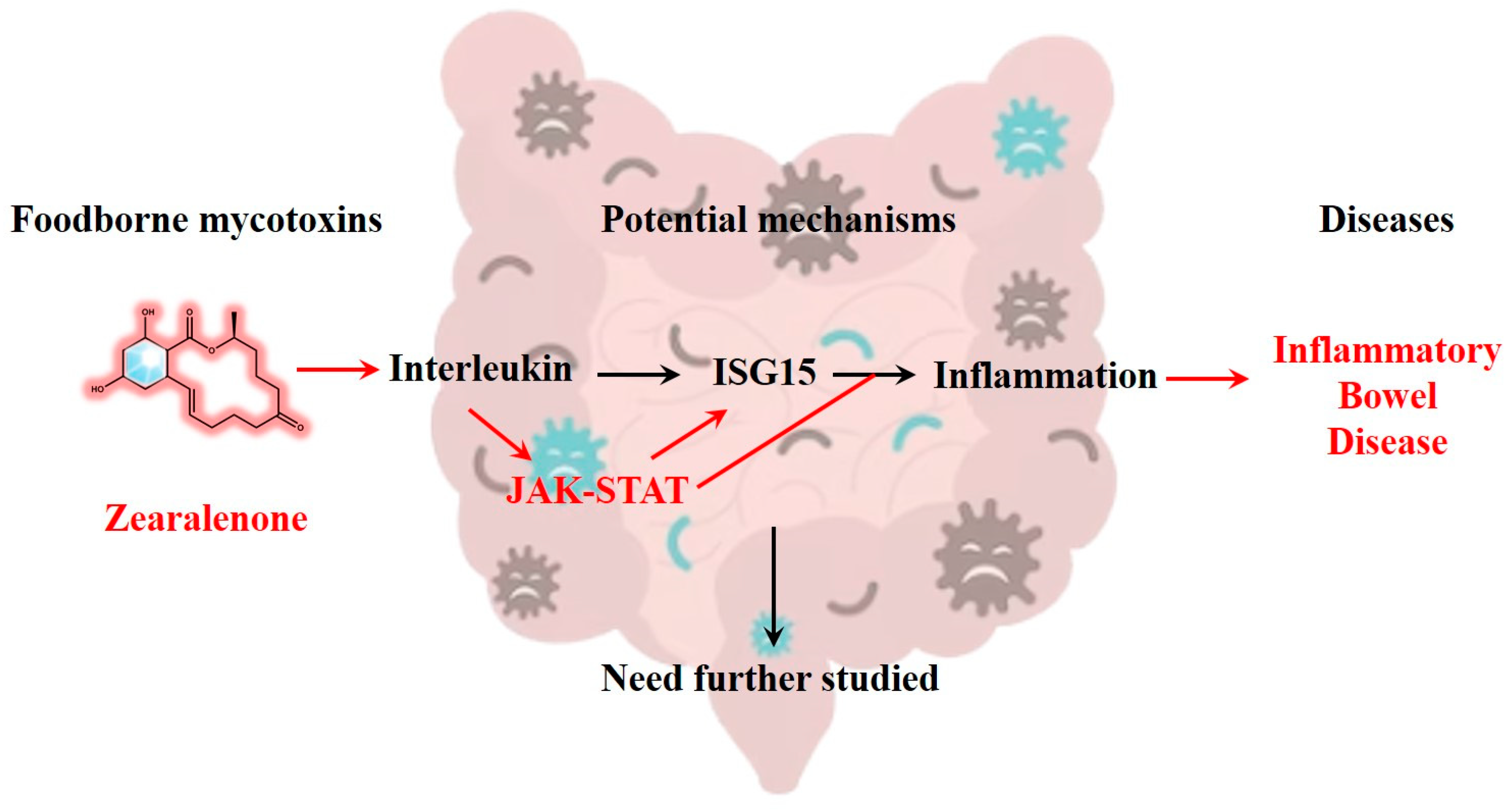
2.5. ZEN Exposure May Increase the Risk of IBD by Disrupting STAT Family
3. Discussion
4. Conclusions
5. Materials and Methods
5.1. Chemicals and Reagents
5.2. Animal Study
5.3. HE Staining
5.4. Proteomic Analysis of Rat Colon
5.5. Gene Expression Data Set
5.6. Identification of DEGs
5.7. Gene Set Enrichment Analysis and PPI Analysis
5.8. Statistical Analyses
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Rai, A.; Das, M.; Tripathi, A. Occurrence and toxicity of a fusarium mycotoxin, zearalenone. Crit. Rev. Food Sci. Nutr. 2020, 60, 2710–2729. [Google Scholar] [CrossRef]
- Hao, W.; Guan, S.; Li, A.; Wang, J.; An, G.; Hofstetter, U.; Schatzmayr, G. Mycotoxin Occurrence in Feeds and Raw Materials in China: A Five-Year Investigation. Toxins 2023, 15, 63. [Google Scholar] [CrossRef] [PubMed]
- Iwase, C.H.T.; Piacentini, K.C.; Silva, N.C.C.; Rebellato, A.P.; Rocha, L.O. Deoxynivalenol and zearalenone in Brazilian barley destined for brewing. Food Addit. Contam. Part B Surveill. 2023, 16, 86–92. [Google Scholar] [CrossRef]
- Ruan, H.; Lu, Q.; Wu, J.; Qin, J.; Sui, M.; Sun, X.; Shi, Y.; Luo, J.; Yang, M. Hepatotoxicity of food-borne mycotoxins: Molecular mechanism, anti-hepatotoxic medicines and target prediction. Crit. Rev. Food Sci. Nutr. 2022, 62, 2281–2308. [Google Scholar] [CrossRef]
- Llorens Castelló, P.; Sacco, M.A.; Aquila, I.; Moltó Cortés, J.C.; Juan García, C. Evaluation of Zearalenones and Their Metabolites in Chicken, Pig and Lamb Liver Samples. Toxins 2022, 14, 782. [Google Scholar] [CrossRef] [PubMed]
- Ruan, H.; Wang, Y.; Hou, Y.; Zhang, J.; Wu, J.; Zhang, F.; Sui, M.; Luo, J.; Yang, M. Zearalenone-14-Glucoside Is Hydrolyzed to Zearalenone by β-Glucosidase in Extracellular Matrix to Exert Intracellular Toxicity in KGN Cells. Toxins 2022, 14, 458. [Google Scholar] [CrossRef] [PubMed]
- E Commission. Commission Recommendation of 17 August 2006 on the Presence of Deoxynivalenol, Zearalenone, Ochratoxin A, T-2 and HT-2 and Fumonisins in Products Intended for Animal Nutrition (2006/576/EC as Amended) (OJ. L./229/7). Off. J. Eur. Union 2006, 229, 7–9. [Google Scholar]
- Wang, T.; Ye, Y.; Ji, J.; Yang, X.; Xu, J.; Wang, J.S.; Han, X.; Zhang, T.; Sun, X. Diet composition affects long-term zearalenone exposure on the gut-blood-liver axis metabolic dysfunction in mice. Ecotoxicol. Environ. Saf. 2022, 236, 113466. [Google Scholar] [CrossRef]
- Chen, Z.; Wang, F.; Zhang, W.; Zhou, S.; Wen, D.; Mu, R. Chronic exposure to zearalenone induces intestinal inflammation and oxidative injury in adult Drosophila melanogaster midgut. Ecotoxicol. Environ. Saf. 2023, 251, 114555. [Google Scholar] [CrossRef]
- Braicu, C.; Cojocneanu-Petric, R.; Jurj, A.; Gulei, D.; Taranu, I.; Gras, A.M.; Marin, D.E.; Berindan-Neagoe, I. Microarray based gene expression analysis of Sus Scrofa duodenum exposed to zearalenone: Significance to human health. BMC Genom. 2016, 17, 646. [Google Scholar] [CrossRef]
- Cai, G.; Zhong, F.; Cao, Q.; Bai, Y.; Zou, H.; Gu, J.; Yuan, Y.; Zhu, G.; Liu, Z.; Bian, J. ZEA and DON inhibited inflammation after L. monocytogenes infection and induced ribosomal hyperfunction. Ecotoxicol. Environ. Saf. 2022, 236, 113470. [Google Scholar] [CrossRef]
- Fan, W.; Lv, Y.; Ren, S.; Shao, M.; Shen, T.; Huang, K.; Zhou, J.; Yan, L.; Song, S. Zearalenone (ZEA)-induced intestinal inflammation is mediated by the NLRP3 inflammasome. Chemosphere 2018, 190, 272–279. [Google Scholar] [CrossRef] [PubMed]
- Wang, S.; Fu, W.; Zhao, X.; Chang, X.; Liu, H.; Zhou, L.; Li, J.; Cheng, R.; Wu, X.; Li, X.; et al. Zearalenone disturbs the reproductive-immune axis in pigs: The role of gut microbial metabolites. Microbiome 2022, 10, 234. [Google Scholar] [CrossRef]
- Massironi, S.; Viganò, C.; Palermo, A.; Pirola, L.; Mulinacci, G.; Allocca, M.; Peyrin-Biroulet, L.; Danese, S. Inflammation and malnutrition in inflammatory bowel disease. Lancet Gastroenterol. Hepatol. 2023, 8, 579–590. [Google Scholar] [CrossRef] [PubMed]
- Hodson, R. Inflammatory bowel disease. Nature 2016, 540, S97. [Google Scholar] [CrossRef] [PubMed]
- Maresca, M.; Fantini, J. Some food-associated mycotoxins as potential risk factors in humans predisposed to chronic intestinal inflammatory diseases. Toxicon 2010, 56, 282–294. [Google Scholar] [CrossRef] [PubMed]
- Payros, D.; Alassane-Kpembi, I.; Laffitte, J.; Lencina, C.; Neves, M.; Bracarense, A.P.; Pinton, P.; Ménard, S.; Oswald, I.P. Dietary Exposure to the Food Contaminant Deoxynivalenol Triggers Colonic Breakdown by Activating the Mitochondrial and the Death Receptor Pathways. Mol. Nutr. Food Res. 2021, 65, e2100191. [Google Scholar] [CrossRef]
- Gao, Y.; Meng, L.; Liu, H.; Wang, J.; Zheng, N. The Compromised Intestinal Barrier Induced by Mycotoxins. Toxins 2020, 12, 619. [Google Scholar] [CrossRef]
- Wang, Z.K.; Yang, Y.S.; Stefka, A.T.; Sun, G.; Peng, L.H. Review article: Fungal microbiota and digestive diseases. Aliment. Pharm. Ther. 2014, 39, 751–766. [Google Scholar] [CrossRef]
- Payros, D.; Ménard, S.; Laffitte, J.; Neves, M.; Tremblay-Franco, M.; Luo, S.; Fouche, E.; Snini, S.P.; Theodorou, V.; Pinton, P.; et al. The food contaminant, deoxynivalenol, modulates the Thelper/Treg balance and increases inflammatory bowel diseases. Arch. Toxicol. 2020, 94, 3173–3184. [Google Scholar] [CrossRef]
- Zhou, J.; Tang, L.; Wang, J.S. Aflatoxin B1 Induces Gut-Inflammation-Associated Fecal Lipidome Changes in F344 Rats. Toxicol. Sci. 2021, 183, 363–377. [Google Scholar] [CrossRef]
- Hermann, M.; Bogunovic, D. ISG15: In Sickness and in Health. Trends Immunol. 2017, 38, 79–93. [Google Scholar] [CrossRef]
- Kao, T.J.; Wu, C.C.; Phan, N.N.; Liu, Y.H.; Ta, H.D.K.; Anuraga, G.; Wu, Y.F.; Lee, K.H.; Chuang, J.Y.; Wang, C.Y. Prognoses and genomic analyses of proteasome 26S subunit, ATPase (PSMC) family genes in clinical breast cancer. Aging 2021, 13, 17970. [Google Scholar] [CrossRef] [PubMed]
- Acuña-Gutiérrez, C.; Jiménez, V.M.; Müller, J. Occurrence of mycotoxins in pulses. Compr. Rev. Food Sci. Food Saf. 2022, 21, 4002–4017. [Google Scholar] [CrossRef] [PubMed]
- Juraschek, L.M.; Kappenberg, A.; Amelung, W. Mycotoxins in soil and environment. Sci. Total Environ. 2022, 814, 152425. [Google Scholar] [CrossRef] [PubMed]
- Khor, B.; Gardet, A.; Xavier, R.J. Genetics and pathogenesis of inflammatory bowel disease. Nature 2011, 474, 307–317. [Google Scholar] [CrossRef]
- Piovani, D.; Danese, S.; Peyrin-Biroulet, L.; Nikolopoulos, G.K.; Lytras, T.; Bonovas, S. Environmental Risk Factors for Inflammatory Bowel Diseases: An Umbrella Review of Meta-analyses. Gastroenterology 2019, 157, 647–659.e4. [Google Scholar] [CrossRef]
- Sokol, H.; Leducq, V.; Aschard, H.; Pham, H.P.; Jegou, S.; Landman, C.; Cohen, D.; Liguori, G.; Bourrier, A.; Nion-Larmurier, I.; et al. Fungal microbiota dysbiosis in IBD. Gut 2017, 66, 1039–1048. [Google Scholar] [CrossRef]
- Ott, S.J.; Kühbacher, T.; Musfeldt, M.; Rosenstiel, P.; Hellmig, S.; Rehman, A.; Drews, O.; Weichert, W.; Timmis, K.N.; Schreiber, S. Fungi and inflammatory bowel diseases: Alterations of composition and diversity. Scand. J. Gastroenterol. 2008, 43, 831–841. [Google Scholar] [CrossRef]
- Wan, M.L.Y.; Co, V.A.; El-Nezami, H. Endocrine disrupting chemicals and breast cancer: A systematic review of epidemiological studies. Crit. Rev. Food Sci. Nutr. 2022, 62, 6549–6576. [Google Scholar] [CrossRef]
- Cai, G.; Xia, S.; Zhong, F.; Liu, S.; Gu, J.; Yuan, Y.; Zhu, G.; Zou, H.; Liu, Z.; Bian, J. Zearalenone and deoxynivalenol reduced Th1-mediated cellular immune response after Listeria monocytogenes infection by inhibiting CD4+ T cell activation and differentiation. Environ. Pollut. 2021, 284, 117514. [Google Scholar] [CrossRef]
- Ma, L.; Jiang, Y.; Lu, F.; Wang, S.; Liu, M.; Liu, F.; Huang, L.; Li, Y.; Jiao, N.; Jiang, S.; et al. Quantitative Proteomic Analysis of Zearalenone-Induced Intestinal Damage in Weaned Piglets. Toxins 2022, 14, 702. [Google Scholar] [CrossRef]
- Zhang, W.; Zhang, S.; Wang, J.; Shan, A.; Xu, L. Changes in intestinal barrier functions and gut microbiota in rats exposed to zearalenone. Ecotoxicol. Environ. Saf. 2020, 204, 111072. [Google Scholar] [CrossRef]
- Taranu, I.; Braicu, C.; Marin, D.E.; Pistol, G.C.; Motiu, M.; Balacescu, L.; Beridan Neagoe, I.; Burlacu, R. Exposure to zearalenone mycotoxin alters in vitro porcine intestinal epithelial cells by differential gene expression. Toxicol. Lett. 2015, 232, 310–325. [Google Scholar] [CrossRef] [PubMed]
- Coskun, M.; Salem, M.; Pedersen, J.; Nielsen, O.H. Involvement of JAK/STAT signaling in the pathogenesis of inflammatory bowel disease. Pharm. Res. 2013, 76, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Ţieranu, C.G.; Dobre, M.; Mănuc, T.E.; Milanesi, E.; Pleşea, I.E.; Popa, C.; Mănuc, M.; Ţieranu, I.; Preda, C.M.; Diculescu, M.M.; et al. Gene expression profile of endoscopically active and inactive ulcerative colitis: Preliminary data. Rom. J. Morphol. Embryol. 2017, 58, 1301–1307. [Google Scholar] [PubMed]
- Cordes, F.; Foell, D.; Ding, J.N.; Varga, G.; Bettenworth, D. Differential regulation of JAK/STAT-signaling in patients with ulcerative colitis and Crohn’s disease. World J. Gastroenterol. 2020, 26, 4055–4075. [Google Scholar] [CrossRef]
- Lee, H.H.; Jang, E.; Kang, S.Y.; Shin, J.S.; Han, H.S.; Kim, T.W.; Lee, D.H.; Lee, J.H.; Jang, D.S.; Lee, K.T. Anti-inflammatory potential of Patrineolignan B isolated from Patrinia scabra in LPS-stimulated macrophages via inhibition of NF-κB, AP-1, and JAK/STAT pathways. Int. Immunopharmacol. 2020, 86, 106726. [Google Scholar] [CrossRef] [PubMed]
- Shouval, D.S.; Rufo, P.A. The Role of Environmental Factors in the Pathogenesis of Inflammatory Bowel Diseases: A Review. JAMA Pediatr. 2017, 171, 999–1005. [Google Scholar] [CrossRef]
- Schreiber, S.; Rosenstiel, P.; Hampe, J.; Nikolaus, S.; Groessner, B.; Schottelius, A.; Kühbacher, T.; Hämling, J.; Fölsch, U.R.; Seegert, D. Activation of signal transducer and activator of transcription (STAT) 1 in human chronic inflammatory bowel disease. Gut 2002, 51, 379–385. [Google Scholar] [CrossRef]
- Labbé, C.; Boucher, G.; Foisy, S.; Alikashani, A.; Nkwimi, H.; David, G.; Beaudoin, M.; Goyette, P.; Charron, G.; Xavier, R.J.; et al. Genome-wide expression profiling implicates a MAST3-regulated gene set in colonic mucosal inflammation of ulcerative colitis patients. Inflamm. Bowel Dis. 2012, 18, 1072–1080. [Google Scholar] [CrossRef]
- Perng, Y.C.; Lenschow, D.J. ISG15 in antiviral immunity and beyond. Nat. Rev. Microbiol. 2018, 16, 423–439. [Google Scholar] [CrossRef]
- Darb-Esfahani, S.; Sinn, B.V.; Rudl, M.; Sehouli, J.; Braicu, I.; Dietel, M.; Denkert, C. Interferon-stimulated gene, 15 kDa (ISG15) in ovarian high-grade serous carcinoma: Prognostic impact and link to NF-κB pathway. Int. J. Gynecol. Pathol. 2014, 33, 16–22. [Google Scholar] [CrossRef]
- Gargan, S.; Ahmed, S.; Mahony, R.; Bannan, C.; Napoletano, S.; O’Farrelly, C.; Borrow, P.; Bergin, C.; Stevenson, N.J. HIV-1 Promotes the Degradation of Components of the Type 1 IFN JAK/STAT Pathway and Blocks Anti-viral ISG Induction. EBioMedicine 2018, 30, 203–216. [Google Scholar] [CrossRef]
- Osiak, A.; Utermöhlen, O.; Niendorf, S.; Horak, I.; Knobeloch, K.P. ISG15, an interferon-stimulated ubiquitin-like protein, is not essential for STAT1 signaling and responses against vesicular stomatitis and lymphocytic choriomeningitis virus. Mol. Cell Biol. 2005, 25, 6338–6345. [Google Scholar] [CrossRef] [PubMed]
- Luo, X.Y.; Liu, Q.; Yang, H.; Tan, Q.; Gan, L.Q.; Ren, F.L.; Wang, H. OSMR gene effect on the pathogenesis of chronic autoimmune Urticaria via the JAK/STAT3 pathway. Mol. Med. 2018, 24, 28. [Google Scholar] [CrossRef]
- Østvik, A.E.; Svendsen, T.D.; Granlund AV, B.; Doseth, B.; Skovdahl, H.K.; Bakke, I.; Thorsvik, S.; Afroz, W.; Walaas, G.A.; Mollnes, T.E.; et al. Intestinal Epithelial Cells Express Immunomodulatory ISG15 During Active Ulcerative Colitis and Crohn’s Disease. J. Crohns Colitis 2020, 14, 920–934. [Google Scholar] [CrossRef] [PubMed]
- Obremski, K.; Trybowski, W.; Wojtacha, P.; Gajęcka, M.; Tyburski, J.; Zielonka, Ł. The Effect of Zearalenone on the Cytokine Environment, Oxidoreductive Balance and Metabolism in Porcine Ileal Peyer’s Patches. Toxins 2020, 12, 350. [Google Scholar] [CrossRef] [PubMed]
- Gan, F.; Lin, Z.; Tang, J.; Chen, X.; Huang, K. Deoxynivalenol at No-Observed Adverse-Effect Levels Aggravates DSS-Induced Colitis through the JAK2/STAT3 Signaling Pathway in Mice. J. Agric. Food Chem. 2023, 71, 4144–4152. [Google Scholar] [CrossRef]
- Liu, G.; Yan, T.; Wang, J.; Huang, Z.; Chen, X.; Jia, G.; Wu, C.; Zhao, H.; Xue, B.; Xiao, L.; et al. Biological system responses to zearalenone mycotoxin exposure by integrated metabolomic studies. J. Agric. Food Chem. 2013, 61, 11212–11221. [Google Scholar] [CrossRef]
- Li, Q.; Cui, Y.; Xu, B.; Wang, Y.; Lv, F.; Li, Z.; Li, H.; Chen, X.; Peng, X.; Chen, Y.; et al. Main active components of Jiawei Gegen Qinlian decoction protects against ulcerative colitis under different dietary environments in a gut microbiota-dependent manner. Pharm. Res. 2021, 170, 105694. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Kong, Q.; Yue, J.; Gou, X.; Xu, M.; Wu, X. Genome-edited skin epidermal stem cells protect mice from cocaine-seeking behaviour and cocaine overdose. Nat. Biomed. Eng. 2019, 3, 105–113. [Google Scholar] [CrossRef] [PubMed]
- Li, H.; Chen, X.; Liu, J.; Chen, M.; Huang, M.; Huang, G.; Chen, X.; Du, Q.; Su, J.; Lin, R. Ethanol extract of Centella asiatica alleviated dextran sulfate sodium-induced colitis: Restoration on mucosa barrier and gut microbiota homeostasis. J. Ethnopharmacol. 2021, 267, 113445. [Google Scholar] [CrossRef]
- Wang, M.; Zhang, Z.; Liu, J.; Song, M.; Zhang, T.; Chen, Y.; Hu, H.; Yang, P.; Li, B.; Song, X.; et al. Gefitinib and fostamatinib target EGFR and SYK to attenuate silicosis: A multi-omics study with drug exploration. Signal Transduct. Target. Ther. 2022, 7, 157. [Google Scholar] [CrossRef]
- Ji, X.; Yue, H.; Li, G.; Sang, N. Maternal smoking-induced lung injuries in dams and offspring via inflammatory cytokines. Environ. Int. 2021, 156, 106618. [Google Scholar] [CrossRef] [PubMed]
- Zhou, Y.; Zhou, B.; Pache, L.; Chang, M.; Khodabakhshi, A.H.; Tanaseichuk, O.; Benner, C.; Chanda, S.K. Metascape provides a biologist-oriented resource for the analysis of systems-level datasets. Nat. Commun. 2019, 10, 1523. [Google Scholar] [CrossRef]
- Szklarczyk, D.; Gable, A.L.; Nastou, K.C.; Lyon, D.; Kirsch, R.; Pyysalo, S.; Doncheva, N.T.; Legeay, M.; Fang, T.; Bork, P.; et al. The STRING database in 2021: Customizable protein-protein networks, and functional characterization of user-uploaded gene/measurement sets. Nucleic Acids Res. 2021, 49, D605–D612. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Proteins | Control-1 | Control-2 | Control-3 | ZEN-1 | ZEN-2 | ZEN-3 | * Log2FC | * p-Value |
|---|---|---|---|---|---|---|---|---|
| A1AG | 0.4694 | 0.3347 | 0.791 | 1.1186 | 2.379 | 1.4882 | 1.6441 | 0.0264 |
| G3V9Z6 | 0.4644 | 0.6189 | 0.3563 | 1.0716 | 1.1777 | 0.8675 | 1.1144 | 0.0206 |
| FAAA | 0.445 | 0.5113 | 0.7153 | 1.6298 | 0.7939 | 1.163 | 1.1015 | 0.0491 |
| HPPD | 0.301 | 0.3293 | 0.3467 | 0.5488 | 0.8308 | 0.5625 | 0.9911 | 0.0292 |
| D3ZMR9 | 0.1233 | 0.146 | 0.1092 | 0.3057 | 0.1749 | 0.2517 | 0.9517 | 0.0402 |
| GSTA3 | 1.475 | 1.5525 | 2.2145 | 3.2378 | 3.0798 | 3.6151 | 0.9221 | 0.025 |
| GSTM1 | 2.166 | 2.2667 | 3.5107 | 5.0322 | 6.2371 | 3.7568 | 0.9197 | 0.0396 |
| F1LXY6 | 0.5733 | 0.4813 | 0.5203 | 1.1055 | 0.9261 | 0.8807 | 0.8869 | 0.0028 |
| F1M609 | 0.0766 | 0.1025 | 0.0857 | 0.1299 | 0.1656 | 0.127 | 0.6741 | 0.0177 |
| GALM | 0.736 | 1.0308 | 0.9585 | 1.5363 | 1.5418 | 1.1391 | 0.6299 | 0.0381 |
| Q5U355 | 0.0961 | 0.1176 | 0.1024 | 0.1379 | 0.1793 | 0.1619 | 0.6 | 0.0146 |
| Q1KQ07 | 0.2377 | 0.2491 | 0.2333 | 0.4148 | 0.3265 | 0.3403 | 0.587 | 0.0258 |
| SAM50 | 0.7642 | 0.7959 | 0.9456 | 1.3768 | 1.0104 | 1.269 | 0.5451 | 0.0351 |
| M0R3V4 | 0.3655 | 0.5018 | 0.4934 | 0.6545 | 0.7441 | 0.5801 | 0.5402 | 0.0452 |
| A0A0G2JTA0 | 0.206 | 0.2746 | 0.2259 | 0.3228 | 0.2976 | 0.4062 | 0.539 | 0.0422 |
| A0A0G2K016 | 0.4296 | 0.4338 | 0.3282 | 0.5172 | 0.5157 | 0.6476 | 0.4961 | 0.0451 |
| PRS6B | 1.6427 | 2.0863 | 1.7524 | 2.3933 | 2.8153 | 2.3946 | 0.4721 | 0.024 |
| Q5XI26 | 0.0948 | 0.0781 | 0.0979 | 0.144 | 0.1224 | 0.1069 | 0.4632 | 0.0479 |
| A0JN30 | 1.9871 | 2.0197 | 1.9874 | 2.634 | 2.6796 | 2.83 | 0.4421 | 0.0033 |
| A0A096MJB3 | 0.1177 | 0.1115 | 0.0988 | 0.1447 | 0.1345 | 0.1658 | 0.4396 | 0.0205 |
| HNRH2 | 0.6268 | 0.7558 | 0.7903 | 0.8522 | 1.0493 | 0.9405 | 0.3873 | 0.0456 |
| A0A0G2K1E5 | 0.079 | 0.0864 | 0.0795 | 0.0965 | 0.0989 | 0.1089 | 0.3126 | 0.0112 |
| D3ZDI7 | 0.4407 | 0.4408 | 0.3788 | 0.4798 | 0.5652 | 0.5167 | 0.3093 | 0.0363 |
| APT | 9.2063 | 10.3972 | 10.1311 | 11.2779 | 12.3006 | 12.4024 | 0.2751 | 0.0175 |
| UB2L6 | 0.3352 | 0.3495 | 0.3854 | 0.4214 | 0.4154 | 0.4573 | 0.2742 | 0.0238 |
| Q569C9 | 0.3141 | 0.3756 | 0.3559 | 0.4466 | 0.4055 | 0.4086 | 0.2698 | 0.0493 |
| PSB3 | 3.8159 | 3.9469 | 3.4117 | 3.152 | 2.917 | 3.221 | −0.2665 | 0.0319 |
| F1M7Y3 | 0.9241 | 0.9139 | 0.8875 | 0.7159 | 0.7745 | 0.6323 | −0.3606 | 0.0453 |
| D4AAM0 | 0.3923 | 0.3254 | 0.3651 | 0.2561 | 0.2751 | 0.28 | −0.4165 | 0.0189 |
| CYBC1 | 0.1593 | 0.1766 | 0.165 | 0.1328 | 0.1227 | 0.1057 | −0.4719 | 0.024 |
| Q6MGA6 | 1.4047 | 1.3431 | 1.0569 | 0.9763 | 0.8123 | 0.9176 | −0.4915 | 0.0413 |
| ERO1A | 0.3429 | 0.2804 | 0.2897 | 0.2111 | 0.1824 | 0.2337 | −0.5415 | 0.0173 |
| CNO10 | 0.1421 | 0.1117 | 0.1166 | 0.0745 | 0.082 | 0.0851 | −0.6162 | 0.0143 |
| Q6AXX5 | 0.6402 | 0.3931 | 0.4837 | 0.3111 | 0.2606 | 0.2541 | −0.8772 | 0.036 |
| F1LUV9 | 0.4489 | 0.2294 | 0.3003 | 0.0857 | 0.1227 | 0.1787 | −1.3379 | 0.032 |
| Score | Appearance |
|---|---|
| 0 None; 1 Mild; 2 Moderate; 3 Severe | Submucosal edema |
| 0 None; 1 Localized; 2 Moderate; 3 Severe | Damage/necrosis |
| 0 None; 1 Mild; 2 Moderate; 3 Severe | Inflammatory cell infiltration |
| 0 None; 1 Mild; 2 Moderate; 3 Severe | Vasculitis |
| 0 No; 1 Yes | Perforation |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ruan, H.; Wu, J.; Zhang, F.; Jin, Z.; Tian, J.; Xia, J.; Luo, J.; Yang, M. Zearalenone Exposure Disrupts STAT-ISG15 in Rat Colon: A Potential Linkage between Zearalenone and Inflammatory Bowel Disease. Toxins 2023, 15, 392. https://doi.org/10.3390/toxins15060392
Ruan H, Wu J, Zhang F, Jin Z, Tian J, Xia J, Luo J, Yang M. Zearalenone Exposure Disrupts STAT-ISG15 in Rat Colon: A Potential Linkage between Zearalenone and Inflammatory Bowel Disease. Toxins. 2023; 15(6):392. https://doi.org/10.3390/toxins15060392
Chicago/Turabian StyleRuan, Haonan, Jiashuo Wu, Fangqing Zhang, Ziyue Jin, Jiao Tian, Jing Xia, Jiaoyang Luo, and Meihua Yang. 2023. "Zearalenone Exposure Disrupts STAT-ISG15 in Rat Colon: A Potential Linkage between Zearalenone and Inflammatory Bowel Disease" Toxins 15, no. 6: 392. https://doi.org/10.3390/toxins15060392
APA StyleRuan, H., Wu, J., Zhang, F., Jin, Z., Tian, J., Xia, J., Luo, J., & Yang, M. (2023). Zearalenone Exposure Disrupts STAT-ISG15 in Rat Colon: A Potential Linkage between Zearalenone and Inflammatory Bowel Disease. Toxins, 15(6), 392. https://doi.org/10.3390/toxins15060392