Phylogeny-Related Variations in Venomics: A Test in a Subset of Habu Snakes (Protobothrops)


Abstract
1. Introduction
2. Results and Discussion
2.1. Chromatographic and Electrophoretic Profiles of Venoms from Six Habu Snakes
2.2. Proteomic Profiles of Venoms from Six Habu Snakes

{kind=link}
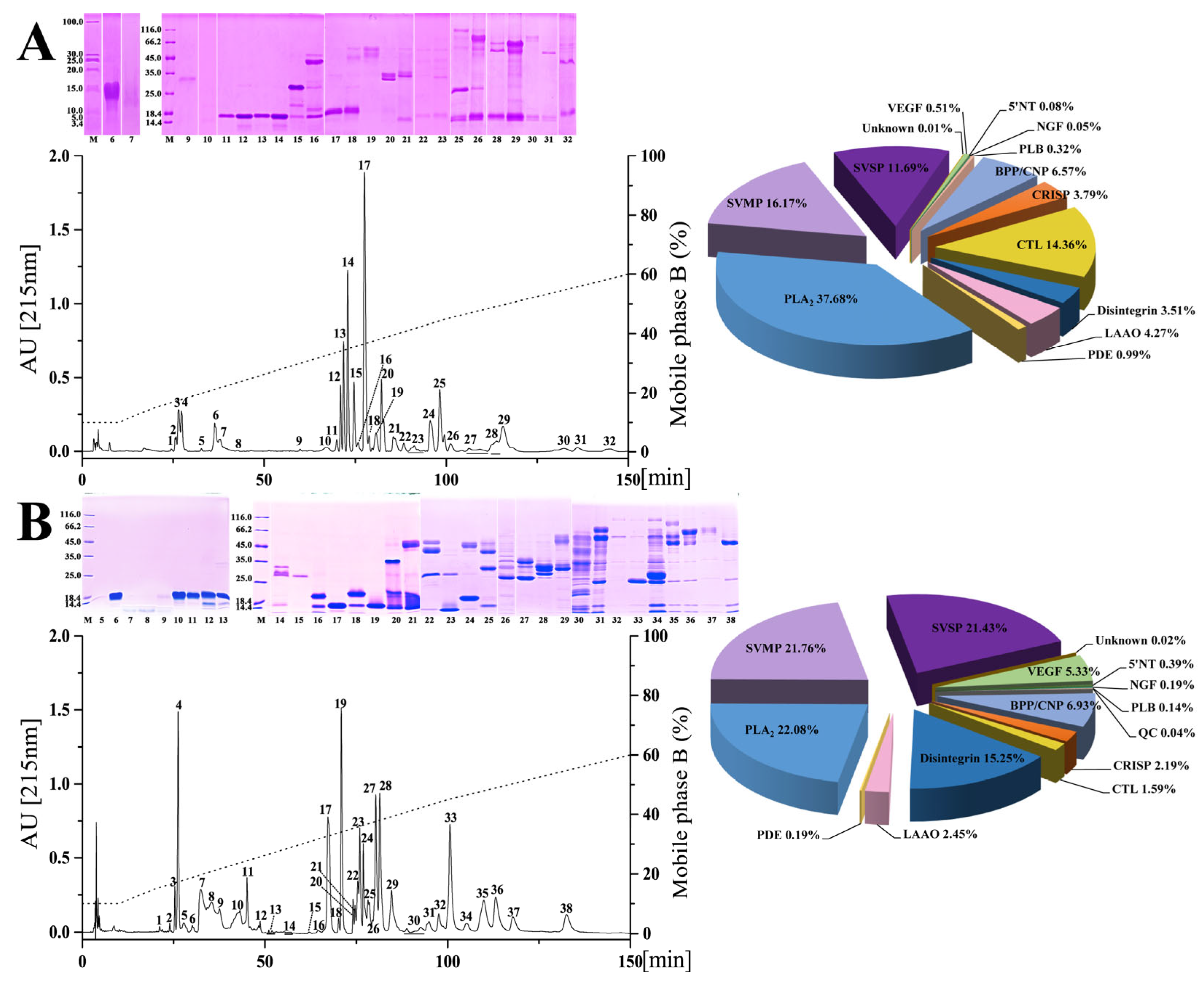{kind=link}
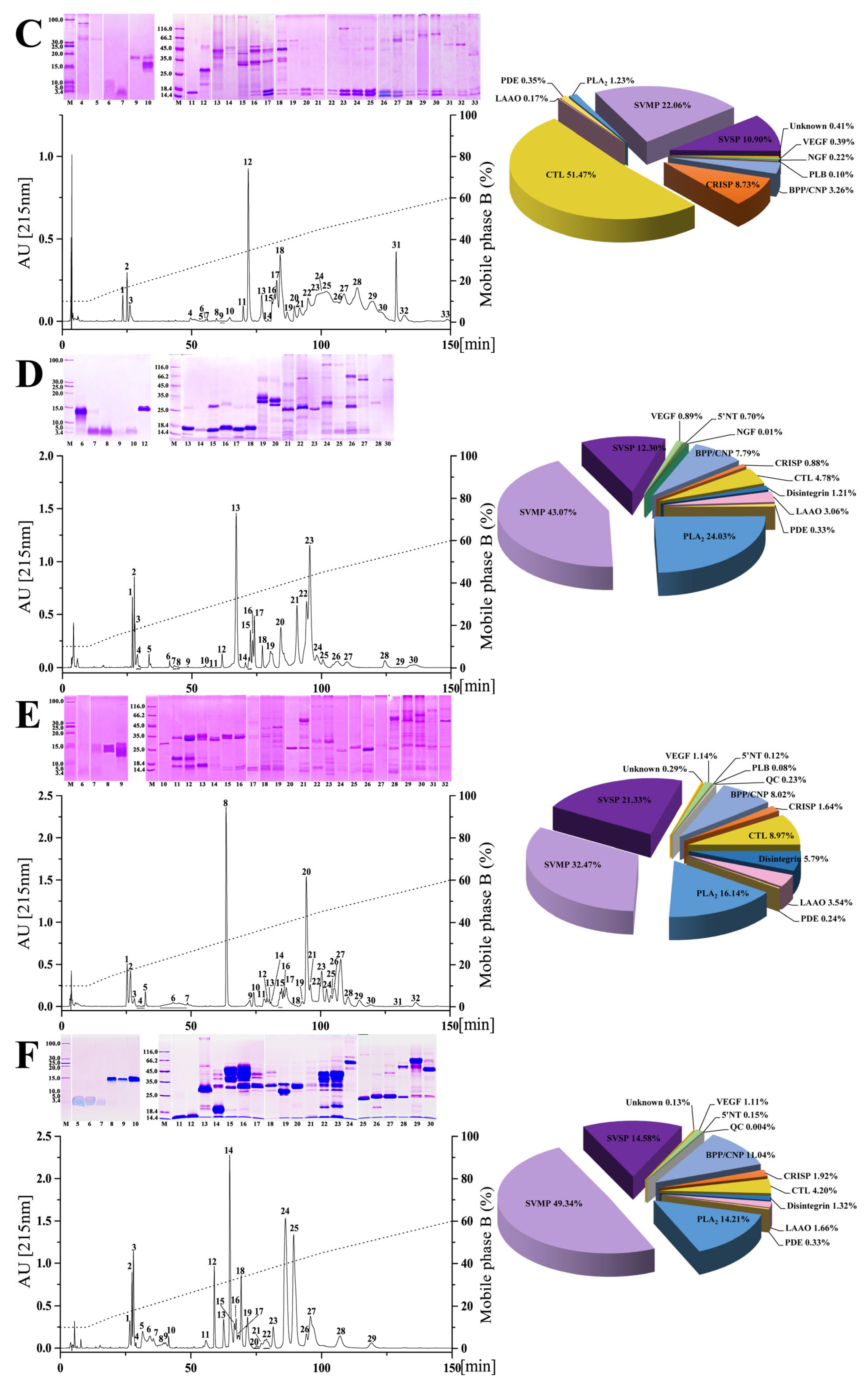{kind=link}
{kind=link}
{kind=link}
| Species | % of Total Venom Proteins | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|
| P. cornutus | P. jerdonii | P. mangshanensis | P. mucrosquamatus | P. sieversorum | P. xiangchengensis | P. elegans | P. flavoviridis | P. kelomohy | ||
| Location | Mainland China | Mainland China | Mainland China | Mainland China | China’s Taiwan | Vietnam | Mainland China | Japan | Japan | Thailand |
| Reference | Current study | Current study | Current study | Current study | Villalta et al. [30] | Current study | Current study | Aird et al. [33], Yamakawa et al. [53] | Huang et al. [54], Damm et al. [31] | Chanhome et al. [36] |
| LD50 (µg/g) | 0.61 (0.46–0.81) | 1.91 (1.73–2.11) | 1.23 (0.98–1.54) | 3.82 (3.50–4.17) | 2.17 | 1.23 (1.07–1.41) | 1.79 (1.51–2.11) | 4.85 | 2.60 | 0.67 (0.58–0.78) |
| Separation | RP-HPLC SDS-PAGE | RP-HPLC SDS-PAGE | RP-HPLC SDS-PAGE | RP-HPLC SDS-PAGE | RP-HPLC SDS-PAGE | RP-HPLC SDS-PAGE | RP-HPLC SDS-PAGE | - | RP-HPLC SDS-PAGE | SDS-PAGE |
| Digestion | In-gel | In-gel | In-gel | In-gel | In-gel | In-gel | In-gel | In-solution | In-gel | In-gel |
| MS | LC-MS/MS | MALDI-TOF LC-MS/MS | LC-MS/MS | LC-MS/MS | MALDI-TOF LC-MS/MS | LC-MS/MS | MALDI-TOF LC-MS/MS | LC-MS/MS | LC-MS/MS | LC-MS/MS |
| SVMP | 16.17 | 21.76 | 22.06 | 43.07 | 43.40 | 32.47 | 49.34 | 39.40 | 31.34 | 40.85 |
| SVSP | 11.69 | 21.43 | 10.90 | 12.30 | 10.40 | 21.33 | 14.58 | 9.50 | 1.44 | 29.93 |
| PLA2 | 37.68 | 22.10 | 1.23 | 24.51 | 23.50 | 16.14 | 14.21 | 22.50 | 55.14 | 15.49 |
| CRISP | 3.79 | 2.19 | 8.73 | 0.88 | 0.80 | 1.64 | 1.92 | 1.80 | 1.83 | 1.41 |
| Disintegrin | 3.51 | 15.25 | - | 1.21 | 0.80 | 5.79 | 1.32 | - | - | 0.35 |
| BPP/CNP | 6.57 | 6.93 | 3.26 | 7.79 | 3.60 | 8.02 | 11.04 | 0.10 | - | - |
| CTL | 14.36 | 1.59 | 51.47 | 4.78 | 3.90 | 8.97 | 4.20 | 2.90 | 2.78 | - |
| LAAO | 4.27 | 2.45 | 0.17 | 3.06 | 2.00 | 3.54 | 1.66 | 9.80 | 0.71 | 3.87 |
| VEGF | 0.51 | 5.33 | 0.39 | 0.89 | - | 1.14 | 1.11 | 1.80 | - | 0.70 |
| NGF | 0.05 | 0.19 | 0.22 | 0.01 | - | - | - | 0.50 | - | 1.76 |
| PDE | 0.99 | 0.19 | 1.06 | 0.80 | - | 0.24 | 0.33 | 4.10 | 0.07 | - |
| 5′NT | 0.08 | 0.39 | - | 0.70 | - | 0.12 | 0.15 | 2.80 | 0.02 | - |
| PLB | 0.32 | 0.14 | 0.10 | - | - | 0.08 | - | 2.20 | - | - |
| QC | - | 0.04 | - | - | - | 0.23 | 0.004 | 1.40 | - | - |
| Others | - | - | - | - | 3.30 | - | - | 1.00 | 6.39 | 5.63 |
| Unknown | 0.01 | 0.02 | 0.41 | - | - | 0.29 | 0.13 | 0.30 | 0.27 | - |
| P. cornutus | P. jerdonii | P. mucrosquamatus | P. sieversorum | P. xiangchengensis | |
|---|---|---|---|---|---|
| P. cornutus | - | 14.4 | 15.8 | 18.9 | 9.6 |
| P. jerdonii | - | - | 15.2 | 14.3 | 23.7 |
| P. mucrosquamatus | - | - | - | 39.2 | 18.0 |
| P. sieversorum | - | - | - | - | 7.6 |
| P. xiangchengensis | - | - | - | - | - |
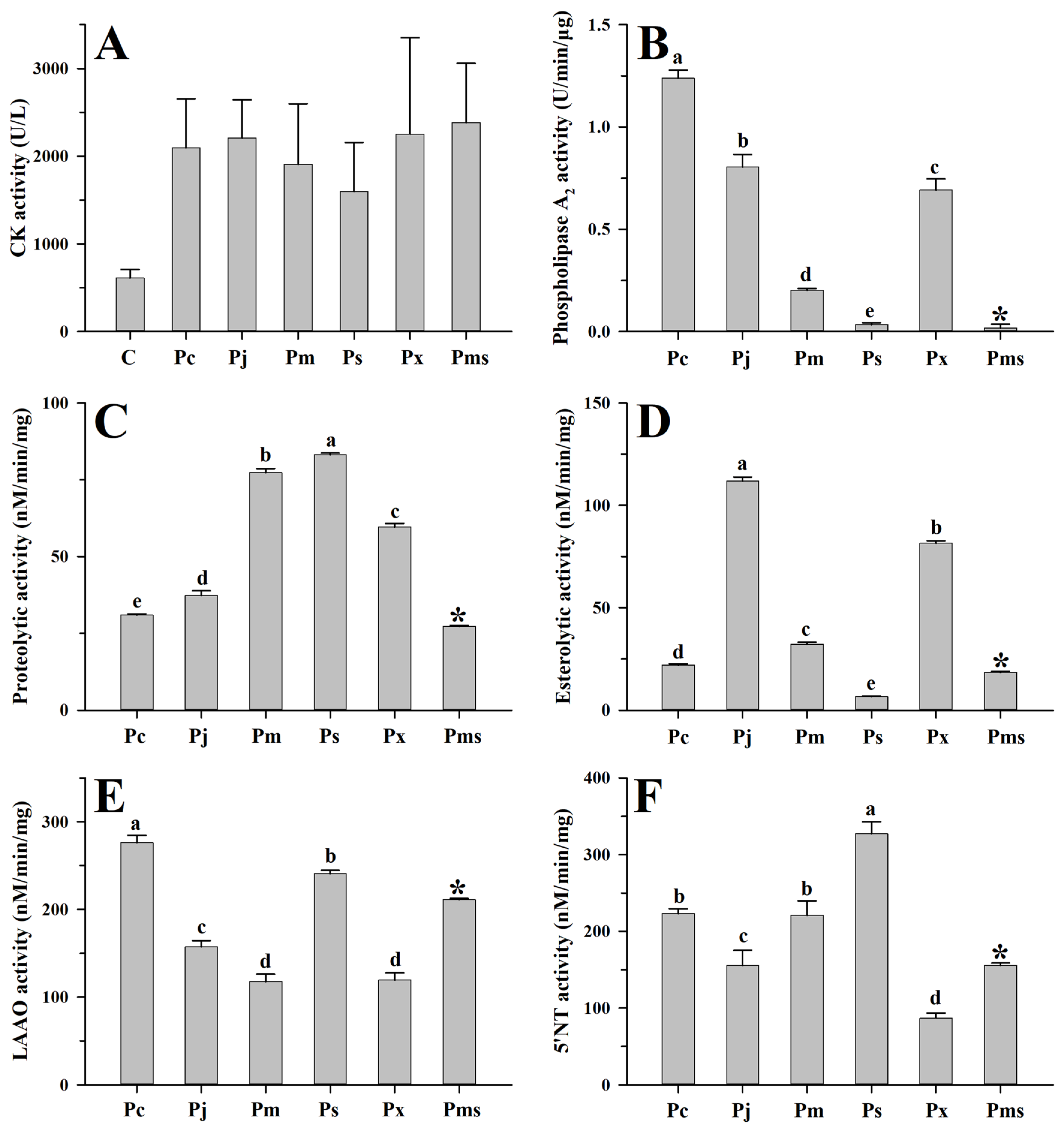
2.3. Toxicological and Enzymatic Activities of Venoms from Six Habu Snakes
2.4. Estimation of the Phylogenetic Signal of the Interspecific Variations in Venom Traits
3. Conclusions
4. Materials and Methods
4.1. Animals, Snake Venom Extraction, and Ethics
4.2. Proteomic Analyses
4.2.1. Reverse-Phase High-Performance Liquid Chromatography (RP-HPLC)
4.2.2. Sodium Dodecyl Sulfate-Polyacrylamide Gel Electrophoresis (SDS-PAGE)
4.2.3. In-Gel and In-Solution Tryptic Digestion
4.2.4. Mass Spectrometry (MS) Identification
4.2.5. Protein Relative Abundance and Similarity Estimation
4.3. Toxicological and Enzymatic Activities of Venoms
4.3.1. Lethality
4.3.2. Myotoxicity
4.3.3. Phospholipase A2 Activity
4.3.4. Proteolytic Activity
4.3.5. Esterolytic Activity
4.3.6. l-Amino Acid Oxidase Activity
4.3.7. 5′-Nucleotidase Activity
4.3.8. Statistical Analyses
4.4. Phylogeny-Based Comparative Analyses
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Calvete, J.J.; Escolano, J.; Sanz, L. Snake venomics of Bitis Species reveals large intragenus venom toxin composition variation: Application to taxonomy of congeneric taxa. J. Proteom. Res. 2007, 6, 2732–2745. [Google Scholar] [CrossRef] [PubMed]
- Casewell, N.R.; Wüster, W.; Vonk, F.J.; Harrison, R.A.; Fry, B.G. Complex cocktails: The evolutionary novelty of venoms. Trends Ecol. Evol. 2013, 28, 219–229. [Google Scholar] [CrossRef] [PubMed]
- Daltry, J.C.; Wuster, W.; Thorpe, R.S. Diet and snake venom evolution. Nature 1996, 379, 537–540. [Google Scholar] [CrossRef]
- Barlow, A.; Pook, C.E.; Harrison, R.A.; Wüster, W. Coevolution of diet and prey-specific venom activity supports the role of selection in snake venom evolution. Proc. R. Soc. B 2009, 276, 2443–2449. [Google Scholar] [CrossRef]
- Healy, K.; Carbone, C.; Jackson, A.L. Snake venom potency and yield are associated with prey-evolution, predator metabolism and habitat structure. Ecol. Lett. 2019, 22, 527–537. [Google Scholar] [CrossRef] [PubMed]
- Holding, M.L.; Strickland, J.L.; Rautsaw, R.M.; Hofmann, E.P.; Mason, A.J.; Hogan, M.P.; Nystrom, G.S.; Ellsworth, S.A.; Colston, T.J.; Borja, M.; et al. Phylogenetically diverse diets favor more complex venoms in North American pitvipers. Proc. Natl. Acad. Sci. USA 2021, 118, e2015579118. [Google Scholar] [CrossRef]
- Siqueira-Silva, T.; de Lima, L.A.G.; Chaves-Silveira, J.; Amado, T.F.; Naipauer, J.; Riul, P.; Martinez, P.A.; Sheard, C. Ecological and biogeographic processes drive the proteome evolution of snake venom. Glob. Ecol. Biogeogr. 2021, 30, 1978–1989. [Google Scholar] [CrossRef]
- Sasa, M. Diet and snake venom evolution: Can local selection alone explain intraspecific venom variation? Toxicon 1999, 37, 249–252. [Google Scholar]
- Mebs, D. Toxicity in animals. Trends in evolution? Toxicon 2001, 39, 87–96. [Google Scholar] [CrossRef]
- Gibbs, H.L.; Sanz, L.; Sovic, M.G.; Calvete, J.J. Phylogeny-based comparative analysis of venom proteome variation in a clade of rattlesnakes (Sistrurus sp.). PLoS ONE 2013, 8, e67220. [Google Scholar] [CrossRef]
- Lomonte, B.; Rey-Suárez, P.; Fernández, J.; Sasa, M.; Pla, D.; Vargas, N.; Bénard-Valle, M.; Sanz, L.; Corrêa-Netto, C.; Núñez, V.; et al. Venoms of Micrurus coral snakes: Evolutionary trends in compositional patterns emerging from proteomic analyses. Toxicon 2016, 122, 7–25. [Google Scholar] [CrossRef] [PubMed]
- Lomonte, B.; Tsai, W.C.; Ureña-Diaz, J.M.; Sanz, L.; Mora-Obando, D.; Sánchez, E.E.; Fry, B.G.; Gutiérrez, J.M.; Gibbs, H.L.; Sovic, M.G.; et al. Venomics of New World pit vipers: Genus-wide comparisons of venom proteomes across Agkistrodon. J. Proteom. 2014, 96, 103–116. [Google Scholar] [CrossRef]
- Aird, S.D.; Arora, J.; Barua, A.; Qiu, L.; Terada, K.; Mikheyev, A.S. Population genomic analysis of a pitviper reveals microevolutionary forces underlying venom chemistry. Genome Biol. Evol. 2017, 9, 2640–2649. [Google Scholar] [CrossRef] [PubMed]
- Qian, L.-F. Evolution of the Mitochondrial Genome Structure in Snake, with the Biogeography Analysis of Protobothrops. Ph.D. Thesis, Anhui Uniersity, Hefei, China, 2018. [Google Scholar]
- Gumprecht, A.; Tillack, F.; Orlov, N.L.; Captain, A.; Ryabov, S. Asian Pitvipers; GeitjeBooks: Berlin, Germany, 2004. [Google Scholar]
- Guo, P.; Malhotra, A.; Li, P.-P.; Pook, C.E.; Creer, S. New evidence on the phylogenetic position of the poorly known Asian pitviper Protobothrops kaulbacki (Serpentes: Viperidae: Crotalinae) with a redescription of the species and a revision of the genus Protobothrops. Herpetol. J. 2007, 17, 237–246. [Google Scholar]
- Orlov, N.; Ryabov, S.A.; Nguyen, T.T. Two new species of genera Protobothrops Hoge et Romano-Hoge, 1983 and Viridovipera Malhotra et Thorpe, 2004 (Ophidia: Viperidae: Crotalinae) from karst region in Northeastern Vietnam. Part I. Description of a new species of Protobothrops. Russ. J. Herpetol. 2009, 16, 69–82. [Google Scholar]
- Yang, J.-H.; Orlov, N.; Wang, Y.-Y. A new species of pitviper of the genus Protobothrops (Squamata: Viperidae) from China. Zootaxa 2011, 2936, 59–68. [Google Scholar] [CrossRef]
- Huang, X.; Pan, T.; Han, D.; Zhang, L.; Hou, Y.; Yu, L.; Zheng, H.; Zhang, B. A new species of the genus Protobothrops (Squamata: Viperidae: Crotalinae) from the Dabie Mountains, Anhui, China. Asian Herpetol. Res. 2012, 3, 213–218. [Google Scholar]
- Sumontha, M.; Vasaruchapong, T.; Chomngam, N.; Suntrarachun, S.; Pawangkhanant, P.; Sompan, W.; Smits, T.; Kunya, K.; Chanhome, L. Protobothrops kelomohy sp. nov. (Squamata Viperidae), the second known species of lance-headed pit viper from Thailand. Trop. Nat. Hist. 2020, 20, 43–59. [Google Scholar]
- Guo, P.; Liu, Q.; Wen, T.; Xiao, R.; Fang, M.; Zhong, G.-H.; Truong, N.Q.; Zhu, F.; Jadin, R.C.; Li, G. Multilocus phylogeny of the Asian Lance-headed pitvipers (Squamata, Viperidae, Protobothrops). Zootaxa 2016, 4093, 382–390. [Google Scholar] [CrossRef]
- Longbottom, J.; Shearer, F.M.; Devine, M.; Alcoba, G.; Chappuis, F.; Weiss, D.J.; Ray, S.E.; Ray, N.; Warrell, D.A.; Ruiz de Castañeda, R.; et al. Vulnerability to snakebite envenoming: A global mapping of hotspots. Lancet 2018, 392, 673–684. [Google Scholar] [CrossRef]
- World Health Organization. Guidelines for the production, control and regulation of snake antivenom immunoglobulins. In WHO Expert Committee on Biological Standardization Sixty-Seventh Report Annex 5 (WHO Technical Report Series, No 964); World Health Organization of the United Nations: Geneva, Switzerland, 2018. [Google Scholar]
- Yasunaga, H.; Horiguchi, H.; Kuwabara, K.; Hashimoto, H.; Matsuda, S. Short report: Venomous snake bites in Japan. Am. J. Trop. Med. Hyg. 2011, 84, 135–136. [Google Scholar] [CrossRef]
- Qin, G.-P. China Poisonous Snake Research; Guangxi Science and Technology Press: Nanning, China, 1998. [Google Scholar]
- Hifumi, T.; Sakai, A.; Kondo, Y.; Yamamoto, A.; Morine, N.; Ato, M.; Shibayama, K.; Umezawa, K.; Kiriu, N.; Kato, H.; et al. Venomous snake bites: Clinical diagnosis and treatment. J. Intensive Care 2015, 3, 16. [Google Scholar] [CrossRef] [PubMed]
- Yan, Z.-R.; Pan, J.-Q.; Hu, X.-J.; Zhang, S.-N.; Shen, S.-X. Epidemiological investigation of the poisonous snake bite around Nanchang area in recent two years. J. Snake 2020, 32, 19–22. [Google Scholar]
- Yang, K.; He, Y.-J.; Yang, S.-Q. Epidemiological investigation of 1167 cases of snake bite. J. Traum. Surg. 2021, 23, 539–541. [Google Scholar]
- Tomari, T.; Wakisaka, I.; Yanagihashi, T. Comparison of the epidemiological features of habu bites among Amamioshima, Tokunoshima and Okinawa. Jpn. J. Health Hum. Ecol. 1987, 53, 87–96. [Google Scholar] [CrossRef]
- Villalta, M.; Pla, D.; Yang, S.L.; Sanz, L.; Segura, A.; Vargas, M.; Chen, P.Y.; Herrera, M.; Estrada, R.; Cheng, Y.F.; et al. Snake venomics and antivenomics of Protobothrops mucrosquamatus and Viridovipera stejnegeri from Taiwan: Keys to understand the variable immune response in horses. J. Proteom. 2012, 75, 5628–5645. [Google Scholar] [CrossRef]
- Damm, M.; Hempel, B.F.; Nalbantsoy, A.; Süssmuth, R.D. Comprehensive snake venomics of the Okinawa habu pit viper, Protobothrops flavoviridis, by complementary mass spectrometry-guided approaches. Molecules 2018, 23, 1893. [Google Scholar] [CrossRef] [PubMed]
- Liu, C.-C.; Wu, C.-J.; Hsiao, Y.-C.; Yang, Y.-H.; Liu, K.-L.; Huang, G.-J.; Hsieh, C.-H.; Chen, C.-K.; Liaw, G.-W. Snake venom proteome of Protobothrops mucrosquamatus in Taiwan: Delaying venom-induced lethality in a rodent model by inhibition of phospholipase A2 activity with varespladib. J. Proteom. 2021, 234, 104084. [Google Scholar] [CrossRef]
- Aird, S.D.; Aggarwal, S.; Villar-Briones, A.; Tin, M.M.-Y.; Terada, K.; Mikheyev, A.S. Snake venoms are integrated systems, but abundant venom proteins evolve more rapidly. BMC Genom. 2015, 16, 647. [Google Scholar] [CrossRef]
- Aird, S.D.; Watanabe, Y.; Villar-Briones, A.; Roy, M.C.; Terada, K.; Mikheyev, A.S. Quantitative high-throughput profiling of snake venom gland transcriptomes and proteomes (Ovophis okinavensis and Protobothrops flavoviridis). BMC Genom. 2013, 14, 790. [Google Scholar] [CrossRef] [PubMed]
- Liu, C.-C.; Lin, C.-C.; Hsiao, Y.-C.; Wang, P.-J.; Yu, J.-S. Proteomic characterization of six Taiwanese snake venoms: Identification of species-specific proteins and development of a SISCAPA-MRM assay for cobra venom factors. J. Proteom. 2018, 187, 59–68. [Google Scholar] [CrossRef]
- Chanhome, L.; Khow, O.; Reamtong, O.; Vasaruchapong, T.; Laoungbua, P.; Tawan, T.; Suntrarachun, S.; Sitprija, S.; Kumkate, S.; Chaiyabutr, N. Biochemical and proteomic analyses of venom from a new pit viper, Protobothrops kelomohy. J. Venom. Anim. Toxins Incl. Trop. Dis. 2022, 28, e20210080. [Google Scholar] [CrossRef]
- Liu, Y.; Wang, X.; Wu, Z.; Chen, P.; Xiang, Y.; Nie, D.; Yi, J.; Yi, J.; Liu, Z. Electrophysiological and proteomic studies of Protobothrops mangshensis venom revealed its high bioactivities and toxicities. J. Integr. OMICS 2012, 2, 132–139. [Google Scholar]
- Calvete, J.J.; Lomonte, B.; Saviola, A.J.; Bonilla, F.; Sasa, M.; Williams, D.J.; Undheim, E.A.B.; Sunagar, K.; Jackson, T.N.W. Mutual enlightenment: A toolbox of concepts and methods for integrating evolutionary and clinical toxinology via snake venomics and the contextual stance. Toxicon X 2021, 9–10, 100070. [Google Scholar] [CrossRef]
- Zheng, S.-R.; Sun, Y.; Zhao, H.-Y.; Wen, L.; Ji, X.; Gao, J.-F. Differences between two groups of Burmese vipers (Viperidae: Azemiops) in the proteomic profiles, immunoreactivity and biochemical functions of their venoms. Toxins 2022, 14, 572. [Google Scholar] [CrossRef]
- Zhao, E.-M. Snakes of China; Anhui Science and Technology Publishing House: Hefei, China, 2006. [Google Scholar]
- Fry, B.G.; Wickramaratna, J.C.; Hodgson, W.C.; Alewood, P.F.; Kini, R.M.; Ho, H.; Wüster, W. Electrospray liquid chromatography/mass spectrometry fingerprinting of Acanthophis (death adder) venoms: Taxonomic and toxinological implications. Rapid Commun. Mass Spectrom. 2002, 16, 600–608. [Google Scholar] [CrossRef] [PubMed]
- Gao, J.-F.; Qu, Y.-F.; Zhang, X.-Q.; Ji, X. Within-clutch variation in venoms from hatchlings of Deinagkistrodon acutus (Viperidae). Toxicon 2011, 57, 970–977. [Google Scholar] [CrossRef] [PubMed]
- Angulo, Y.; Escolano, J.; Lomonte, B.; Gutiérrez, J.M.; Sanz, L.; Calvete, J.J. Snake venomics of central American pitvipers: Clues for rationalizing the distinct envenomation profiles of Atropoides nummifer and Atropoides picadoi. J. Proteom. Res. 2008, 7, 708–719. [Google Scholar] [CrossRef]
- von Reumont, B.M.; Anderluh, G.; Antunes, A.; Ayvazyan, N.; Beis, D.; Caliskan, F.; Crnković, A.; Damm, M.; Dutertre, S.; Ellgaard, L.; et al. Modern venomics-Current insights, novel methods, and future perspectives in biological and applied animal venom research. GigaScience 2022, 11, giac048. [Google Scholar]
- Tasoulis, T.; Isbister, G.K. A review and database of snake venom proteomes. Toxins 2017, 9, 290. [Google Scholar] [CrossRef]
- Calvete, J.J.; Borges, A.; Segura, Á.; Flores-Díaz, M.; Alape-Girón, A.; Gutiérrez, J.M.; Diez, N.; De Sousa, L.; Kiriakos, D.; Sánchez, E.; et al. Snake venomics and antivenomics of Bothrops colombiensis, a medically important pitviper of the Bothrops atrox-asper complex endemic to Venezuela: Contributing to its taxonomy and snakebite management. J. Proteom. 2009, 72, 227–240. [Google Scholar] [CrossRef]
- Tasoulis, T.; Pukala, T.L.; Isbister, G.K. Investigating toxin diversity and abundance in snake venom proteomes. Front. Pharmacol. 2021, 12, 768015. [Google Scholar] [CrossRef]
- Mebs, D.; Kuch, U.; Coronas, F.I.; Batista, C.V.; Gumprecht, A.; Possani, L.D. Biochemical and biological activities of the venom of the Chinese pitviper Zhaoermia mangshanensis, with the complete amino acid sequence and phylogenetic analysis of a novel Arg49 phospholipase A2 myotoxin. Toxicon 2006, 47, 797–811. [Google Scholar] [CrossRef]
- Murakami, M.T.; Kuch, U.; Betzel, C.; Mebs, D.; Arni, R.K. Crystal structure of a novel myotoxic Arg49 phospholipase A2 homolog (zhaoermiatoxin) from Zhaoermia mangshanensis snake venom: Insights into Arg49 coordination and the role of Lys122 in the polarization of the C-terminus. Toxicon 2008, 51, 723–735. [Google Scholar] [CrossRef] [PubMed]
- Tsai, I.-H.; Lan, H.; Chen, Y.-H.; Wang, Y.-M. Proteomic analyses on the venom of an individual Protobothrops mangshanesis. J. Snake 2010, 22, 325–331. [Google Scholar]
- Nie, D.-S.; Liu, Y.; Wu, Z.; Liu, L.-C.; Zhu, Z.-B. Analysis of two-dimensional electrophoresis of toxin from the Ermia mangshanensis. J. Hunan Inst. Sci. Technol. (Nat. Sci.) 2011, 24, 48–51. [Google Scholar]
- Margres, M.J.; Wray, K.P.; Seavy, M.; McGivern, J.J.; Herrera, N.D.; Rokyta, D.R. Expression differentiation is constrained to low-expression proteins over ecological timescales. Genetics 2016, 202, 273–283. [Google Scholar] [CrossRef]
- Yamakawa, M.; Nozaky, M.; Hokama, Z. Fractionation of sakishimahabu (Trimeresurus elegans) venom and lethal, hemorrhagic and edema-forming activities of the fractions. In Animal, Plant and Microbial Toxins; Ohsaka, A., Hayashi, K., Sawai, Y., Eds.; Plenum Press: New York, NY, USA, 1976; Volume 1, pp. 97–109. [Google Scholar]
- Huang, S.; Huang, J.-T. An anti-hemorrhagic factor in Zaocys dhumnades serum: A promising source for antivenom drug. Acta Zool. Sin. 2006, 52, 1113–1118. [Google Scholar]
- Gutiérrez, J.M.; Calvete, J.J.; Habib, A.G.; Harrison, R.A.; Williams, D.J.; Warrell, D.A. Snakebite envenoming. Nat. Rev. Dis. Prim. 2017, 3, 17063. [Google Scholar] [CrossRef] [PubMed]
- Gutiérrez, J.M.; Lomonte, B. Phospholipases A2: Unveiling the secrets of a functionally versatile group of snake venom toxins. Toxicon 2013, 62, 27–39. [Google Scholar] [CrossRef]
- Gutiérrez, J.M.; Ownby, C.L. Skeletal muscle degeneration induced by venom phospholipases A2: Insights into the mechanisms of local and systemic myotoxicity. Toxicon 2003, 42, 915–931. [Google Scholar] [CrossRef]
- Yu, P.-N.; Zhong, H.; Yu, Y.-Z.; Yu, Y.-Z. Zhongguo Sheshang Xue; Shanxi Science and Technology Publishing House: Taiyuan, China, 2009. [Google Scholar]
- Ogawa, T.; Tobishima, Y.; Kamata, S.; Matsuda, Y.; Muramoto, K.; Hidaka, M.; Futai, E.; Kuraishi, T.; Yokota, S.; Ohno, M.; et al. Focused proteomics analysis of habu snake (Protobothrops flavoviridis) venom using antivenom-based affinity chromatography reveals novel myonecrosis-enhancing activity of thrombin-like serine proteases. Front. Pharmacol. 2021, 12, 766406. [Google Scholar] [CrossRef]
- Chiang, L.-C.; Chien, K.-Y.; Su, H.-Y.; Chen, Y.-C.; Mao, Y.-C.; Wu, W.-G. Comparison of protein variation in Protobothrops mucrosquamatus venom between Northern and Southeast Taiwan and association with human envenoming effects. Toxins 2022, 14, 643. [Google Scholar] [CrossRef]
- Serrano, S.M.T. The long road of research on snake venom serine proteinases. Toxicon 2013, 62, 19–26. [Google Scholar] [CrossRef] [PubMed]
- Gao, J.-F.; Wang, J.; He, Y.; Qu, Y.-F.; Lin, L.-H.; Ma, X.-M.; Ji, X. Proteomic and biochemical analyses of short-tailed pit viper (Gloydius brevicaudus) venom: Age-related variation and composition–activity correlation. J. Proteom. 2014, 105, 307–322. [Google Scholar] [CrossRef] [PubMed]
- Lin, C.-C.; Shih, C.-P.; Wang, C.-C.; Ouyang, C.-H.; Liu, C.-C.; Yu, J.-S.; Lo, C.-H. The clinical usefulness of Taiwan bivalent freeze-dried hemorrhagic antivenom in Protobothrops mucrosquamatus- and Viridovipera stejnegeri-envenomed patients. Toxins 2022, 14, 794. [Google Scholar] [CrossRef] [PubMed]
- Lomonte, B.; Fernández, J.; Sanz, L.; Angulo, Y.; Sasa, M.; Gutiérrez, J.M.; Calvete, J.J. Venomous snakes of Costa Rica: Biological and medical implications of their venom proteomic profiles analyzed through the strategy of snake venomics. J. Proteom. 2014, 105, 323–339. [Google Scholar] [CrossRef]
- Souza, G.H.M.F.; Catharino, R.R.; Ifa, D.R.; Eberlin, M.N.; Hyslop, S. Peptide fingerprinting of snake venoms by direct infusion nano-electrospray ionization mass spectrometry: Potential use in venom identification and taxonomy. J. Mass Spectrom. 2008, 43, 594–599. [Google Scholar] [CrossRef]
- Zhao, E.-M.; Wu, G.-F.; Wu, X.-F.; Chen, Y.-C.; Jiang, M.-S.; Zhang, J.-K.; Hsu, K. A comparison of snake venom eletrophoretogram of Agkistrodon species in China and their value in snake taxonomy. Acta Zool. Sin. 1981, 27, 213–217+302. [Google Scholar]
- Blomberg, S.P.; Garland, T., Jr.; Ives, A.R. Testing for phylogenetic signal in comparative data: Behavioral traits are more labile. Evolution 2003, 57, 717–745. [Google Scholar]
- Guo, P.; Liu, Q.; Wu, Y.-Y.; Zhu, F.; Zhong, G.-H. Pitvipers of China; Science Press: Beijing, China, 2022. [Google Scholar]
- Sousa, L.F.; Holding, M.L.; Del-Rei, T.H.M.; Rocha, M.M.T.; Mourao, R.H.V.; Chalkidis, H.M.; Prezoto, B.; Gibbs, H.L.; Moura-da-Silva, A.M. Individual variability in Bothrops atrox snakes collected from different habitats in the Brazilian Amazon: New findings on venom composition and functionality. Toxins 2021, 13, 814. [Google Scholar] [CrossRef]
- Bradford, M.M. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal. Biochem. 1976, 72, 248–254. [Google Scholar] [CrossRef] [PubMed]
- He, Y.; Gao, J.-F.; Lin, L.-H.; Ma, X.-M.; Ji, X. Age-related variation in snake venom: Evidence from two snakes (Naja atra and Deinagkistrodon acutus) in southeastern China. Asian Herpetol. Res. 2014, 5, 119–127. [Google Scholar]
- Tu, A.T.; James, G.P.; Chua, A. Some biochemical evidence in support of the classification of venomous snakes. Toxicon 1965, 3, 5–8. [Google Scholar] [CrossRef] [PubMed]
- Toyama, M.H.; Toyama, D.d.O.; Passero, L.F.D.; Laurenti, M.D.; Corbett, C.E.; Tomokane, T.Y.; Fonseca, F.V.; Antunes, E.; Joazeiro, P.P.; Beriam, L.O.S.; et al. Isolation of a new L-amino acid oxidase from Crotalus durissus cascavella venom. Toxicon 2006, 47, 47–57. [Google Scholar] [CrossRef]
- Dhananjaya, B.L.; Nataraju, A.; Rajesh, R.; Raghavendra Gowda, C.D.; Sharath, B.K.; Vishwanath, B.S.; D’Souza, C.J.M. Anticoagulant effect of Naja naja venom 5′nucleotidase: Demonstration through the use of novel specific inhibitor, vanillic acid. Toxicon 2006, 48, 411–421. [Google Scholar] [CrossRef]
- Wen, L. Interspecific Variations in Venom Proteomic and Venom-Gland Transcriptomic Profiles across the Genus Gloydius. Master’s Thesis, Hangzhou Normal University, Hangzhou, China, 2020. [Google Scholar]
- Li, J.-N.; Liang, D.; Wang, Y.-Y.; Guo, P.; Huang, S.; Zhang, P. A large-scale systematic framework of Chinese snakes based on a unified multilocus marker system. Mol. Phylogenet. Evol. 2020, 148, 106807. [Google Scholar] [CrossRef]
- Lanfear, R.; Calcott, B.; Ho, S.Y.W.; Guindon, S. PartitionFinder: Combined selection of partitioning schemes and substitution models for phylogenetic analyses. Mol. Biol. Evol. 2012, 29, 1695–1701. [Google Scholar] [CrossRef]
- Bouckaert, R.; Heled, J.; Kühnert, D.; Vaughan, T.; Wu, C.-H.; Xie, D.; Suchard, M.A.; Rambaut, A.; Drummond, A.J. BEAST 2: A software platform for Bayesian evolutionary analysis. PLoS Comput. Biol. 2014, 10, e1003537. [Google Scholar] [CrossRef]
- Winemiller, K.O.; Pianka, E.R. Organization in natural assemblages of desert lizards and tropical fishes. Ecol. Monogr. 1990, 60, 27–55. [Google Scholar] [CrossRef]
- Kembel, S.W.; Cowan, P.D.; Helmus, M.R.; Cornwell, W.K.; Morlon, H.; Ackerly, D.D.; Blomberg, S.P.; Webb, C.O. Picante: R tools for integrating phylogenies and ecology. Bioinformatics 2010, 26, 1463–1464. [Google Scholar] [CrossRef] [PubMed]





| Venom Trait | K-Value | p-Value |
|---|---|---|
| SVMP | 0.566 | 0.405 |
| SVSP | 0.932 | 0.026 |
| PLA2 | 0.728 | 0.158 |
| CRISP | 0.789 | 0.153 |
| Disintegrin | 0.324 | 0.694 |
| BPP/CNP | 0.474 | 0.422 |
| CTL | 0.628 | 0.303 |
| LAAO | 0.499 | 0.504 |
| Venom breadth | 0.472 | 0.492 |
| LD50 | 0.621 | 0.311 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhao, H.-Y.; He, N.; Sun, Y.; Wang, Y.-C.; Zhang, H.-B.; Chen, H.-H.; Zhang, Y.-Q.; Gao, J.-F. Phylogeny-Related Variations in Venomics: A Test in a Subset of Habu Snakes (Protobothrops). Toxins 2023, 15, 350. https://doi.org/10.3390/toxins15050350
Zhao H-Y, He N, Sun Y, Wang Y-C, Zhang H-B, Chen H-H, Zhang Y-Q, Gao J-F. Phylogeny-Related Variations in Venomics: A Test in a Subset of Habu Snakes (Protobothrops). Toxins. 2023; 15(5):350. https://doi.org/10.3390/toxins15050350
Chicago/Turabian StyleZhao, Hong-Yan, Na He, Yan Sun, Yong-Chen Wang, Hao-Bing Zhang, Hui-Hui Chen, Ya-Qi Zhang, and Jian-Fang Gao. 2023. "Phylogeny-Related Variations in Venomics: A Test in a Subset of Habu Snakes (Protobothrops)" Toxins 15, no. 5: 350. https://doi.org/10.3390/toxins15050350
APA StyleZhao, H.-Y., He, N., Sun, Y., Wang, Y.-C., Zhang, H.-B., Chen, H.-H., Zhang, Y.-Q., & Gao, J.-F. (2023). Phylogeny-Related Variations in Venomics: A Test in a Subset of Habu Snakes (Protobothrops). Toxins, 15(5), 350. https://doi.org/10.3390/toxins15050350