Model of the Origin of a Ciguatoxic Grouper (Plectropomus leopardus)


Abstract
1. Introduction
2. Results and Discussion
2.1. Framework for Model Development
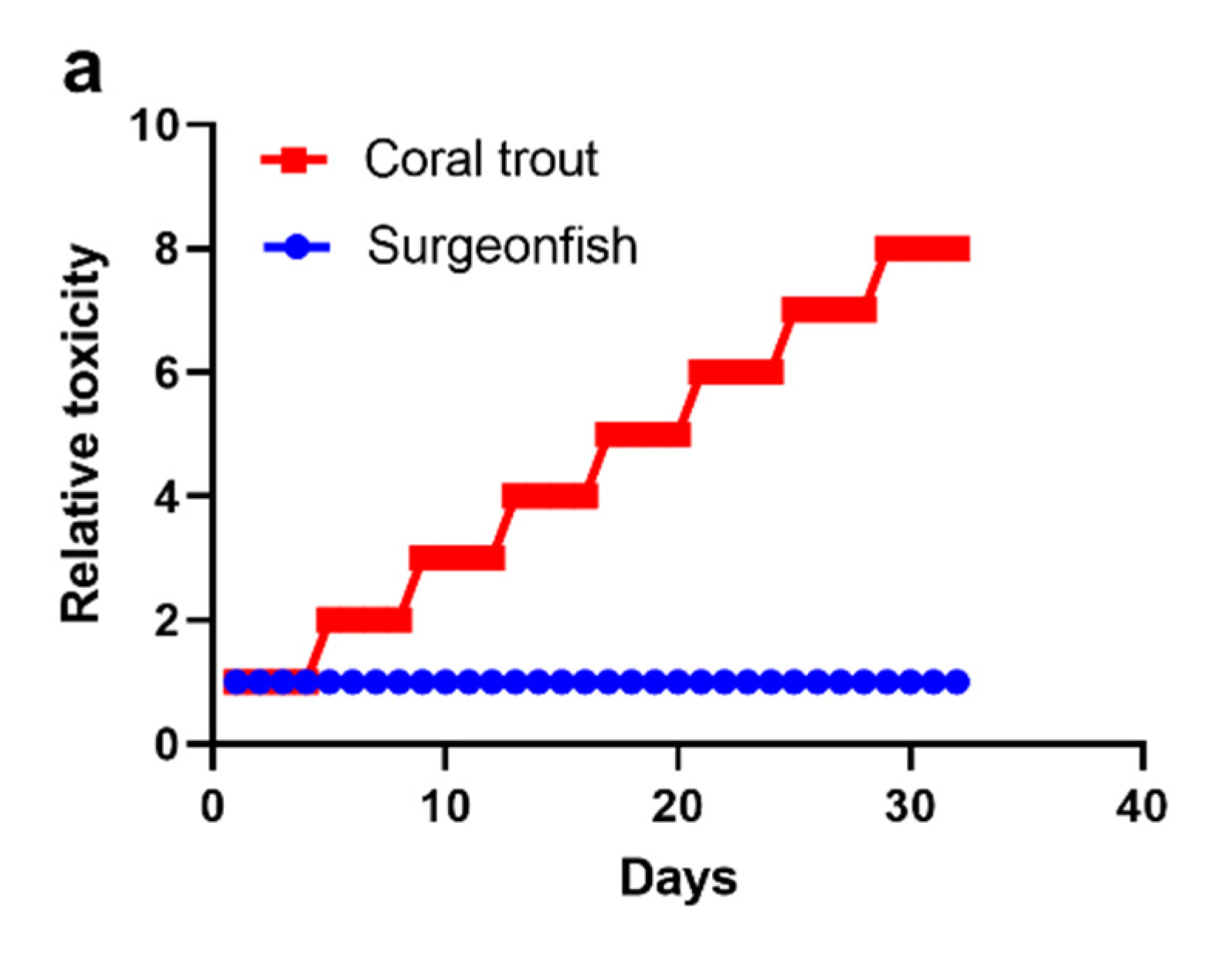
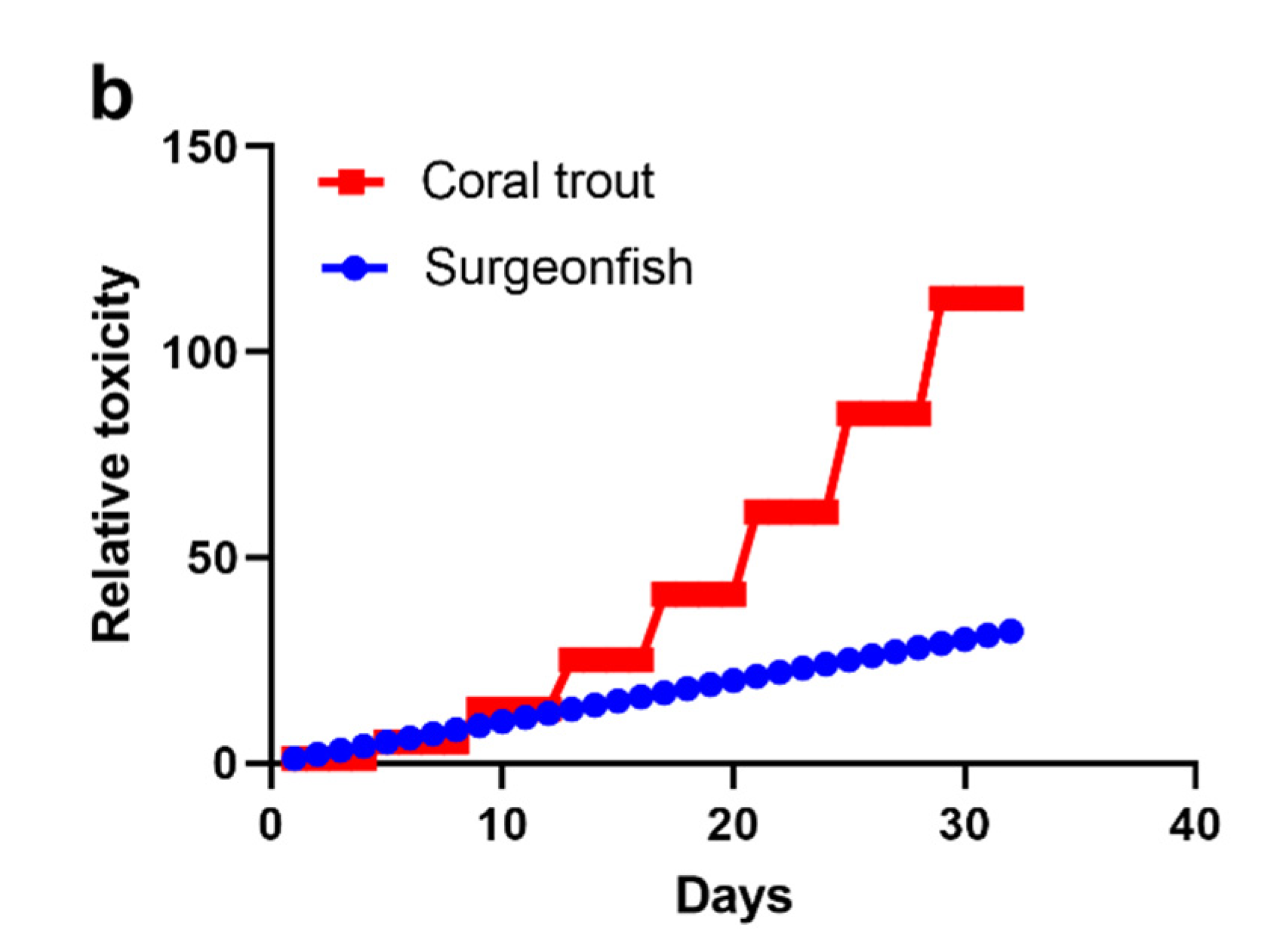
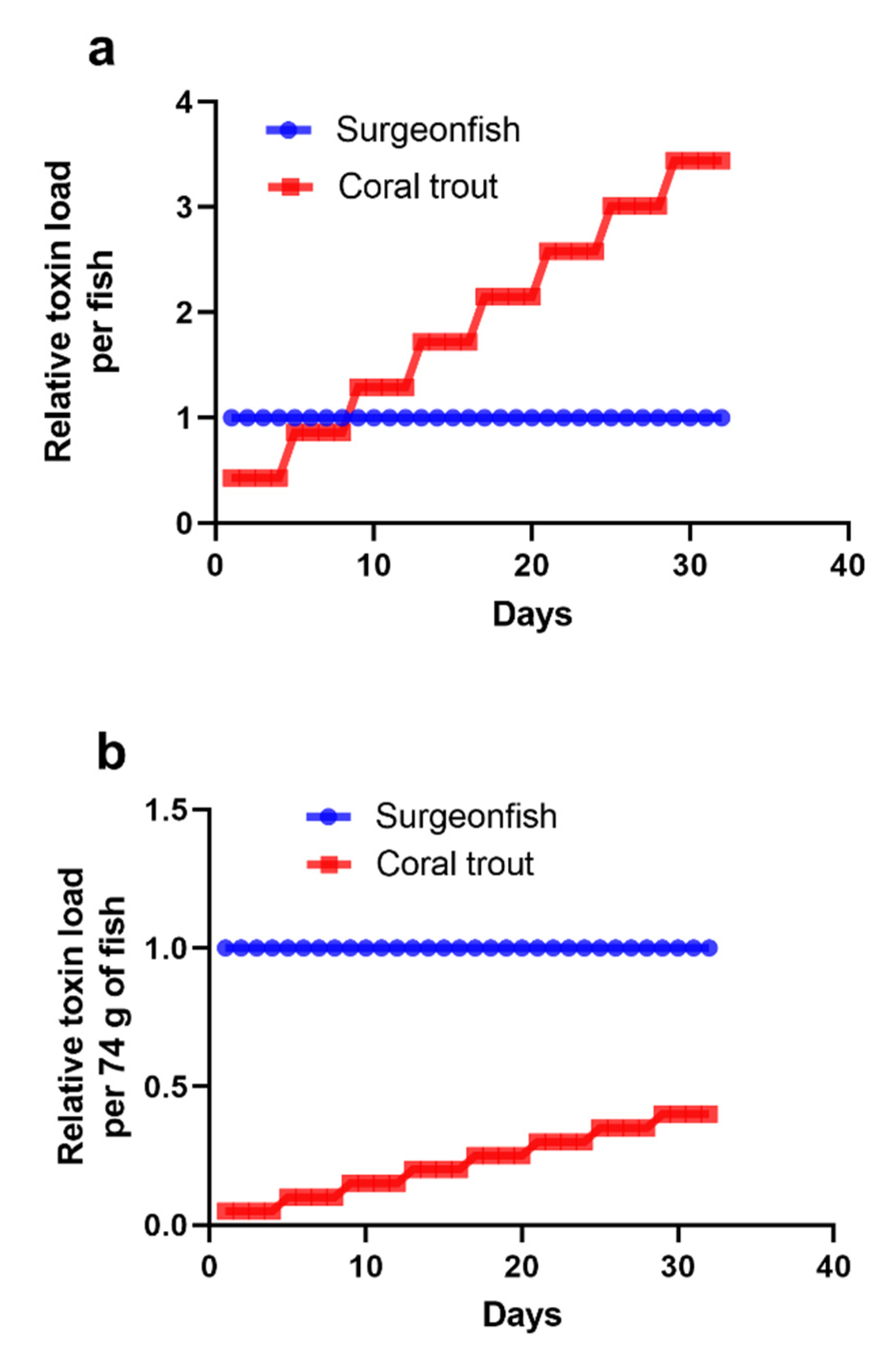
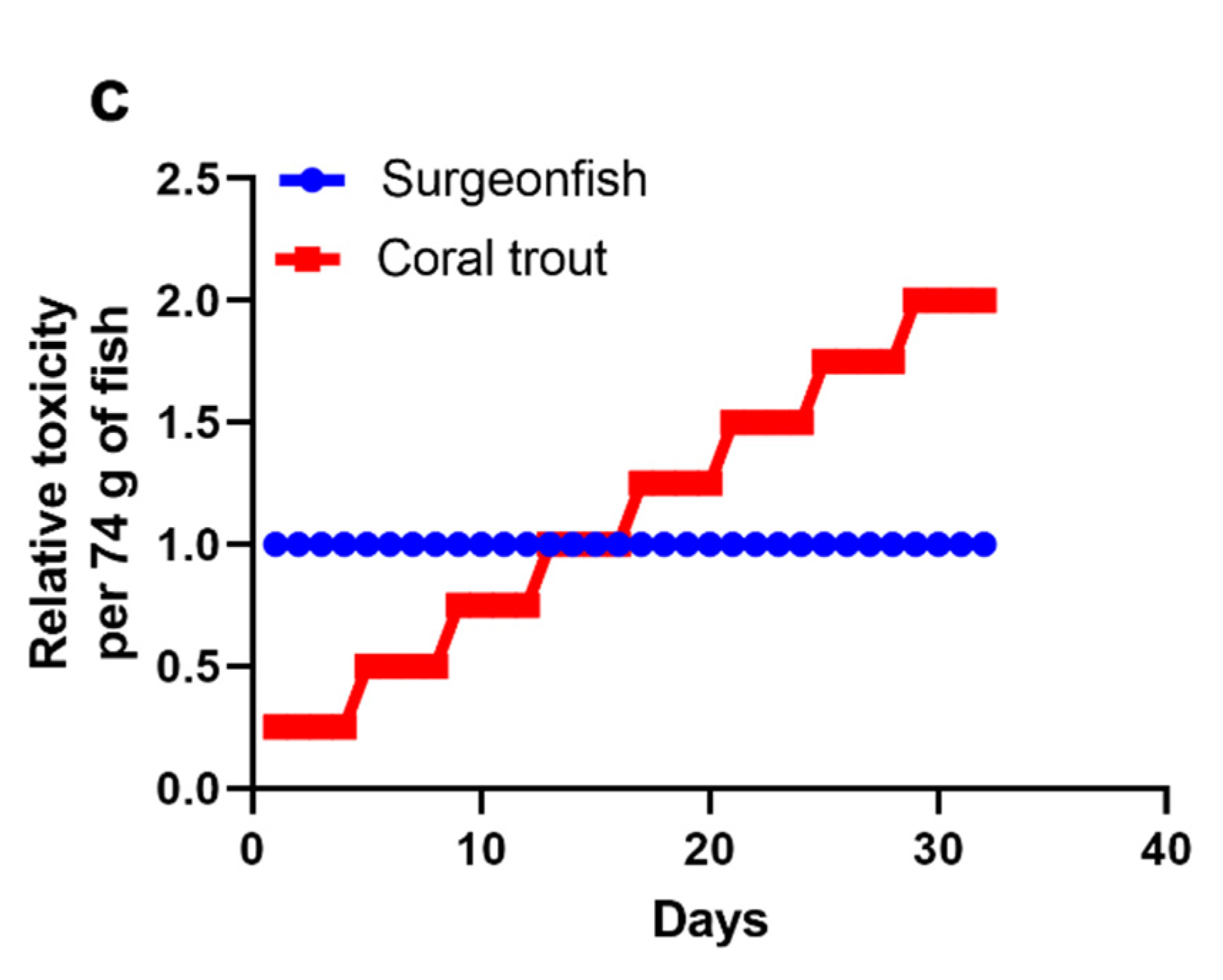
2.2. Scenario 1: 1000 Gambierdiscus/cm2 Epiphytic upon Turf Algae
2.3. Scenario 2: ≤10 Gambierdiscus/cm2 Epiphytic upon Turf Algae
2.4. Scenario 3: 100 Gambierdiscus/cm2 Epiphytic upon Turf Algae
2.5. Accumulation of CTX into Common Coral Trout (P. leopardus)
2.6. The Minimum CTX Concentration in Gambierdiscus to Produce a Ciguateric Grouper
3. Conclusions
4. Materials and Methods
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Gillespie, N.C.; Lewis, R.J.; Pearn, J.H.; Bourke, A.T.C.; Holmes, M.J.; Bourke, J.B.; Shields, W.J. Ciguatera in Australia: Occurrence, clinical features, pathophysiology and management. Med. J. Aust. 1986, 145, 584–590. [Google Scholar] [CrossRef] [PubMed]
- Tonge, J.I.; Battey, Y.; Forbes, J.J.; Grant, E.M. Ciguatera poisoning: A report of two outbreaks and a probable fatal case in Queensland. Med. J. Aust. 1967, 2, 1088–1090. [Google Scholar] [CrossRef]
- Gillespie, N.C. Ciguatera poisoning. In Toxic Plants and Animals, A Guide for Australia; Covacevich, J., Davie, P., Pearn, J., Eds.; Queensland Museum: Brisbane, Australia, 1987; pp. 161–169. [Google Scholar]
- Lewis, R.J. Ciguatera: Australian perspectives on a global problem. Toxicon 2006, 48, 799–809. [Google Scholar] [CrossRef] [PubMed]
- Paradice, W.E.J. Injuries and lesions caused by the bites of animals and insects. Med. J. Aust. 1924, 2, 650–652. [Google Scholar] [CrossRef]
- Whitley, G.P. Ichthyological miscellanea. Mem. Qld. Mus. 1930, 10, 8–31. [Google Scholar]
- Whitley, G.P. A new fish, reputed to be poisonous, from Queensland. Mem. Qld. Mus. 1934, 10, 175–179. [Google Scholar]
- Fenner, P.J.; Lewis, R.J.; Williamson, J.A.; Williams, M.L. A Queensland family with ciguatera after eating coral trout. Med. J. Aust. 1997, 166, 473–475. [Google Scholar] [CrossRef]
- Leigh, G.M.; Campbell, A.B.; Lunow, C.P.; O’Neill, M.F. Stock Assessment of the Queensland East Coast Common Coral Trout (Plectropomus leopardus) Fishery; Queensland Department of Agriculture, Fisheries and Forestry: Brisbane, Australia, 2014; p. 113.
- Campbell, A.; Leigh, G.; Bessell-Browne, P.; Lovett, R. Stock Assessment of the Queensland East Coast Common Coral Trout (Plectropomus leopardus) Fishery; Department of Employment, Economic Development and Innovation, Queensland Government: Brisbane, Australia, 2019; p. 61.
- Campbell, A.B.; Northrop, A.R. Stock Assessment of the Common Coral Trout (Plectropomus leopardus) in Queensland, Australia; Department of Employment, Economic Development and Innovation, Queensland Government: Brisbane, Australia, 2020; p. 45.
- FishBase; Froese, R.; Pauly, D. (Eds.). FishBase. World Wide Web Electronic Publication. Version (08/2022). Available online: www.fishbase.org (accessed on 16 January 2023).
- Holmes, M.J.; Venables, B.; Lewis, R.J. Critical review and conceptual and quantitative models for the transfer and depuration of ciguatoxins in fishes. Toxins 2021, 13, 515. [Google Scholar] [CrossRef]
- FAO; WHO. Report of the Expert Meeting on Ciguatera Poisoning: Rome, 19–23 November 2018; Food Safety and Quality No. 9; Rome, Italy. 2020. Available online: https://books.google.com.sg/books?hl=zh-TW&lr=&id=HhvtDwAAQBAJ&oi=fnd&pg=PR6&dq=10.4060/ca8817en&ots=UUfascmP-s&sig=UbOO36DHmBc8aCkBNQpXUxfuHdk#v=onepage&q=10.4060%2Fca8817en&f=false (accessed on 16 March 2023). [CrossRef]
- Lewis, R.J.; Sellin, M.; Poli, M.A.; Norton, R.S.; MacLeod, J.K.; Sheil, M.A. Purification and characterization of ciguatoxins from moray eel (Lycodontis javanicus, Muraenidae). Toxicon 1991, 29, 1115–1127. [Google Scholar] [CrossRef]
- Lewis, R.J.; Sellin, M. Multiple ciguatoxins in the flesh of fishes. Toxicon 1992, 30, 915–919. [Google Scholar] [CrossRef]
- Chinain, M.; Darius, H.T.; Ung, A.; Cruchet, P.; Wang, Z.; Ponton, D.; Laurent, D.; Pauillac, S. Growth and toxin production in the ciguatera-causing dinoflagellate Gambierdiscus polynesiensis (Dinophyceae) in culture. Toxicon 2010, 56, 739–750. [Google Scholar] [CrossRef] [PubMed]
- Fraga, S.; Rodíguez, F.; Caillaud, A.; Diogène, J.; Raho, N.; Zapata, M. Gambierdiscus excentricus sp. nov. (Dinophyceae), a benthic toxic dinoflagellate from the Canary Islands (NE Atlantic Ocean). Harmful Algae 2011, 11, 10–22. [Google Scholar] [CrossRef]
- Pisapia, F.; Holland, W.C.; Hardison, D.R.; Litaker, R.W.; Fraga, S.; Nishimura, T.; Adachi, M.; Nguyen-Ngoc, L.; Séchet, V.; Amzil, Z.; et al. Toxicity screening of 13 Gambierdiscus strains using neuro-2a and erythrocyte lysis bioassays. Harmful Algae 2017, 63, 173–183. [Google Scholar] [CrossRef] [PubMed]
- Litaker, R.W.; Holland, W.C.; Hardison, D.H.; Pisapia, F.; Hess, P.; Kibler, S.R.; Tester, P.A. Ciguatoxicity of Gambierdiscus and Fukuyoa species from the Caribbean and Gulf of Mexico. PLoS ONE 2017, 12, 173–183. [Google Scholar] [CrossRef] [PubMed]
- Gaiani, G.; Leonardo, S.; Tudó, À.; Toldrà, A.; Rey, M.; Andree, K.B.; Tsumuraya, T.; Hirama, M.; Diogène, J.; O’Sullivan, C.K.; et al. Rapid detection of ciguatoxins in Gambierdiscus and Fukuyoa with immunosensing tools. Ecotoxicoloy Environ. Saf. 2020, 204, 111004. [Google Scholar] [CrossRef] [PubMed]
- Holmes, M.J.; Lewis, R.J.; Poli, M.A.; Gillespie, N.C. Strain dependent production of ciguatoxin precursors (gambiertoxins) by Gambierdiscus toxicus (Dinophyceae) in culture. Toxicon 1991, 29, 761–775. [Google Scholar] [CrossRef] [PubMed]
- Larsson, M.E.; Laczka, O.F.; Harwood, D.T.; Lewis, R.J.; Himaya, S.W.A.; Murray, S.A.; Doblin, M.A. Toxicology of Gambierdiscus spp. (Dinophyceae) from tropical and temperate Australian waters. Mar. Drugs 2018, 16, 7. [Google Scholar] [CrossRef]
- Satake, M.; Ishibashi, Y.; Legrand, A.M.; Yasumoto, T. Isolation and structure of ciguatoxin-4A, a new ciguatoxin precursor, from cultures of dinoflagellate Gambierdiscus toxicus and parrotfish Scarus gibbus. Biosci. Biotechnol. Biochem. 1997, 60, 2103–2105. [Google Scholar] [CrossRef]
- Yasumoto, T.; Igarashi, T.; Legrand, A.-M.; Cruchet, P.; Chinain, M.; Fujita, T.; Naoki, H. Structural elucidation of ciguatoxin congeners by fast-atom bombardment tandem mass spectrometry. J. Am. Chem. Soc. 2000, 122, 4988–4989. [Google Scholar] [CrossRef]
- Ikehara, T.; Kuniyoshi, K.; Oshiro, N.; Yasumoto, T. Biooxidation of ciguatoxins leads to species-specific toxin profiles. Toxins 2017, 9, 205. [Google Scholar] [CrossRef]
- Hamilton, B.; Whittle, N.; Shaw, G.; Eaglesham, G.; Moore, M.R.; Lewis, R.J. Human fatality associated with Pacific ciguatoxin contaminated fish. Toxicon 2010, 56, 668–673. [Google Scholar] [CrossRef] [PubMed]
- Kohli, G.S.; Haslauer, K.; Sarowar, C.; Kretzschmar, A.L.; Boulter, M.; Harwood, D.T.; Laczka, O.; Murray, S.M. Qualitative and quantitative assessment of the presence of ciguatoxin, P-CTX-1b, in Spanish mackerel (Scomberomorus commerson) from waters in New South Wales (Australia). Toxicol. Rep. 2017, 4, 328–334. [Google Scholar] [CrossRef] [PubMed]
- Lewis, R.J.; Holmes, M.J. Origin and transfer of toxins involved in ciguatera. Comp. Biochem. Physiol. 1993, 106C, 615–628. [Google Scholar] [CrossRef] [PubMed]
- Munday, R.; Murray, S.; Rhodes, L.L.; Larsson, M.E.; Harwood, D.T. Ciguatoxins and maitotoxins in extracts of sixteen Gambierdiscus isolates and one Fukuyoa isolate from the South Pacific and their toxicity to mice by intraperitoneal and oral administration. Mar. Drugs 2017, 15, 208. [Google Scholar] [CrossRef]
- Sibat, M.; Herrenknecht, C.; Darius, H.T.; Roué, M.; Chinain, M.; Hess, P. Detection of pacific ciguatoxins using liquid chromatography coupled to either low or high resolution mass spectrometry (LC-MS/MS). J. Chromat. A 2018, 1571, 16–28. [Google Scholar] [CrossRef]
- Longo, S.; Sibat, M.; Viallon, J.; Darius, H.T.; Hess, P.; Chinain, M. Intraspecific variability in the toxin production and toxin profiles of in vitro cultures of Gambierdiscus polynesiensis (Dinophyceae) from French Polynesia. Toxins 2019, 11, 735. [Google Scholar] [CrossRef]
- Darius, H.T.; Revel, T.; Viallon, J.; Sibat, M.; Cruchet, P.; Longo, S.; Hardison, D.R.; Holland, W.C.; Tester, P.A.; Litaker, R.W.; et al. Comparative study on the performance of three detection methods for the quantification of Pacific ciguatoxins in French Polynesian strains of Gambierdiscus polynesiensis. Toxins 2022, 20, 348. [Google Scholar] [CrossRef]
- Laza-Martínez, A.; David, H.; Riobó, P.; Miguel, I.; Orive, E. Characterization of a strain of Fukuyoa paulensis (Dinophyceae) from the Western Mediterranean Sea. J. Eukaryot. Microbiol. 2016, 63, 481–497. [Google Scholar] [CrossRef]
- Yogi, K.; Oshiro, N.; Inafuku, Y.; Hirama, M.; Yasumoto, T. Detailed LC-MS/MS analysis of ciguatoxins revealing distinct regional and species characteristics in fish and causative alga from the Pacific. Anal. Chem. 2011, 83, 8886–8891. [Google Scholar] [CrossRef]
- Yogi, K.; Oshiro, N.; Matsuda, S.; Sakugawa, S.; Matsuo, T.; Yasumoto, T. Toxin profiles in fish implicated in ciguatera fish poisoning in Amami and Kakeroma Islands, Kagoshima Prefecture, Japan. J. Food Hyg. Soc. Jpn. 2013, 54, 385–391. [Google Scholar] [CrossRef]
- Yogi, K.; Sakugawa, S.; Oshiro, N.; Ikehara, T.; Sugiyama, K.; Yasumoto, T. Determination of toxins involved in ciguatera fish poisoning in the Pacific by LC/MS. J. AOAC 2014, 97, 398–402. [Google Scholar] [CrossRef]
- Oshiro, N.; Nagaswa, H.; Kuniyoshi, K.; Kobayashi, N.; Sugita-Konishi, Y.; Asakura, H.; Yasumoto, T. Characteristic distribution of ciguatoxins in the edible parts of a grouper, Variola louti. Toxins 2021, 13, 218. [Google Scholar] [CrossRef] [PubMed]
- Oshiro, N.; Nagasawa, H.; Watanabe, M.; Nishimura, M.; Kuniyoshi, K.; Kobayashi, N.; Sugita-Konishi, Y.; Asakura, H.; Tachihara, K.; Yasumoto, T. An extensive survey of ciguatoxins on grouper Variola louti from the Ryukyu Islands, Japan, using liquid chromatography–tandem mass spectrometry (LC-MS/MS). Mar. Sci. Eng. 2022, 10, 423. [Google Scholar] [CrossRef]
- Tomikawa, T.; Kuniyoshi, K.; Ito, S.; Sakugawa, S.; Ishikawa, A.; Saito, T.; Kojima, T.; Asakura, H.; Ikehara, T.; Oshiro, N. Analysis of ciguatoxins in the Spotted Knifejaw, Oplegnathus punctatus from the waters of Japan. J. Food Hyg. Soc. Jpn. 2022, 63, 190–194. [Google Scholar] [CrossRef]
- Diaz-Pulido, G.; McCook, L.J.; Larkum, A.W.D.; Lotze, H.K.; Raven, J.A.; Schaffelke, B.; Smith, J.E.; Steneck, R.S. Chapter 7: Vulnerability of Macroalgae of the Great Barrier Reef to Climate Change. In Climate Change and the Great Barrier Reef: A Vulnerability Assessment; Johnson, J.E., Marshall, P.A., Eds.; Great Barrier Reef Marine Park Authority: Townsville, Australia, 2007; pp. 153–192. [Google Scholar]
- Wismer, S.; Hoey, A.S.; Bellwood, D.R. Cross-shelf benthic community structure on the Great Barrier Reef: Relationships between macroalgal cover and herbivore biomass. Mar. Ecol. Prog. Ser. 2009, 376, 45–54. [Google Scholar] [CrossRef]
- Tebbett, S.B.; Connolly, S.R.; Bellwood, D.R. Benthic composition changes on coral reefs at global scales. Nat. Ecol. Evol. 2023, 7, 71–81. [Google Scholar] [CrossRef] [PubMed]
- Lewis, R.J.; Sellin, M.; Gillespie, N.C.; Holmes, M.J.; Keys, A.; Street, R.; Smythe, H.; Thaggard, H.; Bryce, S. Ciguatera and herbivores: Uptake and accumulation of ciguatoxins in Ctenochaetus striatus on the Great Barrier Reef. Mem. Qld. Mus. 1994, 34, 565–570. [Google Scholar]
- Yong, H.L.; Mustapa, N.I.; Lee, L.K.; Lim, Z.F.; Tan, T.H.; Usup, G.; Gu, H.; Litaker, R.W.; Tester, P.A.; Lim, P.T.; et al. Habitat complexity affects benthic harmful dinoflagellate assemblages in the fringing reef of Rawa Island, Malaysia. Harmful Algae 2018, 78, 56–68. [Google Scholar] [CrossRef]
- Mustapa, N.I.; Yong, H.L.; Lee, L.K.; Lim, Z.F.; Lim, H.C.; Teng, S.T.; Luo, Z.; Gu, H.; Leaw, C.P.; Lim, P.T. Growth and epiphytic behaviour of three Gambierdiscus species (Dinophyceae) associated with various macroalgal substrates. Harmful Algae 2019, 89, 101671. [Google Scholar] [CrossRef]
- Lee, L.K.; Lim, Z.F.; Gu, H.; Chan, L.L.; Litaker, R.W.; Tester, P.A.; Leaw, C.P.; Lim, P.T. Effects of substratum and depth on benthic harmful dinoflagellate assemblages. Sci. Rep. 2020, 10, 11251. [Google Scholar] [CrossRef]
- Parsons, M.L.; Richlen, M.L.; Smith, T.B.; Solow, A.R.; Anderson, D.M. Evaluation of 24-h screen deployments as a standardized platform to monitor Gambierdiscus populations in the Florida Keys and U.S. Virgin Islands. Harmful Algae 2021, 103, 101998. [Google Scholar] [CrossRef] [PubMed]
- Cheal, A.; Emslie, M.; Miller, I.; Sweatman, H. The distribution of herbivorous fishes on the Great Barrier Reef. Mar. Biol. 2012, 159, 1143–1154. [Google Scholar] [CrossRef]
- Choat, J.H.; Bellwood, D.R. Interactions amongst herbivorous fishes on a coral reef: Influence of spatial variation. Mar. Biol. 1985, 89, 221–234. [Google Scholar] [CrossRef]
- Marshell, A.; Mumby, P.J. The role of surgeonfish (Acanthuridae) in maintaining algal turf biomass on coral reefs. J. Exp. Mar. Biol. Ecol. 2015, 473, 152–160. [Google Scholar] [CrossRef]
- Yasumoto, T.; Hashimoto, Y.; Bagnis, R.; Randall, J.E.; Banner, A.H. Toxicity of the surgeonfishes. Bull. Jap. Soc. Sci. Fish. 1971, 37, 724–734. [Google Scholar] [CrossRef]
- Bagnis, R. Natural versus anthropogenic disturbances to coral reefs: Comparison in epidemiological patterns of ciguatera. Mem. Qld. Mus. 1994, 34, 455–460. [Google Scholar]
- Yasumoto, T. Chemistry, etiology, and food chain dynamics of marine toxins. Proc. Jpn. Acad. 2005, 81, 43–51. [Google Scholar] [CrossRef]
- Chinain, M.; Darius, H.T.; Ung, A.; Fouc, M.T.; Revel, T.; Cruchet, P.; Paullac, S.; Laurent, D. Ciguatera risk management in French Polynesia: The case study of Raivavae Island (Australes Archipelago). Toxicon 2010, 56, 674–690. [Google Scholar] [CrossRef]
- Rongo, T.; van Woesik, R. Ciguatera poisoning in Rarotonga, southern Cook Islands. Harmful Algae 2011, 10, 345–355. [Google Scholar] [CrossRef]
- Tebbett, S.B.; Goatley, C.H.R.; Bellwood, D.R. Clarifying functional roles: Algal removal by the surgeonfishes Ctenochaetus striatus and Acanthurus nigrofuscus. Coral Reefs 2017, 36, 803–813. [Google Scholar] [CrossRef]
- Tebbett, S.B.; Goatley, C.H.R.; Huertas, V.; Mihalitsis, M.; Bellwood, D.R. A functional evaluation of feeding in the surgeonfish Ctenochaetus striatus: The role of soft tissues. R. Soc. Open Sci. 2018, 5, 17111. [Google Scholar] [CrossRef]
- Purcell, S.W.; Bellwood, D.R. A functional analysis of food procurement in two surgeonfish species, Acanthurus nigrofuscus and Ctenochaetus striatus (Acanthuridae). Env. Biol. Fish. 1993, 37, 139–159. [Google Scholar] [CrossRef]
- Marshell, A.; Mumby, P.J. Revisiting the functional roles of the surgeonfish Acanthurus nigrofuscus and Ctenochaetus striatus. Coral Reefs 2012, 31, 1093–1101. [Google Scholar] [CrossRef]
- Choat, J.H.; Clements, K.D.; Robbins, W.D. The trophic status of herbivorous fishes on coral reefs 1: Dietary analyses. Mar. Biol. 2002, 140, 613–623. [Google Scholar]
- Lin, X.; Hu, S.; Liu, Y.; Zhang, L.; Huang, H.; Liu, S. Disturbance-mediated changes in coral reef habitat provoke a positive feeding response in a major coral reef detritovore, Ctenochaetus striatus. Front. Mar. Sci. 2021, 8, 682697. [Google Scholar] [CrossRef]
- Tebbett, S.B.; Goatley, C.H.R.; Bellwood, D.R. The effects of algal turf sediments and organic loads on feeding by coral reef surgeonfishes. PLoS ONE 2017, 12, e0169479. [Google Scholar] [CrossRef]
- Holmes, M.J.; Lewis, R.J. Origin of ciguateric fish: Quantitative modelling of the flow of ciguatoxin through a marine food chain. Toxins 2022, 14, 534. [Google Scholar] [CrossRef]
- Bomber, J.W. Ecological Studies of Benthic Dinoflagellates Associated with Ciguatera from the Florida Keys. Master’s Thesis, Florida Institute of Technology, Melbourne, FL, USA, 1985; p. 104. [Google Scholar]
- Caire, J.F.; Raymond, A.; Bagnis, R. Ciguatera: Study of Setting Up and the Evolution of Gambiediscus toxicus Population on an Artificial Substrate Introduced in an Atoll Lagoon with Follow up of Associated Environmental Factors. In Proceedings of the Fifth International Coral Reef Congress: French Polynesian Coral Reefs, Tahiti, France, 27 May–1 June 1985; Delsalle, B., Galzin, R., Salvat, B., Eds.; Antenne Museum-Ephe: Moorea, France, 1985; Volume 1, pp. 429–435. [Google Scholar]
- Lobel, P.S.; Anderson, D.M.; Durand-Clement, M. Assessment of ciguatera dinoflagellate populations: Sample variability and algal substrate selection. Biol. Bull. 1988, 175, 94–101. [Google Scholar] [CrossRef]
- Parsons, M.L.; Settlemier, C.J.; Bienfang, P.K. A simple model capable of simulating the population dynamics of Gambierdiscus, the benthic dinoflagellate responsible for ciguatera fish poisoning. Harmful Algae 2010, 10, 71–80. [Google Scholar] [CrossRef]
- Loeffler, C.R.; Richlen, M.L.; Brandt, M.E.; Smith, T.B. Effects of grazing, nutrients, and depth on the ciguatera-causing dinoflagellate Gambierdiscus in the US Virgin Islands. Mar. Ecol. Prog. Ser. 2015, 531, 91–104. [Google Scholar] [CrossRef]
- Tester, P.A.; Kibler, S.R.; Holland, W.C.; Usup, G.; Vandersea, M.W.; Leaw, C.P.; Teen, L.P.; Larsen, J.; Mohammad-Noor, N.; Faust, M.A.; et al. Sampling harmful benthic dinoflagellates: Comparison of artificial and natural substrate methods. Harmful Algae 2014, 39, 8–25. [Google Scholar] [CrossRef]
- Tester, P.A.; Litaker, R.W.; Soler-Onís, E.; Fernández-Zabala, J.; Berdalet, E. Using artificial substrates to quantify Gambierdiscus and other toxic benthic dinoflagellates for monitoring purposes. Harmful Algae 2022, 120, 102351. [Google Scholar] [CrossRef] [PubMed]
- Parsons, M.L.; Brandt, A.L.; Ellsworth, A.; Leyne, A.L.; Rains, L.K.; Anderson, D.M. Assessing the use of artificial substrates to monitor Gambierdiscus populations in the Florida Keys. Harmful Algae 2017, 68, 52–66. [Google Scholar] [CrossRef]
- Fernández-Zabala, J.; Amorim, A.; Tuya, F.; Herrera, R.; Soler-Onís, E. Playing hide and seek: Distribution with depth of potentially harmful epibenthic dinoflagellates of Southern El Hierro Island, Canary Islands (NE Atlantic). Harmful Algae 2022, 117, 102271. [Google Scholar] [CrossRef]
- Nguyen-Ngoc, L.; Hai, D.-N.; Luom, P.-T.; Duyen Ngoc, H.-T.; Vinh, N.-T.; Anh, H.P.; Hue Minh, T.-T.; Tester, P.A. Seasonal occurrence of the potentially toxic benthic armoured dinoflagellates in Nha Trang Bay, South Central Coast of Viet Nam. Reg. Stud. Mar. Sci. 2022, 55, 102627. [Google Scholar] [CrossRef]
- Bomber, J.W.; Guillard, R.R.L.; Nelson, W.G. Rôles of temperature, salinity, and light in seasonality, growth, and toxicity of ciguatera-causing Gambierdiscus toxicus Adachi et Fukuyo (Dinophyceae). J. Exp. Mar. Biol. Ecol. 1988, 115, 53–65. [Google Scholar] [CrossRef]
- Kibler, S.R.; Litaker, R.W.; Holland, W.C.; Vandersea, M.W.; Tester, P.A. Growth of eight Gambierdiscus (Dinophyceae) species: Effects of temperature, salinity and irradiance. Harmful Algae 2012, 19, 1–14. [Google Scholar] [CrossRef]
- Xu, Y.; Richlen, M.L.; Liefer, J.D.; Robertson, A.; Kullis, D.; Smith, T.B.; Parsons, M.L.; Anderson, D.M. Influence of environmental variables on Gambierdiscus sp. (Dinophyceae) growth and distribution. PLoS ONE 2016, 11, e0153197. [Google Scholar] [CrossRef]
- Vacarizas, J.; Benico, G.; Austero, N.; Azanza, R. Taxonomy and toxin production of Gambierdiscus carpenteri (Dinophyceae) in a tropical marine ecosystem: The first record from the Philippines. Mar. Pol. Bull. 2018, 137, 430–443. [Google Scholar] [CrossRef]
- Holmes, M.J.; Lewis, R.J.; Gillespie, N.C. Toxicity of Australian and French Polynesian strains of Gambierdiscus toxicus (Dinophyceae) grown in culture: Characterization of a new type of maitotoxin. Toxicon 1990, 28, 1159–1172. [Google Scholar] [CrossRef]
- Sparrow, L.; Momigliano, P.; Russ, G.R.; Heimann, K. Effects of temperature, salinity and composition of the dinoflagellate assemblage on the growth of Gambierdiscus carpenteri isolated from the Great Barrier Reef. Harmful Algae 2017, 65, 52–60. [Google Scholar] [CrossRef] [PubMed]
- Holmes, M.J. Gambierdiscus yasumotoi sp. nov. (Dinophyceae), a toxic benthic dinoflagellate from southeastern Asia. J. Phycol. 1998, 34, 661–668. [Google Scholar] [CrossRef]
- Holmes, M.J.; Lewis, R.J. Bimodal cell size and fusing cells observed in a clonal culture of the ciguatoxin-producing benthic dinoflagellate Gambierdiscus (WC1/1). Toxins 2022, 14, 767. [Google Scholar] [CrossRef]
- Arjunwadkar, C.V.; Tebbett, S.B.; Bellwood, D.R.; Bourne, D.G.; Smith, H.A. Algal turf structure and composition vary with particulate loads on coral reefs. Mar. Pol. Bull. 2022, 181, 113903. [Google Scholar] [CrossRef]
- USFDA Natural Toxins. Fish and Fishery Products Hazards and Control Guidance, Fourth Edition. June 2021; U.S. Department of Health and Human Services, Food and Drug Administration, Center for Food Safety and Applied Nutrition, Table A5–11. Available online: https://www.fda.gov/media/80400/download (accessed on 14 December 2022).
- Lehane, L.; Lewis, R.J. Ciguatera: Recent advances but the risk remains. Int. J. Food Microbiol. 2000, 61, 91–125. [Google Scholar] [CrossRef]
- Anderson, T.A.; Kane, K.; Hart, A.; Appleford, P.; Evans, L.; Bennett, M.; Turner, T.B.; Bennett, S.; Davies, C.R.; Mapstone, B.D. Enhancement of Ship-Board Survivorship of Coral Trout Destined for the Live Fish Market; James Cook University: Townsville, Australia, 2003; p. 26. [Google Scholar]
- Sydney Fish Market Website. Coral Trout. Available online: https://www.sydneyfishmarket.com.au/Home/Seafood/Species-Information/List/coral-trout (accessed on 14 December 2022).
- Bennett, C.T.; Robertson, A. Depuration Kinetics and Growth Dilution of Caribbean Ciguatoxin in the Omnivore Lagodon rhomboides: Implications for Trophic Transfer and Ciguatera Risk. Toxins 2021, 13, 774. [Google Scholar] [CrossRef]
- Johansen, J.L.; Prachett, M.S.; Messmer, V.; Coker, D.J.; Tobin, A.J.; Hoey, A.S. Large predatory coral trout species unlikely to meet increasing energetic demands in a warming ocean. Sci. Rep. 2015, 5, 13830. [Google Scholar] [CrossRef]
- Scott, M.; Heupel, M.; Tobin, A.; Pratchett, M. A large predatory reef fish species moderates feeding and activity patterns in response to seasonal and latitudinal temperature variation. Sci. Rep. 2017, 7, 12966. [Google Scholar] [CrossRef] [PubMed]
- Polunin, N.V.C.; Klumpp, D.W. Ecological correlates of foraging periodicity in herbivorous reef fishes of the Coral Sea. J. Exp. Mar. Biol. Ecol. 1989, 126, 1–20. [Google Scholar] [CrossRef]
- Montgomery, W.L.; Myrberg, A.A., Jr.; Fishelson, L. Feeding ecology of surgeonfishes (Acanthuridae) in the northern Red Sea, with particular reference to Acanthurus nigrofuscus (Forsskål). J. Exp. Mar. Biol. Ecol. 1989, 132, 179–207. [Google Scholar] [CrossRef]
- Polunin, N.V.C.; Harmelin-Vivien, M.; Galzin, R. Contrasts in algal food processing among five herbivorous coral-reef fishes. J. Fish Biol. 1995, 47, 455–465. [Google Scholar] [CrossRef]
- Kelly, E.L.A.; Eynaud, Y.; Williams, I.D.; Sparks, R.T.; Dailer, M.L.; Sandin, S.A.; Smith, J.E. A budget of algal production and consumption by herbivore fisheries management area, Maui Hawaii. Ecosphere 2017, 8, e01899. [Google Scholar] [CrossRef]
- Liefer, J.D.; Richlen, M.L.; Smith, T.B.; DeBose, J.L.; Xu, Y.; Anderson, D.M.; Robertson, A. Asynchrony of Gambierdiscus spp. abundance and toxicity in the U.S. Virgin Islands: Implications for monitoring and management of ciguatera. Toxins 2021, 13, 413. [Google Scholar] [CrossRef]
- St John, J. Temporal variation in the diet of a coral reef piscivore (Pisces: Serranidae) was not seasonal. Coral Reefs 2001, 20, 163–170. [Google Scholar] [CrossRef]
- St John, J.; Russ, G.R.; Brown, I.W.; Squire, L.C. The diet of the large coral reef serranid Plectropomus leopardus in two fishing zones on the Great Barrier Reef, Australia. Fish. Bull. 2001, 99, 180–192. [Google Scholar]
- Frisch, A.J.; Ireland, M.; Baker, R. Trophic ecology of large predatory reef fishes: Energy pathways, trophic level, and implications for fisheries in a changing climate. Mar. Biol. 2014, 161, 61–73. [Google Scholar] [CrossRef]
- Matley, J.K.; Maes, G.E.; Devloo-Delva, F.; Huerlimann, R.; Chua, G.; Tobin, A.J.; Fisk, A.T.; Simpfendorfer, C.A.; Heupel, M.R. Integrating complementary methods to improve diet analysis in fishery-targeted species. Ecol. Evol. 2018, 8, 9503–9515. [Google Scholar] [CrossRef] [PubMed]
- Cheal, A.J.; MacNeill, M.A.; Cripps, E.; Emslie, M.J.; Jonker, M.; Schaffelke, B.; Sweatman, H. Coral-macroalgal phase shifts or reef resilience: Links with diversity and functional roles of herbivorous fishes on the Great Barrier Reef. Coral Reefs 2010, 29, 1005–1015. [Google Scholar] [CrossRef]
- Webley, J.; McInnes, K.; Teixeira, D.; Lawson, A.; Quinn, R. Statewide Recreational Fishing Survey 2013–2014. In Queensland Government Report, 2015; p. 127. Available online: http://era.daf.qld.gov.au/id/eprint/6513/ (accessed on 16 March 2023).
- Davin, W.T.; Kohler, C.C.; Tindall, D.R. Effects of ciguatera toxins on the bluehead. Trans. Am. Fish. Soc. 1986, 115, 908–912. [Google Scholar] [CrossRef]
- Davin, W.T.; Kohler, C.C.; Tindall, D.R. Ciguatera toxins adversely affect piscivorous fishes. Trans. Am. Fish. Soc. 1988, 117, 384–474. [Google Scholar] [CrossRef]
- Magnelia, S.J.; Kohler, C.C.; Tindall, D.R. Acanthurids do not avoid consuming cultured toxic dinoflagellates yet do not become ciguatoxic. Trans. Am. Fish. Soc. 1992, 121, 737–745. [Google Scholar] [CrossRef]
- Ledreux, A.; Brand, H.; Chinain, M.; Bottein, M.-Y.D. Dynamics of ciguatoxins from Gambierdiscus polynesiensis in the benthic herbivore Mugil cephalus: Trophic transfer implications. Harmful Algae 2014, 39, 165–174. [Google Scholar] [CrossRef]
- Clausing, R.J.; Losen, B.; Oberhaensli, F.R.; Darius, H.T.; Sibat, M.; Hess, P.; Swarzenski, P.W.; Chinain, M.; Bottein, M.-Y.D. Experimental evidence of dietary ciguatoxin accumulation in an herbivorous coral reef fish. Aquat. Toxicol. 2018, 200, 257–265. [Google Scholar] [CrossRef] [PubMed]
- Gillespie, N.C.; Holmes, M.J.; Burke, J.B.; Doley, J. Distribution and Periodicity of Gambierdiscus toxicus in Queensland, Australia. In Toxic Dinoflagellates; Anderson, D.M., White, A.W., Baden, D.G., Eds.; Elsevier: Oxford, UK, 1985; pp. 183–188. [Google Scholar]
- Skinner, M.P.; Lewis, R.J.; Morton, S. Ecology of the ciguatera causing dinoflagellates from the Northern Great Barrier Reef: Changes in community distribution and coastal eutrophication. Mar. Poll. Bull. 2013, 77, 210–219. [Google Scholar] [CrossRef] [PubMed]
- Choat, J.H.; Axe, L.M. Growth and longevity in acanthurid fishes; an analysis of otolith increments. Mar. Ecol. Prog. Ser. 1996, 134, 15–26. [Google Scholar] [CrossRef]
- Trip, E.L.; Choat, J.H.; Wilson, D.T.; Robertson, D.R. Inter-oceanic analysis of demographic variation in a widely distributed Indo-Pacific coral reef fish. Mar. Ecol. Prog. Ser. 2008, 373, 97–109. [Google Scholar] [CrossRef]
- Trip, E.D.L.; Craig, P.; Green, A.; Choat, J.H. Recruitment dynamics and first year growth of the coral reef surgeonfish Ctenochaetus striatus, with implications for acanthurid growth models. Coral Reefs 2014, 33, 879–889. [Google Scholar] [CrossRef]
- Craig, P.C.; Choat, J.H.; Axe, L.M.; Saucerman, S. Population biology and harvest of the coral reef surgeonfish Acanthurus lineatus in American Samoa. Fish. Bull. 1997, 95, 680–693. [Google Scholar]
- Doherty, P.J.; Dufour, V.; Galzin, R.; Hixon, M.A.; Meekan, M.G.; Planes, S. High mortality during settlement is a population bottleneck for a tropical surgeonfish. Ecology 2004, 85, 2422–2428. [Google Scholar] [CrossRef]
- Li, J.; Mak, Y.L.; Chang, Y.-H.; Xiao, C.; Chen, Y.-M.; Shen, J.; Wang, Q.; Ruan, Y.; Lam, P.K.S. Uptake and depuration kinetics of Pacific ciguatoxins in orange-spotted grouper (Epinephelus coioides). Environ. Sci. Technol. 2020, 54, 4475–4483. [Google Scholar] [CrossRef]
- Kingsford, M.J. Spatial and temporal variation in predation on reef fishes by coral trout (Plectropomus leopardus, Serranidae). Coral Reefs 1992, 11, 193–198. [Google Scholar] [CrossRef]
- Takahashi, M.; DiBattista, J.D.; Jarman, S.; Newman, S.J.; Wakefeld, C.B.; Harvey, E.S.; Bunce, M. Partitioning of diet between species and life history stages of sympatric and cryptic snappers (Lutjanidae) based on DNA metabarcoding. Sci. Rep. 2020, 10, 4319. [Google Scholar] [CrossRef] [PubMed]
- Russ, G.R. Grazer biomass correlates more strongly with production than with biomass of algal turfs on a coral reef. Coral Reefs 2003, 22, 63–67. [Google Scholar] [CrossRef]
- Tebbett, S.B.; Bellwood, D.R. Algal turf productivity on coral reefs: A meta-analysis. Mar. Environ. Res. 2021, 168, 105311. [Google Scholar] [CrossRef]
- Streit, R.P.; Cumming, G.S.; Bellwood, D.R. Patchy delivery of functions undermines functional redundancy in a high diversity system. Funct. Ecol. 2019, 33, 1144–1155. [Google Scholar] [CrossRef]
- Funaki, H.; Gaonkar, C.C.; Kataoka, T.; Nishimura, T.; Tanaka, K.; Yanagida, I.; Abe, S.; Yamaguchi, H.; Nagasaki, K.; Adachi, M. Horizontal and vertical distribution of Gambierdiscus spp. (Dinophyceae) including novel phylotypes in Japan identified by 18S rDNA metabarcoding. Harmful Algae 2022, 111, 102163. [Google Scholar] [CrossRef]
- Funaki, H.; Nishimura, T.; Yoshioka, T.; Ataka, T.; Tanii, Y.; Hasimoto, K.; Yamuguchi, H.; Adachi, M. Toxicity and growth characteristics of epiphytic dinoflagellate Gambierdiscus silvae in Japan. Harmful Algae 2022, 115, 102230. [Google Scholar] [CrossRef]
- Qld Health Notifiable Conditions Reports, Notifiable Conditions Annual Reporting. Available online: https://www.health.qld.gov.au/clinical-practice/guidelines-procedures/diseases-infection/surveillance/reports/notifiable/annual (accessed on 14 January 2023).
- Sanchez-Henao, A.; García-Àlvarez, N.; Padilla, D.; Ramos-Sosa, M.; Sergent, F.S.; Fernández, A.; Estévez, P.; Gago-Martínez, A.; Diogène, J.; Real, F. Accumulation of C-CTX1 in muscle tissue of goldfish (Carassius auratus) by dietary exposure. Animals 2021, 11, 242. [Google Scholar] [CrossRef]
- Mak, Y.L.; Wai, T.-C.; Murphy, M.B.; Chan, W.H.; Wu, J.J.; Lam, J.C.W.; Chan, L.L.; Lam, P.K.S. Pacific ciguatoxins in food web components of coral reef systems in the Republic of Kiribati. Environ. Sci. Tech. 2013, 47, 14070–14079. [Google Scholar] [CrossRef]
- Chan, T.Y.K. Regional variations in the risk and severity of ciguatera caused by eating moray eels. Toxins 2017, 9, 201. [Google Scholar] [CrossRef]
- Zhu, J.; Lee, W.-H.; Wu, J.; Zhou, S.; Yip, K.-C.; Liu, X.; Kirata, T.; Chan, L.-L. The occurrence, distribution, and toxicity of high-risk ciguatera fish species (grouper and snapper) in Kiritimati Island and Marakei Island of the Republic of Kiribati. Toxins 2022, 14, 208. [Google Scholar] [CrossRef] [PubMed]
- Bagnis, R. Naissance et développment d’une flambée de ciguatera dans un atoll des Tuamotu. Rev. Corps St. 1969, 10, 783–795. [Google Scholar]
- Darius, H.T.; Ponton, D.; Revel, T.; Cruchet, P.; Ung, A.; Tchou Fouc, M.; Chinain, M. Ciguatera risk assessment in two toxic sites of French Polynesia using the receptor-binding assay. Toxicon 2007, 50, 612–626. [Google Scholar] [CrossRef] [PubMed]
- Gaboriau, M.; Ponton, D.; Darius, H.T.; Chinain, M. Ciguatera fish toxicity in French Polynesia: Size does not always matter. Toxicon 2014, 84, 41–50. [Google Scholar] [CrossRef] [PubMed]
- Satake, M.; Fukui, M.; Legrand, A.-M.; Cruchet, P.; Yasumoto, T. Isolation and structures of new ciguatoxin analogs, 2,3-dihydroxyCTX3C and 51-hydroxyCTX3C, accumulated in tropical reef fish. Tetrahedron Lett. 1998, 39, 1197–1198. [Google Scholar] [CrossRef]
- Dechraoui, M.Y.; Naar, J.; Pauillac, S.; Legrand, A.M. Ciguatoxins and brevetoxins, neurotoxic polyether compounds active on sodium channels. Toxicon 1999, 37, 125–143. [Google Scholar] [CrossRef]
- Murata, M.; Legrand, A.-M.; Ishibashi, Y.; Fukui, M.; Yasumoto, T. Structures and Configurations of Ciguatoxin from the Moray Eel Gymnothorax javanicus and Its Likely Precursor from the Dinoflagellate Gambierdiscus toxicus. J. Am. Chem. Soc. 1990, 112, 4380–4386. [Google Scholar] [CrossRef]
- Oshiro, N.; Tomikawa, T.; Kuniyoshi, K.; Ishikawa, A.; Toyofuku, H.; Kojima, T.; Asakura, H. LC–MS/MS analysis of ciguatoxins revealing the regional and species distinction of fish in the tropical Western Pacific. J. Mar. Sci. Eng. 2021, 9, 299. [Google Scholar] [CrossRef]
- Spielmeyer, A.; Loeffler, C.R.; Kappenstein, O. Identical ciguatoxin-3C group profiles in Lutjanus bohar from the Pacific and Indian Oceans-indicating the need to re-evaluate geographical CTX classifications. Front. Mar. Sci. 2022, 9, 937438. [Google Scholar] [CrossRef]
- Anguilar-Medrano, R. Ecomorphology and evolution of the pharyngeal apparatus of benthic damselfishes (Pomacentridae, subfamily Stegastinae). Mar. Biol. 2017, 164, 21. [Google Scholar] [CrossRef]
- McCord, C.L.; Nash, C.M.; Cooper, W.J.; Westneat, M.W. Phylogeny of the damselfishes (Pomacentridae) and patterns of asymmetrical diversification in body size and feeding ecology. PLoS ONE 2021, 16, e0258889. [Google Scholar] [CrossRef] [PubMed]
- Díaz-Ascenio, L.; Clausing, R.J.; Vandersea, M.; Chamero-Lago, D.; Gómez-Batista, M.; Hernández-Albernas, J.I.; Chomérat, N.; Rojas-Abrahantes, G.; Litaker, R.W.; Tester, P.; et al. Ciguatoxin occurrence in food-web components of a Cuban coral reef ecosystem: Risk-assessment implications. Toxins 2019, 11, 722. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| P-CTX-4B and/or P-CTX-4A/Gambierdiscus Cell (pg/Cell) | Days to Consume Turf Algae with 10 Gambierdiscus/cm2 | Days to Consume Turf Algae with 100 Gambierdiscus/cm2 | Days to Consume Turf Algae with 1000 Gambierdiscus/cm2 |
|---|---|---|---|
| 1.6 | 36–143 | 4–14 | 0.4–1.4 |
| 0.6 | 95–382 | 10–38 | 1–3.8 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Holmes, M.J.; Lewis, R.J. Model of the Origin of a Ciguatoxic Grouper (Plectropomus leopardus). Toxins 2023, 15, 230. https://doi.org/10.3390/toxins15030230
Holmes MJ, Lewis RJ. Model of the Origin of a Ciguatoxic Grouper (Plectropomus leopardus). Toxins. 2023; 15(3):230. https://doi.org/10.3390/toxins15030230
Chicago/Turabian StyleHolmes, Michael J., and Richard J. Lewis. 2023. "Model of the Origin of a Ciguatoxic Grouper (Plectropomus leopardus)" Toxins 15, no. 3: 230. https://doi.org/10.3390/toxins15030230
APA StyleHolmes, M. J., & Lewis, R. J. (2023). Model of the Origin of a Ciguatoxic Grouper (Plectropomus leopardus). Toxins, 15(3), 230. https://doi.org/10.3390/toxins15030230