Abstract
The relative lack of marine venom pharmaceuticals can be anecdotally attributed to difficulties in working with venomous marine animals, including how to maintain venom bioactivity during extraction and purification. The primary aim of this systematic literature review was to examine the key factors for consideration when extracting and purifying jellyfish venom toxins to maximise their effectiveness in bioassays towards the characterisation of a single toxin.An up-to-date database of 119 peer-reviewed research articles was established for all purified and semi-purified venoms across all jellyfish, including their level of purification, LD50, and the types of experimental toxicity bioassay used (e.g., whole animal and cell lines). We report that, of the toxins successfully purified across all jellyfish, the class Cubozoa (i.e., Chironex fleckeri and Carybdea rastoni) was most highly represented, followed by Scyphozoa and Hydrozoa. We outline the best practices for maintaining jellyfish venom bioactivity, including strict thermal management, using the “autolysis” extraction method and two-step liquid chromatography purification involving size exclusion chromatography. To date, the box jellyfish C. fleckeri has been the most effective jellyfish venom model with the most referenced extraction methods and the most isolated toxins, including CfTX-A/B. In summary, this review can be used as a resource for the efficient extraction, purification, and identification of jellyfish venom toxins.
Key Contribution:
A database of all purified and semi-purified venoms across all jellyfish, including their LD50, current status of venom purification, and existing bioassay models that have reported the purification of toxins of jellyfish supported by bioassays. Identification of extraction and purification methods of venom used during isolation of single toxins.
1. Introduction
Animal venom is a proven source of potent bioactive compounds, many of which have been adapted into pharmaceuticals. Specifically, 11 venom-derived drugs are commercially available and approved by both the United States Food and Drug Administration and the European Medicines Agency [1]. The wide range of venom bioactivity can be seen in these different drugs being utilised in the treatment of coronary syndrome, chronic pain treatment, diabetes, hypertension, and sciatica [1]. Only one of these drugs, Prialt®, comes from a marine organism (Conus magus), which is symptomatic of the widening gap between terrestrial and marine venom research. This deficiency in marine venom research can be anecdotally attributed to the difficulties of working with marine venom in contrast to terrestrial venom. These difficulties include the availability of target organisms in the wild (often seasonal) and poor knowledge of aquaculture of the organisms and extraction procedures from tissue(s) that maintain bioactivity. Regarding the extraction of biomolecules from tissues, a significant diversity of methodologies exists in the literature for jellyfish venom.
Medusozoa represents the clade of organisms within the phylum Cnidaria that contain a dominant free-swimming phase in their life cycle, known as a medusa stage [2,3]. It is this swimming behaviour that gives these organisms the colloquial name of “jellyfish” (also known as sea jellies). There are however several non-jellyfish exceptions within Hydrozoa such as the hydra included within this review, falling under the sub-phylum Medusozoa. These jellyfish have spread across four classes; Cubozoa (box jellyfish), Hydrozoa (hydra and siphonophores), Scyphozoa (true swimming jellyfish), and Staurozoa (stalked jellyfish) [2,3]. Medusozoans deliver their venom using cells called cnidocytes, containing a ballistically discharging organelle called a nematocyst to inject venom into their prey [4]. The cnidocytes capsules are discharged as a response to either a chemical signal or mechanical stimulus that is triggered by contact with prey organisms [5].
Jellyfish venoms are composed of many complex proteins, peptides, and small molecules, each with extensive and sometimes specific bioactivities [4]. This is mainly attributed to the inability for cognitive selection during their predation, requiring their venom to have cytotoxic, cardiotoxic, and neurotoxic effects across a wider range of prey [6]. As such, some jellyfish have venom so benign to humans they are referred to as “non-venomous”, while others such as box jellyfish and Irukandji have venom so powerful, they are well known to cause death in humans within hours [7]. This wide range of bioactivities observed across both the proteinaceous and non-proteinaceous components of jellyfish venom is of emerging focus towards not only biodiscovery but also the development of more effective first aid and clinical treatments, involving the development of an anti-venom [8,9,10,11].
While many toxins have been reported across various different jellyfish species using bottom-up proteomic investigations, only a few have been confirmed as venomous in vitro via purification and subsequently had their native function characterised. Venoms can often exhibit complex structures, such as multiple disulphide bridges and post-translational modifications and, as such, often require a purification from the whole native venom to confirm bioactivity, prior to commitment towards synthetic or recombinant production. Although multiple reviews have comprehensively covered the properties and specific function of known jellyfish venoms [12,13,14,15,16,17,18,19], to date, no systematic review has compared the methods used to extract individual venom, nor compared 50% lethal dose (LD50) of functional assays to identify the best purification method. As such, it is unclear if there are key factors to consider for primary extraction or purification that may ensure the native structure is maintained.
The primary aim of this systematic [20] literature review was to examine the key factors for consideration for the extraction and purification of native venom toxins to maximise their effectiveness in bioassays towards the development of a more standardised methodology. This knowledge could make the biodiscovery of jellyfish venom toxins more appealing to researchers, particularly those unable to justify the financial commitment of peptide synthesis based entirely on either in silico or bottom-up proteomic analysis. In the process, we have developed an up-to-date database of all purified and semi-purified venoms across all venoms within the literature captured (up until 2023), including their LD50, the current status of venom purification, and existing bioassay models.
2. Results
2.1. Captured Literature
The captured literature database contained 119 peer-reviewed research articles and 5 literature reviews that have reported the purification of toxins of jellyfish supported by bioassays. Of the reviewed literature, the first reported purification of a jellyfish venom and its assay was in 1975 (Figure 1), in which venom from Stomoiophus meieagris was fractionated and Na+ transporting influencing factors were identified [21]. Relevant literature gradually increased from 2001, peaking in 2012 (13 publications), and then trending downwards up to now. This decrease may be attributed to the inherent difficulties associated with jellyfish venom extraction and purifications relative to terrestrial venoms which is further discussed later in this review. This is best highlighted by the vast majority of the captured articles including a recommendation for the continuation of the research, yet the venom and toxin fractions they identified did not receive any subsequent publications detailing further refinement. While speculative, this demonstrates the difficulty of jellyfish purification in the initial stages of research, regularly causes the delivery of results that do not justify further research into the venom when more favourable sources of bioactive molecules may be present. Overall, the class Scyphozoa represented the most common jellyfish class for venom purifications (90 publications), followed by Cubozoa (39 publications) and Hydrozoa (12 publications). No venom had been reported purified from any Staurozoa.
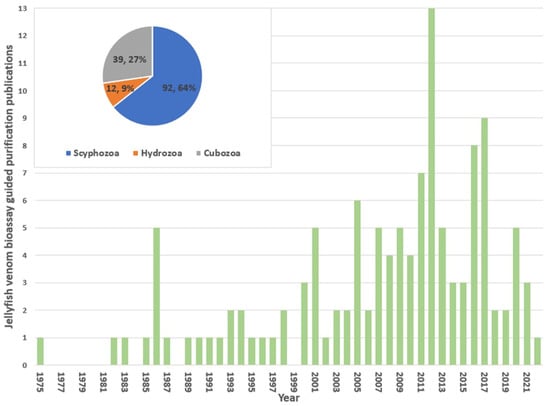
Figure 1.
The bar graph shows the number of jellyfish venom purification papers identified per year (1975–2022). In the inset, the pie chart shows the relative distribution (number and percentage) represented by jellyfish class.
2.2. Jellyfish Venom Identification
A total of 202 venom proteins have been reported from jellyfish, with varying degrees of purification (Table 1), which could be classified into (1) crude venom, (2) a venom fraction, and (3) isolated venom toxins. Crude venom was defined as venom that had been removed from nematocysts, filtered and/or sonicated and/or centrifuged, and intended to be representative of a jellyfish’s entire venom toxin repertoire (i.e., the entire venome). A venom fraction was defined as crude venom that had had its components separated (e.g., liquid chromatography) to refine the potential number of venom toxin products. Finally, isolated toxins were defined as a single homogenous product, pure enough to obtain a single kilodalton (kDa) measurement, which varied from 1 to 600 kDa. For each classification of purification, the venom had additionally been tested in toxicity assays to obtain an LD50 (or LD50 equivalent for cellular assays). Of the 202 reported venoms, 99 (49%) were crude venom, 62 (31%) were venom fractions, and 38 (20%) were isolated venom toxins.

Table 1.
Summary of venom or venom toxins isolated from jellyfish, including name, size (kDa), purification level, LD50, and/or reported primary activity. Toxin names have been left abbreviated for clear reference to the original articles. * Non-jellyfish species (hydra).
3. Discussion
3.1. Purification Levels of Jellyfish Venom
A total of 31 jellyfish species had at least one venom purification level reported (Table 1 and Figure 2), with Chironex fleckeri, Chrysaora quinquecirrha (Atlantic Sea nettle), and Cyanea capillata (Lion’s mane jellyfish) most extensively investigated. C. quinquecirrha shares C. fleckeri’s status as a ‘medically relevant’ jellyfish [31], due to its rapidly acting cardiotoxic venom [40] and powerful haemolytic properties [36,37]. In addition, it inhabits the East Coast of the United States and the Gulf of Mexico, thereby serving as a convenient model for venom researchers in North America. The observed abundance of C. capillata venom research follows a similar logic, with a powerful cardiotoxic venom [40,52,54,57] and a niche distribution within the northern oceans, extending to the Artic, which helps to explain their disproportionately large representation in the literature.
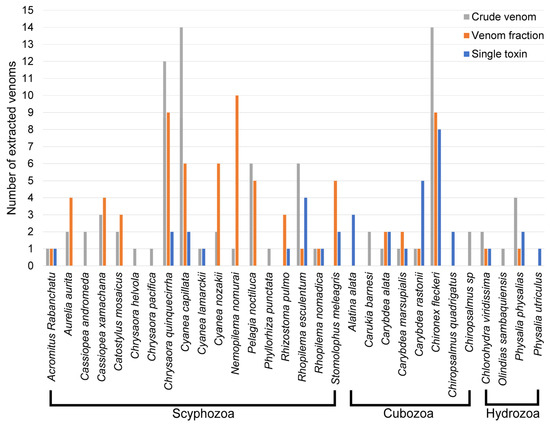
Figure 2.
Graph showing a summary of jellyfish species by class (in alphabetical order) and level of their venom purification.
The data highlight that Aurelia aurita, Cassiopea xamachana, Catostylus mosaicus, Cyanea nozakii, Nemopilema nomurai, and Pelagia noctiluca have had toxic fractions identified, yet not a single toxin purified. These jellyfish represent an excellent opportunity for research as there exists a strong foundation of effective venom purification and fractionation. Moreover, these jellyfish only represent 32 species of over 3500 known jellyfish species from which no, or very little, research has been performed on their venom. Although hydrozoans represent around 77% of all medusozoan species [2,3], they only occupied 12% of the venom purifications reported.
3.2. Jellyfish Venom Extraction
Jellyfish venom for subsequent purification of single toxins and toxicity bioassay confirmation have been successfully obtained using a variety of different extraction and purification procedures (Table 2). As categorised by Carrette and Seymour (2004) [33] venom extraction methods can be separated into, natural discharge of intact nematocysts, mechanical disruption of nematocysts, chemically induced discharge of intact nematocysts and removal of liquid from individual intact nematocysts [33]. Only natural and mechanic methods of extraction returned individual toxins.
Table 2.
Summary of 38 toxins successfully isolated, including extraction and purification procedures. SEC, size-exclusion chromatography; CEX, cation exchange chromatography; AEX anion exchange chromatography; HPLC, high-performance liquid chromatography; RP-HPLC, reverse phase high performance liquid chromatography.
The most commonly referenced isolation methods [22,23,37,89,94,113,114] were developed by Burnett et al. (1992) [43] and Bloom et al. (1998) [26], the latter of which had been refined from Burnett’s original method. Briefly, jellyfish tentacles were freshly removed from beachside individuals, then immediately refrigerated within two volumes of seawater for 1–4 days. For venom extraction, the tentacle samples were shaken, and an aliquot of water was taken daily for filtration with filter paper prior to being viewed under a microscope to confirm the release of intact cnidocytes from tissue. The cnidocytes were then lyophilised and stored (−70 °C) prior to extraction. The venom was extracted from the cnidocytes via sonication (3 × 20 s with 1 min intervals) in ice-cold water (4 °C) before centrifugation at 20,000× g for 1 h at 4 °C and then immediately used in toxicity assays. This approach was regularly referred to as the ‘autolysis’ venom extraction technique since it relied on autolysis of tentacle tissue (but not the cnidocytes themselves). Variations in autolysis venom extraction included tentacle incubation times of one day [88] to up to 6 weeks [82]. However, the longest time of autolysis that yielded a single proteinaceous toxin was 4 days. Therefore, it is recommended that autolysis be no longer than 4 days.
The efficiency of the venom extraction autolysis technique may be attributed to a few factors. First, it has been speculated that the intracellular environment of cnidocytes, including high pressure, contributes to venom stability. In addition, venom is known to act rapidly to facilitate processes that disable prey; therefore, they do not require a high degree of structural stability that could maintain a longer half-life outside of the cellular environment [26].
A variation of the autolysis technique involved the agitation of jellyfish tentacles in a buffered solution for up to 6 weeks to allow for maximal separation of the cnidocytes. Cnidocytes were then chilled in a pressure cell at 12,000 psi for 15 min, followed by a freeze–thaw cycle to help liberate the venom [82]. While this method is a conventional approach for liberating the intracellular components of a cell for small molecule analysis, it should not be used when isolating proteinaceous toxins from jellyfish venom, as multiple studies have reported that freeze–thaw cycles cause a significant decrease in bioactivity [65,83] particularly for proteinaceous compounds. This is relatively well known, as the phase change of an aqueous environment can cause alterations at the tertiary and quaternary levels of proteins by mechanical disruption of the structure during freezing.
A strict thermal regulation regime appears to play a critical role in the preservation of jellyfish venom bioactivity and, therefore, its potency. This is irrespective of cnidocyte separation since a large number of jellyfish toxins have been successfully isolated using whole tentacle homogenisation [84,91,93,95,98,102] and whole tentacle lysis [83]. Thermal denaturation of jellyfish toxins has been studied to investigate their sensitivity. For example, the activity of the CnPH toxin was significantly reduced when exposed to temperatures above 45 °C [116] and the venom activity of the flame jellyfish (Rhopilema esculentum) was found to be reduced at 40 °C [8]. It was additionally inferred during the purification of ClGp1 from lysed cnidocytes that it should be performed as fast as possible to preserve bioactivity [98].
Whole tentacle homogenisation and lysis use mechanical, electrical, or chemical means to break open cells, and in doing so, cause nematocysts to fire, releasing the venom. Thus, whole tentacle extraction undoubtedly adds ambiguity as to the true source of the toxin and introduces background biomolecules to the subsequent purification process. For instance, the enzyme hydralysin was identified from a Chlorohydra viridissima tentacle homogenate [91], yet it is a cytolytic/neurotoxic proteinaceous toxin that originates from a non-cnidocyte source. This, as well as other examples, indicates the possibility that not all jellyfish “venom” is delivered from nematocysts [15,91]. Whole tentacle extraction may be a better choice if the sample size is low as it will provide a more concentrated venom extract. However, if the jellyfish venom is of low toxicity and if the venom readily loses bioactivity under extraction conditions, it may be significantly impacted during the hours or days that the tentacles are suspended in the separation solution.
3.3. Jellyfish Venom Purification
A total of 38 individual jellyfish venom toxins have been identified (Table 2). In all of the isolated toxins within the literature-captured jellyfish venom purification studies, liquid chromatography was the primary method of choice, while most of them utilised a combination of at least two chromatography techniques, including HPLC [93], RP-HPLC [6], SEC [7], CEX [94], and AEX [102]. Only two studies required additional chromatography to ensure the purity of a single toxin [7,59]. Non-chromatography purification processes have also been used in tandem with liquid chromatography, using acidic precipitation [102] and a “salting out” protein precipitation method [103]. While these methods were shown to have produced single toxins from crude venom, they should only be used as a final purification step for identification, when the activity and purity of the toxin have been confirmed. This is due to protein precipitation causing the denaturation of proteins, thereby removing any chance to confirm its native function in bioassays.
Size exclusion chromatography was used in more than 70% of the jellyfish toxin purification studies, thereby representing the most common liquid chromatography method. SEC is particularly useful when purifying unknown toxins, as it does not require organic solvents. Organic solvents, such as acetonitrile, methanol, and ethanol, commonly used in RP-HPLC and chloroform in HPLC, are known to modify protein structure, thereby potentially affecting a toxin’s function. Unlike CEX and AEX, SEC does not require prior knowledge of the proteins’ solubility, charge, or isoelectric point and will provide better resolution across a sample with many compounds. SEC can be performed as high-throughput [23] or used as a last step to “polish” toxins [98].
3.4. Toxicity Assays of Venom towards Proposed Function
The combined data obtained, from jellyfish purification to identification, indicate that overall direct comparisons should not be performed due to the lack of commonality between studies regarding the toxicity assay (Figure 3). For jellyfish venoms, mice have been the most used animal model system for toxicity characterisation (to establish lethal dosage; Figure 3A). This is consistent with studies on other types of venom (derived from marine or terrestrial organisms), whereby functional toxicity assays using mice are considered relatively cheap with accessible ethical requirements, as well thiersimilar biological, physiological, and symptomatic effects to humans. However, as animal ethics for vertebrate experimentation become stricter, invertebrate animal models (e.g., crayfish, flies, moths, and spiders) may be more widely used. In addition, jellyfish predate most commonly on small fish and crustaceans making them the most biologically relevant models for whole animal toxicity testing.
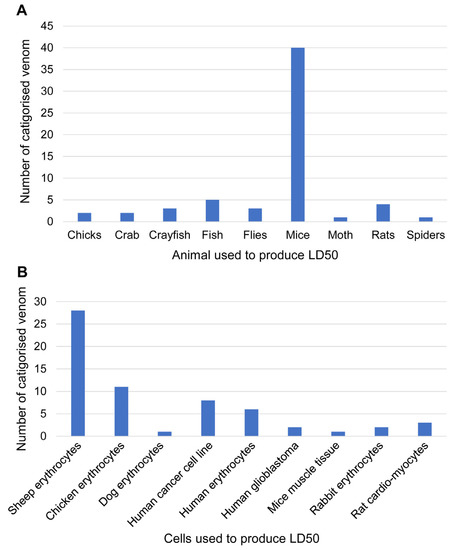
Figure 3.
Summary of animal and cell toxicity assays used for bioassay-guided identification of jellyfish venom, including LD50. (A) Graph showing the number of venom toxicity assays using whole animal models. (B) Graph showing the number of venom toxicity assays using cell-based toxicity assays.
Erythrocytes are excellent for cell-based toxicity assays to establish haemolytic activity. To assess jellyfish venom toxicity, sheep erythrocytes have been the most used, likely due to their ethical sourcing, relatively low cost, and high availability (Figure 3B). Chicken erythrocytes have also been commonly used, which although non-mammalian, are still nucleated cells that have demonstrated clear comparative results against different types of jellyfish venom [8,40,45,78,88]. Human erythrocytes have also been well used in cell-based toxicity assays, primarily with relevance to investigating cardiovascular and haemolytic impacts on humans; they are also more sensitive to venom compared to sheep erythrocytes [92]. The haemolytic effects have been studied in human erythrocytes to reveal venom protein interactions with membrane lipids, which were proposed to be directly and most likely involved in pore formation [34,71,92,109]. Research investigating these impacts has used various venom purification methods as this model gives autonomy in method designation, although the HU50s (one haemolytic unit, the amount of protein sample required to induce 50% haemolysis, referred to as LD50 in this review) were quite different between methods conducted on the same species [34]. However, the most dependable method was presented by Yanagihara and Shohet (2012) [34], where human erythrocytes were recognised as being a reproducible source for experiments. They showed specific membrane breakdown when interacting with venom toxins; although considering that different jellyfish venoms contain different repertoires of toxins, they consequently have different haemolytic activity that might not be possible to compare; in addition, reportedly, careful consideration should be taken with the enzymes used in bioassays to avoid inhibitory effects on venom proteins [91].
Human cancer cell lines are often used to elucidate the harmful effects of venoms on humans and to understand the molecular mechanisms of venom toxicity, and they have been used on numerous occasions for jellyfish venom toxins [57,65,72,118,123,124,129]. They provide a reliable, high-throughput, convenient, and relatively cheap approach [124], which is primarily relevant for medical and pharmaceutical applications, specifically when it comes to first aid treatments regarding understudied jellyfish venom stings [123,124]. This model, however, has its own limitation, with a bias for the most accessible cell lines available, thus causing variation in the obtained results [57,65,72,118,123,124,129]. The most common mechanistic function reported using these cell lines involved reactive oxygen species (ROS) and cell apoptosis.
However, it was revealed that comparisons using the same animal model and the crude venom of the same species may not aid in elucidating the most effective purification or extraction technique. Separate investigations of Chironex fleckeri “crude venom”, described in the articles as crude tentacle venom [28], stock venom [9], nematocyst venom [26], and Chironex venom [32], all used near identical methodologies while producing significantly different LD50s in mice of 11, 12, 23.4, and 150 µg/mL, respectively. This variation in LD50 is most likely due to individual variations in venom potency between jellyfish or venom degradation that may have occurred prior to individual capture and isolation. As jellyfish are difficult to capture while in the ocean, especially smaller species, specimens are most often collected after they have washed up on the beach, where animal death and thermal desiccation quickly degrade the proteinaceous compounds within the venom. Furthermore, as very few jellyfish species are able to be maintained in captivity, repeated bioassays using the same individual animal’s venom is not possible, thus compounding the source of error due to individual variability of venom potency. Although not captured in the scope of this review, it is important to highlight a 2019 study of Nemopilema nomurai which showed that individual nomurai venom showed significantly different enzymatic metalloproteinase activity, with up to a 77-fold difference in haemolytic activity [130]. Moreover, the geographical location and the jellyfish’s age are correlated with significant variations in jellyfish venom of the same species [131].
Published studies have noted difficulties associated with the attempted analysis of reported jellyfish venom due to accidental misreporting of the specific activity of venom and the vastly different methodologies involved in the preparation and bioassays [82]. Complicating analysis further, jellyfish venom can have a unique haemolytic activity profile that can be highly variable between jellyfish species [65,116] and that very rarely aligns with general cytotoxicity assay results. Furthermore, the objective of many research studies was not to classify the general function or toxicity of the venom towards the discovery of a toxin. Instead, their objective was focused on the biodiscovery of a compound with a predetermined, specific function: for example, studies of jellyfish venom targeting angiotensin I, converting enzyme (ACE) inhibitors [76,93,112]. As such, jellyfish venom studies currently lack a standard methodology for toxicity measurements in cells and animal models using LD50 measurements. Function-specific bioassays of toxins are too specific and lack any type of standardisation, meaning that the comparison of bioassay data between articles for the purpose of elucidating the most effective purification techniques may not be possible. This highlights that jellyfish venom LD50s should not be used as a means to compare the effectiveness of venom purification or extraction methods. The limitations of venom potency can be best mitigated by obtaining the largest possible sample size when collecting venom for extraction and excluding juvenile or desiccated jellyfish from extractions.
3.5. Chironex fleckeri, a Model for Jellyfish Venom Purification
The box jellyfish, Chironex fleckeri, is often referred to as the most medically relevant jellyfish [17,31,111], given that its powerful haemolytic and cardiotoxic toxins have, to date, been prime sources for biodiscovery. In fact, the increase in jellyfish venom purification studies can generally be attributed to the attention brought to the field by extensive research performed on cardiotoxic venoms derived from C. fleckeri (Figure 2). The box jellyfish is well known for an extremely lethal and fast-acting venom that can be lethal to humans and prey alike. Thus, C. fleckeri represents the most researched jellyfish, with the most venomous LD50 score, and from which the most jellyfish toxins have been identified. Although there are outcomes that have reported on the toxin properties and lethality of the whole venom [9,10,23,24,27,28,29,30,32,33,34,99] leading to many bioactive fractions [9,24,25,26,27], the primary lethal toxin responsible for human fatalities has not been reported. Nonetheless, C. fleckeri represents a well-established model for the purification of jellyfish toxins and could be further used by researchers for methods of venom characterisation of Medusozoan.
4. Conclusions
This review has systematically compiled all kinds of Medusozoan purified venoms (crude venom, toxic fractions, and single toxins), which may serve as a valuable resource for venom researchers. Despite initial intensive foundational research since 1974, with research peaking in 2011, very few toxins have been successfully purified across the Medusozoans, with the exceptions being C. fleckeri and C. rastoni. Importantly, in order to characterise a single toxin when guided by bioassays, a strict thermal management plan is required to avoid loss of potency, thereby improving the success rate of purifications. Although a clear universal methodology for venom purification could not be elucidated, there is a clear trend for two-step liquid chromatography involving SEC to isolate a single toxin. This variation in the methodologies used, as well as a lack of duplication and the natural variation in individual species’ venom, currently precludes our ability to compare bioassay data. This points towards jellyfish venom extraction and purification methods being entirely dependent upon the observed chemical and functional behaviour of the toxins of interest. Therefore, they should be experimentally determined per species via functional bioassays of the toxin, to confirm the conservation of the bioactivity at each step. Performing comparisons of venom LD50, even when using the same model organism and same jellyfish species, has been shown to be of questionable value given the discussed limitations. To date, C. fleckeri has been the most effective jellyfish venom model, from which researchers have developed widely used extraction and purification methods, leading to a large number of venoms being categorised. While jellyfish venom research continues to trail behind terrestrial venom research, this does provide ample opportunity for novel discovery.
5. Method
Only literature captured using the methodology described herein is referenced in this review to maintain the integrity of the analysis, unless specifically stated. The inclusion criteria were as follows: Scopus, Web of Science, and Pub Med databases were searched using the keywords “purification or purified” and “venom” in “all fields” using the access provided by the University of the Sunshine Coast. Additionally, five secondary search terms, including “Jellyfish”, “Cubozoa” or “Cubozoan”, “Hydrozoa” or “Hydrozoan”, “Scyphozoa” or “Scyphozoan”, and “Staurozoa” or “Staurozoan”, were added and combined to ensure the capture of all jellyfish purification publications for analysis (Figure 4), with duplicate articles removed. The exclusion criteria were applied as follows: (1) if no venom purification was detailed in the publication, (2) if the venom detailed in the publication is not a single Medusozoan venom, and (3) if the venom purification detailed in the publication was non-guided with no bioassays. Using the above methodology, the literature from between 1 January 1975–15 December 2022 was captured as the earliest and latest articles. This methodology followed the Transparent Reporting of Systematic Reviews and Meta-Analysis (PRISMA) [20]; the database of all of the literature information is provided in File S1.
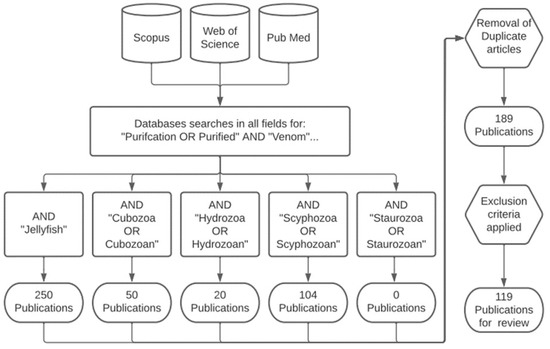
Figure 4.
Flowcharts of the systematic method used for this review, producing the 119 publications for analysis.
Supplementary Materials
The following supporting information can be downloaded at: https://www.mdpi.com/article/10.3390/toxins15030170/s1. File S1: Captured literature database and reporting of inclusion and exclusion reasons.
Author Contributions
Conceptualization, A.A., B.L., S.C. and T.W.; methodology, B.L.; validation, B.L., S.C. and T.W.; formal analysis, B.L.; investigation, A.A. and B.L.; resources, B.L., S.C. and T.W.; writing—original draft preparation, B.L. and A.A.; writing—review and editing, A.A., S.C. and T.W.; visualization, B.L.; supervision, S.C. and T.W.; project administration, S.C. and T.W.; funding acquisition, S.C. All authors have read and agreed to the published version of the manuscript.
Funding
This study was supported by the Australian Research Council Discovery Project (DP220103933).
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
All research data are included within the Supplementary Materials.
Acknowledgments
We thank Abigail Elizur of the Centre for Bioinovation for her valuable advice and support.
Conflicts of Interest
The authors declare no conflict of interest.
References
- De Castro Figueiredo Bordon, K.; Cologna, C.T.; Fornari-Baldo, E.C.; Pinheiro-Júnior, E.L.; Cerni, F.A.; Amorim, F.G.; Anjolette, F.A.P.; Cordeiro, F.A.; Wiezel, G.A.; Cardoso, I.A.; et al. From Animal Poisons and Venoms to Medicines: Achievements, Challenges and Perspectives in Drug Discovery. Front. Pharmacol. 2020, 11, 1132. [Google Scholar] [CrossRef] [PubMed]
- Podobnik, M.; Anderluh, G. Pore-forming toxins in Cnidaria. Semin. Cell Dev. Biol. 2017, 72, 133–141. [Google Scholar] [CrossRef] [PubMed]
- Kayal, E.; Bentlage, B.; Pankey, M.S.; Ohdera, A.H.; Medina, M.; Plachetzki, D.C.; Collins, A.G.; Ryan, J.F. Phylogenomics provides a robust topology of the major cnidarian lineages and insights on the origins of key organismal traits. BMC Evol. Biol. 2018, 18, 68. [Google Scholar] [CrossRef]
- Yue, Y.; Yu, H.; Li, R.; Xing, R.; Liu, S.; Li, K.; Wang, X.; Chen, X.; Li, P. Functional Elucidation of Nemopilema nomurai and Cyanea nozakii Nematocyst Venoms’ Lytic Activity Using Mass Spectrometry and Zymography. Toxins 2017, 9, 47. [Google Scholar] [CrossRef]
- Sánchez-Rodríguez, J.; Torrens, E.; Segura-Puertas, L. Partial purification and characterization of a novel neurotoxin and three cytolysins from box jellyfish (Carybdea marsupialis) nematocyst venom. Arch. Toxicol. 2005, 80, 163–168. [Google Scholar] [CrossRef] [PubMed]
- Lassen, S.; Wiebring, A.; Helmholz, H.; Ruhnau, C.; Prange, A. Isolation of a Nav channel blocking polypeptide from Cyanea capillata medusae—A neurotoxin contained in fishing tentacle isorhizas. Toxicon 2012, 59, 610–616. [Google Scholar] [CrossRef] [PubMed]
- Endean, R. Separation of two myotoxins from nematocysts of the box jellyfish (Chironex fleckeri). Toxicon 1987, 25, 483–492. [Google Scholar] [CrossRef] [PubMed]
- Yu, H.; Li, C.; Li, R.; Xing, R.; Liu, S.; Li, P. Factors influencing hemolytic activity of venom from the jellyfish Rhopilema esculentum Kishinouye. Food Chem. Toxicol. 2007, 45, 1173–1178. [Google Scholar] [CrossRef]
- Calton, G.J.; Burnett, J.W. Partial purification of Chironex fleckeri (sea wasp) venom by immunochromatography with antivenom. Toxicon 1986, 24, 416–420. [Google Scholar] [CrossRef]
- Ramasamy, S.; Isbister, G.K.; Seymour, J.E.; Hodgson, W.C. The in vitro effects of two chirodropid (Chironex fleckeri and Chiropsalmus sp.) venoms: Efficacy of box jellyfish antivenom. Toxicon 2003, 41, 703–711. [Google Scholar] [CrossRef] [PubMed]
- Li, R.; Yu, H.; Li, A.; Yu, C.; Li, P. Refinement and Neutralization Evaluation of the F(ab’)2 Type of Antivenom against the Deadly Jellyfish Nemopilema nomurai Toxins. Int. J. Mol. Sci. 2021, 22, 12672. [Google Scholar] [CrossRef]
- D’Ambra, I.; Lauritano, C. A Review of Toxins from Cnidaria. Mar. Drugs 2020, 18, 507. [Google Scholar] [CrossRef] [PubMed]
- Nisa, S.A.; Vinu, D.; Krupakar, P.; Govindaraju, K.; Sharma, D.; Vivek, R. Jellyfish venom proteins and their pharmacological potentials: A review. Int. J. Biol. Macromol. 2021, 176, 424–436. [Google Scholar] [CrossRef] [PubMed]
- Jouiaei, M.; Yanagihara, A.A.; Madio, B.; Nevalainen, T.J.; Alewood, P.F.; Fry, B.G. Ancient Venom Systems: A Review on Cnidaria Toxins. Toxins 2015, 7, 2251–2271. [Google Scholar] [CrossRef] [PubMed]
- Badre, S. Bioactive toxins from stinging jellyfish. Toxicon Off. J. Int. Soc. Toxinology 2014, 91, 114–125. [Google Scholar] [CrossRef] [PubMed]
- Brinkman, D.L.; Burnell, J.N. Biochemical and molecular characterisation of cubozoan protein toxins. Toxicon 2009, 54, 1162–1173. [Google Scholar] [CrossRef] [PubMed]
- Klompen, A.M.L.; Kayal, E.; Collins, A.G.; Cartwright, P. Phylogenetic and Selection Analysis of an Expanded Family of Putatively Pore-Forming Jellyfish Toxins (Cnidaria: Medusozoa). Genome Biol. Evol. 2021, 13, 1–17. [Google Scholar] [CrossRef] [PubMed]
- Morabito, R.; La Spada, G.; Crupi, R.; Esposito, E.; Marino, A. Crude Venom from Nematocysts of the Jellyfish Pelagia noctiluca as a Tool to Study Cell Physiology. Cent. Nerv. Syst. Agents Med. Chem. 2015, 15, 68–73. [Google Scholar] [CrossRef] [PubMed]
- Frazão, B.; Antunes, A. Jellyfish Bioactive Compounds: Methods for Wet-Lab Work. Mar. Drugs 2016, 14, 75. [Google Scholar] [CrossRef]
- Page, M.J.; McKenzie, J.E.; Bossuyt, P.M.; Boutron, I.; Hoffmann, T.C.; Mulrow, C.D.; Shamseer, L.; Tetzlaff, J.M.; Akl, E.A.; Brennan, S.E.; et al. The PRISMA 2020 Statement: An Updated Guideline for Reporting Systematic Reviews. BMJ 2021, 372, n71. [Google Scholar] [CrossRef]
- Toom, P.M.; Phillips, T.D. Effects of purified components of jellyfish toxin (Stomolophus meleagris) on active sodium transport. Toxicon 1975, 13, 261–268. [Google Scholar] [CrossRef] [PubMed]
- Brinkman, D.L.; Konstantakopoulos, N.; McInerney, B.V.; Mulvenna, J.; Seymour, J.E.; Isbister, G.K.; Hodgson, W.C. Chironex fleckeri (box jellyfish) venom proteins: Expansion of a cnidarian toxin family that elicits variable cytolytic and cardiovascular effects. J. Biol. Chem. 2014, 289, 4798–4812. [Google Scholar] [CrossRef] [PubMed]
- Brinkman, D.; Burnell, J. Partial purification of cytolytic venom proteins from the box jellyfish, Chironex fleckeri. Toxicon 2008, 51, 853–863. [Google Scholar] [CrossRef]
- Saggiomo, S.L.; Seymour, J.E. Cardiotoxic effects of venom fractions from the Australian box jellyfish Chironex fleckeri on human myocardiocytes. Toxicon 2012, 60, 391–395. [Google Scholar] [CrossRef] [PubMed]
- Endean, R.; Monks, S.A.; Cameron, A.M. Toxins from the box-jellyfish Chironex fleckeri. Toxicon 1993, 31, 397–410. [Google Scholar] [CrossRef] [PubMed]
- Bloom, D.A.; Burnett, J.W.; Alderslade, P. Partial purification of box jellyfish (Chironex fleckeri) nematocyst venom isolated at the beachside. Toxicon 1998, 36, 1075–1085. [Google Scholar] [CrossRef] [PubMed]
- Bloom, D.; Radwan, F.; Burnett, J. Toxinological and immunological studies of capillary electrophoresis fractionated Chrysaora quinquecirrha (Desor) fishing tentacle and Chironex fleckeri Southcott nematocyst venoms. Comp. Biochem. Physiol. Part C Toxicol. Pharmacol. 2001, 128, 75–90. [Google Scholar] [CrossRef]
- Othman, I.; Burnett, J.W. Techniques applicable for purifying Chironex fleckeri (box-jellyfish) venom. Toxicon 1990, 28, 821–835. [Google Scholar] [CrossRef] [PubMed]
- Kintner, A.H.; Seymour, J.E.; Edwards, S.L. Variation in lethality and effects of two Australian chirodropid jellyfish venoms in fish. Toxicon 2005, 46, 699–708. [Google Scholar] [CrossRef] [PubMed]
- Ramasamy, S.; Isbister, G.K.; Seymour, J.E.; Hodgson, W.C. Pharmacologically distinct cardiovascular effects of box jellyfish (Chironex fleckeri) venom and a tentacle-only extract in rats. Toxicol. Lett. 2005, 155, 219–226. [Google Scholar] [CrossRef] [PubMed]
- Ponce, D.; Brinkman, D.L.; Luna-Ramírez, K.; Wright, C.E.; Dorantes-Aranda, J.J. Comparative study of the toxic effects of Chrysaora quinquecirrha (Cnidaria: Scyphozoa) and Chironex fleckeri (Cnidaria: Cubozoa) venoms using cell-based assays. Toxicon 2015, 106, 57–67. [Google Scholar] [CrossRef] [PubMed]
- Cao, C.; Eldefrawi, M.; Eldefrawi, A.; Burnett, J.; Mioduszewski, R.; Menking, D.; Valdes, J. Toxicity of sea nettle toxin to human hepatocytes and the protective effects of phosphorylating and alkylating agents. Toxicon 1998, 36, 269–281. [Google Scholar] [CrossRef] [PubMed]
- Carrette, T.; Seymour, J. A rapid and repeatable method for venom extraction from Cubozoan nematocysts. Toxicon Off. J. Int. Soc. Toxinology 2004, 44, 135–139. [Google Scholar] [CrossRef]
- Yanagihara, A.A.; Shohet, R.V. Cubozoan Venom-Induced Cardiovascular Collapse Is Caused by Hyperkalemia and Prevented by Zinc Gluconate in Mice. PLoS ONE 2012, 7, e51368. [Google Scholar] [CrossRef]
- Pereira, P.; Seymour, J.E. In vitro effects on human heart and skeletal cells of the venom from two cubozoans, Chironex fleckeri and Carukia barnesi. Toxicon 2013, 76, 310–315. [Google Scholar] [CrossRef] [PubMed]
- Long-Rowe, K.O.; Burnett, J.W. Sea nettle (Chrysaora quinquecirrha) lethal factor: Purification by recycling on m-aminophenyl boronic acid acrylic beads. Toxicon 1994, 32, 467–478. [Google Scholar] [CrossRef] [PubMed]
- Long, K.O.; Burnett, J.W. Isolation, characterization, and comparison of hemolytic peptides in nematocyst venoms of two species of jellyfish (Chrysaora quinquecirrha and Cyanea capillata). Comp. Biochem. Physiol. Part B Comp. Biochem. 1989, 94, 641–646. [Google Scholar] [CrossRef]
- Suganthi, K.; Bragadeeswaran, S.; Kumaran, N.S.; Thenmozhi, C.; Thangaraj, S. In vitro antioxidant activities of jelly fish Chrysaora quinquecirrha venom from southeast coast of India. Asian Pac. J. Trop. Biomed. 2012, 2, S347–S351. [Google Scholar] [CrossRef]
- Long-Rowe, K.O.; Burnett, J.W. Characteristics of hyaluronidase and hemolytic activity in fishing tentacle nematocyst venom of Chrysaora quinquecirrha. Toxicon 1994, 32, 165–174. [Google Scholar] [CrossRef]
- Olson, C.E.; Heard, M.G.; Calton, G.J.; Burnett, J.W. Interrelationships between toxins: Studies on the cross-reactivity between bacterial or animal toxins and monoclonal antibodies to two jellyfish venoms. Toxicon 1985, 23, 307–316. [Google Scholar] [CrossRef] [PubMed]
- Burnett, J.W.; Calton, G.J. Sea nettle and man o’war venoms: A chemical comparison of their venoms and studies on the pathogenesis of the sting. J. Investig. Dermatol. 1974, 62, 372–377. [Google Scholar] [CrossRef]
- Russo, A.J.; Calton, G.J.; Burnett, J.W. The relationship of the possible allergic response to jellyfish envenomation and serum antibody titers. Toxicon 1983, 21, 475–480. [Google Scholar] [CrossRef] [PubMed]
- Burnett, J.W.; Long, K.O.; Rubinstein, H.M. Beachside preparation of jellyfish nematocyst tentacles. Toxicon 1992, 30, 794–796. [Google Scholar] [CrossRef] [PubMed]
- Ishikawa, T.; Vucenik, I.; Shamsuddin, A.; Niculescu, F.; Burnett, J.W. Two new actions of sea nettle (Chrysaora quinquecirrha) nematocyst venom: Studies on the mechanism of actions on complement activation and on the central nervous system. Toxicon 2004, 44, 895–899. [Google Scholar] [CrossRef] [PubMed]
- Helmholz, H. Selective toxin–lipid membrane interactions of natural, haemolytic Scyphozoan toxins analyzed by surface plasmon resonance. Biochim. Biophys. Acta (BBA) Biomembr. 2010, 1798, 1944–1952. [Google Scholar] [CrossRef] [PubMed]
- Lassen, S.; Helmholz, H.; Ruhnau, C.; Prange, A. Characterisation of neurotoxic polypeptides from Cyanea capillata medusae (Scyphozoa). Hydrobiologia 2010, 645, 213–221. [Google Scholar] [CrossRef]
- Wang, B.; Liu, D.; Wang, C.; Wang, Q.; Zhang, H.; Liu, G.; Tao, X.; Zhang, L. Mechanism of endothelial nitric oxide synthase phosphorylation and activation by tentacle extract from the jellyfish Cyanea capillata. PeerJ 2017, 5, e3172. [Google Scholar] [CrossRef] [PubMed]
- Wang, B.; Zhang, L.; Zheng, J.; Wang, Q.; Wang, T.; Lu, J.; Wen, X.; Zhang, B.; Liu, G.; Zhang, W.; et al. Multiple organ dysfunction: A delayed envenomation syndrome caused by tentacle extract from the jellyfish Cyanea capillata. Toxicon 2013, 61, 54–61. [Google Scholar] [CrossRef] [PubMed]
- Wang, Q.; Xiao, L.; He, Q.; Liu, S.; Zhang, J.; Li, Y.; Zhang, Z.; Nie, F.; Guo, Y.; Zhang, L. Comparison of haemolytic activity of tentacle-only extract from jellyfish Cyanea capillata in diluted whole blood and erythrocyte suspension: Diluted whole blood is a valid test system for haemolysis study. Exp. Toxicol. Pathol. 2011, 64, 831–835. [Google Scholar] [CrossRef]
- Wang, Q.; Zhang, H.; Wang, B.; Wang, C.; Xiao, L.; Zhang, L. β adrenergic receptor/cAMP/PKA signaling contributes to the intracellular Ca2+ release by tentacle extract from the jellyfish Cyanea capillata. BMC Pharmacol. Toxicol. 2017, 18, 60. [Google Scholar] [CrossRef]
- Wang, T.; He, Q.; Xiao, L.; Wang, Q.; Zhang, B.; Wang, B.; Liu, G.; Zheng, J.; Yu, B.; Zhang, L. Mitochondrial dysfunction contributes to the cytotoxicity induced by tentacle extract from the jellyfish Cyanea capillata in rat renal tubular epithelial NRK-52E cells. Toxicon 2013, 74, 1–7. [Google Scholar] [CrossRef]
- Xiao, L.; He, Q.; Guo, Y.; Zhang, J.; Nie, F.; Li, Y.; Ye, X.; Zhang, L. Cyanea capillata tentacle-only extract as a potential alternative of nematocyst venom: Its cardiovascular toxicity and tolerance to isolation and purification procedures. Toxicon 2009, 53, 146–152. [Google Scholar] [CrossRef] [PubMed]
- Xiao, L.; Liu, S.; He, Q.; Wang, Q.; Ye, X.; Liu, G.; Nie, F.; Zhao, J.; Zhang, L. The Acute Toxicity and Hematological Characterization of the Effects of Tentacle-Only Extract from the Jellyfish Cyanea capillata. Mar. Drugs 2011, 9, 526–534. [Google Scholar] [CrossRef] [PubMed]
- Liang, X.; Beilei, W.; Ying, L.; Qianqian, W.; Sihua, L.; Yang, W.; Guoyan, L.; Jia, L.; Xuting, Y.; Liming, Z. Cardiovascular Effect Is Independent of Hemolytic Toxicity of Tentacle-Only Extract from the Jellyfish Cyanea capillata. PLoS ONE 2012, 7, e43096. [Google Scholar] [CrossRef]
- Xiao, L.; Zhang, J.; Wang, Q.-Q.; He, Q.; Liu, S.-H.; Li, Y.; Zhang, L.-M. In vitro and in vivo haemolytic studies of tentacle-only extract from jellyfish Cyanea capillata. Toxicol. Vitr. 2010, 24, 1203–1207. [Google Scholar] [CrossRef] [PubMed]
- Zhang, H.; Wang, Q.Q.; Xiao, L.; Zhang, L.M. Intervention effects of five cations and their correction on hemolytic activity of tentacle extract from the jellyfish Cyanea capillata. PeerJ 2017, 5, e3338. [Google Scholar] [CrossRef] [PubMed]
- Wang, C.; Wang, B.; Wang, B.; Wang, Q.; Liu, G.; Wang, T.; He, Q.; Zhang, L. Unique Diversity of Sting-Related Toxins Based on Transcriptomic and Proteomic Analysis of the Jellyfish Cyanea capillata and Nemopilema nomurai (Cnidaria: Scyphozoa). J. Proteome Res. 2019, 18, 436–448. [Google Scholar] [CrossRef] [PubMed]
- Helmholz, H.; Ruhnau, C.; Schütt, C.; Prange, A. Comparative study on the cell toxicity and enzymatic activity of two northern scyphozoan species Cyanea capillata (L.) and Cyanea lamarckii (Péron & Léslieur). Toxicon 2007, 50, 53–64. [Google Scholar] [CrossRef] [PubMed]
- Lassen, S.; Helmholz, H.; Ruhnau, C.; Prange, A. A novel proteinaceous cytotoxin from the northern Scyphozoa Cyanea capillata (L.) with structural homology to cubozoan haemolysins. Toxicon 2011, 57, 721–729. [Google Scholar] [CrossRef] [PubMed]
- Li, R.F.; Yu, H.H.; Li, T.; Li, P.C. Comprehensive Proteome Reveals the Key Lethal Toxins in the Venom of Jellyfish Nemopilema nomurai. J. Proteome Res. 2020, 19, 2491–2500. [Google Scholar] [CrossRef] [PubMed]
- Bae, S.K.; Lee, H.; Heo, Y.; Pyo, M.J.; Choudhary, I.; Han, C.H.; Yoon, W.D.; Kang, C.; Kim, E. In vitro characterization of jellyfish venom fibrin(ogen)olytic enzymes from Nemopilema nomurai. J. Venom. Anim. Toxins Incl. Trop. Dis. 2017, 23, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Pang, M.; Xu, J.T.; Liu, Y.L.; Zhang, X.L. Comparative study of the hemolytic and cytotoxic activities of nematocyst venoms from the jellyfish Cyanea nozakii Kishinouye and Nemopilema nomurai Kishinouye. J. Oceanol. Limnol. 2018, 36, 1255–1265. [Google Scholar] [CrossRef]
- Yu, H.; Yue, Y.; Dong, X.; Li, R.; Li, P. The Acaricidal Activity of Venom from the Jellyfish Nemopilema nomurai against the Carmine Spider Mite Tetranychus cinnabarinus. Toxins 2016, 8, 179. [Google Scholar] [CrossRef] [PubMed]
- Kang, C.; Kim, Y.K.; Lee, H.; Cha, M.; Sohn, E.-T.; Jung, E.-S.; Song, C.; Kim, M.; Lee, H.C.; Kim, J.-S.; et al. Target organ identification of jellyfish envenomation using systemic and integrative analyses in anesthetized dogs. J. Pharmacol. Toxicol. Methods 2011, 64, 173–179. [Google Scholar] [CrossRef]
- Kang, C.; Munawir, A.; Cha, M.; Sohn, E.-T.; Lee, H.; Kim, J.-S.; Yoon, W.D.; Lim, D.; Kim, E. Cytotoxicity and hemolytic activity of jellyfish Nemopilema nomurai (Scyphozoa: Rhizostomeae) venom. Comp. Biochem. Physiol. Part C Toxicol. Pharmacol. 2009, 150, 85–90. [Google Scholar] [CrossRef] [PubMed]
- Prakash, R.L.M.; Hwang, D.H.; Hong, I.-H.; Chae, J.; Kang, C.; Kim, E. Danio rerio as an alternative vertebrate model for jellyfish venom study: The toxinological aspects of Nemopilema nomurai venom. Toxicol. Lett. 2020, 335, 91–97. [Google Scholar] [CrossRef] [PubMed]
- Hwang, D.H.; Lee, H.; Choudhary, I.; Kang, C.; Chae, J.; Kim, E. Protective effect of epigallocatechin-3-gallate (EGCG) on toxic metalloproteinases-mediated skin damage induced by Scyphozoan jellyfish envenomation. Sci. Rep. 2020, 10, 1–10. [Google Scholar] [CrossRef]
- Yue, Y.; Yu, H.; Li, R.; Xing, R.; Liu, S.; Li, K.; Wang, X.; Chen, X.; Li, P. Biochemical and kinetic evaluation of the enzymatic toxins from two stinging scyphozoans Nemopilema nomurai and Cyanea nozakii. Toxicon 2017, 125, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Ayed, Y.; Bousabbeh, M.; Mabrouk, H.B.; Morjen, M.; Marrakchi, N.; Bacha, H. Impairment of the cell-to-matrix adhesion and cytotoxicity induced by the Mediterranean jellyfish Pelagia noctiluca venom and its fractions in cultured glioblastoma cells. Lipids Heal. Dis. 2012, 11, 84. [Google Scholar] [CrossRef] [PubMed]
- Ayed, Y.; Dellai, A.; Ben Mansour, H.; Bacha, H.; Abid, S. Analgesic and antibutyrylcholinestrasic activities of the venom prepared from the Mediterranean jellyfish Pelagia noctiluca (Forsskal, 1775). Ann. Clin. Microbiol. Antimicrob. 2012, 11, 15. [Google Scholar] [CrossRef] [PubMed]
- Sánchez-Rodríguez, J.; Lucio-Martínez, N. Isolation and prepurification of active compounds in venom from Pelagia noctiluca (Scyphozoa: Pelagiidae) from the Caribbean Sea. Cienc. Mar. 2011, 37, 369–377. [Google Scholar] [CrossRef]
- Ayed, Y.; Boussabbeh, M.; Zakhama, W.; Bouaziz, C.; Abid, S.; Bacha, H. Induction of cytotoxicity of Pelagia noctiluca venom causes reactive oxygen species generation, lipid peroxydation induction and DNA damage in human colon cancer cells. Lipids Heal. Dis. 2011, 10, 232. [Google Scholar] [CrossRef]
- Marino, A.; Morabito, R.; Pizzata, T.; La Spada, G. Effect of crude venom from nematocysts of Pelagia noctiluca (Scyphozoa) on spread discharge of acontia of Calliactis parasitica (Anthozoa). Chem. Ecol. 2008, 24, 9–17. [Google Scholar] [CrossRef]
- Marino, A.; Morabito, R.; Pizzata, T.; La Spada, G. Effect of various factors on Pelagia noctiluca (Cnidaria, Scyphozoa) crude venom-induced haemolysis. Comp. Biochem. Physiol. Part A Mol. Integr. Physiol. 2008, 151, 144–149. [Google Scholar] [CrossRef] [PubMed]
- Marino, A.; Crupi, R.; Rizzo, G.; Morabito, R.; Musci, G.; LA Spada, G. The unusual toxicity and stability properties of crude venom from isolated nematocysts of Pelagia noctiluca (Cnidaria, Scyphozoa). Cell. Mol. Biol. 2007, 53, 994–1002. [Google Scholar]
- Liu, X.; Zhang, M.S.; Zhang, C.; Liu, C.H. Angiotensin converting enzyme (ACE) inhibitory, antihypertensive and antihyperlipidaemic activities of protein hydrolysates from Rhopilema esculentum. Food Chem. 2012, 134, 2134–2140. [Google Scholar] [CrossRef] [PubMed]
- Li, C.P.; Yu, H.H.; Liu, S.; Xing, R.E.; Guo, Z.Y.; Li, P.C. Factors affecting the protease activity of venom from jellyfish Rhopilema esculentum Kishinouye. Bioorganic Med. Chem. Lett. 2005, 15, 5370–5374. [Google Scholar] [CrossRef] [PubMed]
- Feng, J.; Yu, H.; Li, C.; Xing, R.; Liu, S.; Wang, L.; Cai, S.; Li, P. Isolation and characterization of venom from nematocysts of jellyfish Rhopilema esculentum Kishinouye. Chin. J. Oceanol. Limnol. 2009, 27, 869–874. [Google Scholar] [CrossRef]
- Li, C.; Yu, H.; Feng, J.; Chen, X.; Li, P. Comparative analysis of methods for concentrating venom from jellyfish Rhopilema esculentum Kishinouye. Chin. J. Oceanol. Limnol. 2009, 27, 172–176. [Google Scholar] [CrossRef]
- Yu, H.H.; Xing, R.E.; Liu, S.; Li, C.P.; Guo, Z.Y.; Li, P.C. Studies on the hemolytic activity of tentacle extracts of jellyfish Rhopilema esculentum Kishinouye: Application of orthogonal test. Int. J. Biol. Macromol. 2007, 40, 276–280. [Google Scholar] [CrossRef] [PubMed]
- Yu, H.; Liu, X.; Dong, X.; Li, C.; Xing, R.; Liu, S.; Li, P. Insecticidal activity of proteinous venom from tentacle of jellyfish Rhopilema esculentum Kishinouye. Bioorganic Med. Chem. Lett. 2005, 15, 4949–4952. [Google Scholar] [CrossRef]
- Reinicke, J.; Kitatani, R.; Masoud, S.S.; Galbraith, K.K.; Yoshida, W.; Igarashi, A.; Nagasawa, K.; Berger, G.; Yanagihara, A.; Nagai, H.; et al. Isolation, Structure Determination, and Synthesis of Cyclic Tetraglutamic Acids from Box Jellyfish Species Alatina alata and Chironex yamaguchii. Molecules 2020, 25, 883. [Google Scholar] [CrossRef] [PubMed]
- Nagai, H.; Takuwa, K.; Nakao, M.; Ito, E.; Miyake, M.; Noda, M.; Nakajima, T. Novel Proteinaceous Toxins from the Box Jellyfish (Sea Wasp) Carybdea rastoni. Biochem. Biophys. Res. Commun. 2000, 275, 582–588. [Google Scholar] [CrossRef] [PubMed]
- Azuma, H.; Sekizaki, S.; Satoh, A.; Nakajima, T. Platelet Aggregation Caused by Carybdea Rastonii Toxins (CrTX-I, II, and III) Obtained from a Jellyfish, Carybdea rastonii. Exp. Biol. Med. 1986, 182, 34–42. [Google Scholar] [CrossRef]
- Azuma, H.; Sekizaki, S.; Satoh, A.; Nakajima, T.; Ishikawa, M. Platelet aggregation caused by a partially purified jellyfish toxin from Carybdea rastonii. Toxicon 1986, 24, 489–499. [Google Scholar] [CrossRef] [PubMed]
- Rottini, G.; Gusmani, L.; Parovel, E.; Avian, M.; Patriarca, P. Purification and properties of a cytolytic toxin in venom of the jellyfish Carybdea marsupialis. Toxicon 1995, 33, 315–326. [Google Scholar] [CrossRef] [PubMed]
- Li, R.; Yu, H.; Xing, R.; Liu, S.; Qing, Y.; Li, K.; Li, B.; Meng, X.; Cui, J.; Li, P. Isolation, identification and characterization of a novel antioxidant protein from the nematocyst of the jellyfish Stomolophus meleagris. Int. J. Biol. Macromol. 2012, 51, 274–278. [Google Scholar] [CrossRef]
- Li, R.; Yu, H.; Xing, R.; Liu, S.; Qing, Y.; Li, K.; Li, B.; Meng, X.; Cui, J.; Li, P. Isolation and in vitro partial characterization of hemolytic proteins from the nematocyst venom of the jellyfish Stomolophus meleagris. Toxicol. Vitr. 2013, 27, 1620–1625. [Google Scholar] [CrossRef] [PubMed]
- Nagai, H.; Takuwa, K.; Nakao, M.; Sakamoto, B.; Crow, G.L.; Nakajima, T. Isolation and Characterization of a Novel Protein Toxin from the Hawaiian Box Jellyfish (Sea Wasp) Carybdea alata. Biochem. Biophys. Res. Commun. 2000, 275, 589–594. [Google Scholar] [CrossRef]
- Chung, J.J.; Ratnapala, L.A.; Cooke, I.M.; Yanagihara, A.A. Partial purification and characterization of a hemolysin (CAH1) from Hawaiian box jellyfish (Carybdea alata) venom. Toxicon 2001, 39, 981–990. [Google Scholar] [CrossRef] [PubMed]
- Lotan, A.; Fishman, L.; Zlotkin, E. Toxin compartmentation and delivery in the Cnidaria: The nematocyst’s tubule as a multiheaded poisonous arrow. J. Exp. Zool. 1996, 275, 444–451. [Google Scholar] [CrossRef] [PubMed]
- Gusmani, L.; Avian, M.; Galil, B.; Patriarca, P.; Rottini, G. Biologically active polypeptides in the venom of the jellyfish Rhopilema nomadica. Toxicon 1997, 35, 637–648. [Google Scholar] [CrossRef] [PubMed]
- Liu, X.; Zhang, M.S.; Shi, Y.P.; Qiao, R.J.; Tang, W.; Sun, Z.L. Production of the angiotensin I converting enzyme inhibitory peptides and isolation of four novel peptides from jellyfish (Rhopilema esculentum) protein hydrolysate. J. Sci. Food Agric. 2016, 96, 3240–3248. [Google Scholar] [CrossRef] [PubMed]
- Alam, J.M.; Sultana, I.; Mahmood, S.R.; Alam, S.M. Pneumotoxic activity of crude venom and a cytolytic protein, PuTX-IVC, from a coelenterate, Physalia utriculus (Blue Bottle). Pak. J. Zool. 2006, 38, 159–165. [Google Scholar]
- Diaz-Garcia, C.; Fuentes-Silva, D.; Sanchez-Soto, C.; Pérez, D.D.; Garcia-Delgado, N.; Varela, C.; Mendoza-Hernandez, G.; Rodriguez-Romero, A.; Castaneda, O.; Hiriart, M. Toxins from Physalia physalis (Cnidaria) Raise the Intracellular Ca2+ of Beta-Cells and Promote Insulin Secretion. Curr. Med. Chem. 2012, 19, 5414–5423. [Google Scholar] [CrossRef] [PubMed]
- Gaur, P.K.; Anthony, R.L.; Calton, G.J.; Burnett, J.W. Isolation of hybridomas secreting monoclonal antibodies against Physalia physalis (Portuguese man-o’war) nematocyst venom. Toxicon 1982, 20, 419–425. [Google Scholar] [CrossRef] [PubMed]
- Burnett, J.W.; Ordonez, J.V.; Calton, G.J. Differential toxicity of Physalia physalis (Portuguese man-o’war) nematocysts separated by flow cytometry. Toxicon 1986, 24, 514–518. [Google Scholar] [CrossRef] [PubMed]
- Helmholz, H.; Naatz, S.; Lassen, S.; Prange, A. Isolation of a cytotoxic glycoprotein from the Scyphozoa Cyanea lamarckii by lectin-affinity chromatography and characterization of molecule interactions by surface plasmon resonance. J. Chromatogr. B 2008, 871, 60–66. [Google Scholar] [CrossRef] [PubMed]
- Radwan, F.F.; Burnett, J.W.; Bloom, D.A.; Coliano, T.; Eldefrawi, M.E.; Erderly, H.; Aurelian, L.; Torres, M.; la Cotera, E.P.H.-D. A comparison of the toxinological characteristics of two Cassiopea and Aurelia species. Toxicon 2000, 39, 245–257. [Google Scholar] [CrossRef]
- Ponce, D.; López-Vera, E.; Aguilar, M.B.; Sanchez-Rodriguez, J. Preliminary Results of the in Vivo and in Vitro Characterization of a Tentacle Venom Fraction from the Jellyfish Aurelia aurita. Toxins 2013, 5, 2420–2433. [Google Scholar] [CrossRef]
- Rastogi, A.; Biswas, S.; Sarkar, A.; Chakrabarty, D. Anticoagulant activity of Moon jellyfish (Aurelia aurita) tentacle extract. Toxicon 2012, 60, 719–723. [Google Scholar] [CrossRef] [PubMed]
- Ghosh, T.K.; Gomes, A.; Nag Chaudhuri, A.K. Isolation of a toxin from jellyfish Acromitus rabanchatu and its effect on skeletal muscle. Toxicon 1993, 31, 873–880. [Google Scholar] [CrossRef] [PubMed]
- Rastogi, A.; Sarkar, A.; Chakrabarty, D. Partial purification and identification of a metalloproteinase with anticoagulant activity from Rhizostoma pulmo (Barrel Jellyfish). Toxicon Off. J. Int. Soc. Toxinology 2017, 132, 29–39. [Google Scholar] [CrossRef] [PubMed]
- Azuma, H.; Ishikawa, M.; Nakajima, T.; Satoh, A.; Sekizaki, S. Calcium-dependent contractile response of arterial smooth muscle to a jellyfish toxin (pCrTX: Carybdea rastonii). Br. J. Pharmacol. 1986, 88, 549–559. [Google Scholar] [CrossRef]
- Zhang, M.; Fishman, Y.; Sher, D.; Zlotkin, E. Hydralysin, a novel animal group-selective paralytic and cytolytic protein from a noncnidocystic origin in hydra. Biochemistry 2003, 42, 8939–8944. [Google Scholar] [CrossRef] [PubMed]
- Lazcano-Pérez, F.; Arellano, R.O.; Garay, E.; Arreguín-Espinosa, R.; Sánchez-Rodríguez, J. Electrophysiological activity of a neurotoxic fraction from the venom of box jellyfish Carybdea marsupialis. Comp. Biochem. Physiol. Part C Toxicol. Pharmacol. 2017, 191, 177–182. [Google Scholar] [CrossRef] [PubMed]
- Radwan, F.F.Y.; Roman, L.G.; Baksi, K.; Burnett, J.W. Toxicity and mAChRs binding activity of Cassiopea xamachana venom from Puerto Rican coasts. Toxicon 2005, 45, 107–112. [Google Scholar] [CrossRef] [PubMed]
- Radwan, F.F.; Burnett, J.W. Toxinological studies of the venom from Cassiopea xamachana nematocysts isolated by flow cytometry. Comp. Biochem. Physiol. Part C: Toxicol. Pharmacol. 2001, 128, 65–73. [Google Scholar] [CrossRef]
- Torres, M.; Aguilar, M.B.; Falcón, A.; Sánchez, L.; Radwan, F.F.; Burnett, J.W.; la Cotera, E.P.H.-D.; Arellano, R.O. Electrophysiological and hemolytic activity elicited by the venom of the jellyfish Cassiopea xamachana. Toxicon 2001, 39, 1297–1307. [Google Scholar] [CrossRef]
- Azila, N.; Siao, F.K.; Othman, I. Haemolytic, oedema and haemorrhage inducing activities of tentacular extract of the blubber jellyfish (Catostylus mosaicus). Comp. Biochem. Physiol. Part C Comp. Pharmacol. 1991, 99, 153–156. [Google Scholar] [CrossRef]
- Wiltshire, C.J.; Sutherland, S.K.; Fenner, P.J.; Young, A.R. Optimization and preliminary characterization of venom isolated from 3 medically important jellyfish: The box (Chironex fleckeri), Irukandji (Carukia barnesi), and blubber (Catostylus mosaicus) jellyfish. Wilderness Environ. Med. 2000, 11, 241–250. [Google Scholar] [CrossRef] [PubMed]
- So, P.B.T.; Rubio, P.; Lirio, S.; Macabeo, A.P.; Huang, H.-Y.; Corpuz, M.J.-A.T.; Villaflores, O.B. In vitro angiotensin I converting enzyme inhibition by a peptide isolated from Chiropsalmus quadrigatus Haeckel (box jellyfish) venom hydrolysate. Toxicon 2016, 119, 77–83. [Google Scholar] [CrossRef] [PubMed]
- Nagai, H.; Takuwa-Kuroda, K.; Nakao, M.; Oshiro, N.; Iwanaga, S.; Nakajima, T. A Novel Protein Toxin from the Deadly Box Jellyfish (Sea Wasp, Habu-kurage) Chiropsalmus quadrigatus. Biosci. Biotechnol. Biochem. 2002, 66, 97–102. [Google Scholar] [CrossRef] [PubMed]
- Horiike, T.; Nagai, H.; Kitani, S. Identification of Allergens in the Box Jellyfish Chironex yamaguchii That Cause Sting Dermatitis. Int. Arch. Allergy Immunol. 2015, 167, 73–82. [Google Scholar] [CrossRef] [PubMed]
- Li, R.; Yu, H.; Yue, Y.; Li, P. Combined Proteome and Toxicology Approach Reveals the Lethality of Venom Toxins from Jellyfish Cyanea nozakii. J. Proteome Res. 2018, 17, 3904–3913. [Google Scholar] [CrossRef]
- Li, R.; Yu, H.; Feng, J.; Xing, R.; Liu, S.; Wang, L.; Qin, Y.; Li, K.; Li, P. Two-step purification and in vitro characterization of a hemolysin from the venom of jellyfish Cyanea nozakii Kishinouye. Int. J. Biol. Macromol. 2011, 49, 14–19. [Google Scholar] [CrossRef]
- Feng, J.; Yu, H.; Li, C.; Xing, R.; Liu, S.; Wang, L.; Cai, S.; Li, P. Isolation and characterization of lethal proteins in nematocyst venom of the jellyfish Cyanea nozakii Kishinouye. Toxicon 2010, 55, 118–125. [Google Scholar] [CrossRef]
- Li, C.P.; Li, P.C.; Feng, J.H.; Li, R.F.; Yu, H.H. Cytotoxicity of the venom from the nematocysts of jellyfish Cyanea nozakii Kishinouye. Toxicol. Ind. Health 2012, 28, 186–192. [Google Scholar] [CrossRef] [PubMed]
- De Domenico, S.; De Rinaldis, G.; Paulmery, M.; Piraino, S.; Leone, A. Barrel Jellyfish (Rhizostoma pulmo) as Source of Antioxidant Peptides. Mar. Drugs 2019, 17, 134. [Google Scholar] [CrossRef]
- Winkel, K.D.; Tibballs, J.; Molenaar, P.; Lambert, G.; Coles, P.; Ross-Smith, M.; Wiltshire, C.; Fenner, P.J.; Gershwin, L.-A.; Hawdon, G.M. Cardiovascular actions of the venom from the Irukandji (Carukia Barnesi) jellyfish: Effects in human, rat and guinea-pig tissues in vitro and in pigs in vitro. Clin. Exp. Pharmacol. Physiol. 2005, 32, 777–788. [Google Scholar] [CrossRef]
- Nabipour, I.; Mohebbi, G.; Vatanpour, H.; Vazirizadeh, A. Hematological parameters on the effect of the jellyfish venom Cassiopea andromeda in animal models. Data Brief 2017, 11, 517–521. [Google Scholar] [CrossRef] [PubMed]
- Mohebbi, G.; Farzadinia, P.; Vatanpour, H.; Hashemi, A.; Vazirizadeh, A.; Khazaei, Z. Sub-acute toxicity of the alien Cassiopea andromeda (forsskal, 1775) jellyfish venom, in rats. Entomol. Appl. Sci. Lett. 2016, 3, 65–71. [Google Scholar]
- Qu, X.; Fan, L.; Zhong, T.; Li, G.; Xia, X.; Long, H.; Huang, D.; Shu, W. The nematocysts venom of Chrysaora helvola Brandt leads to apoptosis-like cell death accompanied by uncoupling of oxidative phosphorylation. Toxicon 2016, 110, 74–78. [Google Scholar] [CrossRef]
- Qu, X.; Xia, X.; Lai, Z.; Zhong, T.; Li, G.; Fan, L.; Shu, W. Apoptosis-like cell death induced by nematocyst venom from Chrysaora helvola Brandt jellyfish and an in vitro evaluation of commonly used antidotes. Comp. Biochem. Physiol. Part C Toxicol. Pharmacol. 2016, 180, 31–39. [Google Scholar] [CrossRef]
- Kim, H.-J.; Noh, J.-W.; Amarsanaa, K.; Jeon, S.-C.; Yang, Y.-S.; Hwang, N.-H.; Ko, E.-A.; Kang, Y.-J.; Jung, S.-C. Peripheral Pain Modulation of Chrysaora pacifica Jellyfish Venom Requires Both Ca2+ Influx and TRPA1 Channel Activation in Rats. Neurotox. Res. 2020, 38, 900–913. [Google Scholar] [CrossRef]
- Carneiro, R.F.V.; Nascimento, N.R.F.D.; Costa, P.P.C.; Gomes, V.M.; de Souza, A.J.F.; de Oliveira, S.C.B.; Filho, E.B.D.S.D.; Zara, F.J.; Fonteles, M.C.; Toyama, D.D.O.; et al. The extract of the jellyfish Phyllorhiza punctata promotes neurotoxic effects. J. Appl. Toxicol. 2011, 31, 720–729. [Google Scholar] [CrossRef]
- Knittel, P.S.; Long, P.F.; Brammall, L.; Marques, A.C.; Almeida, M.T.; Padilla, G.; Moura-Da-Silva, A.M. Characterising the enzymatic profile of crude tentacle extracts from the South Atlantic jellyfish Olindias sambaquiensis (Cnidaria: Hydrozoa). Toxicon 2016, 119, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Becerra-Amezcua, M.P.; Guerrero-Legarreta, I.; Gonzalez-Marquez, H.; Guzman-Garcia, X. In vivo analysis of effects of venom from the jellyfish Chrysaora sp. in zebrafish (Danio rerio). Toxicon 2016, 113, 49–54. [Google Scholar] [CrossRef] [PubMed]
- Yang, F.; Ye, R.; Ma, C.; Wang, Y.; Wang, Y.; Chen, J.; Yang, J.; Höfer, J.; Zhu, Y.; Xiao, L.; et al. Toxicity evaluation, toxin screening and its intervention of the jellyfish Phacellophora camtschatica based on a combined transcriptome-proteome analysis. Ecotoxicol. Environ. Saf. 2022, 233, 113315. [Google Scholar] [CrossRef] [PubMed]
- Yue, Y.; Yu, H.; Li, R.; Liu, S.; Xing, R.; Li, P. Insights into individual variations in nematocyst venoms from the giant jellyfish Nemopilema nomurai in the Yellow Sea. Sci. Rep. 2019, 9, 3361. [Google Scholar] [CrossRef] [PubMed]
- Yu, C.; Yu, H.; Li, P. Highlights of animal venom research on the geographical variations of toxin components, toxicities and envenomation therapy. Int. J. Biol. Macromol. 2020, 165, 2994–3006. [Google Scholar] [CrossRef] [PubMed]




Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).