Antifungal and Antibacterial Activities of Isolated Marine Compounds

 ,
,  , ,
, ,  ,
,  and
and 
Abstract
1. Introduction
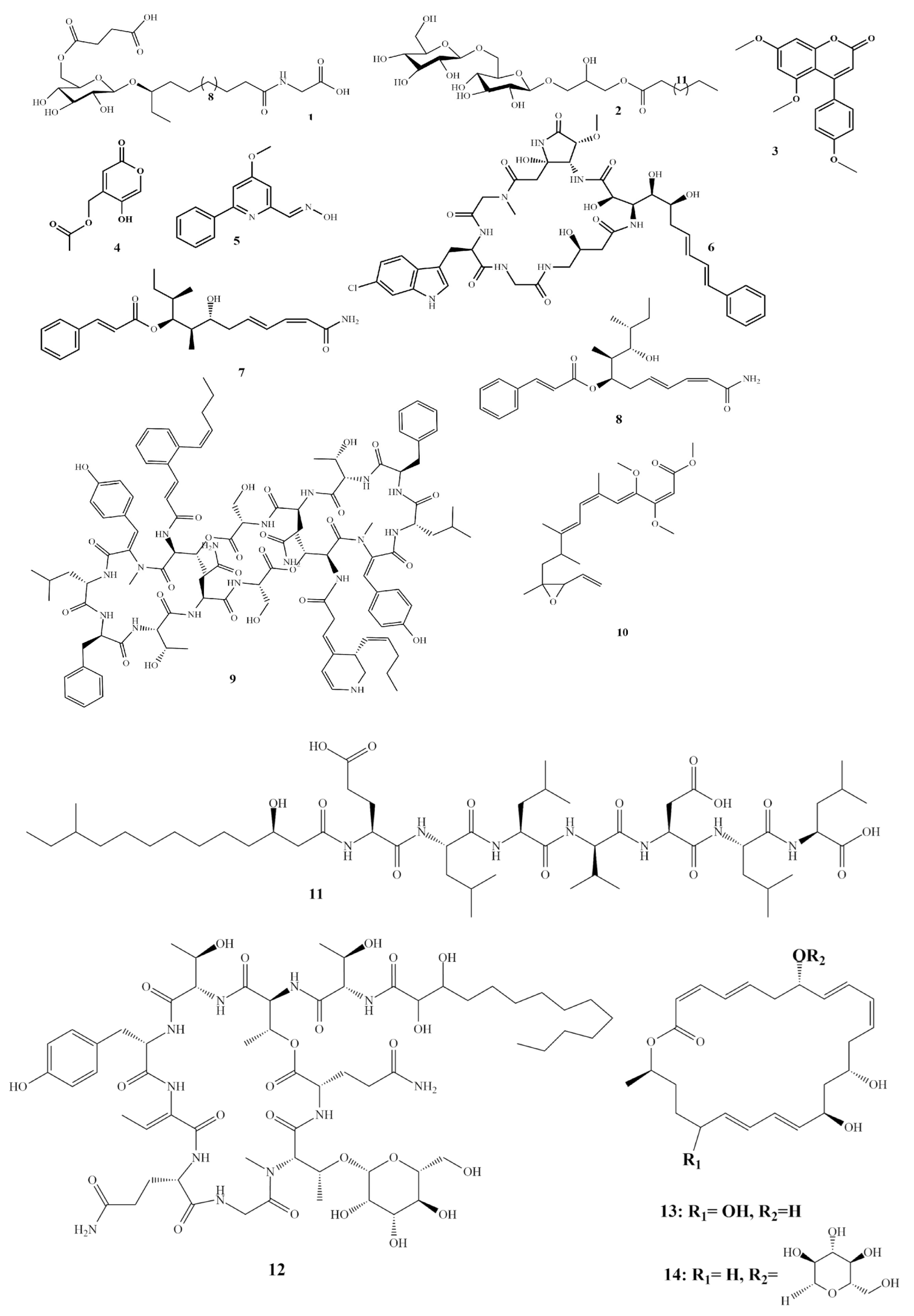
2. Isolated Marine Compounds with Antifungal Activity
2.1. Antifungal Compounds Isolated from Marine Bacteria
2.2. Antifungal Compounds Isolated from Marine Fungi
2.3. Antifungal Compounds Isolated from Marine Algae
2.4. Antifungal Compounds Isolated from Sea Cucumbers
2.5. Antifungal Compounds Isolated from Sea Sponges
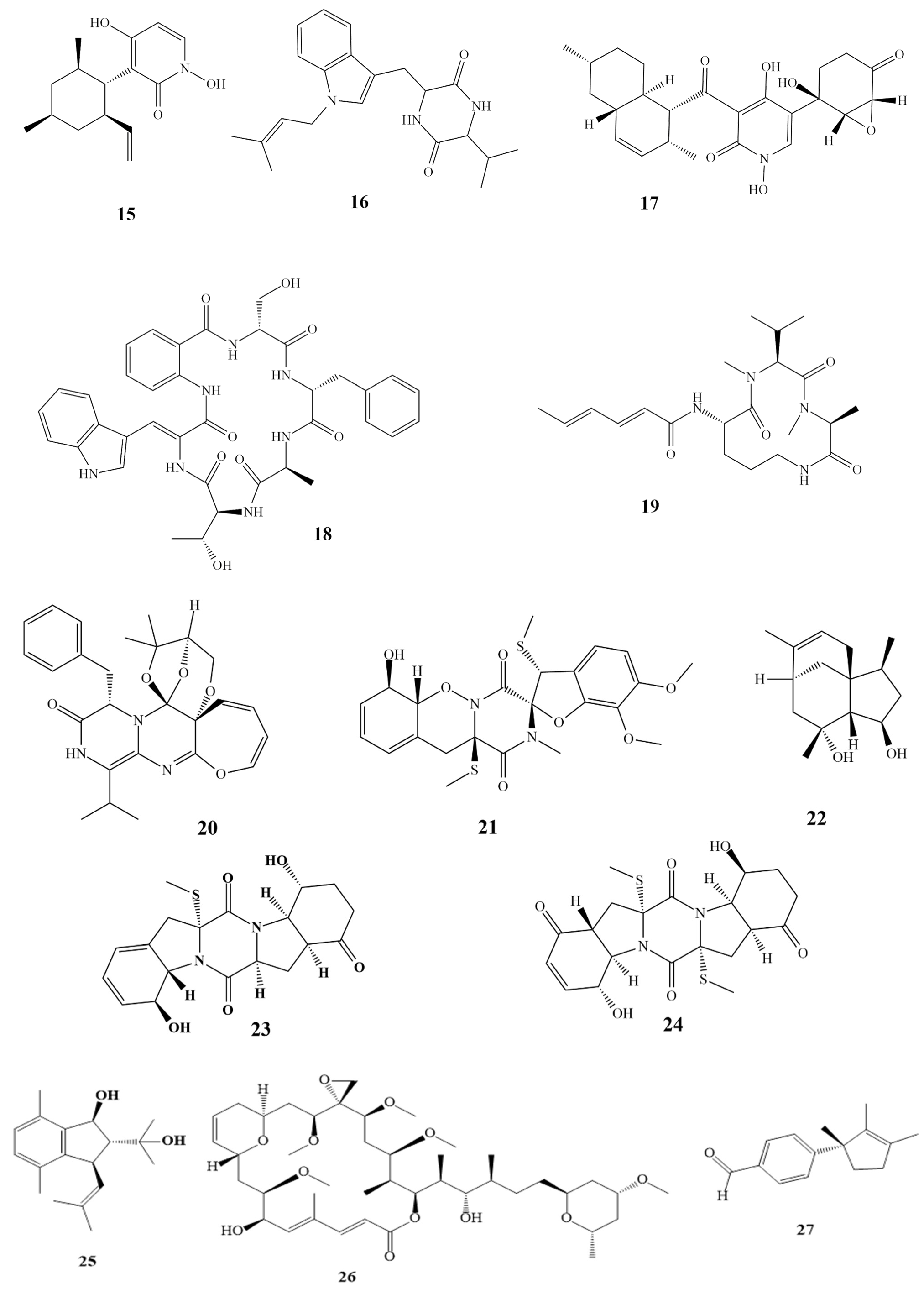
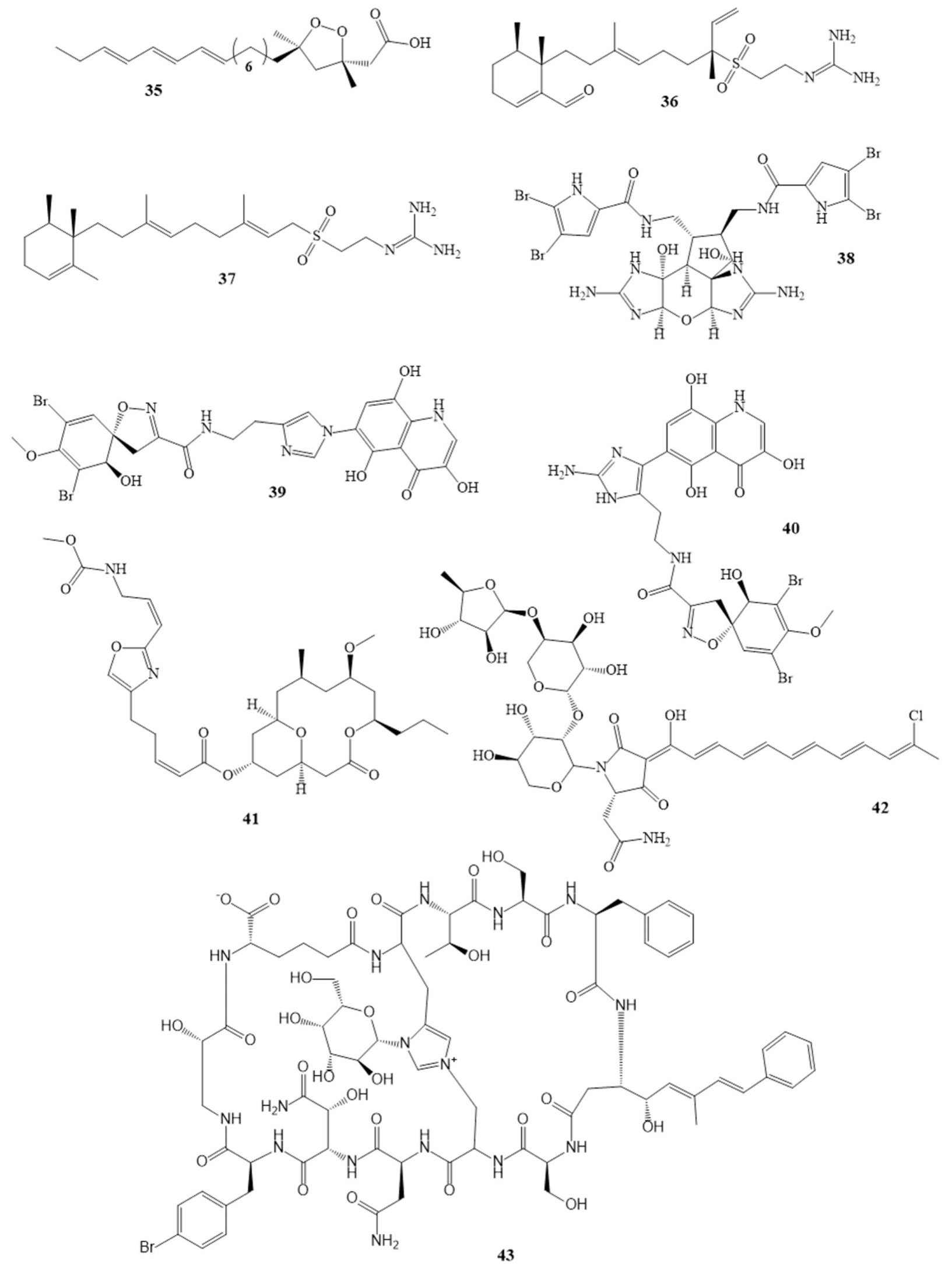
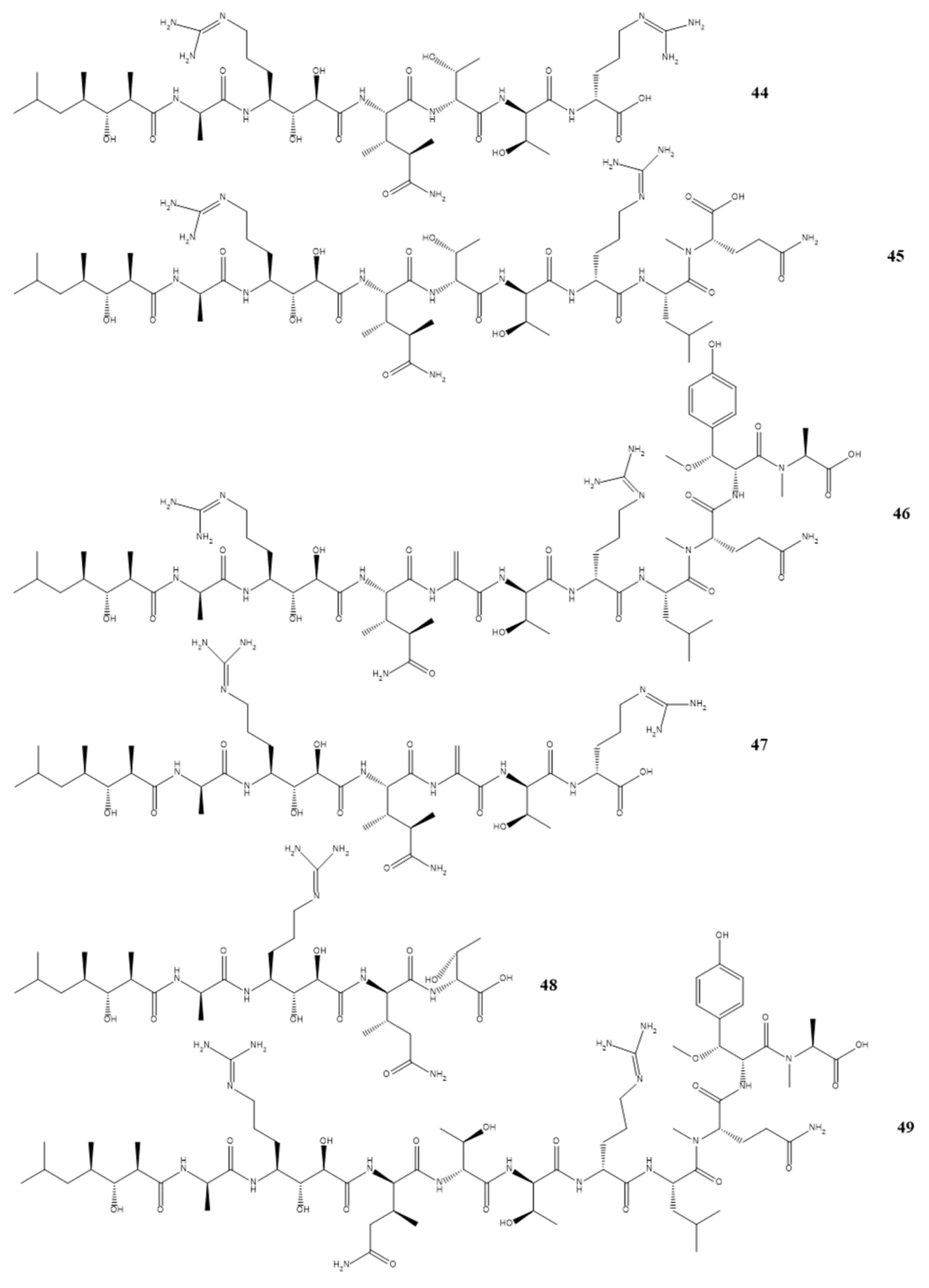
3. Isolated Marine Compounds with Antibacterial Activity
3.1. Ribosomal Peptides—Antimicrobial Peptides
3.2. Nonribosomal Peptides
3.3. Polyketides
3.4. Alkaloids
3.5. Terpenes
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Centers for Disease Control and Prevention. Antibiotic Resistance Threats in the United States, 2019; US Department of Health and Human Services, CDC: Atlanta, GA, USA, 2019.
- Okada, B.K.; Seyedsayamdost, M.R. Antibiotic dialogues: Induction of silent biosynthetic gene clusters by exogenous small molecules. FEMS Microbiol. Rev. 2017, 41, 19–33. [Google Scholar] [CrossRef]
- Farha, M.A.; Brown, E.D. Strategies for target identification of antimicrobial natural products. Nat. Prod. Rep. 2016, 33, 668–680. [Google Scholar] [CrossRef] [PubMed]
- Bengtsson-Palme, J.; Kristiansson, E.; Larsson, D.J. Environmental factors influencing the development and spread of antibiotic resistance. FEMS Microbiol. Rev. 2018, 42, fux053. [Google Scholar] [CrossRef] [PubMed]
- Fair, R.J.; Tor, Y. Antibiotics and bacterial resistance in the 21st century. Perspect. Med. Chem. 2014, 6, S14459. [Google Scholar] [CrossRef] [PubMed]
- Velmurugan, P.; Venil, C.K.; Veera Ravi, A.; Dufossé, L. Marine bacteria are the cell factory to produce bioactive pigments: A prospective pigment source in the ocean. Front. Sustain. Food Syst. 2020, 4, 589655. [Google Scholar] [CrossRef]
- Gomes, N.G.; Dasari, R.; Chandra, S.; Kiss, R.; Kornienko, A. Marine invertebrate metabolites with anticancer activities: Solutions to the “supply problem”. Mar. Drugs 2016, 14, 98. [Google Scholar] [CrossRef]
- Fan, Y.; Wang, Y.; Liu, P.; Fu, P.; Zhu, T.; Wang, W.; Zhu, W. Indole-diterpenoids with anti-H1N1 activity from the aciduric fungus Penicillium camemberti OUCMDZ-1492. J. Nat. Prod. 2013, 76, 1328–1336. [Google Scholar] [CrossRef]
- Gong, K.K.; Tang, X.L.; Zhang, G.; Cheng, C.L.; Zhang, X.W.; Li, P.L.; Li, G.Q. Polyhydroxylated steroids from the South China Sea soft coral Sarcophyton sp. and their cytotoxic and antiviral activities. Mar. Drugs 2013, 11, 4788–4798. [Google Scholar] [CrossRef]
- Manivasagan, P.; Venkatesan, J.; Sivakumar, K.; Kim, S.K. Pharmaceutically active secondary metabolites of marine actinobacteria. Microbiol. Res. 2014, 169, 262–278. [Google Scholar] [CrossRef]
- Plouguerné, E.; De Souza, L.M.; Sassaki, G.L.; Cavalcanti, J.F.; Romanos, M.T.V.; Da Gama, B.A.; Barreto-Bergter, E. Antiviral sulfoquinovosyldiacylglycerols (SQDGs) from the Brazilian brown seaweed Sargassum vulgare. Mar. Drugs 2013, 11, 4628–4640. [Google Scholar] [CrossRef]
- Tajima, H.; Wakimoto, T.; Takada, K.; Ise, Y.; Abe, I. Revised structure of cyclolithistide A, a cyclic depsipeptide from the marine sponge Discodermia japonica. J. Nat. Prod. 2014, 77, 154–158. [Google Scholar] [CrossRef] [PubMed]
- Wang, W.; Wang, S.X.; Guan, H.S. The antiviral activities and mechanisms of marine polysaccharides: An overview. Mar. Drugs 2012, 10, 2795–2816. [Google Scholar] [CrossRef] [PubMed]
- Malve, H. Exploring the ocean for new drug developments: Marine pharmacology. J. Pharm. Bioallied Sci. 2016, 8, 83–91. [Google Scholar] [CrossRef] [PubMed]
- Venkatesan, J.; Anil, S.; Kim, S.K.; Shim, M.S. Marine fish proteins and peptides for cosmeceuticals: A review. Mar. Drugs 2017, 15, 143. [Google Scholar] [CrossRef]
- Williams, P.G. Panning for chemical gold: Marine bacteria as a source of new therapeutics. Trends Biotechnol. 2009, 27, 45–52. [Google Scholar] [CrossRef]
- Gulder, T.A.; Moore, B.S. Chasing the treasures of the sea—Bacterial marine natural products. Curr. Opin. Microbiol. 2009, 12, 252–260. [Google Scholar] [CrossRef]
- Joseph, A. Oceans: Abode of Nutraceuticals, Pharmaceuticals, and Biotoxins. In Investigating Seafloors and Oceans; Joseph, A., Ed.; Candice Janco: Goa, India, 2016; pp. 493–554. ISBN 9780128093573. [Google Scholar]
- Tareq, F.S.; Lee, H.S.; Lee, Y.J.; Lee, J.S.; Shin, H.J. Ieodoglucomide C and Ieodoglycolipid, New Glycolipids from a Marine-Derived Bacterium Bacillus licheniformis 09IDYM23. Lipids 2015, 50, 513–519. [Google Scholar] [CrossRef]
- Choudhary, A.; Naughton, L.M.; Montánchez, I.; Dobson, A.D.; Rai, D.K. Current status and future prospects of marine natural products (MNPs) as antimicrobials. Mar. Drugs 2017, 15, 272. [Google Scholar] [CrossRef] [PubMed]
- El-Gendy, M.M.; El-Bondkly, A.M. Production and genetic improvement of a novel antimycotic agent, saadamycin, against dermatophytes and other clinical fungi from endophytic Streptomyces sp. Hedaya48. J. Ind. Microbiol. Biotechnol. 2010, 37, 831–841. [Google Scholar] [CrossRef] [PubMed]
- Gouda, S.; Das, G.; Sen, S.K.; Shin, H.S.; Patra, J.K. Endophytes: A treasure house of bioactive compounds of medicinal importance. Front. Microbiol. 2016, 7, 1538. [Google Scholar] [CrossRef]
- Ambavane, V.; Tokdar, P.; Parab, R.; Sreekumar, E.S.; Mahajan, G.B.; Mishra, P.D.; Ranadive, P. Caerulomycin A—An antifungal compound isolated from marine actinomycetes. Adv. Microbiol. 2014, 4, 567–578. [Google Scholar] [CrossRef]
- El-Hossary, E.M.; Cheng, C.; Hamed, M.M.; Hamed, A.N.E.S.; Ohlsen, K.; Hentschel, U.; Abdelmohsen, U.R. Antifungal potential of marine natural products. Eur. J. Med. Chem. 2017, 126, 631–651. [Google Scholar] [CrossRef] [PubMed]
- Shin, H.J. Natural products from marine fungi. Mar. Drugs 2020, 18, 230. [Google Scholar] [CrossRef] [PubMed]
- Kumar, V.; Sarma, V.V.; Thambugala, K.M.; Huang, J.J.; Li, X.Y.; Hao, G.F. Ecology and evolution of marine fungi with their adaptation to climate change. Front. Microbiol. 2021, 12, 719000. [Google Scholar] [CrossRef] [PubMed]
- Wu, B.; Oesker, V.; Wiese, J.; Schmaljohann, R.; Imhoff, J.F. Two new antibiotic pyridones produced by a marine fungus, Trichoderma sp. strain MF106. Mar. Drugs 2014, 12, 1208–1219. [Google Scholar] [CrossRef]
- Imhoff, J.F. Natural products from marine fungi—Still an underrepresented resource. Mar. Drugs 2016, 14, 19. [Google Scholar] [CrossRef] [PubMed]
- Borthwick, A.D. 2, 5-Diketopiperazines: Synthesis, reactions, medicinal chemistry, and bioactive natural products. Chem. Rev. 2012, 112, 3641–3716. [Google Scholar] [CrossRef]
- Hu, J.; Li, Z.; Gao, J.; He, H.; Dai, H.; Xia, X.; Liu, C.; Zhang, L.; Song, F. New diketopiperazines from a marine-derived fungus strain Aspergillus versicolor MF180151. Mar. Drugs 2019, 17, 262. [Google Scholar] [CrossRef]
- Xu, L.; Meng, W.; Cao, C.; Wang, J.; Shan, W.; Wang, Q. Antibacterial and antifungal compounds from marine fungi. Mar. Drugs 2015, 13, 3479–3513. [Google Scholar] [CrossRef]
- Haga, A.; Tamoto, H.; Ishino, M.; Kimura, E.; Sugita, T.; Kinoshita, K.; Koyama, K. Pyridone alkaloids from a marine-derived fungus, Stagonosporopsis cucurbitacearum, and their activities against azole-resistant Candida albicans. J. Nat. Prod. 2013, 76, 750–754. [Google Scholar] [CrossRef]
- Sun, C.; Zhang, Z.; Ren, Z.; Yu, L.; Zhou, H.; Han, Y.; Shah, M.; Che, Q.; Zhang, G.; Li, D.; et al. Antibacterial cyclic tripeptides from Antarctica-sponge-derived fungus Aspergillus insulicola HDN151418. Mar. Drugs 2020, 18, 532. [Google Scholar] [CrossRef]
- Liu, J.; Gu, B.; Yang, L.; Yang, F.; Lin, H. New anti-inflammatory cyclopeptides from a sponge-derived fungus Aspergillus violaceofuscus. Front. Chem. 2018, 6, 226. [Google Scholar] [CrossRef]
- Jin, L.; Quan, C.; Hou, X.; Fan, S. Potential pharmacological resources: Natural bioactive compounds from marine-derived fungi. Mar. Drugs 2016, 14, 76. [Google Scholar] [CrossRef]
- Liu, Y.; Mándi, A.; Li, X.M.; Meng, L.H.; Kurtán, T.; Wang, B.G. Peniciadametizine A, a dithiodiketopiperazine with a unique spiro [furan-2,7′-pyrazino [1,2-b][1,2] oxazine] skeleton, and a related analogue, Peniciadametizine B, from the marine sponge-derived fungus Penicillium adametzioides. Mar. Drugs 2015, 13, 3640–3652. [Google Scholar] [CrossRef]
- Meng, L.H.; Li, X.M.; Liu, Y.; Wang, B.G. Penicibilaenes A and B, sesquiterpenes with a tricyclo [6.3. 1.01, 5] dodecane skeleton from the marine isolate of Penicillium bilaiae MA-267. Org. Lett. 2014, 16, 6052–6055. [Google Scholar] [CrossRef]
- Meng, L.H.; Zhang, P.; Li, X.M.; Wang, B.G. Penicibrocazines A–E, five new sulfide diketopiperazines from the marine-derived endophytic fungus Penicillium brocae. Mar. Drugs 2015, 13, 276–287. [Google Scholar] [CrossRef]
- Mehner, T.; Krienitz, L. Encyclopedia of Inland Waters; Likens, G.E., Ed.; Academic Press: Cambridge, MA, USA, 2009; pp. 103–113. [Google Scholar]
- Zerrifi, S.E.A.; El Khalloufi, F.; Oudra, B.; Vasconcelos, V. Seaweed bioactive compounds against pathogens and microalgae: Potential uses on pharmacology and harmful algae bloom control. Mar. Drugs 2018, 16, 55. [Google Scholar] [CrossRef]
- Alarif, W.M.; Al-Lihaibi, S.S.; Ayyad, S.E.N.; Abdel-Rhman, M.H.; Badria, F.A. Laurene-type sesquiterpenes from the Red Sea red alga Laurencia obtusa as potential antitumor–antimicrobial agents. Eur. J. Med. Chem. 2012, 55, 462–466. [Google Scholar] [CrossRef]
- Raeesossadati, M.J.; Ahmadzadeh, H.; McHenry, M.P.; Moheimani, N.R. CO2 bioremediation by microalgae in photobioreactors: Impacts of biomass and CO2 concentrations, light, and temperature. Algal Res. 2014, 6, 78–85. [Google Scholar] [CrossRef]
- De Almeida, C.L.F.; Falcão, H.D.S.; Lima, G.R.D.M.; Montenegro, C.D.A.; Lira, N.S.; de Athayde-Filho, P.F.; Rodrigues, L.C.; Souza, M.D.F.V.D.; Barbosa-Filho, J.M.; Batista, L.M. Bioactivities from marine algae of the genus Gracilaria. Int. J. Mol. Sci. 2011, 12, 4550–4573. [Google Scholar] [CrossRef]
- Lee, J.C.; Hou, M.F.; Huang, H.W.; Chang, F.R.; Yeh, C.C.; Tang, J.Y.; Chang, H.W. Marine algal natural products with anti-oxidative, anti-inflammatory, and anti-cancer properties. Cancer Cell Int. 2013, 13, 55–57. [Google Scholar] [CrossRef]
- Wang, X.H.; Zou, Z.R.; Yi, Y.H.; Han, H.; Li, L.; Pan, M.X. Variegatusides: New non-sulphated triterpene glycosides from the sea cucumber Stichopus variegates semper. Mar. Drugs 2014, 12, 2004–2018. [Google Scholar] [CrossRef]
- Bahrami, Y.; Franco, C.M. Acetylated triterpene glycosides and their biological activity from holothuroidea reported in the past six decades. Mar. Drugs 2016, 14, 147. [Google Scholar] [CrossRef]
- Hua, H.A.N.; Ling, L.I.; Yi, Y.H.; Wang, X.H.; Pan, M.X. Triterpene glycosides from sea cucumber Holothuria scabra with cytotoxic activity. Chin. Herb. Med. 2012, 4, 183–188. [Google Scholar]
- Wang, Z.N.; Yuan, X. Concurrent effects of hot streak and gas species concentration on aerothermal characteristics in a turbine stage. In Turbo Expo: Power for Land, Sea, and Air; American Society of Mechanical Engineers: New York, NY, USA, 2012; Volume 44748, pp. 1431–1441. [Google Scholar]
- Elbandy, M.; Rho, J.R.; Afifi, R. Analysis of saponins as bioactive zoochemicals from the marine functional food sea cucumber Bohadschia cousteaui. Eur. Food Res. Technol. 2014, 238, 937–955. [Google Scholar] [CrossRef]
- Bordbar, S.; Anwar, F.; Saari, N. High-value components and bioactives from sea cucumbers for functional foods—A review. Mar. Drugs 2011, 9, 1761–1805. [Google Scholar] [CrossRef]
- Jamison, M.T.; Dalisay, D.S.; Molinski, T.F. Peroxide natural products from plakortis zyggompha and the sponge association plakortis halichondrioides–xestospongia deweerdtae: Antifungal activity against Cryptococcus gattii. J. Nat. Prod. 2016, 79, 555–563. [Google Scholar] [CrossRef]
- Stout, E.P.; Yu, L.C.; Molinski, T.F. Antifungal diterpene alkaloids from the Caribbean sponge Agelas citrina: Unified configurational assignments of agelasidines and agelasines. Eur. J. Org. Chem. 2012, 2012, 5131–5135. [Google Scholar] [CrossRef]
- Zhou, X.; Hartman, S.V.; Born, E.J.; Smits, J.P.; Holstein, S.A.; Wiemer, D.F. Triazole-based inhibitors of geranylgeranyltransferase II. Bioorganic Med. Chem. Lett. 2013, 23, 764–766. [Google Scholar] [CrossRef]
- Gotsbacher, M.P.; Karuso, P. New antimicrobial bromotyrosine analogues from the sponge Pseudoceratina purpurea and its predator Tylodina corticalis. Mar. Drugs 2015, 13, 1389–1409. [Google Scholar] [CrossRef]
- Olatunji, O.J. Bromotyrosines from the sponges Acanthodendrilla sp. and Pseudoceratina cf. Ph.D. Thesis, Prince of Songkla University, Hat Yai, Thailand, 2014. [Google Scholar]
- Fuwa, H. Contemporary strategies for the synthesis of tetrahydropyran derivatives: Application to total synthesis of neopeltolide, a marine macrolide natural product. Mar. Drugs 2016, 14, 65. [Google Scholar] [CrossRef] [PubMed]
- Harvey, A.L.; Edrada-Ebel, R.; Quinn, R.J. The re-emergence of natural products for drug discovery in the genomics era. Nat. Rev. Drug Discov. 2015, 14, 111–129. [Google Scholar] [CrossRef] [PubMed]
- Youssef, D.T.; Shaala, L.A.; Mohamed, G.A.; Badr, J.M.; Bamanie, F.H.; Ibrahim, S.R. Theonellamide G, a potent antifungal and cytotoxic bicyclic glycopeptide from the Red Sea marine sponge Theonella swinhoei. Mar. Drugs 2014, 12, 1911–1923. [Google Scholar] [CrossRef] [PubMed]
- Kumar, R.; Subramani, R.; Feussner, K.D.; Aalbersberg, W. Aurantoside K, a new antifungal tetramic acid glycoside from a Fijian marine sponge of the genus Melophlus. Mar. Drugs 2012, 10, 200–208. [Google Scholar] [CrossRef] [PubMed]
- Kikuchi, M.; Nosaka, K.; Akaji, K.; Konno, H. Solid phase total synthesis of callipeltin isolated from marine sponge Latrunculia sp. Tetrahedron Lett. 2011, 52, 3872–3875. [Google Scholar] [CrossRef]
- Stierhof, M.; Hansen, K.Ø.; Sharma, M.; Feussner, K.; Subko, K.; Díaz-Rullo, F.F.; Isaksson, J.; Pérez-Victoria, I.; Clarke, D.; Hansen, E.; et al. New cytotoxic callipeltins from the Solomon Island marine sponge Asteropus sp. Tetrahedron 2016, 72, 6929–6934. [Google Scholar] [CrossRef]
- El-Amraoui, B.; Biard, J.F.; Fassouane, A. Haliscosamine: A new antifungal sphingosine derivative from the Moroccan marine sponge Haliclona viscosa. Springerplus 2013, 2, 252. [Google Scholar] [CrossRef]
- Xu, T.; Feng, Q.; Jacob, M.R.; Avula, B.; Mask, M.M.; Baerson, S.R.; Tripathi, S.K.; Mohammed, R.; Hamann, M.T.; Khan, I.A.; et al. The marine sponge-derived polyketide endoperoxide plakortide F acid mediates its antifungal activity by interfering with calcium homeostasis. Antimicrob. Agents Chemother. 2011, 55, 1611–1621. [Google Scholar] [CrossRef]
- Liu, X.F.; Shen, Y.; Yang, F.; Hamann, M.T.; Jiao, W.H.; Zhang, H.J.; Chen, W.-S.; Lin, H.W. Simplexolides A–E and plakorfuran A, six butyrate derived polyketides from the marine sponge Plakortis simplex. Tetrahedron 2012, 68, 4635–4640. [Google Scholar] [CrossRef]
- Tripathi, S.K.; Feng, Q.; Liu, L.; Levin, D.E.; Roy, K.K.; Doerksen, R.J.; Baerson, S.R.; Shi, X.; Pan, X.; Xu, W.-H.; et al. Puupehenone, a marine-sponge-derived sesquiterpene quinone, potentiates the antifungal drug caspofungin by disrupting Hsp90 activity and the cell wall integrity pathway. Msphere 2020, 5, e00818-19. [Google Scholar] [CrossRef]
- Piao, S.J.; Song, Y.L.; Jiao, W.H.; Yang, F.; Liu, X.F.; Chen, W.S.; Han, B.-N.; Lin, H.W. Hippolachnin A, a new antifungal polyketide from the South China sea sponge Hippospongia lachne. Org. Lett. 2013, 15, 3526–3529. [Google Scholar] [CrossRef]
- Arevabini, C.; Crivelenti, Y.D.; de Abreu, M.H.; Bitencourt, T.A.; Santos, M.F.; Berlinck, R.G.; Marins, M. Antifungal activity of metabolites from the marine sponges Amphimedon sp. and Monanchora arbuscula against Aspergillus flavus strains isolated from peanuts (Arachis hypogaea). Nat. Prod. Commun. 2014, 9, 33–36. [Google Scholar] [CrossRef]
- Mosey, R.A.; Floreancig, P.E. Isolation, biological activity, synthesis, and medicinal chemistry of the pederin/mycalamide family of natural products. Nat. Prod. Rep. 2012, 29, 980–995. [Google Scholar] [CrossRef]
- Abdelmohsen, U.R.; Balasubramanian, S.; Oelschlaeger, T.A.; Grkovic, T.; Pham, N.B.; Quinn, R.J.; Hentschel, U. Potential of marine natural products against drug-resistant fungal, viral, and parasitic infections. Lancet Infect. Dis. 2017, 17, e30–e41. [Google Scholar] [CrossRef]
- Martins, A.; Vieira, H.; Gaspar, H.; Santos, S. Marketed marine natural products in the pharmaceutical and cosmeceutical industries: Tips for success. Mar. Drugs 2014, 12, 1066–1101. [Google Scholar] [CrossRef]
- Aslam, B.; Wang, W.; Arshad, M.I.; Khurshid, M.; Muzammil, S.; Nisar, M.A.; Alvi, R.F.; Aslam, M.A.; Qamar, M.U.; Salamat, M.K.F.; et al. Antibiotic resistance: A rundown of a global crisis. Infect. Drug Resist. 2018, 11, 1645. [Google Scholar] [CrossRef]
- Travkova, O.G.; Moehwald, H.; Brezesinski, G. The interaction of antimicrobial peptides with membranes. Adv. Colloid Interface Sci. 2017, 247, 521–532. [Google Scholar] [CrossRef]
- Cho, J.; Lee, D.G. The antimicrobial peptide arenicin-1 promotes generation of reactive oxygen species and induction of apoptosis. Biochim. Et Biophys. Acta (BBA)-Gen. Subj. 2011, 1810, 1246–1251. [Google Scholar] [CrossRef]
- Choi, H.; Lee, D.G. Synergistic effect of antimicrobial peptide arenicin-1 in combination with antibiotics against pathogenic bacteria. Res. Microbiol. 2012, 163, 479–486. [Google Scholar] [CrossRef]
- Panteleev, P.V.; Bolosov, I.A.; Balandin, S.V.; Ovchinnikova, T.V. Design of antimicrobial peptide arenicin analogs with improved therapeutic indices. J. Pept. Sci. 2015, 21, 105–113. [Google Scholar] [CrossRef] [PubMed]
- Han, J.; Jyoti, M.A.; Song, H.Y.; Jang, W.S. Antifungal activity and action mechanism of histatin 5-halocidin hybrid peptides against Candida ssp. PLoS ONE 2016, 11, e0150196. [Google Scholar] [CrossRef] [PubMed]
- Shin, S.H.; Lee, Y.S.; Shin, Y.P.; Kim, B.; Kim, M.H.; Chang, H.R.; Jang, W.S.; Lee, I.H. Therapeutic efficacy of halocidin-derived peptide HG1 in a mouse model of Candida albicans oral infection. J. Antimicrob. Chemother. 2013, 68, 1152–1160. [Google Scholar] [CrossRef] [PubMed]
- Jeong, J.E.; Kang, S.W.; Shin, Y.K.; Jun, J.C.; Kim, Y.O.; Hur, Y.B.; Kim, J.-H.; Chae, S.-H.; Lee, J.-S.; Choi, I.H.; et al. Comparative analysis of expressed sequence tags (ESTs) between normal group and softness syndrome group in Halocynthia roretzi. Mol. Cell. Toxicol. 2011, 7, 357–365. [Google Scholar] [CrossRef]
- Cuvillier-Hot, V.; Boidin-Wichlacz, C.; Tasiemski, A. Polychaetes as annelid models to study ecoimmunology of marine organisms. J. Mar. Sci. Technol. 2014, 22, 9–14. [Google Scholar] [CrossRef]
- Rajanbabu, V.; Chen, J.Y.; Wu, J.L. Antimicrobial peptides from marine organisms. In Springer Handbook of Marine Biotechnology; Springer: Berlin/Heidelberg, Germany, 2015; pp. 747–758. [Google Scholar]
- de Miranda, J.L.; Oliveira, M.D.; Oliveira, I.S.; Frias, I.A.; Franco, O.L.; Andrade, C.A. A simple nanostructured biosensor based on clavanin A antimicrobial peptide for gram-negative bacteria detection. Biochem. Eng. J. 2017, 124, 108–114. [Google Scholar] [CrossRef]
- Silva, O.N.; Fensterseifer, I.C.; Rodrigues, E.A.; Holanda, H.H.; Novaes, N.R.; Cunha, J.P.; Rezende, T.M.B.; Magalhães, K.G.; Moreno, S.E.; Jerônimo, M.S.; et al. Clavanin A improves outcome of complications from different bacterial infections. Antimicrob. Agents Chemother. 2015, 59, 1620–1626. [Google Scholar] [CrossRef]
- Andrade, C.A.; Nascimento, J.M.; Oliveira, I.S.; de Oliveira, C.V.; de Melo, C.P.; Franco, O.L.; Oliveira, M.D. Nanostructured sensor based on carbon nanotubes and clavanin A for bacterial detection. Colloids Surf. BBiointerfaces 2015, 135, 833–839. [Google Scholar] [CrossRef]
- Nguyen, L.T.; Haney, E.F.; Vogel, H.J. The expanding scope of antimicrobial peptide structures and their modes of action. Trends Biotechnol. 2011, 29, 464–472. [Google Scholar] [CrossRef] [PubMed]
- Miller, A.; Matera-Witkiewicz, A.; Mikołajczyk, A.; Wieczorek, R.; Rowinska-Zyrek, M. Chemical “butterfly effect” explaining the coordination chemistry and antimicrobial properties of clavanin complexes. Inorg. Chem. 2021, 60, 12730–12734. [Google Scholar] [CrossRef]
- Saúde, A.C.; Ombredane, A.S.; Silva, O.N.; Barbosa, J.A.; Moreno, S.E.; Araujo, A.C.G.; Franco, O.L. Clavanin bacterial sepsis control using a novel methacrylate nanocarrier. Int. J. Nanomed. 2014, 9, 5055–5069. [Google Scholar]
- Hur, G.H.; Vickery, C.R.; Burkart, M.D. Explorations of catalytic domains in non-ribosomal peptide synthetase enzymology. Nat. Prod. Rep. 2012, 29, 1074–1098. [Google Scholar] [CrossRef]
- Liu, Y.; Ding, S.; Shen, J.; Zhu, K. Nonribosomal antibacterial peptides that target multidrug-resistant bacteria. Nat. Prod. Rep. 2019, 36, 573–592. [Google Scholar] [CrossRef]
- Aldholmi, M.; Wilkinson, B.; Ganesan, A. Epigenetic modulation of secondary metabolite profiles in Aspergillus calidoustus and Aspergillus westerdijkiae through histone deacetylase (HDAC) inhibition by vorinostat. J. Antibiot. 2020, 73, 410–413. [Google Scholar] [CrossRef] [PubMed]
- Lukassen, M.B.; Saei, W.; Sondergaard, T.E.; Tamminen, A.; Kumar, A.; Kempken, F.; Wiebe, M.G.; Sørensen, J.L. Identification of the scopularide biosynthetic gene cluster in Scopulariopsis brevicaulis. Mar. Drugs 2015, 13, 4331–4343. [Google Scholar] [CrossRef]
- Pradhan, T.K.; Reddy, K.M.; Ghosh, S. Total synthesis of emericellamides A and B. TetrahedronAsymmetry 2013, 24, 1042–1051. [Google Scholar] [CrossRef]
- Bleich, R.; Watrous, J.D.; Dorrestein, P.C.; Bowers, A.A.; Shank, E.A. Thiopeptide antibiotics stimulate biofilm formation in Bacillus subtilis. Proc. Natl. Acad. Sci. USA 2015, 112, 3086–3091. [Google Scholar] [CrossRef] [PubMed]
- Indraningrat, A.A.G.; Smidt, H.; Sipkema, D. Bioprospecting sponge-associated microbes for antimicrobial compounds. Mar. Drugs 2016, 14, 87. [Google Scholar] [CrossRef] [PubMed]
- Yamashita, Y.; Hirano, Y.; Takada, A.; Takikawa, H.; Suzuki, K. Total Synthesis of Bis-anthraquinone Antibiotic BE-43472B. Synthesis 2018, 50, 2490–2515. [Google Scholar] [CrossRef]
- Wang, Q.; Song, F.; Xiao, X.; Huang, P.; Li, L.; Monte, A.; Zhang, L. Abyssomicins from the South China Sea deep-sea sediment Verrucosispora sp.: Natural thioether Michael addition adducts as antitubercular prodrugs. Angew. Chem. 2013, 125, 1269–1272. [Google Scholar] [CrossRef]
- Augner, D.; Krut, O.; Slavov, N.; Gerbino, D.C.; Sahl, H.G.; Benting, J.; Nising, C.F.; Hillebrand, S.; Krönke, M.; Schmalz, H.-G. On the antibiotic and antifungal activity of pestalone, pestalachloride A, and structurally related compounds. J. Nat. Prod. 2013, 76, 1519–1522. [Google Scholar] [CrossRef]
- Liu, S.; Dai, H.; Makhloufi, G.; Heering, C.; Janiak, C.; Hartmann, R.; Mándi, A.; Kurtán, T.; Müller, W.E.G.; Kassack, M.U.; et al. Cytotoxic 14-membered macrolides from a mangrove-derived endophytic fungus, Pestalotiopsis microspora. J. Nat. Prod. 2016, 79, 2332–2340. [Google Scholar] [CrossRef] [PubMed]
- Linares-Otoya, L.; Linares-Otoya, V.; Armas-Mantilla, L.; Blanco-Olano, C.; Crüsemann, M.; Ganoza-Yupanqui, M.L.; Campos-Florian, J.; König, G.M.; Schäberle, T.F. Diversity and antimicrobial potential of predatory bacteria from the Peruvian coastline. Mar. Drugs 2017, 15, 308. [Google Scholar] [CrossRef]
- Mayer, A.M.; Rodríguez, A.D.; Berlinck, R.G.; Fusetani, N. Marine pharmacology in 2007–8: Marine compounds with antibacterial, anticoagulant, antifungal, anti-inflammatory, antimalarial, antiprotozoal, antituberculosis, and antiviral activities; affecting the immune and nervous system, and other miscellaneous mechanisms of action. Comp. Biochem. Physiol. Part C Toxicol. Pharmacol. 2011, 153, 191–222. [Google Scholar]
- Rateb, M.E.; Ebel, R. Secondary metabolites of fungi from marine habitats. Nat. Prod. Rep. 2011, 28, 290–344. [Google Scholar] [CrossRef]
- Wang, T.; Yang, S.; Li, H.; Lu, A.; Wang, Z.; Yao, Y.; Wang, Q. Discovery, structural optimization, and mode of action of essramycin alkaloid and its derivatives as anti-tobacco mosaic virus and anti-phytopathogenic fungus agents. J. Agric. Food Chem. 2019, 68, 471–484. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.; Hesek, D.; Lee, M.; Lastochkin, E.; Oliver, A.G.; Chang, M.; Mobashery, S. The natural product essramycin and three of its isomers are devoid of antibacterial activity. J. Nat. Prod. 2016, 79, 1219–1222. [Google Scholar] [CrossRef] [PubMed]
- Mohapatra, D.K.; Reddy, D.P.; Dash, U.; Yadav, J.S. Total synthesis of Z-isomer of phomolide B. Tetrahedron Lett. 2011, 52, 151–154. [Google Scholar] [CrossRef]
- Arunpanichlert, J.; Rukachaisirikul, V.; Sukpondma, Y.; Phongpaichit, S.; Supaphon, O.; Sakayaroj, J. A β-resorcylic macrolide from the seagrass-derived fungus Fusarium sp. PSU-ES73. Arch. Pharmacal Res. 2011, 34, 1633–1637. [Google Scholar] [CrossRef]
- Thawabteh, A.; Juma, S.; Bader, M.; Karaman, D.; Scrano, L.; Bufo, S.A.; Karaman, R. The biological activity of natural alkaloids against herbivores, cancerous cells and pathogens. Toxins 2019, 11, 656. [Google Scholar] [CrossRef]
- Das, J.; Bhandari, M.; Lovely, C.J. Isolation, Bioactivity, and Synthesis of Nagelamides. Stud. Nat. Prod. Chem. 2016, 50, 341–371. [Google Scholar]
- Swain, S.S.; Paidesetty, S.K.; Padhy, R.N. Antibacterial, antifungal and antimycobacterial compounds from cyanobacteria. Biomed. Pharmacother. 2017, 90, 760–776. [Google Scholar] [CrossRef]
- Bian, M.; Li, L.; Ding, H. Recent Advances on the Application of Electrocyclic Reactions in Complex Natural Product Synthesis. Synthesis 2017, 49, 4383–4413. [Google Scholar] [CrossRef]
- Pessoa, C.; dos Santos, M.F.C.; Berlinck, R.G.S.; Ferreira, P.M.P.; Cavalcanti, B.C. Cytotoxic batzelladine L from the Brazilian marine sponge Monanchora arbuscula. Planta Med. 2013, 79, PK6. [Google Scholar] [CrossRef]
- Saurav, K.; Zhang, W.; Saha, S.; Zhang, H.; Li, S.; Zhang, Q.; Wu, Z.; Zhang, G.; Zhu, Y.; Verma, G. In silico molecular docking, preclinical evaluation of spiroindimicins AD, lynamicin A and D isolated from deep marine sea derived Streptomyces sp. SCSIO 03032. Interdiscip. Sci. Comput. Life Sci. 2014, 6, 187–196. [Google Scholar] [CrossRef] [PubMed]
- Clive, D.L.; Cheng, P. The marinopyrroles. Tetrahedron 2013, 69, 5067–5078. [Google Scholar] [CrossRef]
- Thomson, R.H. (Ed.) The Chemistry of Natural Products; Springer Science & Business Media: Berlin/Heidelberg, Germany, 2012. [Google Scholar]
- Thawabteh, A.M.; Thawabteh, A.; Lelario, F.; Bufo, S.A.; Scrano, L. Classification, toxicity and bioactivity of natural diterpenoid alkaloids. Molecules 2021, 26, 4103. [Google Scholar] [CrossRef] [PubMed]
- Rocha, J.; Peixe, L.; Gomes, N.C.; Calado, R. Cnidarians as a source of new marine bioactive compounds—An overview of the last decade and future steps for bioprospecting. Mar. Drugs 2011, 9, 1860–1886. [Google Scholar] [CrossRef] [PubMed]
- APA American Psychological Association. National Center for Biotechnology Information. Pubchem Compound Summary for CID 12699, N-Nitroso-N-methylurea. Retrieved 24. 2020. Available online: https://pubchem.ncbi.nlm.nih.gov/compound/N-Nitroso-N-methylurea (accessed on 6 January 2023).
- Incerti-Pradillos, C.A.; Kabeshov, M.A.; O’Hora, P.S.; Shipilovskikh, S.A.; Rubtsov, A.E.; Drobkova, V.A.; Balandina, S.Y.; Malkov, A.V. Asymmetric Total Synthesis of (−)-Erogorgiaene and Its C-11 Epimer and Investigation of Their Antimycobacterial Activity. Chem. A Eur. J. 2016, 22, 14390–14396. [Google Scholar] [CrossRef]
- Pour, P.M.; Behzad, S.; Asgari, S.; Khankandi, H.P.; Farzaei, M.H. Sesterterpenoids. In Recent Advances in Natural Products Analysis; Elsevier: Amsterdam, The Netherlands, 2020; pp. 347–391. [Google Scholar]
- Chen, Y.; Zhao, J.; Li, S.; Xu, J. Total synthesis of sesterterpenoids. Nat. Prod. Rep. 2019, 36, 263–288. [Google Scholar] [CrossRef] [PubMed]
- McCulloch, M.W.; Haltli, B.; Marchbank, D.H.; Kerr, R.G. Evaluation of pseudopteroxazole and pseudopterosin derivatives against Mycobacterium tuberculosis and other pathogens. Mar. Drugs 2012, 10, 1711–1728. [Google Scholar] [CrossRef] [PubMed]
- Ibañez, E.; Herrero, M.; Mendiola, J.A.; Castro-Puyana, M. Extraction and characterization of bioactive compounds with health benefits from marine resources: Macro and micro algae, cyanobacteria, and invertebrates. In Marine Bioactive Compounds; Springer: Berlin/Heidelberg, Germany, 2012; pp. 55–98. [Google Scholar]
- Dumas, F.; Kousara, M.; Chen, L.; Wei, L.; Le Bideau, F. Nonhalogenated heterotricyclic sesquiterpenes from marine origin i: Fused systems. Stud. Nat. Prod. Chem. 2017, 52, 269–302. [Google Scholar]
- Yende, S.R.; Harle, U.N.; Chaugule, B.B. Therapeutic potential and health benefits of Sargassum species. Pharmacogn. Rev. 2014, 8, 1–7. [Google Scholar] [CrossRef] [PubMed]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
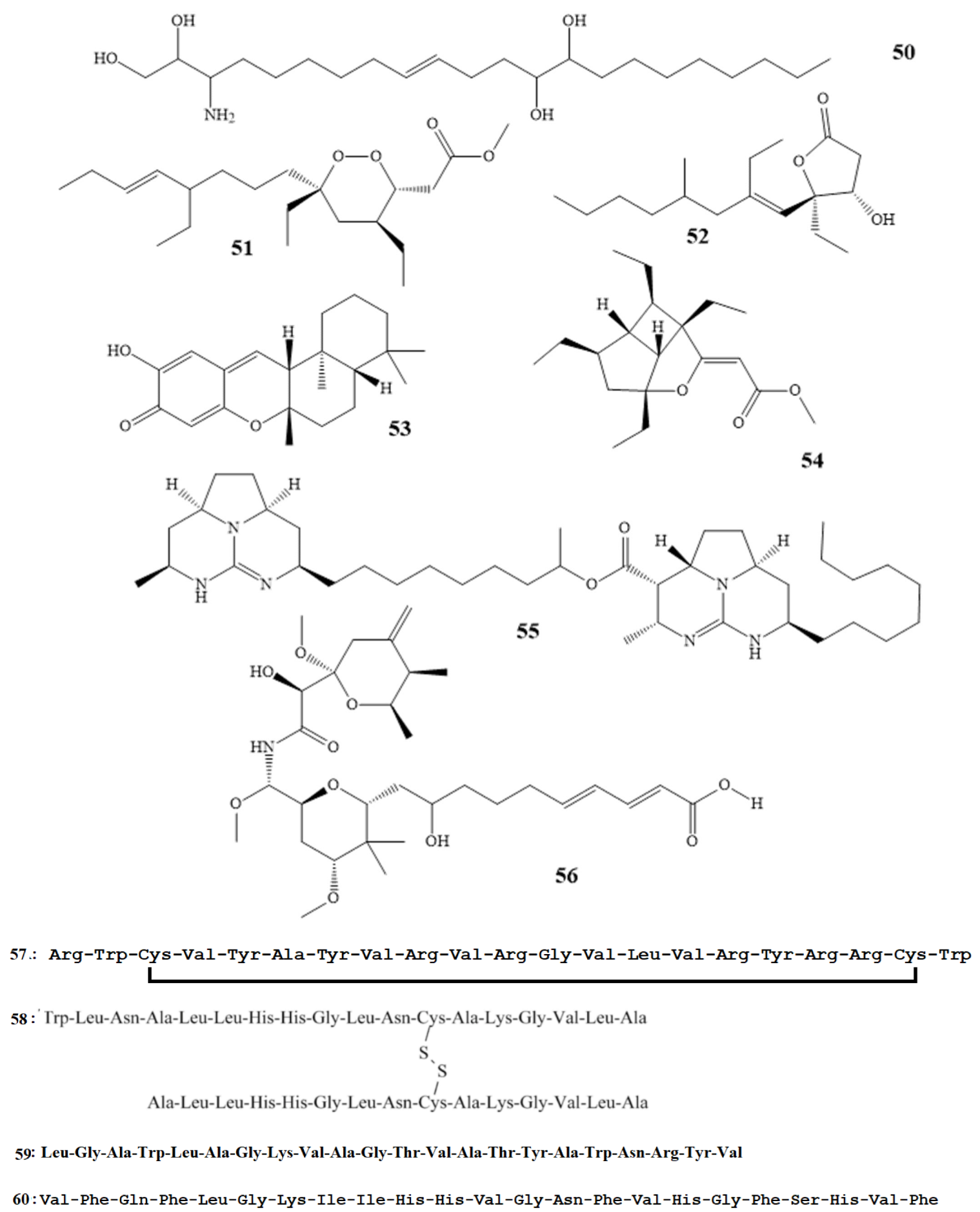
| Isolated Compound | Marine Sources | Activities | MIC µg/mL |
|---|---|---|---|
| Basiliskamides A (7) Basiliskamides B (8) [22] | Bacillus laterosporus | C. albicans A. fumigatus | 1.0–5.0 |
| Mohangamide A (9) [23] | Streptomyces sp. | C. albicans | 4.14 |
| Haliangicin (10) [24,25] | Haliangium luteum | Botrytis cinerea Pythium ultimum Saprolegnia parasitica | 3.1 0.4 0.1 |
| Gageostatin A (11) [26,27] | B. subtilis 109GGC020 | Rhizoctonia solani B. cinerea | 4 |
| Hassallidin A (12) [28] | epilithic cyanobacteria | A. fumigatus C. albicans | 4.8 |
| Macrolactins T (13) Macrolactins B (14) [29] | Bufo marinus | Pyricularia oryzae Alternaria. solani | 0.8–4.8 |
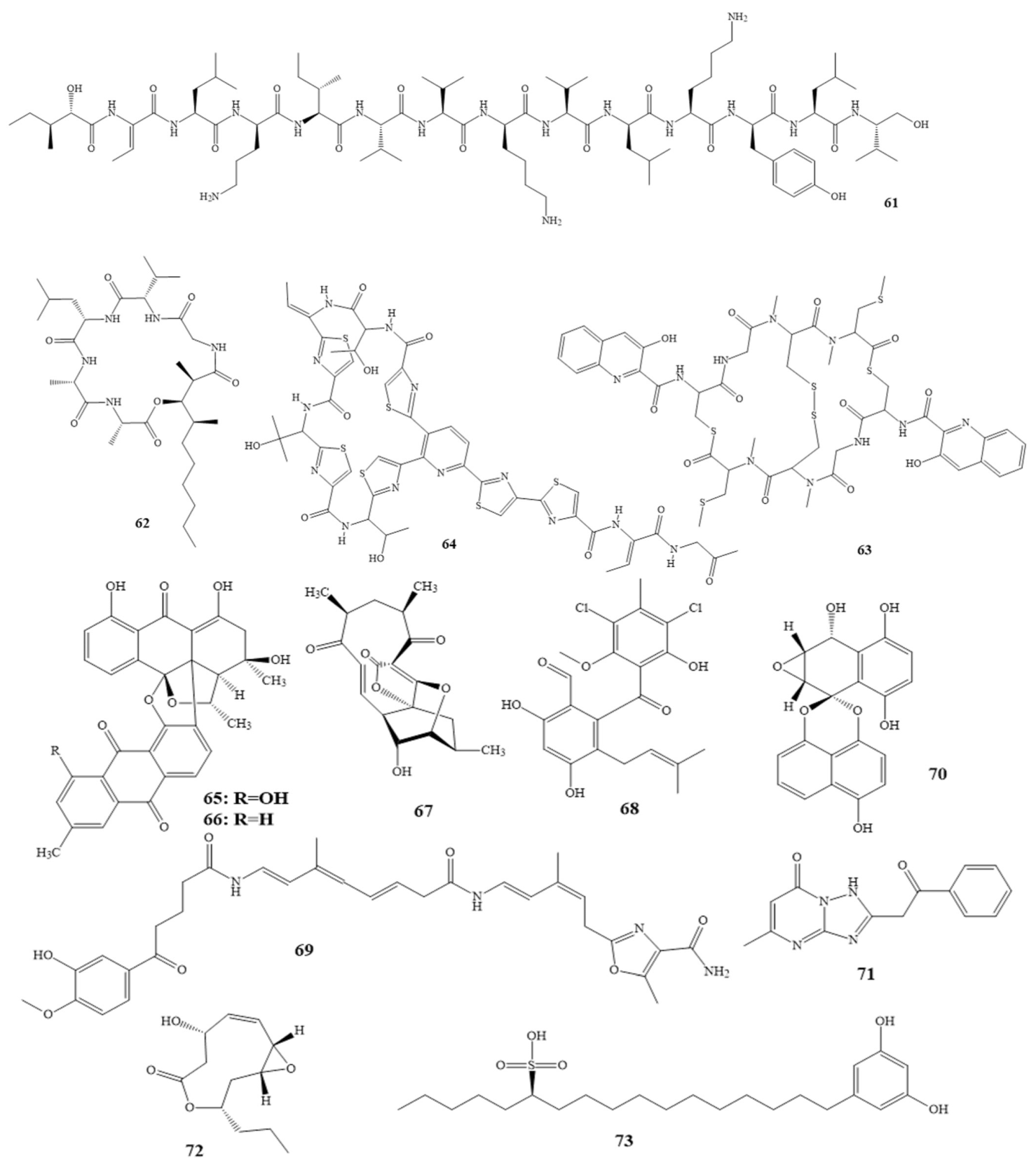
| Isolated Compound | Marine Sources | Ref. | Conc. of Inhibition |
|---|---|---|---|
| Ceratinadins A (39) Ceratinadins B (40) | Pseudoceratina sp. | [54,55] | MIC 2 µg/mL 4 µg/mL |
| Neopeltolide (41) | Okinawan sponge | [56,57] | MIC 0.62 µg/mL |
| Theonellamide G (43) | Theonella swinhoei in the red sea | [58] | IC50 4.49 µM |
| Aurantoside K (42) * | Melophlus sp. | [59] | MIC 31.25 μg/mL |
| Callipeltins peptides F (44) Callipeltins peptides G (45) Callipeltins peptides H (46) Callipeltins peptides I (47) Callipeltins peptides J (48) Callipeltins peptides K (49) | Latrunculia sp. sponge usually found in Vanuatu islands and South Pacific | [60,61] | MIC 100 µM |
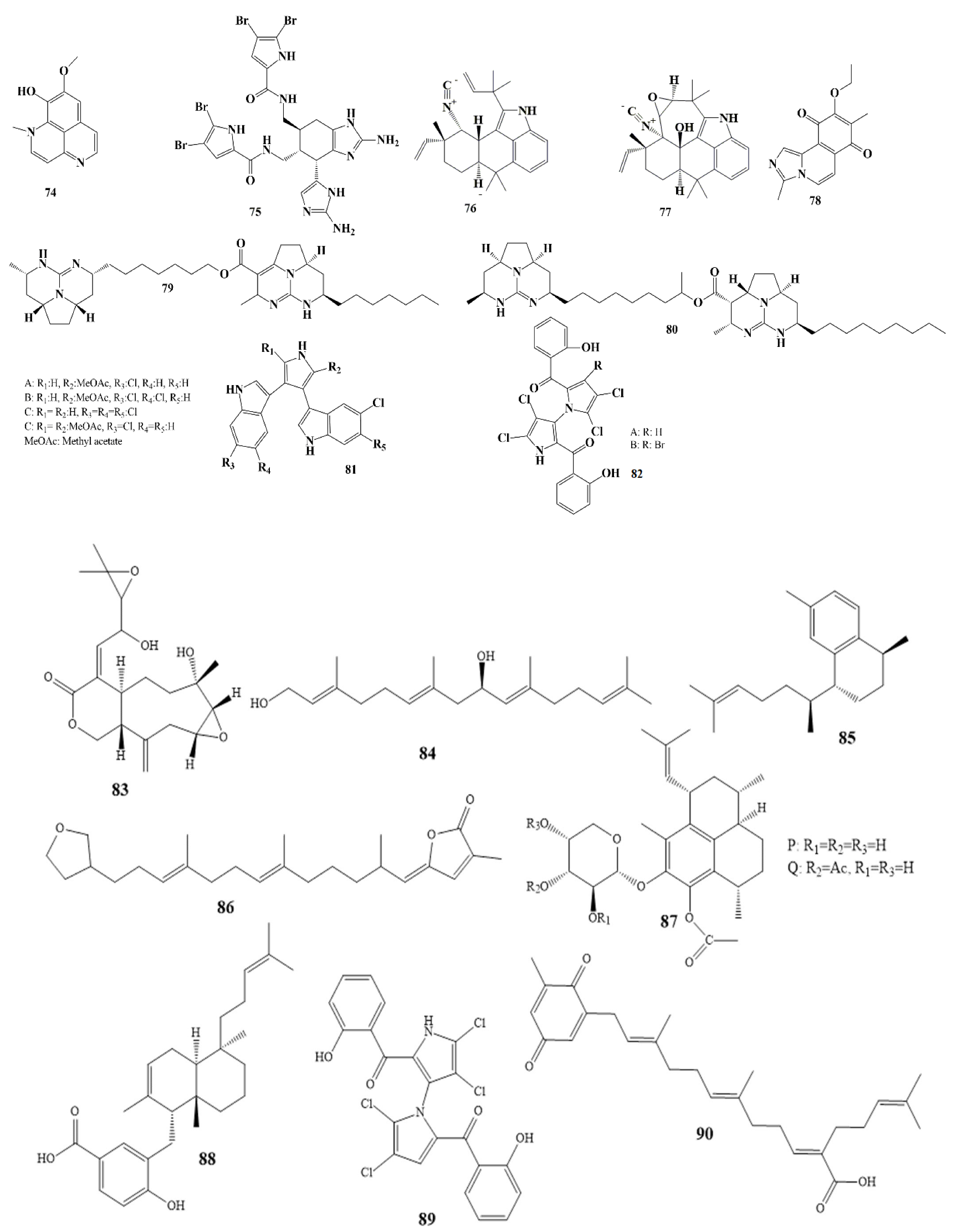
| Isolated Compound | Marine Sources | Activities | MIC µg/mL |
|---|---|---|---|
| Ariakemicins A (69) [98,99] | Rapidithrix sp. (marine gliding bacterium) | Brevibacterium sp. S. aureus B. subtilis | 830.4683 |
| Ascochytatin (70) [100] | marine-derived fungus, Ascochyta sp. | B. subtilis | 4.8 |
| Essramycin (71) [101,102] | culture broth of the marine Streptomyces sp. | E. coli P. aeruginosa B. subtilis S. aureus M. luteus | 1–8 |
| Phomolides B (72) [103] | culture of Phomopsis sp. | E. coli strains B. subtilis | 5–101.25 |
| Sulfoalkylresorcinol (73) [104] | marine-derived fungus Zygosporium sp. | S. aureus | 12.5 |
| Isolated Compound | Marine Sources | Terpenes Class | Activities | MIC µg/mL |
|---|---|---|---|---|
| Erogorgiaene (85) [116] | Pseudopterogorgia elisabethae | Serrulatane Diterpenes | M. tuberculosis | 12.5 |
| 22-deoxyvariabilin (86), [117,118] | Sponge Ircinia variabilis | Sesterterpene | S. aureus B. subtilis | 50 100 |
| Pseudopterosin P and Q, (87), [119] | Pseudopterogorgia elisabethae | diterpene glycosides | S. pyogenes S. aureus E. faecalis | 0.8 and 1 2 and 2.3 3.5 and 3.6 |
| Isojaspic acid (88), [120] | sponge Cacospongia | meroditerpene | S. epidermis | 2.5 |
| (–)-Microcionin, (89) [121] | Fasciospongia sp. | furanosesquiterpenes | M. luteus | 6 |
| Sargaquinoic acid, (90), [122] | Sargassum sagamianum | Plastoquinones | S. aureus | 2 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Thawabteh, A.M.; Swaileh, Z.; Ammar, M.; Jaghama, W.; Yousef, M.; Karaman, R.; A. Bufo, S.; Scrano, L. Antifungal and Antibacterial Activities of Isolated Marine Compounds. Toxins 2023, 15, 93. https://doi.org/10.3390/toxins15020093
Thawabteh AM, Swaileh Z, Ammar M, Jaghama W, Yousef M, Karaman R, A. Bufo S, Scrano L. Antifungal and Antibacterial Activities of Isolated Marine Compounds. Toxins. 2023; 15(2):93. https://doi.org/10.3390/toxins15020093
Chicago/Turabian StyleThawabteh, Amin Mahmood, Zain Swaileh, Marwa Ammar, Weam Jaghama, Mai Yousef, Rafik Karaman, Sabino A. Bufo, and Laura Scrano. 2023. "Antifungal and Antibacterial Activities of Isolated Marine Compounds" Toxins 15, no. 2: 93. https://doi.org/10.3390/toxins15020093
APA StyleThawabteh, A. M., Swaileh, Z., Ammar, M., Jaghama, W., Yousef, M., Karaman, R., A. Bufo, S., & Scrano, L. (2023). Antifungal and Antibacterial Activities of Isolated Marine Compounds. Toxins, 15(2), 93. https://doi.org/10.3390/toxins15020093