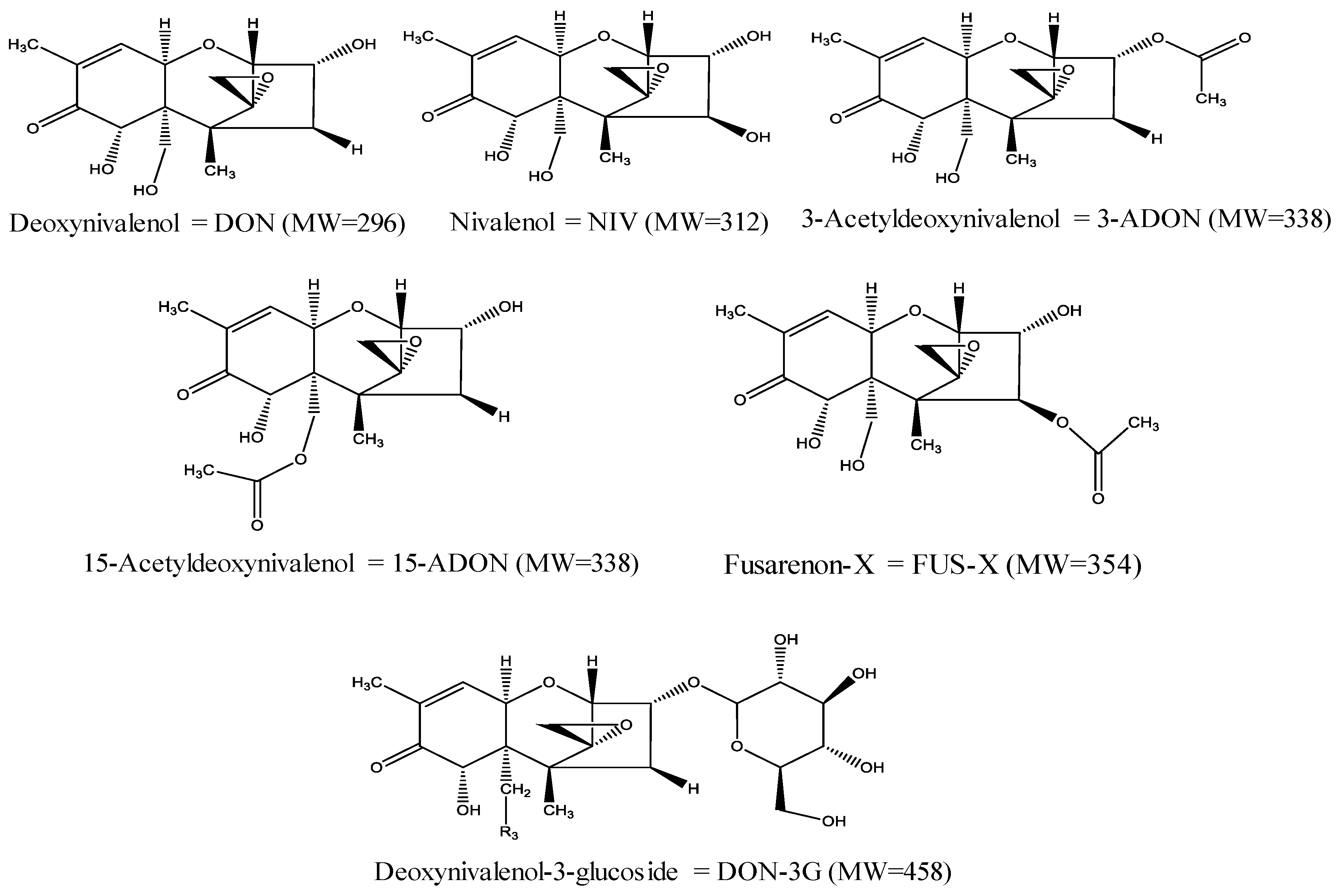
Type B Trichothecenes in Cereal Grains and Their Products: Recent Advances on Occurrence, Toxicology, Analysis and Post-Harvest Decontamination Strategies



Abstract
1. Introduction
2. Factors Affecting Type B Trichothecenes Production
3. Toxicity of Type B Trichothecenes
4. Legislation of Type B Trichothecenes in Foodstuffs

{kind=link}
{kind=link}
{kind=link}
| Regulation Authority | Foodstuff | Maximum Level μg/kg | Reference |
|---|---|---|---|
| European Commission | Unprocessed durum wheat, maize, and oats | 1750 | [86] |
| Unprocessed cereals | 1250 | ||
| Cereal flour for human consumption and dry pasta | 750 | ||
| Bread, pastries, breakfast cereals, and cereal snacks | 500 | ||
| Cereal-based foods and baby foods for infants and young children | 200 | ||
| United States Food and Drug Administration | Flour, germ, and bran derived from wheat cereal intended for human consumption | 1000 | [87] |
| Food and Agricultural Organization of the United States/Codex Alimentarius | Raw grains such as maize, wheat, and barley | 2000 | [87] |
| Maize, wheat, and barley-derived foods (flour, meal, semolina, and flakes) | 1000 | ||
| Cereal-based foods and baby foods for infants and young children | 500 | ||
| Republic of Korea | Grain and their processed foods | 1000 | [92] |
| Corn and their processed foods | 2000 | ||
| Cereals | 500 | ||
| China | Corn, corn flour (grits, flake) | 1000 | [93] |
| Barley, wheat, cereal, wheat flour | 1000 |
5. Occurrence of Type B Trichothecenes and DON-3G in Cereal Grains and Their Products
5.1. Wheat
5.2. Corn
5.3. Oats
5.4. Barley
5.5. Rice
5.6. Sorghum and Rye
6. Sample Preparation Methods for Analysis
6.1. Sampling Tactics
6.2. Extraction Methods
6.3. Clean-Up Methods
7. Separation and Detection Techniques
7.1. Chromatographic Methods
7.2. Immunochemical Techniques
7.2.1. Enzyme-Linked Immunosorbent Assay
7.2.2. Lateral Flow Immunochromatographic Assay
7.2.3. Fluorescence Polarization Immunoassay
7.3. Biosensors
7.4. Infrared Spectroscopy
7.5. Other Emerging Detection Technologies
7.5.1. Electronic Nose
7.5.2. Capillary Electrophoresis
8. Advances in Management and Control Strategies
8.1. Pre-Harvest Practices
8.2. Harvest and Post-Harvest Practices
9. Conclusions and Future Outlook
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Abbreviations
| 15-ADON | 15-Acetyldeoxynivalenol |
| 3-ADON | 3-Acetyldeoxynivalenol |
| AFs | Aflatoxins |
| ARfD | Acute reference dose |
| ASE | Accelerated solvent extraction |
| CAs | Chromosomal aberrations |
| CE | Capillary electrophoresis |
| CEN | European Committee of Standardization |
| DON-3G | Deoxynivalenol-3-glucoside |
| DNA | Deoxyribonucleic acid |
| DON | Deoxynivalenol |
| DSPE | Dispersive solid-phase extraction |
| dsRNA | Double-stranded RNA |
| EC | European Commission |
| ECD | Electron capture detection |
| EFSA | European Food Safety Authority |
| EN | Electronic nose |
| ELIZA | Enzyme-linked immunosorbent assay |
| EU | European Union |
| FAO | Food and Agriculture Organization |
| FHB | Fusarium head blight |
| FID | Flame ionization detection |
| FPIA | Fluorescence polarization immunoassay |
| FTNIR | Fourier transform NIR |
| FUS-X | Fusarenon-X |
| GAPs | Good agricultural practices |
| GC | Gas chromatography |
| GCB | Graphitized carbon black |
| GC-MS | Gas chromatography–mass spectrometry |
| GC-MS/MS | Gas chromatography–tandem mass spectrometry |
| GHPs | Good hygiene practices |
| GMPs | Good management practices |
| GSPs | Good storage practices |
| Hck | Hematopoietic cell kinase |
| HLB | Hydrophilic–lipophilic balanced |
| HPLC-PDA | High-performance liquid chromatography–photodiode array detection |
| HPLC-UV | High-performance liquid chromatography–ultraviolet detection |
| HRMS | High-resolution mass spectrometry |
| HS-GC/MS | Headspace–gas chromatography/mass spectrometry |
| HT2 | HT-2 toxin |
| IACs | Immunoaffinity columns |
| IARC | International Agency for Research on Cancer |
| IECs | Intestinal epithelial cells |
| JECFA | Joint FAO/WHO Expert Committee on Food Additives |
| LC | Liquid chromatography |
| LC-FLD | Liquid chromatography–fluorescence detection |
| LC-MS | Liquid chromatography–mass spectrometry |
| LC-MS/MS | Liquid chromatography–tandem mass spectrometry |
| LFIA | Lateral flow immunochromatographic assay |
| LLE | Liquid–liquid extraction |
| LOD | Limit of detection |
| LOQ | Limit of quantification |
| mAb | Monoclonal antibody |
| MAE | Microwave-assisted extraction |
| MAPKs | Mitogen-activated protein kinases |
| MOS | Metal oxide semiconductors |
| MFC | Multifunctional clean-up columns |
| MIR | Mid-infrared spectroscopy |
| MOF | Metallic organic framework |
| MRLs | Maximum regulatory limits |
| mRNA | Messenger ribonucleic acid |
| MS | Mass spectrometry |
| MWFPIA | Multi-wavelength fluorescence polarization immunoassay |
| NA | Not available |
| ND | Not detected |
| NIR | Near-infrared spectroscopy |
| NIR-HIS | Near-infrared spectroscopy–hyperspectral imaging |
| NIV | Nivalenol |
| PCA | Principal component analysis |
| PLE | Pressurized liquid extraction |
| PLS-DA | Partial least squares discriminant analysis |
| PMTDI | Provisional maximum tolerable daily intake |
| PRRSV | Porcine reproductive and respiratory syndrome virus |
| PSA | Primary secondary amine |
| PTC | Peptidyl transferase center |
| QuEChERS | Quick, easy, cheap, effective, rugged, and safe |
| RNA | Ribonucleic acid |
| RSD | Relative standard deviation |
| SCF | Scientific Committee on Food |
| SFE | Supercritical fluid extraction |
| SLE | Solid–liquid extraction |
| SPE | Solid-phase extraction |
| T2 | T-2 toxin |
| TDI | Tolerable daily intake |
| TJP | Tight junction proteins |
| TLC | Thin layer chromatography |
| UPLC | Ultra-performance liquid chromatography |
| UPLC-MS/MS | Ultra-performance liquid chromatography–tandem mass spectrometry |
| WHO | World Health Organization |
| ZEN | Zearalenone |
References
- Leite, M.; Freitas, A.; Silva, A.S.; Barbosa, J.; Ramos, F. Maize (Zea mays L.) and mycotoxins: A review on optimization and validation of analytical methods by liquid chromatography coupled to mass spectrometry. Trends Food Sci. Technol. 2020, 99, 542–565. [Google Scholar] [CrossRef]
- Turner, N.W.; Subrahmanyam, S.; Piletsky, S.A. Analytical methods for determination of mycotoxins: A review. Anal. Chim. Acta 2009, 632, 168–180. [Google Scholar] [CrossRef] [PubMed]
- Peng, W.X.; Marchal, J.L.M.; van der Poel, A.F.B. Strategies to prevent and reduce mycotoxins for compound feed manufacturing. Anim. Feed Sci. Technol. 2018, 237, 129–153. [Google Scholar] [CrossRef]
- de Nijs, M.; van den Top, H.; de Stoppelaar, J.; Lopez, P.; Mol, H. Fate of enniatins and deoxynivalenol during pasta cooking. Food Chem. 2016, 213, 763–767. [Google Scholar] [CrossRef] [PubMed]
- Gab-Allah, M.A.; Choi, K.; Kim, B. Development of isotope dilution–liquid chromatography/tandem mass spectrometry as a candidate reference method for the accurate determination of patulin in apple products. Anal. Bioanal. Chem. 2022, 414, 1867–1879. [Google Scholar] [CrossRef]
- Lijalem, Y.G.; Gab-Allah, M.A.; Choi, K.; Kim, B. Development of isotope dilution-liquid chromatography/tandem mass spectrometry for the accurate determination of zearalenone and its metabolites in corn. Food Chem. 2022, 384, 132483. [Google Scholar] [CrossRef]
- Ran, R.; Wang, C.; Han, Z.; Wu, A.; Zhang, D.; Shi, J. Determination of deoxynivalenol (DON) and its derivatives: Current status of analytical methods. Food Control 2013, 34, 138–148. [Google Scholar] [CrossRef]
- Romera, D.; Mateo, E.M.; Mateo-Castro, R.; Gomez, J.V.; Gimeno-Adelantado, J.V.; Jimenez, M. Determination of multiple mycotoxins in feedstuffs by combined use of UPLC–MS/MS and UPLC–QTOF–MS. Food Chem. 2018, 267, 140–148. [Google Scholar] [CrossRef]
- Arraché, E.R.S.; Fontes, M.R.V.; Buffon, J.G.; Badiale-Furlong, E. Trichothecenes in wheat: Methodology, occurrence and human exposure risk. J. Cereal Sci. 2018, 82, 129–137. [Google Scholar] [CrossRef]
- Haque, M.A.; Wang, Y.; Shen, Z.; Li, X.; Saleemi, M.K.; He, C. Mycotoxin contamination and control strategy in human, domestic animal and poultry: A review. Microb. Pathog. 2020, 142, 104095. [Google Scholar] [CrossRef]
- Ferrigo, D.; Raiola, A.; Causin, R. Fusarium toxins in cereals: Occurrence, legislation, factors promoting the appearance and their management. Molecules 2016, 21, 627. [Google Scholar] [CrossRef]
- Yoshinari, T.; Ohashi, H.; Abe, R.; Kaigome, R.; Ohkawa, H.; Sugita-Konishi, Y. Development of a rapid method for the quantitative determination of deoxynivalenol using Quenchbody. Anal. Chim. Acta 2015, 888, 126–130. [Google Scholar] [CrossRef]
- Ibáñez-Vea, M.; Martínez, R.; González-Peñas, E.; Lizarraga, E.; López de Cerain, A. Co-occurrence of aflatoxins, ochratoxin A and zearalenone in breakfast cereals from spanish market. Food Control 2011, 22, 1949–1955. [Google Scholar] [CrossRef]
- Nathanail, A.V.; Syvähuoko, J.; Malachová, A.; Jestoi, M.; Varga, E.; Michlmayr, H.; Adam, G.; Sieviläinen, E.; Berthiller, F.; Peltonen, K. Simultaneous determination of major type A and B trichothecenes, zearalenone and certain modified metabolites in Finnish cereal grains with a novel liquid chromatography-tandem mass spectrometric method. Anal. Bioanal. Chem. 2015, 407, 4745–4755. [Google Scholar] [CrossRef]
- Nathanail, A.V.; Sarikaya, E.; Jestoi, M.; Godula, M.; Peltonen, K. Determination of deoxynivalenol and deoxynivalenol-3-glucoside in wheat and barley using liquid chromatography coupled to mass spectrometry: On-line clean-up versus conventional sample preparation techniques. J. Chromatogr. A 2014, 1374, 31–39. [Google Scholar] [CrossRef]
- Gab-Allah, M.A.; Choi, K.; Kim, B. Development of isotope dilution-liquid chromatography/tandem mass spectrometry for the accurate determination of type-A trichothecenes in grains. Food Chem. 2021, 344, 128698. [Google Scholar] [CrossRef]
- Zou, Z.; He, Z.; Li, H.; Han, P.; Tang, J.; Xi, C.; Li, Y.; Zhang, L.; Li, X. Development and application of a method for the analysis of two trichothecenes: Deoxynivalenol and T-2 toxin in meat in China by HPLC–MS/MS. Meat Sci. 2012, 90, 613–617. [Google Scholar] [CrossRef]
- Gab-Allah, M.A.; Choi, K.; Kim, B. Accurate determination of type B trichothecenes and conjugated deoxynivalenol in grains by isotope dilution–liquid chromatography tandem mass spectrometry. Food Control 2021, 121, 107557. [Google Scholar] [CrossRef]
- Zhao, Z.; Rao, Q.; Song, S.; Liu, N.; Han, Z.; Hou, J.; Wu, A. Simultaneous determination of major type B trichothecenes and deoxynivalenol-3-glucoside in animal feed and raw materials using improved DSPE combined with LC-MS/MS. J. Chromatogr. B 2014, 963, 75–82. [Google Scholar] [CrossRef]
- Berthiller, F.; Dall’Asta, C.; Schuhmacher, R.; Lemmens, M.; Adam, G.; Krska, R. Masked mycotoxins: Determination of a deoxynivalenol glucoside in artificially and naturally contaminated wheat by liquid chromatography−tandem mass spectrometry. J. Agric. Food Chem. 2005, 53, 3421–3425. [Google Scholar] [CrossRef]
- Berthiller, F.; Krska, R.; Domig, K.J.; Kneifel, W.; Juge, N.; Schuhmacher, R.; Adam, G. Hydrolytic fate of deoxynivalenol-3-glucoside during digestion. Toxicol. Lett. 2011, 206, 264–267. [Google Scholar] [CrossRef] [PubMed]
- Berthiller, F.; Sulyok, M.; Krska, R.; Schuhmacher, R. Chromatographic methods for the simultaneous determination of mycotoxins and their conjugates in cereals. Int. J. Food Microbiol. 2007, 119, 33–37. [Google Scholar] [CrossRef] [PubMed]
- Lancova, K.; Hajslova, J.; Poustka, J.; Krplova, A.; Zachariasova, M.; Dostálek, P.; Sachambula, L. Transfer of Fusarium mycotoxins and ‘masked’deoxynivalenol (deoxynivalenol-3-glucoside) from field barley through malt to beer. Food Addit. Contam. 2008, 25, 732–744. [Google Scholar] [CrossRef] [PubMed]
- Rasmussen, P.; Nielsen, K.F.; Ghorbani, F.; Spliid, N.H.; Nielsen, G.; Jørgensen, L. Occurrence of different trichothecenes and deoxynivalenol-3-β-D-glucoside in naturally and artificially contaminated Danish cereal grains and whole maize plants. Mycotoxin Res. 2012, 28, 181–190. [Google Scholar] [CrossRef] [PubMed]
- Zachariasova, M.; Hajslova, J.; Kostelanska, M.; Poustka, J.; Krplova, A.; Cuhra, P.; Hochel, I. Deoxynivalenol and its conjugates in beer: A critical assessment of data obtained by enzyme-linked immunosorbent assay and liquid chromatography coupled to tandem mass spectrometry. Anal. Chim. Acta 2008, 625, 77–86. [Google Scholar] [CrossRef] [PubMed]
- Gab-Allah, M.A.; Mekete, K.G.; Choi, K.; Kim, B. Occurrence of major type-B trichothecenes and deoxynivalenol-3-glucoside in cereal-based products from Republic of Korea. J. Food Compos. Anal. 2021, 99, 103851. [Google Scholar] [CrossRef]
- Awad, W.; Ghareeb, K.; Böhm, J.; Zentek, J. The toxicological impacts of the Fusarium mycotoxin, deoxynivalenol, in poultry flocks with special reference to immunotoxicity. Toxins 2013, 5, 912–925. [Google Scholar] [CrossRef]
- Cinar, A.; Onbaşı, E. Mycotoxins: The hidden danger in foods. Mycotoxins Food Saf. 2019, 1–21. [Google Scholar] [CrossRef]
- Smith, M.-C.; Madec, S.; Coton, E.; Hymery, N. Natural co-occurrence of mycotoxins in foods and feeds and their in vitro combined toxicological effects. Toxins 2016, 8, 94. [Google Scholar] [CrossRef]
- Magan, N.; Olsen, M. Mycotoxins in Food: Detection and Control; Woodhead Publishing: Cambridge, UK, 2004. [Google Scholar]
- Shwab, E.K.; Keller, N.P. Regulation of secondary metabolite production in filamentous ascomycetes. Mycol. Res. 2008, 112, 225–230. [Google Scholar] [CrossRef]
- Mannaa, M.; Kim, K.D. Influence of temperature and water activity on deleterious fungi and mycotoxin production during grain storage. Mycobiology 2017, 45, 240–254. [Google Scholar] [CrossRef]
- D’Mello, J. Contaminants and toxins in animal feeds. FAO Feed and Food Safety page. In Animal Production and Health Division; FAO: Rome, Italy, 2001; Available online: https://agris.fao.org/agris-search/search.do?recordID=XF2013001395 (accessed on 2 December 2022).
- Degirmencioglu, N.; Esecali, H.; Cokal, Y.; Bilgic, M. From safety feed to safety food: The application of HACCP in mycotoxin control. Arch Zootech 2005, 8, 19–32. [Google Scholar]
- Devegowda, G.; Murthy, T. Mycotoxins: Their effects in poultry and some practical solutions. In The Mycotoxin Blue Book; Nottingham University Press: Nottingham, UK, 2005; pp. 25–56. [Google Scholar]
- Daou, R.; Joubrane, K.; Maroun, R.G.; Khabbaz, L.R.; Ismail, A.; El Khoury, A. Mycotoxins: Factors influencing production and control strategies. AIMS Agric. Food 2021, 6, 416–447. [Google Scholar] [CrossRef]
- Bryła, M.; Waśkiewicz, A.; Ksieniewicz-Woźniak, E.; Szymczyk, K.; Jędrzejczak, R. Modified Fusarium mycotoxins in cereals and their products—Metabolism, occurrence, and toxicity: An updated review. Molecules 2018, 23, 963. [Google Scholar] [CrossRef]
- Zain, M.E. Impact of mycotoxins on humans and animals. J. Saudi Chem. Soc. 2011, 15, 129–144. [Google Scholar] [CrossRef]
- Zhang, H.; Wang, B. Fate of deoxynivalenol and deoxynivalenol-3-glucoside during wheat milling and Chinese steamed bread processing. Food Control 2014, 44, 86–91. [Google Scholar] [CrossRef]
- Vidal, A.; Sanchis, V.; Ramos, A.J.; Marín, S. The fate of deoxynivalenol through wheat processing to food products. Curr. Opin. Food Sci. 2016, 11, 34–39. [Google Scholar] [CrossRef]
- Vidal, A.; Ambrosio, A.; Sanchis, V.; Ramos, A.J.; Marín, S. Enzyme bread improvers affect the stability of deoxynivalenol and deoxynivalenol-3-glucoside during breadmaking. Food Chem. 2016, 208, 288–296. [Google Scholar] [CrossRef]
- Zhang, H.; Wang, B. Fates of deoxynivalenol and deoxynivalenol-3-glucoside during bread and noodle processing. Food Control 2015, 50, 754–757. [Google Scholar] [CrossRef]
- Polak-Śliwińska, M.; Paszczyk, B. Trichothecenes in Food and Feed, Relevance to Human and Animal Health and Methods of Detection: A Systematic Review. Molecules 2021, 26, 454. [Google Scholar] [CrossRef]
- Pestka, J.J. Deoxynivalenol: Toxicity, mechanisms and animal health risks. Anim. Feed Sci. Technol. 2007, 137, 283–298. [Google Scholar] [CrossRef]
- Pinton, P.; Accensi, F.; Beauchamp, E.; Cossalter, A.-M.; Callu, P.; Grosjean, F.; Oswald, I.P. Ingestion of deoxynivalenol (DON) contaminated feed alters the pig vaccinal immune responses. Toxicol. Lett. 2008, 177, 215–222. [Google Scholar] [CrossRef] [PubMed]
- Khaneghah, A.M.; Martins, L.M.; von Hertwig, A.M.; Bertoldo, R.; Sant’Ana, A.S. Deoxynivalenol and its masked forms: Characteristics, incidence, control and fate during wheat and wheat based products processing-A review. Trends Food Sci. Technol. 2018, 71, 13–24. [Google Scholar] [CrossRef]
- Arunachalam, C.; Doohan, F.M. Trichothecene toxicity in eukaryotes: Cellular and molecular mechanisms in plants and animals. Toxicol. Lett. 2013, 217, 149–158. [Google Scholar] [CrossRef] [PubMed]
- Payros, D.; Alassane-Kpembi, I.; Pierron, A.; Loiseau, N.; Pinton, P.; Oswald, I.P. Toxicology of deoxynivalenol and its acetylated and modified forms. Arch. Toxicol. 2016, 90, 2931–2957. [Google Scholar] [CrossRef]
- Pestka, J.J. Mechanisms of deoxynivalenol-induced gene expression and apoptosis. Food Addit. Contam. 2008, 25, 1128–1140. [Google Scholar] [CrossRef]
- Pierron, A.; Alassane-Kpembi, I.; Oswald, I.P. Impact of mycotoxin on immune response and consequences for pig health. Anim. Nutr. 2016, 2, 63–68. [Google Scholar] [CrossRef]
- Savard, C.; Pinilla, V.; Provost, C.; Gagnon, C.A.; Chorfi, Y. In vivo effect of deoxynivalenol (DON) naturally contaminated feed on porcine reproductive and respiratory syndrome virus (PRRSV) infection. Vet. Microbiol. 2014, 174, 419–426. [Google Scholar] [CrossRef]
- Akbari, P.; Braber, S.; Varasteh, S.; Alizadeh, A.; Garssen, J.; Fink-Gremmels, J. The intestinal barrier as an emerging target in the toxicological assessment of mycotoxins. Arch. Toxicol. 2017, 91, 1007–1029. [Google Scholar] [CrossRef]
- Murugesan, G.; Ledoux, D.; Naehrer, K.; Berthiller, F.; Applegate, T.; Grenier, B.; Phillips, T.; Schatzmayr, G. Prevalence and effects of mycotoxins on poultry health and performance, and recent development in mycotoxin counteracting strategies. Poult. Sci. 2015, 94, 1298–1315. [Google Scholar] [CrossRef]
- Van De Walle, J.; Sergent, T.; Piront, N.; Toussaint, O.; Schneider, Y.-J.; Larondelle, Y. Deoxynivalenol affects in vitro intestinal epithelial cell barrier integrity through inhibition of protein synthesis. Toxicol. Appl. Pharmacol. 2010, 245, 291–298. [Google Scholar] [CrossRef]
- Hsia, C.C.; Wu, Z.Y.; Li, Y.S.; Zhang, F.; Sun, Z.T. Nivalenol, a main Fusarium toxin in dietary foods from high-risk areas of cancer of esophagus and gastric cardia in China, induced benign and malignant tumors in mice. Oncol. Rep. 2004, 12, 449–456. [Google Scholar] [CrossRef]
- Wu, W.; Flannery, B.M.; Sugita-Konishi, Y.; Watanabe, M.; Zhang, H.; Pestka, J.J. Comparison of murine anorectic responses to the 8-ketotrichothecenes 3-acetyldeoxynivalenol, 15-acetyldeoxynivalenol, fusarenon X and nivalenol. Food Chem. Toxicol. 2012, 50, 2056–2061. [Google Scholar] [CrossRef]
- Wu, W.; Bates, M.A.; Bursian, S.J.; Link, J.E.; Flannery, B.M.; Sugita-Konishi, Y.; Watanabe, M.; Zhang, H.; Pestka, J.J. Comparison of emetic potencies of the 8-ketotrichothecenes deoxynivalenol, 15-acetyldeoxynivalenol, 3-acetyldeoxynivalenol, fusarenon X, and nivalenol. Toxicol. Sci. 2013, 131, 279–291. [Google Scholar] [CrossRef]
- Ueno, Y.; Nakajima, M.; Sakai, K.; Ishii, K.; Sato, N.; Shimada, N. Comparative toxicology of trichothec mycotoxins: Inhibition of protein synthesis in animal cells. J. Biochem. 1973, 74, 285–296. [Google Scholar]
- Garaleviciene, D.; Pettersson, H.; Elwinger, K. Effects on health and blood plasma parameters of laying hens by pure nivalenol in the diet. J. Anim. Physiol. Anim. Nutr. 2002, 86, 389–398. [Google Scholar] [CrossRef]
- EFSA Panel on Contaminants in the Food Chain (CONTAM). Scientific Opinion on the risks for human and animal health related to the presence of modified forms of certain mycotoxins in food and feed. EFSA J. 2014, 12, 3916. [Google Scholar]
- Pettersson, H.; Hedman, R.; Engström, B.; Elwinger, K.; Fossum, O. Nivalenol in Swedish cereals1—Occurrence, production and toxicity towards chickens. Food Addit. Contam. 1995, 12, 373–376. [Google Scholar] [CrossRef]
- EFSA Panel on Contaminants in the Food Chain (CONTAM). Scientific Opinion on risks for animal and public health related to the presence of nivalenol in food and feed. EFSA J. 2013, 11, 3262. [Google Scholar] [CrossRef]
- Claeys, L.; Romano, C.; De Ruyck, K.; Wilson, H.; Fervers, B.; Korenjak, M.; Zavadil, J.; Gunter, M.J.; De Saeger, S.; De Boevre, M. Mycotoxin exposure and human cancer risk: A systematic review of epidemiological studies. Compr. Rev. Food Sci. Food Saf. 2020, 19, 1449–1464. [Google Scholar] [CrossRef]
- Maul, R.; Müller, C.; Rieß, S.; Koch, M.; Methner, F.-J.; Irene, N. Germination induces the glucosylation of the Fusarium mycotoxin deoxynivalenol in various grains. Food Chem. 2012, 131, 274–279. [Google Scholar] [CrossRef]
- Pinton, P.; Tsybulskyy, D.; Lucioli, J.; Laffitte, J.; Callu, P.; Lyazhri, F.; Grosjean, F.; Bracarense, A.P.; Kolf-Clauw, M.; Oswald, I.P. Toxicity of deoxynivalenol and its acetylated derivatives on the intestine: Differential effects on morphology, barrier function, tight junction proteins, and mitogen-activated protein kinases. Toxicol. Sci. 2012, 130, 180–190. [Google Scholar] [CrossRef] [PubMed]
- Ajandouz, E.H.; Berdah, S.; Moutardier, V.; Bege, T.; Birnbaum, D.J.; Perrier, J.; Di Pasquale, E.; Maresca, M. Hydrolytic fate of 3/15-acetyldeoxynivalenol in humans: Specific deacetylation by the small intestine and liver revealed using in vitro and ex vivo approaches. Toxins 2016, 8, 232. [Google Scholar] [CrossRef] [PubMed]
- Pinton, P.; Braicu, C.; Nougayrede, J.-P.; Laffitte, J.; Taranu, I.; Oswald, I.P. Deoxynivalenol impairs porcine intestinal barrier function and decreases the protein expression of claudin-4 through a mitogen-activated protein kinase-dependent mechanism. J. Nutr. 2010, 140, 1956–1962. [Google Scholar] [CrossRef] [PubMed]
- Juan-García, A.; Juan, C.; Tolosa, J.; Ruiz, M.-J. Effects of deoxynivalenol, 3-acetyl-deoxynivalenol and 15-acetyl-deoxynivalenol on parameters associated with oxidative stress in HepG2 cells. Mycotoxin Res. 2019, 35, 197–205. [Google Scholar] [CrossRef]
- Juan-García, A.; Taroncher, M.; Font, G.; Ruiz, M.-J. Micronucleus induction and cell cycle alterations produced by deoxynivalenol and its acetylated derivatives in individual and combined exposure on HepG2 cells. Food Chem. Toxicol. 2018, 118, 719–725. [Google Scholar] [CrossRef]
- Sugita-Konishi, Y.; Pestka, J.J. Differential upregulation of TNF-α, IL-6, and IL-8 production by deoxynivalenol (vomitoxin) and other 8-ketotrichothecenes in a human macrophage model. J. Toxicol. Environ. Health Part A 2001, 64, 619–636. [Google Scholar] [CrossRef]
- Tomar, R.; Blakley, B.; Schiefer, H.; DeCoteau, W. In vitro effects of 3-acetyl-deoxynivalenol on the immune response of human peripheral blood lymphocytes. Int. J. Immunopharmacol. 1986, 8, 125–130. [Google Scholar] [CrossRef]
- Forsell, J.; Jensen, R.; Tai, J.-H.; Witt, M.; Lin, W.; Pestka, J. Comparison of acute toxicities of deoxynivalenol (vomitoxin) and 15-acetyldeoxynivalenol in the B6C3F1 mouse. Food Chem. Toxicol. 1987, 25, 155–162. [Google Scholar] [CrossRef]
- Kadota, T.; Furusawa, H.; Hirano, S.; Tajima, O.; Kamata, Y.; Sugita-Konishi, Y. Comparative study of deoxynivalenol, 3-acetyldeoxynivalenol, and 15-acetyldeoxynivalenol on intestinal transport and IL-8 secretion in the human cell line Caco-2. Toxicol. Vitr. 2013, 27, 1888–1895. [Google Scholar] [CrossRef]
- He, Y.; Yin, X.; Dong, J.; Yang, Q.; Wu, Y.; Gong, Z. Transcriptome Analysis of Caco-2 Cells upon the Exposure of Mycotoxin Deoxynivalenol and Its Acetylated Derivatives. Toxins 2021, 13, 167. [Google Scholar] [CrossRef]
- Alassane-Kpembi, I.; Puel, O.; Oswald, I.P. Toxicological interactions between the mycotoxins deoxynivalenol, nivalenol and their acetylated derivatives in intestinal epithelial cells. Arch. Toxicol. 2015, 89, 1337–1346. [Google Scholar] [CrossRef]
- Yin, S.; Liu, X.; Fan, L.; Hu, H. Mechanisms of cell death induction by food-borne mycotoxins. Crit. Rev. Food Sci. Nutr. 2018, 58, 1406–1417. [Google Scholar] [CrossRef]
- Wu, W.; Zhou, H.-R.; Bursian, S.J.; Pan, X.; Link, J.E.; Berthiller, F.; Adam, G.; Krantis, A.; Durst, T.; Pestka, J.J. Comparison of anorectic and emetic potencies of deoxynivalenol (vomitoxin) to the plant metabolite deoxynivalenol-3-glucoside and synthetic deoxynivalenol derivatives EN139528 and EN139544. Toxicol. Sci. 2014, 142, 167–181. [Google Scholar] [CrossRef]
- Wu, W.; He, K.; Zhou, H.-R.; Berthiller, F.; Adam, G.; Sugita-Konishi, Y.; Watanabe, M.; Krantis, A.; Durst, T.; Zhang, H. Effects of oral exposure to naturally-occurring and synthetic deoxynivalenol congeners on proinflammatory cytokine and chemokine mRNA expression in the mouse. Toxicol. Appl. Pharmacol. 2014, 278, 107–115. [Google Scholar] [CrossRef]
- Pierron, A.; Mimoun, S.; Murate, L.S.; Loiseau, N.; Lippi, Y.; Bracarense, A.-P.F.; Liaubet, L.; Schatzmayr, G.; Berthiller, F.; Moll, W.-D. Intestinal toxicity of the masked mycotoxin deoxynivalenol-3-β-D-glucoside. Arch. Toxicol. 2016, 90, 2037–2046. [Google Scholar] [CrossRef]
- Dall’Erta, A.; Cirlini, M.; Dall’Asta, M.; Del Rio, D.; Galaverna, G.; Dall’Asta, C. Masked mycotoxins are efficiently hydrolyzed by human colonic microbiota releasing their aglycones. Chem. Res. Toxicol. 2013, 26, 305–312. [Google Scholar] [CrossRef]
- Gareis, M.; Bauer, J.; Thiem, J.; Plank, G.; Grabley, S.; Gedek, B. Cleavage of zearalenone-glycoside, a “masked” mycotoxin, during digestion in swine. J. Vet. Med. Ser. B 1990, 37, 236–240. [Google Scholar] [CrossRef]
- Joint, F.; World Health Organization; WHO Expert Committee on Food Additives. Evaluation of Certain Contaminants in Food: Seventy-Second [72nd] Report of the Joint FAO/WHO Expert Committee on Food Additives; World Health Organization: Geneva, Switzerland, 2011. [Google Scholar]
- Juan, C.; Ritieni, A.; Mañes, J. Occurrence of Fusarium mycotoxins in Italian cereal and cereal products from organic farming. Food Chem. 2013, 141, 1747–1755. [Google Scholar] [CrossRef]
- Ok, H.E.; Choi, S.-W.; Chung, S.H.; Kang, Y.-W.; Kim, D.-S.; Chun, H.S. Natural occurrence of type-B trichothecene mycotoxins in Republic of Korea cereal-based products. Food Addit. Contam. Part B 2011, 4, 132–140. [Google Scholar] [CrossRef]
- Piacentini, K.C.; Rocha, L.; Savi, G.; Carnielli-Queiroz, L.; Almeida, F.; Minella, E.; Corrêa, B. Occurrence of deoxynivalenol and zearalenone in brewing barley grains from Brazil. Mycotoxin Res. 2018, 34, 173–178. [Google Scholar] [CrossRef] [PubMed]
- Kyprianou, M. Commission Regulation (EC) No 1126/2007. of 28 September 2007. Amending regulation (EC) no 1881/2006 setting maximum levels for certain contaminants in foodstuffs as regards Fusarium toxins in maize and maize products. J. Eur. Union 2007, 255, 14–17. [Google Scholar]
- Mishra, S.; Srivastava, S.; Dewangan, J.; Divakar, A.; Kumar Rath, S. Global occurrence of deoxynivalenol in food commodities and exposure risk assessment in humans in the last decade: A survey. Crit. Rev. Food Sci. Nutr. 2022, 60, 1346–1374. [Google Scholar] [CrossRef]
- Kamle, M.; Mahato, D.K.; Gupta, A.; Pandhi, S.; Sharma, B.; Dhawan, K.; Mishra, S.; Kumar, M.; Tripathi, A.D.; Rasane, P. Deoxynivalenol: An Overview on Occurrence, Chemistry, Biosynthesis, Health Effects and Its Detection, Management, and Control Strategies in Food and Feed. Microbiol. Res. 2022, 13, 292–314. [Google Scholar] [CrossRef]
- Scientific Committee on Food (SCF). Opinion of the Scientific Committee on Food on Fusarium Toxins. Part 6: Group Evaluation of T-2 Toxin, HT-2 Toxin, Nivalenol and Deoxynivalenol; Scientific Opinions European Commission: Brussels, Belgium, 2002; pp. 1–12. [Google Scholar]
- Authority, E.F.S. Deoxynivalenol in food and feed: Occurrence and exposure. EFSA J. 2013, 11, 3379. [Google Scholar]
- Knutsen, H.K.; Alexander, J.; Barregård, L.; Bignami, M.; Brüschweiler, B.; Ceccatelli, S.; Cottrill, B.; Dinovi, M.; Grasl-Kraupp, B. Risks to human and animal health related to the presence of deoxynivalenol and its acetylated and modified forms in food and feed. EFSA J. 2017, 15, e04718. [Google Scholar]
- Chun, H. Risk assessment of mycotoxin contamination in Republic of Korea foods. In Proceedings of FFTCeKU 2011 Conference, International Seminar on Risk Assessment and Risk Management of Mycotoxins for Food Safety in Asia; Kasetsart University: Bangkok, Thailand, 2011. [Google Scholar]
- Ji, F.; Xu, J.; Liu, X.; Yin, X.; Shi, J. Natural occurrence of deoxynivalenol and zearalenone in wheat from Jiangsu province, China. Food Chem. 2014, 157, 393–397. [Google Scholar] [CrossRef]
- Juan, C.; Ritieni, A.; Mañes, J. Determination of trichothecenes and zearalenones in grain cereal, flour and bread by liquid chromatography tandem mass spectrometry. Food Chem. 2012, 134, 2389–2397. [Google Scholar] [CrossRef]
- Liu, Y.; Lu, Y.; Wang, L.; Chang, F.; Yang, L. Occurrence of deoxynivalenol in wheat, Hebei Province, China. Food Chem. 2016, 197, 1271–1274. [Google Scholar] [CrossRef]
- Xu, W.; Han, X.; Li, F. Co-occurrence of multi-mycotoxins in wheat grains harvested in Anhui province, China. Food Control 2019, 96, 180–185. [Google Scholar] [CrossRef]
- Bryła, M.; Ksieniewicz-Woźniak, E.; Waśkiewicz, A.; Szymczyk, K.; Jędrzejczak, R. Natural occurrence of nivalenol, deoxynivalenol, and deoxynivalenol-3-glucoside in Polish winter wheat. Toxins 2018, 10, 81. [Google Scholar] [CrossRef]
- Palacios, S.A.; Erazo, J.G.; Ciasca, B.; Lattanzio, V.M.; Reynoso, M.M.; Farnochi, M.C.; Torres, A.M. Occurrence of deoxynivalenol and deoxynivalenol-3-glucoside in durum wheat from Argentina. Food Chem. 2017, 230, 728–734. [Google Scholar] [CrossRef]
- Iqbal, S.Z.; Usman, S.; Razis, A.F.A.; Basheir Ali, N.; Saif, T.; Asi, M.R. Assessment of deoxynivalenol in wheat, corn and its products and estimation of dietary intake. Int. J. Environ. Res. Public Health 2020, 17, 5602. [Google Scholar] [CrossRef]
- Pleadin, J.; Sokolović, M.; Perši, N.; Zadravec, M.; Jaki, V.; Vulić, A. Contamination of maize with deoxynivalenol and zearalenone in Croatia. Food Control 2012, 28, 94–98. [Google Scholar] [CrossRef]
- Berthiller, F.; Dall’Asta, C.; Corradini, R.; Marchelli, R.; Sulyok, M.; Krska, R.; Adam, G.; Schuhmacher, R. Occurrence of deoxynivalenol and its 3-β-D-glucoside in wheat and maize. Food Addit. Contam. 2009, 26, 507–511. [Google Scholar] [CrossRef]
- Gab-Allah, M.A.; Tahoun, I.F.; Yamani, R.N.; Rend, E.A.; Shehata, A.B. Natural occurrence of deoxynivalenol, nivalenol and deoxynivalenol-3-glucoside in cereal-derived products from Egypt. Food Control 2022, 137, 108974. [Google Scholar] [CrossRef]
- Adejumo, T.O.; Hettwer, U.; Karlovsky, P. Occurrence of Fusarium species and trichothecenes in Nigerian maize. Int. J. Food Microbiol. 2007, 116, 350–357. [Google Scholar] [CrossRef]
- Egbuta, M.; Wanza, M.; Dutton, M. Evaluation of five major mycotoxins co-contaminating two cereal grains from Nigeria. Int. J. Biochem. Res. Rev. 2015, 6, 160–169. [Google Scholar] [CrossRef]
- Jajić, I.; Jurić, V.; Glamočić, D.; Abramović, B. Occurrence of deoxynivalenol in maize and wheat in Serbia. Int. J. Mol. Sci. 2008, 9, 2114–2126. [Google Scholar] [CrossRef]
- Camardo Leggieri, M.; Bertuzzi, T.; Pietri, A.; Battilani, P. Mycotoxin occurrence in maize produced in Northern Italy over the years 2009–2011: Focus on the role of crop related factors. Phytopathol. Mediterr. 2015, 212–221. [Google Scholar] [CrossRef]
- Kamala, A.; Ortiz, J.; Kimanya, M.; Haesaert, G.; Donoso, S.; Tiisekwa, B.; De Meulenaer, B. Multiple mycotoxin co-occurrence in maize grown in three agro-ecological zones of Tanzania. Food Control 2015, 54, 208–215. [Google Scholar] [CrossRef]
- Czembor, E.; Stępień, Ł.; Waśkiewicz, A. Effect of environmental factors on Fusarium species and associated mycotoxins in maize grain grown in Poland. PLoS ONE 2015, 10, e0133644. [Google Scholar] [CrossRef] [PubMed]
- Tima, H.; Brückner, A.; Mohácsi-Farkas, C.; Kiskó, G. Fusarium mycotoxins in cereals harvested from Hungarian fields. Food Addit. Contam. Part B 2016, 9, 127–131. [Google Scholar] [CrossRef] [PubMed]
- Mishra, S.; Ansari, K.M.; Dwivedi, P.D.; Pandey, H.P.; Das, M. Occurrence of deoxynivalenol in cereals and exposure risk assessment in Indian population. Food Control 2013, 30, 549–555. [Google Scholar] [CrossRef]
- Pleadin, J.; Vahčić, N.; Perši, N.; Ševelj, D.; Markov, K.; Frece, J. Fusarium mycotoxins’ occurrence in cereals harvested from Croatian fields. Food Control 2013, 32, 49–54. [Google Scholar]
- Tarazona, A.; Gómez, J.V.; Mateo, F.; Jiménez, M.; Mateo, E.M. Potential Health Risk Associated with Mycotoxins in Oat Grains Consumed in Spain. Toxins 2021, 13, 421. [Google Scholar] [CrossRef]
- Schöneberg, T.; Jenny, E.; Wettstein, F.E.; Bucheli, T.D.; Mascher, F.; Bertossa, M.; Musa, T.; Seifert, K.; Gräfenhan, T.; Keller, B. Occurrence of Fusarium species and mycotoxins in Swiss oats—Impact of cropping factors. Eur. J. Agron. 2018, 92, 123–132. [Google Scholar] [CrossRef]
- Ji, X.; Yang, H.; Wang, J.; Li, R.; Zhao, H.; Xu, J.; Xiao, Y.; Tang, B.; Qian, M. Occurrence of deoxynivalenol (DON) in cereal-based food products marketed through e-commerce stores and an assessment of dietary exposure of Chinese consumers to DON. Food Control 2018, 92, 391–398. [Google Scholar] [CrossRef]
- Islam, M.N.; Tabassum, M.; Banik, M.; Daayf, F.; Fernando, W.D.; Harris, L.J.; Sura, S.; Wang, X. Naturally Occurring Fusarium Species and Mycotoxins in Oat Grains from Manitoba, Canada. Toxins 2021, 13, 670. [Google Scholar] [CrossRef]
- Lee, S.Y.; Woo, S.Y.; Tian, F.; Song, J.; Michlmayr, H.; Kim, J.-B.; Chun, H.S. Occurrence of deoxynivalenol, nivalenol, and their glucosides in Republic of Korea market foods and estimation of their population exposure through food consumption. Toxins 2020, 12, 89. [Google Scholar] [CrossRef]
- Fredlund, E.; Gidlund, A.; Sulyok, M.; Börjesson, T.; Krska, R.; Olsen, M.; Lindblad, M. Deoxynivalenol and other selected Fusarium toxins in Swedish oats—Occurrence and correlation to specific Fusarium species. Int. J. Food Microbiol. 2013, 167, 276–283. [Google Scholar] [CrossRef]
- Edwards, S.G. Impact of agronomic and climatic factors on the mycotoxin content of harvested oats in the United Kingdom. Food Addit. Contam. Part A 2017, 34, 2230–2241. [Google Scholar] [CrossRef]
- Soleimany, F.; Jinap, S.; Abas, F. Determination of mycotoxins in cereals by liquid chromatography tandem mass spectrometry. Food Chem. 2012, 130, 1055–1060. [Google Scholar] [CrossRef]
- Sliková, S.; Gavurníková, S.; Hasana, R.; Mináriková, M.; Gregová, E. Deoxynivalenol in grains of oats and wheat produced in Slovakia. Poljopr. I Sumar. 2016, 62, 343. [Google Scholar] [CrossRef]
- Ok, H.; Chang, H.-J.; Choi, S.-W.; Cho, T.; Oh, K.; Chun, H. Occurrence and intake of deoxynivalenol in cereal-based products marketed in Republic of Korea during 2007–2008. Food Addit. Contam. 2009, 2, 154–161. [Google Scholar] [CrossRef]
- Bryła, M.; Ksieniewicz-Woźniak, E.; Waśkiewicz, A.; Szymczyk, K.; Jędrzejczak, R. Co-occurrence of nivalenol, deoxynivalenol and deoxynivalenol-3-glucoside in beer samples. Food Control 2018, 92, 319–324. [Google Scholar] [CrossRef]
- Golge, O.; Kabak, B. Occurrence of deoxynivalenol and zearalenone in cereals and cereal products from Turkey. Food Control 2020, 110, 106982. [Google Scholar] [CrossRef]
- Majeed, S.; De Boevre, M.; De Saeger, S.; Rauf, W.; Tawab, A.; Rahman, M.; Iqbal, M. Multiple mycotoxins in rice: Occurrence and health risk assessment in children and adults of Punjab, Pakistan. Toxins 2018, 10, 77. [Google Scholar] [CrossRef]
- Ok, H.E.; Lee, S.Y.; Chun, H.S. Occurrence and simultaneous determination of nivalenol and deoxynivalenol in rice and bran by HPLC-UV detection and immunoaffinity cleanup. Food Control 2018, 87, 53–59. [Google Scholar] [CrossRef]
- Rodríguez-Carrasco, Y.; Moltó, J.C.; Berrada, H.; Mañes, J. A survey of trichothecenes, zearalenone and patulin in milled grain-based products using GC–MS/MS. Food Chem. 2014, 146, 212–219. [Google Scholar] [CrossRef]
- Moreira, G.M.; Nicolli, C.P.; Gomes, L.B.; Ogoshi, C.; Scheuermann, K.K.; Silva-Lobo, V.L.; Schurt, D.A.; Ritieni, A.; Moretti, A.; Pfenning, L.H. Nationwide survey reveals high diversity of Fusarium species and related mycotoxins in Brazilian rice: 2014 and 2015 harvests. Food Control 2020, 113, 107171. [Google Scholar] [CrossRef]
- Chilaka, C.A.; De Boevre, M.; Atanda, O.O.; De Saeger, S. Occurrence of Fusarium mycotoxins in cereal crops and processed products (Ogi) from Nigeria. Toxins 2016, 8, 342. [Google Scholar] [CrossRef] [PubMed]
- Olopade, B.K.; Oranusi, S.U.; Nwinyi, O.C.; Gbashi, S.; Njobeh, P.B. Occurrences of Deoxynivalenol, Zearalenone and some of their masked forms in selected cereals from Southwest Nigeria. NFS J. 2021, 23, 24–29. [Google Scholar] [CrossRef]
- Ssepuuya, G.; Van Poucke, C.; Ediage, E.N.; Mulholland, C.; Tritscher, A.; Verger, P.; Kenny, M.; Bessy, C.; De Saeger, S. Mycotoxin contamination of sorghum and its contribution to human dietary exposure in four sub-Saharan countries. Food Addit. Contam. Part A 2018, 35, 1384–1393. [Google Scholar] [CrossRef] [PubMed]
- Ayalew, A.; Fehrmann, H.; Lepschy, J.; Beck, R.; Abate, D. Natural occurrence of mycotoxins in staple cereals from Ethiopia. Mycopathologia 2006, 162, 57–63. [Google Scholar] [CrossRef]
- Hanvi, D.M.; Lawson-Evi, P.; De Boevre, M.; Goto, C.; De Saeger, S.; Eklu-Gadegbeku, K. Natural occurrence of mycotoxins in maize and sorghum in Togo. Mycotoxin Res. 2019, 35, 321–327. [Google Scholar] [CrossRef]
- Serrano, A.; Font, G.; Ruiz, M.; Ferrer, E. Co-occurrence and risk assessment of mycotoxins in food and diet from Mediterranean area. Food Chem. 2012, 135, 423–429. [Google Scholar] [CrossRef]
- Oueslati, S.; Blesa, J.; Moltó, J.C.; Ghorbel, A.; Mañes, J. Presence of mycotoxins in sorghum and intake estimation in Tunisia. Food Addit. Contam. Part A 2014, 31, 307–318. [Google Scholar] [CrossRef]
- Martos, P.; Thompson, W.; Diaz, G. Multiresidue mycotoxin analysis in wheat, barley, oats, rye and maize grain by high-performance liquid chromatography-tandem mass spectrometry. World Mycotoxin J. 2010, 3, 205–223. [Google Scholar] [CrossRef]
- Gottschalk, C.; Barthel, J.; Engelhardt, G.; Bauer, J.; Meyer, K. Simultaneous determination of type A, B and D trichothecenes and their occurrence in cereals and cereal products. Food Addit. Contam. 2009, 26, 1273–1289. [Google Scholar] [CrossRef]
- Stuper-Szablewska, K.; Perkowski, J. Level of contamination with mycobiota and contents of mycotoxins from the group of trichothecenes in grain of wheat, oats, barley, rye and triticale harvested in Poland in 2006–2008. Ann. Agric. Environ. Med. 2017, 24, 49–55. [Google Scholar] [CrossRef]
- Rasmussen, P.H.; Ghorbani, F.; Berg, T. Deoxynivalenol and other Fusarium toxins in wheat and rye flours on the Danish market. Food Addit. Contam. 2003, 20, 396–404. [Google Scholar] [CrossRef]
- Jin, Z.; Gillespie, J.; Barr, J.; Wiersma, J.J.; Sorrells, M.E.; Zwinger, S.; Gross, T.; Cumming, J.; Bergstrom, G.C.; Brueggeman, R. Malting of Fusarium head blight-infected rye (Secale cereale): Growth of Fusarium graminearum, trichothecene production, and the impact on malt quality. Toxins 2018, 10, 369. [Google Scholar] [CrossRef]
- Zhao, J.; Cheng, T.; Xu, W.; Han, X.; Zhang, J.; Zhang, H.; Wang, C.; Fanning, S.; Li, F. Natural co-occurrence of multi-mycotoxins in unprocessed wheat grains from China. Food Control 2021, 130, 108321. [Google Scholar] [CrossRef]
- Vidal, A.; Marín, S.; Ramos, A.J.; Cano-Sancho, G.; Sanchis, V. Determination of aflatoxins, deoxynivalenol, ochratoxin A and zearalenone in wheat and oat based bran supplements sold in the Spanish market. Food Chem. Toxicol. 2013, 53, 133–138. [Google Scholar] [CrossRef]
- Alkadri, D.; Rubert, J.; Prodi, A.; Pisi, A.; Mañes, J.; Soler, C. Natural co-occurrence of mycotoxins in wheat grains from Italy and Syria. Food Chem. 2014, 157, 111–118. [Google Scholar] [CrossRef]
- Blesa, J.; Moltó, J.-C.; El Akhdari, S.; Mañes, J.; Zinedine, A. Simultaneous determination of Fusarium mycotoxins in wheat grain from Morocco by liquid chromatography coupled to triple quadrupole mass spectrometry. Food Control 2014, 46, 1–5. [Google Scholar] [CrossRef]
- Yan, P.; Liu, Z.; Liu, S.; Yao, L.; Liu, Y.; Wu, Y.; Gong, Z. Natural occurrence of deoxynivalenol and its acetylated derivatives in Chinese maize and wheat collected in 2017. Toxins 2020, 12, 200. [Google Scholar] [CrossRef]
- Tralamazza, S.M.; Bemvenuti, R.H.; Zorzete, P.; de Souza Garcia, F.; Corrêa, B. Fungal diversity and natural occurrence of deoxynivalenol and zearalenone in freshly harvested wheat grains from Brazil. Food Chem. 2016, 196, 445–450. [Google Scholar] [CrossRef]
- Tanaka, H.; Sugita-Konishi, Y.; Takino, M.; Tanaka, T.; Toriba, A.; Hayakawa, K. A survey of the occurrence of Fusarium mycotoxins in biscuits in Japan by using LC/MS. J. Health Sci. 2010, 56, 188–194. [Google Scholar] [CrossRef]
- Vogelgsang, S.; Musa, T.; Bänziger, I.; Kägi, A.; Bucheli, T.D.; Wettstein, F.E.; Pasquali, M.; Forrer, H.-R. Fusarium mycotoxins in Swiss wheat: A survey of growers’ samples between 2007 and 2014 shows strong year and minor geographic effects. Toxins 2017, 9, 246. [Google Scholar] [CrossRef] [PubMed]
- Abdallah, M.F.; Girgin, G.; Baydar, T.; Krska, R.; Sulyok, M. Occurrence of multiple mycotoxins and other fungal metabolites in animal feed and maize samples from Egypt using LC-MS/MS. J. Sci. Food Agric. 2017, 97, 4419–4428. [Google Scholar] [CrossRef] [PubMed]
- Binder, E.; Tan, L.; Chin, L.; Handl, J.; Richard, J. Worldwide occurrence of mycotoxins in commodities, feeds and feed ingredients. Anim. Feed Sci. Technol. 2007, 137, 265–282. [Google Scholar] [CrossRef]
- Castillo, M.-Á.; Montes, R.; Navarro, A.; Segarra, R.; Cuesta, G.; Hernández, E. Occurrence of deoxynivalenol and nivalenol in Spanish corn-based food products. J. Food Compos. Anal. 2008, 21, 423–427. [Google Scholar] [CrossRef]
- Abia, W.A.; Warth, B.; Sulyok, M.; Krska, R.; Tchana, A.N.; Njobeh, P.B.; Dutton, M.F.; Moundipa, P.F. Determination of multi-mycotoxin occurrence in cereals, nuts and their products in Cameroon by liquid chromatography tandem mass spectrometry (LC-MS/MS). Food Control 2013, 31, 438–453. [Google Scholar] [CrossRef]
- Xing, F.; Liu, X.; Wang, L.; Selvaraj, J.N.; Jin, N.; Wang, Y.; Zhao, Y.; Liu, Y. Distribution and variation of fungi and major mycotoxins in pre-and post-nature drying maize in North China Plain. Food Control 2017, 80, 244–251. [Google Scholar] [CrossRef]
- Tutelyan, V.; Zakharova, L.; Sedova, I.; Perederyaev, O.; Aristarkhova, T.; Eller, K. Fusariotoxins in Russian Federation 2005–2010 grain harvests. Food Addit. Contam. Part B 2013, 6, 139–145. [Google Scholar] [CrossRef]
- Hofgaard, I.; Aamot, H.; Torp, T.; Jestoi, M.; Lattanzio, V.; Klemsdal, S.; Waalwijk, C.; Van der Lee, T.; Brodal, G. Associations between Fusarium species and mycotoxins in oats and spring wheat from farmers’ fields in Norway over a six-year period. World Mycotoxin J. 2016, 9, 365–378. [Google Scholar] [CrossRef]
- Bensassi, F.; Rjiba, I.; Zarrouk, A.; Rhouma, A.; Hajlaoui, M.; Bacha, H. Deoxynivalenol contamination in Tunisian barley in the 2009 harvest. Food Addit. Contam. Part B 2011, 4, 205–211. [Google Scholar] [CrossRef]
- Al-Julaifi, M.Z.; Al-Falih, A.M. Detection of trichothecenes in animal feeds and foodstuffs during the years 1997 to 2000 in Saudi Arabia. J. Food Prot. 2001, 64, 1603–1606. [Google Scholar] [CrossRef]
- Makun, H.A.; Dutton, M.F.; Njobeh, P.B.; Mwanza, M.; Kabiru, A.Y. Natural multi-occurrence of mycotoxins in rice from Niger State, Nigeria. Mycotoxin Res. 2011, 27, 97–104. [Google Scholar] [CrossRef]
- Roscoe, V.; Lombaert, G.; Huzel, V.; Neumann, G.; Melietio, J.; Kitchen, D.; Kotello, S.; Krakalovich, T.; Trelka, R.; Scott, P. Mycotoxins in breakfast cereals from the Canadian retail market: A 3-year survey. Food Addit. Contam. 2008, 25, 347–355. [Google Scholar] [CrossRef]
- Lee, M.J.; Wee, C.-D.; Ham, H.; Choi, J.-H.; Baek, J.S.; Lim, S.B.; Lee, T.; Kim, J.-S.; Jang, J.Y. Survey on Fusarium mycotoxin contamination in oat, sorghum, adlay, and proso millet during the harvest season in Republic of Korea. J. Food Hyg. Saf. 2020, 35, 13–22. [Google Scholar] [CrossRef]
- Błajet-Kosicka, A.; Twarużek, M.; Kosicki, R.; Sibiorowska, E.; Grajewski, J. Co-occurrence and evaluation of mycotoxins in organic and conventional rye grain and products. Food Control 2014, 38, 61–66. [Google Scholar] [CrossRef]
- Rahmani, A.; Jinap, S.; Soleimany, F. Qualitative and quantitative analysis of mycotoxins. Compr. Rev. Food Sci. Food Saf. 2009, 8, 202–251. [Google Scholar] [CrossRef]
- Meneely, J.P.; Ricci, F.; van Egmond, H.P.; Elliott, C.T. Current methods of analysis for the determination of trichothecene mycotoxins in food. TrAC Trends Anal. Chem. 2011, 30, 192–203. [Google Scholar] [CrossRef]
- Alshannaq, A.; Yu, J.-H. Occurrence, toxicity, and analysis of major mycotoxins in food. Int. J. Environ. Res. Public Health 2017, 14, 632. [Google Scholar] [CrossRef]
- Thompson, M.; Ellison, S.L.; Wood, R. Harmonized guidelines for single-laboratory validation of methods of analysis (IUPAC Technical Report). Pure Appl. Chem. 2002, 74, 835–855. [Google Scholar] [CrossRef]
- Shehata, A.; Mohamed, G.; Gab-Allah, M. Development of crude oil reference material certified for the concentrations of sulfur, iron, nickel, vanadium and magnesium. Mapan 2017, 32, 101–112. [Google Scholar] [CrossRef]
- Pereira, V.; Fernandes, J.; Cunha, S. Mycotoxins in cereals and related foodstuffs: A review on occurrence and recent methods of analysis. Trends Food Sci. Technol. 2014, 36, 96–136. [Google Scholar] [CrossRef]
- Shanakhat, H.; Sorrentino, A.; Raiola, A.; Romano, A.; Masi, P.; Cavella, S. Current methods for mycotoxins analysis and innovative strategies for their reduction in cereals: An overview. J. Sci. Food Agric. 2018, 98, 4003–4013. [Google Scholar] [CrossRef] [PubMed]
- Boyd, R.K.; Basic, C.; Bethem, R.A. Trace Quantitative Analysis by Mass Spectrometry; John Wiley & Sons: Hoboken, NJ, USA, 2011. [Google Scholar]
- Commission Regulation (EC). Commission Regulation (EC) No 401/2006 of 23 February 2006 laying down the methods of sampling and analysis for the official control of the levels of mycotoxins in foodstuffs. Off. J. Eur. Union 2006, 70, 12–34. [Google Scholar]
- Gab-Allah, M.; Goda, E.S.; Shehata, A.; Gamal, H. Critical review on the analytical methods for the determination of sulfur and trace elements in crude oil. Crit. Rev. Anal. Chem. 2020, 50, 161–178. [Google Scholar] [CrossRef] [PubMed]
- Shehata, A.; Mohamed, G.; Gab-Allah, M. Simple spectrophotometric method for determination of iron in crude oil. Pet. Chem. 2017, 57, 1007–1011. [Google Scholar] [CrossRef]
- Tahoun, I.F.; Gab-Allah, M.A.; Yamani, R.N.; Shehata, A.B. Development and validation of a reliable LC-MS/MS method for simultaneous determination of deoxynivalenol and T-2 toxin in maize and oats. Microchem. J. 2021, 169, 106599. [Google Scholar] [CrossRef]
- Tahoun, I.F.; Rend, E.A.; Gab-Allah, M.A. Preparation and value assignment of parabens and phenoxyethanol in cosmetic cream certified reference material. J. Chem. Metrol. 2021, 15, 1–10. [Google Scholar] [CrossRef]
- Tahoun, I.F.; Gab-Allah, M.A. Development of ledipasvir and sofosbuvir pure certified reference materials for improving quality of pharmaceutical analysis. J. Chem. Metrol. 2022, 16, 68–77. [Google Scholar] [CrossRef]
- Chen, P.; Xiang, B.; Shi, H.; Yu, P.; Song, Y.; Li, S. Recent advances on type A trichothecenes in food and feed: Analysis, prevalence, toxicity, and decontamination techniques. Food Control 2020, 118, 107371. [Google Scholar] [CrossRef]
- Dong, M.; Si, W.; Jiang, K.; Nie, D.; Wu, Y.; Zhao, Z.; De Saeger, S.; Han, Z. Multi-walled carbon nanotubes as solid-phase extraction sorbents for simultaneous determination of type A trichothecenes in maize, wheat and rice by ultra-high performance liquid chromatography-tandem mass spectrometry. J. Chromatogr. A 2015, 1423, 177–182. [Google Scholar] [CrossRef]
- Kirimker, S.E.; Turksoy, S.; Kabak, B. Assessment of dietary exposure to deoxynivalenol and fumonisin in the population of infants and toddlers in Turkey. Food Chem. Toxicol. 2020, 140, 111304. [Google Scholar] [CrossRef]
- Giménez, I.; Herrera, M.; Escobar, J.; Ferruz, E.; Lorán, S.; Herrera, A.; Ariño, A. Distribution of deoxynivalenol and zearalenone in milled germ during wheat milling and analysis of toxin levels in wheat germ and wheat germ oil. Food Control 2013, 34, 268–273. [Google Scholar] [CrossRef]
- Deng, Y.; Wang, Y.; Deng, Q.; Sun, L.; Wang, R.; Wang, X.; Liao, J.; Gooneratne, R. Simultaneous quantification of aflatoxin B1, T-2 toxin, ochratoxin A and deoxynivalenol in dried seafood products by LC-MS/MS. Toxins 2020, 12, 488. [Google Scholar] [CrossRef]
- González-Osnaya, L.; Cortés, C.; Soriano, J.; Moltó, J.; Mañes, J. Occurrence of deoxynivalenol and T-2 toxin in bread and pasta commercialised in Spain. Food Chem. 2011, 124, 156–161. [Google Scholar] [CrossRef]
- Jin, P.; Han, Z.; Cai, Z.; Wu, Y.; Ren, Y. Simultaneous determination of 10 mycotoxins in grain by ultra-high-performance liquid chromatography–tandem mass spectrometry using 13C15-deoxynivalenol as internal standard. Food Addit. Contam. Part A 2010, 27, 1701–1713. [Google Scholar] [CrossRef]
- Nagl, V.; Schatzmayr, G. Deoxynivalenol and its masked forms in food and feed. Curr. Opin. Food Sci. 2015, 5, 43–49. [Google Scholar] [CrossRef]
- Barros, G.; García, D.; Oviedo, M.; Ramirez, M.; Torres, A.; Lattanzio, V.; Pascale, M.; Chulze, S. Survey of T-2 and HT-2 toxins in soybean and soy meal from Argentina using immunoaffinity clean-up and high performance liquid chromatography. World Mycotoxin J. 2011, 4, 189–197. [Google Scholar] [CrossRef]
- Valle-Algarra, F.M.; Mateo, E.M.; Mateo, R.; Gimeno-Adelantado, J.V.; Jiménez, M. Determination of type A and type B trichothecenes in paprika and chili pepper using LC-triple quadrupole–MS and GC–ECD. Talanta 2011, 84, 1112–1117. [Google Scholar] [CrossRef]
- Sulyok, M.; Berthiller, F.; Krska, R.; Schuhmacher, R. Development and validation of a liquid chromatography/tandem mass spectrometric method for the determination of 39 mycotoxins in wheat and maize. Rapid Commun. Mass Spectrom. 2006, 20, 2649–2659. [Google Scholar] [CrossRef]
- Beltrán, E.; Ibáñez, M.; Sancho, J.V.; Hernández, F. Determination of mycotoxins in different food commodities by ultra-high-pressure liquid chromatography coupled to triple quadrupole mass spectrometry. Rapid Commun. Mass Spectrom. 2009, 23, 1801–1809. [Google Scholar] [CrossRef]
- Snyder, L.R.; Kirkland, J.J.; Glajch, J.L. Practical HPLC Method Development; John Wiley & Sons: Hoboken, NJ, USA, 2012. [Google Scholar]
- Montes, R.; Segarra, R.; Castillo, M.-Á. Trichothecenes in breakfast cereals from the Spanish retail market. J. Food Compos. Anal. 2012, 27, 38–44. [Google Scholar] [CrossRef]
- Ibáñez-Vea, M.; Lizarraga, E.; González-Peñas, E. Simultaneous determination of type-A and type-B trichothecenes in barley samples by GC–MS. Food Control 2011, 22, 1428–1434. [Google Scholar] [CrossRef]
- Muscarella, M.; Iammarino, M.; Nardiello, D.; Palermo, C.; Centonze, D. Determination of deoxynivalenol and nivalenol by liquid chromatography and fluorimetric detection with on-line chemical post-column derivatization. Talanta 2012, 97, 145–149. [Google Scholar] [CrossRef] [PubMed]
- Zuo, H.G.; Zhu, J.X.; Shi, L.; Zhan, C.R.; Guo, P.; Wang, Y.; Zhang, Y.; Liu, J. Development of a novel immunoaffinity column for the determination of deoxynivalenol and its acetylated derivatives in cereals. Food Anal. Methods 2018, 11, 2252–2260. [Google Scholar] [CrossRef]
- Pascale, M.; Panzarini, G.; Powers, S.; Visconti, A. Determination of deoxynivalenol and nivalenol in wheat by ultra-performance liquid chromatography/photodiode-array detector and immunoaffinity column cleanup. Food Anal. Methods 2014, 7, 555–562. [Google Scholar] [CrossRef]
- Trombete, F.; Barros, A.; Vieira, M.; Saldanha, T.; Venâncio, A.; Fraga, M. Simultaneous determination of deoxynivalenol, deoxynivalenol-3-glucoside and nivalenol in wheat grains by HPLC-PDA with immunoaffinity column cleanup. Food Anal. Methods 2016, 9, 2579–2586. [Google Scholar] [CrossRef]
- Royer, D.; Humpf, H.-U.; Guy, P. Quantitative analysis of Fusarium mycotoxins in maize using accelerated solvent extraction before liquid chromatography/atmospheric pressure chemical ionization tandem mass spectrometry. Food Addit. Contam. 2004, 21, 678–692. [Google Scholar] [CrossRef]
- Josephs, R.; Krska, R.; Grasserbauer, M.; Broekaert, J. Determination of trichothecene mycotoxins in wheat by use of supercritical fluid extraction and high-performance liquid chromatography with diode array detection or gas chromatography with electron capture detection. J. Chromatogr. A 1998, 795, 297–304. [Google Scholar] [CrossRef]
- Panasiuk, Ł.; Jedziniak, P.; Pietruszka, K.; Posyniak, A. Simultaneous determination of deoxynivalenol, its modified forms, nivalenol and fusarenone-X in feedstuffs by the liquid chromatography–tandem mass spectrometry method. Toxins 2020, 12, 362. [Google Scholar] [CrossRef]
- Razzazi-Fazeli, E.; Reiter, E. Sample preparation and clean up in mycotoxin analysis: Principles, applications and recent developments. Determ. Mycotoxins Mycotoxigenic Fungi Food Feed 2011, 37–70. [Google Scholar] [CrossRef]
- Tomczyk, Ł.; Szablewski, T.; Stuper-Szablewska, K.; Nowaczewski, S.; Cegielska-Radziejewska, R. The influence of the conditions of acquisition and storage of table eggs on changes in their quality and the presence of mycobiota and Fusarium mycotoxins. Poult. Sci. 2019, 98, 2964–2971. [Google Scholar] [CrossRef]
- Twarużek, M.; Błajet-Kosicka, A.; Kosicki, R.; Grajewski, J. Mycotoxin Analytical Methods. In Environmental Mycology in Public Health; Elsevier: Amsterdam, The Netherlands, 2016; pp. 363–386. [Google Scholar]
- Han, Z.; Ren, Y.; Zhu, J.; Cai, Z.; Chen, Y.; Luan, L.; Wu, Y. Multianalysis of 35 mycotoxins in traditional Chinese medicines by ultra-high-performance liquid chromatography–tandem mass spectrometry coupled with accelerated solvent extraction. J. Agric. Food Chem. 2012, 60, 8233–8247. [Google Scholar] [CrossRef]
- Mahmoud, A.F.; Escrivá, L.; Rodríguez-Carrasco, Y.; Moltó, J.C.; Berrada, H. Determination of trichothecenes in chicken liver using gas chromatography coupled with triple-quadrupole mass spectrometry. LWT 2018, 93, 237–242. [Google Scholar] [CrossRef]
- Gab-Allah, M.A.; Tahoun, I.F.; Yamani, R.N.; Rend, E.A.; Shehata, A.B. Eco-friendly and sensitive analytical method for determination of T-2 toxin and HT-2 toxin in cereal products using UPLC-MS/MS. J. Food Compos. Anal. 2022, 107, 104395. [Google Scholar] [CrossRef]
- Dong, H.; Xian, Y.; Xiao, K.; Wu, Y.; Zhu, L.; He, J. Development and comparison of single-step solid phase extraction and QuEChERS clean-up for the analysis of 7 mycotoxins in fruits and vegetables during storage by UHPLC-MS/MS. Food Chem. 2019, 274, 471–479. [Google Scholar] [CrossRef]
- Klötzel, M.; Lauber, U.; Humpf, H.U. A new solid phase extraction clean-up method for the determination of 12 type A and B trichothecenes in cereals and cereal-based food by LC-MS/MS. Mol. Nutr. Food Res. 2006, 50, 261–269. [Google Scholar] [CrossRef]
- McMaster, N.; Acharya, B.; Harich, K.; Grothe, J.; Mehl, H.L.; Schmale, D.G. Quantification of the mycotoxin deoxynivalenol (DON) in sorghum using GC-MS and a stable isotope dilution assay (SIDA). Food Anal. Methods 2019, 12, 2334–2343. [Google Scholar] [CrossRef]
- Sasanya, J.; Hall, C.; Wolf-Hall, C. Analysis of deoxynivalenol, masked deoxynivalenol, and Fusarium graminearum pigment in wheat samples, using liquid chromatography–UV–mass spectrometry. J. Food Prot. 2008, 71, 1205–1213. [Google Scholar] [CrossRef]
- Pereira, V.; Fernandes, J.; Cunha, S. Comparative assessment of three cleanup procedures after QuEChERS extraction for determination of trichothecenes (type A and type B) in processed cereal-based baby foods by GC–MS. Food Chem. 2015, 182, 143–149. [Google Scholar] [CrossRef]
- Anastassiades, M.; Lehotay, S.J.; Štajnbaher, D.; Schenck, F.J. Fast and easy multiresidue method employing acetonitrile extraction/partitioning and “dispersive solid-phase extraction” for the determination of pesticide residues in produce. J. AOAC Int. 2003, 86, 412–431. [Google Scholar] [CrossRef]
- Perestrelo, R.; Silva, P.; Porto-Figueira, P.; Pereira, J.A.; Silva, C.; Medina, S.; Câmara, J.S. QuEChERS-Fundamentals, relevant improvements, applications and future trends. Anal. Chim. Acta 2019, 1070, 1–28. [Google Scholar] [CrossRef]
- Tamura, M.; Mochizuki, N.; Nagatomi, Y.; Harayama, K.; Toriba, A.; Hayakawa, K. A method for simultaneous determination of 20 Fusarium toxins in cereals by high-resolution liquid chromatography-orbitrap mass spectrometry with a pentafluorophenyl column. Toxins 2015, 7, 1664–1682. [Google Scholar] [CrossRef] [PubMed]
- Miró-Abella, E.; Herrero, P.; Canela, N.; Arola, L.; Borrull, F.; Ras, R.; Fontanals, N. Determination of mycotoxins in plant-based beverages using QuEChERS and liquid chromatography–tandem mass spectrometry. Food Chem. 2017, 229, 366–372. [Google Scholar] [CrossRef] [PubMed]
- Rodriguez-Carrasco, Y.; Fattore, M.; Albrizio, S.; Berrada, H.; Manes, J. Occurrence of Fusarium mycotoxins and their dietary intake through beer consumption by the European population. Food Chem. 2015, 178, 149–155. [Google Scholar] [CrossRef] [PubMed]
- Dall’Asta, C.; Dall’Erta, A.; Mantovani, P.; Massi, A.; Galaverna, G. Occurrence of deoxynivalenol and deoxynivalenol-3-glucoside in durum wheat. World Mycotoxin J. 2013, 6, 83–91. [Google Scholar] [CrossRef]
- Cunha, S.C.; Fernandes, J.O. Development and validation of a method based on a QuEChERS procedure and heart-cutting GC-MS for determination of five mycotoxins in cereal products. J. Sep. Sci. 2010, 33, 600–609. [Google Scholar] [CrossRef]
- Frenich, A.G.; Romero-González, R.; Gómez-Pérez, M.L.; Vidal, J.L.M. Multi-mycotoxin analysis in eggs using a QuEChERS-based extraction procedure and ultra-high-pressure liquid chromatography coupled to triple quadrupole mass spectrometry. J. Chromatogr. A 2011, 1218, 4349–4356. [Google Scholar] [CrossRef]
- Zhou, Q.; Li, F.; Chen, L.; Jiang, D. Quantitative analysis of 10 mycotoxins in wheat flour by ultrahigh performance liquid chromatography-tandem mass spectrometry with a modified QuEChERS strategy. J. Food Sci. 2016, 81, T2886–T2890. [Google Scholar] [CrossRef]
- Lim, C.W.; Tai, S.H.; Lee, L.M.; Chan, S.H. Analytical method for the accurate determination of tricothecenes in grains using LC-MS/MS: A comparison between MRM transition and MS3 quantitation. Anal. Bioanal. Chem. 2012, 403, 2801–2806. [Google Scholar] [CrossRef]
- Neira, M.; Pacin, A.; Martinez, E.; Moltó, G.; Resnik, S. The effects of bakery processing on natural deoxynivalenol contamination. Int. J. Food Microbiol. 1997, 37, 21–25. [Google Scholar] [CrossRef]
- Tutelyan, V.A. Deoxynivalenol in cereals in Russia. Toxicol. Lett. 2004, 153, 173–179. [Google Scholar] [CrossRef]
- Njobeh, P.B.; Dutton, M.F.; Koch, S.H.; Chuturgoon, A.A.; Stoev, S.D.; Mosonik, J.S. Simultaneous occurrence of mycotoxins in human food commodities from Cameroon. Mycotoxin Res. 2010, 26, 47–57. [Google Scholar] [CrossRef]
- Rocha, D.F.d.L.; Oliveira, M.d.S.; Furlong, E.B.; Junges, A.; Paroul, N.; Valduga, E.; Backes, G.T.; Zeni, J.; Cansian, R.L. Evaluation of the TLC quantification method and occurrence of deoxynivalenol in wheat flour of southern Brazil. Food Addit. Contam. Part A 2017, 34, 2220–2229. [Google Scholar] [CrossRef]
- Lee, S.Y.; Woo, S.Y.; Malachová, A.; Michlmayr, H.; Kim, S.-H.; Kang, G.J.; Chun, H.S. Simple validated method for simultaneous determination of deoxynivalenol, nivalenol, and their 3-β-D-glucosides in baby formula and Republic of Korea rice wine via HPLC-UV with immunoaffinity cleanup. Food Addit. Contam. Part A 2019, 36, 964–975. [Google Scholar] [CrossRef]
- Almeida, A.P.d.; Lamardo, L.C.A.; Shundo, L.; Silva, S.A.d.; Navas, S.A.; Alaburda, J.; Ruvieri, V.; Sabino, M. Occurrence of deoxynivalenol in wheat flour, instant noodle and biscuits commercialised in Brazil. Food Addit. Contam. Part B 2016, 9, 251–255. [Google Scholar] [CrossRef]
- Gonçalves, C.; Mischke, C.; Stroka, J. Determination of deoxynivalenol and its major conjugates in cereals using an organic solvent-free extraction and IAC clean-up coupled in-line with HPLC-PCD-FLD. Food Addit. Contam. Part A 2020, 37, 1765–1776. [Google Scholar] [CrossRef]
- Pascale, M.N. Detection methods for mycotoxins in cereal grains and cereal products. Zbornik Matice Srpske za Prirodne Nauke 2009, 15–25. [Google Scholar] [CrossRef]
- Zheng, Y.; Hossen, S.M.; Sago, Y.; Yoshida, M.; Nakagawa, H.; Nagashima, H.; Okadome, H.; Nakajima, T.; Kushiro, M. Effect of milling on the content of deoxynivalenol, nivalenol, and zearalenone in Japanese wheat. Food Control 2014, 40, 193–197. [Google Scholar] [CrossRef]
- Skendi, A.; Irakli, M.N.; Papageorgiou, M.D. Optimized and validated high-performance liquid chromatography method for the determination of deoxynivalenol and aflatoxins in cereals. J. Sep. Sci. 2016, 39, 1425–1432. [Google Scholar] [CrossRef]
- Montanha, F.P.; Anater, A.; Burchard, J.F.; Luciano, F.B.; Meca, G.; Manyes, L.; Pimpão, C.T. Mycotoxins in dry-cured meats: A review. Food Chem. Toxicol. 2018, 111, 494–502. [Google Scholar] [CrossRef]
- Desmarchelier, A.; Tessiot, S.; Bessaire, T.; Racault, L.; Fiorese, E.; Urbani, A.; Chan, W.-C.; Cheng, P.; Mottier, P. Combining the quick, easy, cheap, effective, rugged and safe approach and clean-up by immunoaffinity column for the analysis of 15 mycotoxins by isotope dilution liquid chromatography tandem mass spectrometry. J. Chromatogr. A 2014, 1337, 75–84. [Google Scholar] [CrossRef]
- Narváez, A.; Rodríguez-Carrasco, Y.; Castaldo, L.; Izzo, L.; Ritieni, A. Ultra-high-performance liquid chromatography coupled with quadrupole Orbitrap high-resolution mass spectrometry for multi-residue analysis of mycotoxins and pesticides in botanical nutraceuticals. Toxins 2020, 12, 114. [Google Scholar] [CrossRef] [PubMed]
- Gab-Allah, M.A.; Lijalem, Y.G.; Yu, H.; Lee, S.; Baek, S.-Y.; Han, J.; Choi, K.; Kim, B. Development of a certified reference material for the accurate determination of type B trichothecenes in corn. Food Chem. 2022, 404, 134542. [Google Scholar] [CrossRef] [PubMed]
- Jia, W.; Shi, L.; Zhang, F.; Fan, C.; Chang, J.; Chu, X. Multiplexing data independent untargeted workflows for mycotoxins screening on a quadrupole-Orbitrap high resolution mass spectrometry platform. Food Chem. 2019, 278, 67–76. [Google Scholar] [CrossRef] [PubMed]
- Nakagawa, H.; Ohmichi, K.; Sakamoto, S.; Sago, Y.; Kushiro, M.; Nagashima, H.; Yoshida, M.; Nakajima, T. Detection of a new Fusarium masked mycotoxin in wheat grain by high-resolution LC–Orbitrap™ MS. Food Addit. Contam. Part A 2011, 28, 1447–1456. [Google Scholar] [CrossRef] [PubMed]
- Narváez, A.; Rodríguez-Carrasco, Y.; Castaldo, L.; Izzo, L.; Graziani, G.; Ritieni, A. Occurrence and exposure assessment of mycotoxins in ready-to-eat tree nut products through ultra-high performance liquid chromatography coupled with high resolution q-orbitrap mass spectrometry. Metabolites 2020, 10, 344. [Google Scholar] [CrossRef]
- Zachariasova, M.; Lacina, O.; Malachova, A.; Kostelanska, M.; Poustka, J.; Godula, M.; Hajslova, J. Novel approaches in analysis of Fusarium mycotoxins in cereals employing ultra performance liquid chromatography coupled with high resolution mass spectrometry. Anal. Chim. Acta 2010, 662, 51–61. [Google Scholar] [CrossRef]
- Eke, Z.; Kende, A.; Torkos, K. Simultaneous detection of A and B trichothecenes by gas chromatography with flame ionization or mass selective detection. Microchem. J. 2004, 78, 211–216. [Google Scholar] [CrossRef]
- Schothorst, R.C.; Jekel, A.A. Determination of trichothecenes in wheat by capillary gas chromatography with flame ionisation detection. Food Chem. 2001, 73, 111–117. [Google Scholar] [CrossRef]
- Furlong, E.B.; Valente Soares, L.M. Gas chromatographic method for quantitation and confirmation of trichothecenes in wheat. J. AOAC Int. 1995, 78, 386–390. [Google Scholar] [CrossRef]
- Simsek, S.; Burgess, K.; Whitney, K.L.; Gu, Y.; Qian, S.Y. Analysis of deoxynivalenol and deoxynivalenol-3-glucoside in wheat. Food Control 2012, 26, 287–292. [Google Scholar] [CrossRef]
- Tanaka, T.; Hasegawa, A.; Matsuki, Y.; Ishii, K.; Ueno, Y. Improved methodology for the simultaneous detection of the trichothecene mycotoxins deoxynivalenol and nivalenol in cereals. Food Addit. Contam. 1985, 2, 125–137. [Google Scholar] [CrossRef]
- Rodríguez-Carrasco, Y.; Berrada, H.; Font, G.; Mañes, J. Multi-mycotoxin analysis in wheat semolina using an acetonitrile-based extraction procedure and gas chromatography–tandem mass spectrometry. J. Chromatogr. A 2012, 1270, 28–40. [Google Scholar] [CrossRef]
- Rodríguez-Carrasco, Y.; Font, G.; Moltó, J.C.; Berrada, H. Quantitative determination of trichothecenes in breadsticks by gas chromatography-triple quadrupole tandem mass spectrometry. Food Addit. Contam. Part A 2014, 31, 1422–1430. [Google Scholar] [CrossRef]
- Mahato, D.K.; Pandhi, S.; Kamle, M.; Gupta, A.; Sharma, B.; Panda, B.K.; Srivastava, S.; Kumar, M.; Selvakumar, R.; Pandey, A.K. Trichothecenes in food and feed: Occurrence, impact on human health and their detection and management strategies. Toxicon 2022, 208, 62–77. [Google Scholar] [CrossRef]
- dos Santos, J.S.; Souza, T.M.; Ono, E.Y.S.; Hashimoto, E.H.; Bassoi, M.C.; de Miranda, M.Z.; Itano, E.N.; Kawamura, O.; Hirooka, E.Y. Natural occurrence of deoxynivalenol in wheat from Paraná State, Brazil and estimated daily intake by wheat products. Food Chem. 2013, 138, 90–95. [Google Scholar] [CrossRef]
- Noda, K.; Hirakawa, Y.; Nishino, T.; Sekizuka, R.; Kishimoto, M.; Furukawa, T.; Sawane, S.; Matsunaga, A.; Kobayashi, N.; Sugita, K. Preparation of Monoclonal Antibodies Specifically Reacting with the Trichothecene Mycotoxins Nivalenol and 15-Acetylnivalenol via the Introduction of a Linker Molecule into Its C-15 Position. Toxins 2022, 14, 747. [Google Scholar] [CrossRef]
- Dzuman, Z.; Vaclavikova, M.; Polisenska, I.; Veprikova, Z.; Fenclova, M.; Zachariasova, M.; Hajslova, J. Enzyme-linked immunosorbent assay in analysis of deoxynivalenol: Investigation of the impact of sample matrix on results accuracy. Anal. Bioanal. Chem. 2014, 406, 505–514. [Google Scholar] [CrossRef]
- Zhang, Y.; Yang, J.; Lu, Y.; Ma, D.-Y.; Qi, M.G.; Wang, S. A competitive direct enzyme-linked immunosorbent assay for the rapid detection of deoxynivalenol: Development and application in agricultural products and feedstuff. Food Agric. Immunol. 2017, 28, 516–527. [Google Scholar] [CrossRef]
- Yoshizawa, T.; Kohno, H.; Ikeda, K.; Shinoda, T.; Yokohama, H.; Morita, K.; Kusada, O.; Kobayashi, Y. A practical method for measuring deoxynivalenol, nivalenol, and T-2+ HT-2 toxin in foods by an enzyme-linked immunosorbent assay using monoclonal antibodies. Biosci. Biotechnol. Biochem. 2004, 68, 2076–2085. [Google Scholar] [CrossRef]
- Liu, J.; Zanardi, S.; Powers, S.; Suman, M. Development and practical application in the cereal food industry of a rapid and quantitative lateral flow immunoassay for deoxynivalenol. Food Control 2012, 26, 88–91. [Google Scholar] [CrossRef]
- Yu, Q.; Li, H.; Li, C.; Zhang, S.; Shen, J.; Wang, Z. Gold nanoparticles-based lateral flow immunoassay with silver staining for simultaneous detection of fumonisin B1 and deoxynivalenol. Food Control 2015, 54, 347–352. [Google Scholar] [CrossRef]
- Goryacheva, I.Y.; Saeger, S.d.; Eremin, S.A.; Peteghem, C.V. Immunochemical methods for rapid mycotoxin detection: Evolution from single to multiple analyte screening: A review. Food Addit. Contam. 2007, 24, 1169–1183. [Google Scholar] [CrossRef] [PubMed]
- Nan, M.; Xue, H.; Bi, Y. Contamination, Detection and Control of Mycotoxins in Fruits and Vegetables. Toxins 2022, 14, 309. [Google Scholar] [CrossRef] [PubMed]
- Song, S.; Liu, N.; Zhao, Z.; Njumbe Ediage, E.; Wu, S.; Sun, C.; De Saeger, S.; Wu, A. Multiplex lateral flow immunoassay for mycotoxin determination. Anal. Chem. 2014, 86, 4995–5001. [Google Scholar] [CrossRef] [PubMed]
- Foubert, A.; Beloglazova, N.V.; Gordienko, A.; Tessier, M.D.; Drijvers, E.; Hens, Z.; De Saeger, S. Development of a rainbow lateral flow immunoassay for the simultaneous detection of four mycotoxins. J. Agric. Food Chem. 2017, 65, 7121–7130. [Google Scholar] [CrossRef]
- Kim, K.Y.; Shim, W.B.; Kim, J.S.; Chung, D.H. Development of a simultaneous lateral flow strip test for the rapid and simple detection of deoxynivalenol and zearalenone. J. Food Sci. 2014, 79, M2048–M2055. [Google Scholar] [CrossRef]
- Liu, Z.; Hua, Q.; Wang, J.; Liang, Z.; Li, J.; Wu, J.; Shen, X.; Lei, H.; Li, X. A smartphone-based dual detection mode device integrated with two lateral flow immunoassays for multiplex mycotoxins in cereals. Biosens. Bioelectron. 2020, 158, 112178. [Google Scholar] [CrossRef]
- Li, C.; Wen, K.; Mi, T.; Zhang, X.; Zhang, H.; Zhang, S.; Shen, J.; Wang, Z. A universal multi-wavelength fluorescence polarization immunoassay for multiplexed detection of mycotoxins in maize. Biosens. Bioelectron. 2016, 79, 258–265. [Google Scholar] [CrossRef]
- Maragos, C.; Jolley, M.; Nasir, M. Fluorescence polarization as a tool for the determination of deoxynivalenol in wheat. Food Addit. Contam. 2002, 19, 400–407. [Google Scholar] [CrossRef]
- Goda, E.S.; Gab-Allah, M.; Singu, B.S.; Yoon, K.R. Halloysite nanotubes based electrochemical sensors: A review. Microchem. J. 2019, 147, 1083–1096. [Google Scholar] [CrossRef]
- Wang, K.; He, B.; Xie, L.; Li, L.; Yang, J.; Liu, R.; Wei, M.; Jin, H.; Ren, W. Exonuclease III-assisted triple-amplified electrochemical aptasensor based on PtPd NPs/PEI-rGO for deoxynivalenol detection. Sens. Actuators B Chem. 2021, 349, 130767. [Google Scholar] [CrossRef]
- Ong, C.C.; Sangu, S.S.; Illias, N.M.; Gopinath, S.C.B.; Saheed, M.S.M. Iron nanoflorets on 3D-graphene-nickel: A ‘Dandelion’nanostructure for selective deoxynivalenol detection. Biosens. Bioelectron. 2020, 154, 112088. [Google Scholar] [CrossRef]
- Wen, X.; Huang, Q.; Nie, D.; Zhao, X.; Cao, H.; Wu, W.; Han, Z. A multifunctional n-doped cu–mofs (N–cu–mof) nanomaterial-driven electrochemical aptasensor for sensitive detection of deoxynivalenol. Molecules 2021, 26, 2243. [Google Scholar] [CrossRef]
- Zhang, Q.; Cai, Y.; Li, H.; Kong, D.-M.; Shen, H.-X. Sensitive dual DNAzymes-based sensors designed by grafting self-blocked G-quadruplex DNAzymes to the substrates of metal ion-triggered DNA/RNA-cleaving DNAzymes. Biosens. Bioelectron. 2012, 38, 331–336. [Google Scholar] [CrossRef]
- De Girolamo, A.; Cervellieri, S.; Cortese, M.; Porricelli, A.C.R.; Pascale, M.; Longobardi, F.; von Holst, C.; Ciaccheri, L.; Lippolis, V. Fourier transform near-infrared and mid-infrared spectroscopy as efficient tools for rapid screening of deoxynivalenol contamination in wheat bran. J. Sci. Food Agric. 2019, 99, 1946–1953. [Google Scholar] [CrossRef]
- Kamruzzaman, M.; ElMasry, G.; Sun, D.-W.; Allen, P. Non-destructive prediction and visualization of chemical composition in lamb meat using NIR hyperspectral imaging and multivariate regression. Innov. Food Sci. Emerg. Technol. 2012, 16, 218–226. [Google Scholar] [CrossRef]
- Shen, G.; Cao, Y.; Yin, X.; Dong, F.; Xu, J.; Shi, J.; Lee, Y.-W. Rapid and nondestructive quantification of deoxynivalenol in individual wheat kernels using near-infrared hyperspectral imaging and chemometrics. Food Control 2022, 131, 108420. [Google Scholar] [CrossRef]
- Femenias, A.; Bainotti, M.B.; Gatius, F.; Ramos, A.J.; Marín, S. Standardization of near infrared hyperspectral imaging for wheat single kernel sorting according to deoxynivalenol level. Food Res. Int. 2021, 139, 109925. [Google Scholar] [CrossRef]
- Tyska, D.; Mallmann, A.; Gressler, L.T.; Mallmann, C.A. Near-infrared spectroscopy as a tool for rapid screening of deoxynivalenol in wheat flour and its applicability in the industry. Food Addit. Contam. Part A 2021, 38, 1958–1968. [Google Scholar] [CrossRef]
- Lippolis, V.; Pascale, M.; Cervellieri, S.; Damascelli, A.; Visconti, A. Screening of deoxynivalenol contamination in durum wheat by MOS-based electronic nose and identification of the relevant pattern of volatile compounds. Food Control 2014, 37, 263–271. [Google Scholar] [CrossRef]
- Lippolis, V.; Cervellieri, S.; Damascelli, A.; Pascale, M.; Di Gioia, A.; Longobardi, F.; De Girolamo, A. Rapid prediction of deoxynivalenol contamination in wheat bran by MOS-based electronic nose and characterization of the relevant pattern of volatile compounds. J. Sci. Food Agric. 2018, 98, 4955–4962. [Google Scholar] [CrossRef] [PubMed]
- Campagnoli, A.; Cheli, F.; Polidori, C.; Zaninelli, M.; Zecca, O.; Savoini, G.; Pinotti, L.; Dell’Orto, V. Use of the electronic nose as a screening tool for the recognition of durum wheat naturally contaminated by deoxynivalenol: A preliminary approach. Sensors 2011, 11, 4899–4916. [Google Scholar] [CrossRef] [PubMed]
- Bueno, D.; Istamboulie, G.; Muñoz, R.; Marty, J.L. Determination of mycotoxins in food: A review of bioanalytical to analytical methods. Appl. Spectrosc. Rev. 2015, 50, 728–774. [Google Scholar] [CrossRef]
- Pascale, M.; Lippolis, V.; Maragos, C.M.; Visconti, A. Recent Developments in Trichothecene Analysis; ACS Publications: Washington, DC, USA, 2008. [Google Scholar]
- Kumar, P.; Mahato, D.K.; Gupta, A.; Pandey, S.; Paul, V.; Saurabh, V.; Pandey, A.K.; Selvakumar, R.; Barua, S.; Kapri, M. Nivalenol Mycotoxin Concerns in Foods: An Overview on Occurrence, Impact on Human and Animal Health and Its Detection and Management Strategies. Toxins 2022, 14, 527. [Google Scholar] [CrossRef] [PubMed]
- Beloglazova, N.V.; Sobolev, A.M.; Tessier, M.D.; Hens, Z.; Goryacheva, I.Y.; De Saeger, S. Fluorescently labelled multiplex lateral flow immunoassay based on cadmium-free quantum dots. Methods 2017, 116, 141–148. [Google Scholar] [CrossRef]
- Hamad, G.M.; Mehany, T.; Simal-Gandara, J.; Abou-Alella, S.; Esua, O.J.; Abdel-Wahhab, M.A.; Hafez, E.E. A review of recent innovative strategies for controlling mycotoxins in foods. Food Control 2022, 144, 109350. [Google Scholar] [CrossRef]
- Leslie, J.F.; Moretti, A.; Mesterházy, Á.; Ameye, M.; Audenaert, K.; Singh, P.K.; Richard-Forget, F.; Chulze, S.N.; Ponte, E.M.D.; Chala, A. Key global actions for mycotoxin management in wheat and other small grains. Toxins 2021, 13, 725. [Google Scholar] [CrossRef]
- Peivasteh-Roudsari, L.; Pirhadi, M.; Shahbazi, R.; Eghbaljoo-Gharehgheshlaghi, H.; Sepahi, M.; Mirza Alizadeh, A.; Tajdar-oranj, B.; Jazaeri, S. Mycotoxins: Impact on health and strategies for prevention and detoxification in the food chain. Food Rev. Int. 2021, 1–32. [Google Scholar] [CrossRef]
- Paul, P.; Lipps, P.; Hershman, D.; McMullen, M.; Draper, M.; Madden, L. Efficacy of triazole-based fungicides for Fusarium head blight and deoxynivalenol control in wheat: A multivariate meta-analysis. Phytopathology 2008, 98, 999–1011. [Google Scholar] [CrossRef]
- Beyer, M.; Klix, M.B.; Klink, H.; Verreet, J.-A. Quantifying the effects of previous crop, tillage, cultivar and triazole fungicides on the deoxynivalenol content of wheat grain—A review. J. Plant Dis. Prot. 2006, 113, 241–246. [Google Scholar] [CrossRef]
- Dill-Macky, R.; Jones, R. The effect of previous crop residues and tillage on Fusarium head blight of wheat. Plant Dis. 2000, 84, 71–76. [Google Scholar] [CrossRef]
- Wenda-Piesik, A.; Lemańczyk, G.; Twarużek, M.; Błajet-Kosicka, A.; Kazek, M.; Grajewski, J. Fusarium head blight incidence and detection of Fusarium toxins in wheat in relation to agronomic factors. Eur. J. Plant Pathol. 2017, 149, 515–531. [Google Scholar] [CrossRef]
- Nada, S.; Nikola, T.; Bozidar, U.; Ilija, D.; Andreja, R. Prevention and practical strategies to control mycotoxins in the wheat and maize chain. Food Control 2022, 136, 108855. [Google Scholar] [CrossRef]
- Codex Alimentarius Commission. Codex Alimentarius Commission. Code of practice for the prevention and reduction of mycotoxin contamination in cereals, including annexes on ochratoxin A, zearalenone, fumonisins and tricothecenes (CAC/RCP 51-2003). In Prevention and Reduction of Food and Feed Contamination; FAO/WHO: Rome Italy, 2003; pp. 1–13. [Google Scholar]
- Hofer, K.; Barmeier, G.; Schmidhalter, U.; Habler, K.; Rychlik, M.; Hückelhoven, R.; Hess, M. Effect of nitrogen fertilization on Fusarium head blight in spring barley. Crop Prot. 2016, 88, 18–27. [Google Scholar] [CrossRef]
- Ariño, A.; Juan, T.; Estopañan, G.; González-Cabo, J.F. Natural occurrence of Fusarium species, fumonisin production by toxigenic strains, and concentrations of fumonisins B1 and B2 in conventional and organic maize grown in Spain. J. Food Prot. 2007, 70, 151–156. [Google Scholar] [CrossRef]
- Lemmens, M.; Haim, K.; Lew, H.; Ruckenbauer, P. The effect of nitrogen fertilization on Fusarium head blight development and deoxynivalenol contamination in wheat. J. Phytopathol. 2004, 152, 1–8. [Google Scholar] [CrossRef]
- Aldred, D.; Magan, N. Prevention strategies for trichothecenes. Toxicol. Lett. 2004, 153, 165–171. [Google Scholar] [CrossRef]
- Bai, G.H.; Plattner, R.; Desjardins, A.; Kolb, F.; McIntosh, R. Resistance to Fusarium head blight and deoxynivalenol accumulation in wheat. Plant Breed. 2001, 120, 1–6. [Google Scholar] [CrossRef]
- Gaffar, F.Y.; Imani, J.; Karlovsky, P.; Koch, A.; Kogel, K.-H. Different components of the RNA interference machinery are required for conidiation, ascosporogenesis, virulence, deoxynivalenol production, and fungal inhibition by exogenous double-stranded RNA in the head blight pathogen Fusarium graminearum. Front. Microbiol. 2019, 10, 1662. [Google Scholar] [CrossRef]
- Bawa, A.; Anilakumar, K. Genetically modified foods: Safety, risks and public concerns—A review. J. Food Sci. Technol. 2013, 50, 1035–1046. [Google Scholar] [CrossRef]
- Torres, A.M.; Barros, G.G.; Palacios, S.A.; Chulze, S.N.; Battilani, P. Review on pre-and post-harvest management of peanuts to minimize aflatoxin contamination. Food Res. Int. 2014, 62, 11–19. [Google Scholar] [CrossRef]
- Munkvold, G. Crop management practices to minimize the risk of mycotoxins contamination in temperate-zone maize. Mycotoxin Reduct. Grain Chain. 2014, 1, 59–77. [Google Scholar]
- Awuah, R.; Ellis, W. Effects of some groundnut packaging methods and protection with Ocimum and Syzygium powders on kernel infection by fungi. Mycopathologia 2002, 154, 29–36. [Google Scholar] [CrossRef] [PubMed]
- Pitt, J.; Taniwaki, M.H.; Cole, M. Mycotoxin production in major crops as influenced by growing, harvesting, storage and processing, with emphasis on the achievement of Food Safety Objectives. Food Control 2013, 32, 205–215. [Google Scholar] [CrossRef]
- Belizán, M.M.; Gomez, A.d.l.A.; Baptista, Z.P.T.; Jimenez, C.M.; Matías, M.d.H.S.; Catalán, C.A.; Sampietro, D.A. Influence of water activity and temperature on growth and production of trichothecenes by Fusarium graminearum sensu stricto and related species in maize grains. Int. J. Food Microbiol. 2019, 305, 108242. [Google Scholar] [CrossRef]
- Magan, N.; Lacey, J. Water relations of some Fusarium species from infected wheat ears and grain. Trans. Br. Mycol. Soc. 1984, 83, 281–285. [Google Scholar] [CrossRef]
- Vidal, A.; Sanchis, V.; Ramos, A.J.; Marín, S. Thermal stability and kinetics of degradation of deoxynivalenol, deoxynivalenol conjugates and ochratoxin A during baking of wheat bakery products. Food Chem. 2015, 178, 276–286. [Google Scholar] [CrossRef]
- Stadler, D.; Berthiller, F.; Suman, M.; Schuhmacher, R.; Krska, R. Novel analytical methods to study the fate of mycotoxins during thermal food processing. Anal. Bioanal. Chem. 2020, 412, 9–16. [Google Scholar] [CrossRef]
- Vila-Donat, P.; Marín, S.; Sanchis, V.; Ramos, A. A review of the mycotoxin adsorbing agents, with an emphasis on their multi-binding capacity, for animal feed decontamination. Food Chem. Toxicol. 2018, 114, 246–259. [Google Scholar] [CrossRef]
- Elella, M.H.A.; Goda, E.S.; Gab-Allah, M.A.; Hong, S.E.; Pandit, B.; Lee, S.; Gamal, H.; ur Rehman, A.; Yoon, K.R. Xanthan gum-derived materials for applications in environment and eco-friendly materials: A review. J. Environ. Chem. Eng. 2021, 9, 104702. [Google Scholar] [CrossRef]
- Devreese, M.; Antonissen, G.; De Backer, P.; Croubels, S. Efficacy of active carbon towards the absorption of deoxynivalenol in pigs. Toxins 2014, 6, 2998–3004. [Google Scholar] [CrossRef]
- Gab-Allah, M.A.; Shehata, A.B. Determination of iron, nickel, and vanadium in crude oil by inductively coupled plasma optical emission spectrometry following microwave-assisted wet digestion. Chem. Pap. 2021, 75, 4239–4248. [Google Scholar] [CrossRef]
- Santos Alexandre, A.P.; Vela-Paredes, R.S.; Santos, A.S.; Costa, N.S.; Canniatti-Brazaca, S.G.; Calori-Domingues, M.A.; Augusto, P.E.D. Ozone treatment to reduce deoxynivalenol (DON) and zearalenone (ZEN) contamination in wheat bran and its impact on nutritional quality. Food Addit. Contam. Part A 2018, 35, 1189–1199. [Google Scholar] [CrossRef]
- Cromey, M.; Lauren, D.; Parkes, R.; Sinclair, K.; Shorter, S.; Wallace, A. Control of Fusarium head blight of wheat with fungicides. Australas. Plant Pathol. 2001, 30, 301–308. [Google Scholar] [CrossRef]
- Yao, Y.; Long, M. The biological detoxification of deoxynivalenol: A review. Food Chem. Toxicol. 2020, 145, 111649. [Google Scholar] [CrossRef]
- Elella, M.H.A.; Goda, E.; Gab-Allah, M.; Hong, S.E.; Lijalem, Y.G.; Yoon, K.R. Biodegradable Polymeric Nanocomposites for Wastewater Treatment. In Advances in Nanocomposite Materials for Environmental and Energy Harvesting Applications; Springer: Berlin/Heidelberg, Germany, 2022; pp. 245–298. [Google Scholar]
- Jard, G.; Liboz, T.; Mathieu, F.; Guyonvarc’h, A.; Lebrihi, A. Review of mycotoxin reduction in food and feed: From prevention in the field to detoxification by adsorption or transformation. Food Addit. Contam. Part A 2011, 28, 1590–1609. [Google Scholar] [CrossRef]
- Wachowska, U.; Packa, D.; Wiwart, M. Microbial inhibition of Fusarium pathogens and biological modification of trichothecenes in cereal grains. Toxins 2017, 9, 408. [Google Scholar] [CrossRef]
- He, C.; Fan, Y.; Liu, G.; Zhang, H. Isolation and identification of a strain of Aspergillus tubingensis with deoxynivalenol biotransformation capability. Int. J. Mol. Sci. 2008, 9, 2366–2375. [Google Scholar] [CrossRef]
- Tian, Y.; Tan, Y.; Liu, N.; Yan, Z.; Liao, Y.; Chen, J.; De Saeger, S.; Yang, H.; Zhang, Q.; Wu, A. Detoxification of deoxynivalenol via glycosylation represents novel insights on antagonistic activities of Trichoderma when confronted with Fusarium graminearum. Toxins 2016, 8, 335. [Google Scholar] [CrossRef]
- Garda-Buffon, J.; Kupski, L.; Badiale-Furlong, E. Deoxynivalenol (DON) degradation and peroxidase enzyme activity in submerged fermentation. Food Sci. Technol. 2011, 31, 198–203. [Google Scholar] [CrossRef]
- EFSA Panel on Additives and Products or Substances used in Animal Feed (FEEDAP). Scientific Opinion on the safety and efficacy of micro-organism DSM 11798 when used as a technological feed additive for pigs. EFSA J. 2013, 11, 3203. [Google Scholar]
- Juodeikiene, G.; Bartkiene, E.; Cernauskas, D.; Cizeikiene, D.; Zadeike, D.; Lele, V.; Bartkevics, V. Antifungal activity of lactic acid bacteria and their application for Fusarium mycotoxin reduction in malting wheat grains. LWT 2018, 89, 307–314. [Google Scholar] [CrossRef]
- Wang, S.; Hou, Q.; Guo, Q.; Zhang, J.; Sun, Y.; Wei, H.; Shen, L. Isolation and characterization of a deoxynivalenol-degrading bacterium Bacillus licheniformis YB9 with the capability of modulating intestinal microbial flora of mice. Toxins 2020, 12, 184. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Qin, X.; Guo, Y.; Zhang, Q.; Ma, Q.; Ji, C.; Zhao, L. Enzymatic degradation of deoxynivalenol by a novel bacterium, Pelagibacterium halotolerans ANSP101. Food Chem. Toxicol. 2020, 140, 111276. [Google Scholar] [CrossRef] [PubMed]
- Gao, X.; Mu, P.; Wen, J.; Sun, Y.; Chen, Q.; Deng, Y. Detoxification of trichothecene mycotoxins by a novel bacterium, Eggerthella sp. DII-9. Food Chem. Toxicol. 2018, 112, 310–319. [Google Scholar] [CrossRef]
- Garda, J.; Macedo, R.M.; Faria, R.; Bernd, L.; Dors, G.C.; Badiale-Furlong, E. Alcoholic fermentation effects on malt spiked with trichothecenes. Food Control 2005, 16, 423–428. [Google Scholar] [CrossRef]



| Cereal Grain | Year of Study | Detection Technique | Number of Samples | Type B Trichothecene | Incidence (%) | Mean (µg/kg) | Range (µg/kg) | Country | Reference |
|---|---|---|---|---|---|---|---|---|---|
| Wheat flour | 2012–2013 | UPLC-MS/MS | 672 | DON | 91.5 | 178 | 2.4–1130 | China | [95] |
| 3-ADON | 1.64 | 2.1 | 1.5–2.6 | ||||||
| 15-ADON | 18.0 | 1.3 | 0.62–6.0 | ||||||
| Wheat | 2018 | LC-MS/MS | 338 | DON | 90.8 | 2628 | 2.0–59278 | China | [140] |
| 3-ADON | 69.2 | 135 | 28.8–13409 | ||||||
| 15-ADON | 49.4 | 50.2 | 27.0–805 | ||||||
| NIV | 45.3 | 266 | 13.1–3044 | ||||||
| Wheat | 2016 | HPLC-UV | 92 | DON | 70.0 | 140 | 5.1–373 | Poland | [97] |
| NIV | 83.0 | 35.0 | 10.5–1265 | ||||||
| DON-3G | 27.0 | 41.9 | 15.8–138 | ||||||
| Wheat flour | 2019–2020 | UPLC-MS/MS | 27 | DON | 96.0 | 38.1 | 0.74–154 | Republic of Korea | [26] |
| DON-3G | 96.0 | 6.8 | 0.25–24.7 | ||||||
| NIV | 85.0 | 23.7 | 0.45–126 | ||||||
| 3-ADON | 11.0 | 6.34 | 2.3–10.3 | ||||||
| 15-ADON | 18.5 | 14.3 | 6.0–30.6 | ||||||
| FUS-X | 37.0 | 2.4 | 0.80–4.6 | ||||||
| Wheat flour | 2020 | UPLC-PDA | 50 | DON | 56.0 | 188 | <LOQ–389 | Egypt | [102] |
| NIV | 34.0 | 100 | <LOQ–179 | ||||||
| DON-3G | 24.0 | 84 | <LOQ–120 | ||||||
| Wheat | 2012–2014 | LC-MS/MS | 84 | DON | 100.0 | 1762 | <LOQ–9480 | Argentina | [98] |
| 3/15-ADON | 49.0 | 52 | <LOQ–190 | ||||||
| DON-3G | 93.0 | 198 | <LOQ–850 | ||||||
| Wheat | 2015–2018 | HPLC-PDA | 50 | DON | 26.0 | 433 | 58.0–1092 | Turkey | [123] |
| Wheat flour | 50 | 6.0 | 116 | 92.0–151 | |||||
| Wheat bread | 60 | ND | - | - | |||||
| Wheat (summer) | 2018–2019 | HPLC-UV | 232 | DON | 44.8 | 1051 | <LOQ–2145 | Pakistan | [99] |
| Wheat (winter) | 217 | 41.9 | 840 | <LOQ–2050 | |||||
| Wheat bran | 2013 | HPLC-UV | 37 | DON | 62.0 | 1308 | NA–6178 | Spain | [141] |
| Wheat | 2013 | LC-MS/MS | 57 | DON | 16.0 | 11.0 | 9.6–99.6 | Italy | [83] |
| 3-ADON | ND | - | - | ||||||
| 15-ADON | 2.0 | 0.64 | 10.8–29.1 | ||||||
| FUS-X | 14.0 | 18.4 | 12.5–102 | ||||||
| NIV | 11.0 | 8.9 | 12.0–106 | ||||||
| Wheat | 2015 | HPLC-MS/MS | 368 | DON | 100.0 | 17754 | 109.6–86,255 | China | [96] |
| DON-3G | 99.5 | 414 | 28.3–2957 | ||||||
| NIV | 87.8 | 250 | 0.60–2399 | ||||||
| 3-ADON | 80.0 | 39.6 | 0.60–284 | ||||||
| 15-ADON | 67.3 | 13.2 | 3.0–185 | ||||||
| FUS-X | 35.2 | 8.3 | 1.8–48.2 | ||||||
| Wheat | 2015 | LC-MS/MS | 30 | 3-ADON | 46.7 | 24.6 | NA–71.0 | Finland | [14] |
| DON | 96.7 | 866 | NA–5510 | ||||||
| DON-3G | 83.3 | 174 | NA–922 | ||||||
| NIV | 43.3 | 48.9 | NA–74.0 | ||||||
| Wheat | 2014 | HPLC-MS/MS | 47 | DON | 59.6 | 172 | 13.0–1230 | Italy | [142] |
| 3-ADON | 8.5 | 23.0 | 4.0–33.0 | ||||||
| 15-ADON | 32.0 | 45.0 | 12.0–105 | ||||||
| NIV | 6.3 | 183 | 67.0–290 | ||||||
| FUS-X | 6.3 | 8.0 | 5.0–14.0 | ||||||
| Wheat | 2014 | HPLC-MS/MS | 40 | DON | 22.5 | 172 | 9.0-550 | Syria | [142] |
| Wheat | 2012 | LC-MS/MS | 20 | DON | 50.0 | - | 22.8–113 | Malaysia | [119] |
| Wheat | 2010 | LC-MS/MS | 41 | DON | 100.0 | 1075 | 152–2550 | China | [93] |
| 2011 | 64 | 32.8 | 82.1 | 14.5–1580 | |||||
| 2012 | 75 | 96.0 | 307 | 16.3–41157 | |||||
| Wheat | 2014 | LC-MS/MS | 80 | DON | 5.0 | 653 | NA–1480 | Morocco | [143] |
| Wheat | 2009 | LC-MS/MS | 23 | DON | 100.0 | 1500 | 203–4130 | Austria, Germany, and Slovakia | [101] |
| DON-3G | 100.0 | 393 | 76.0–1070 | ||||||
| Wheat | 2017 | UPLC-MS/MS | 579 | DON | 100.0 | 166 | 12.2–6436 | China | [144] |
| 3-ADON | 4.2 | 1.2 | <LOQ–150 | ||||||
| 15-ADON | 0.52 | 0.20 | <LOQ–24.5 | ||||||
| Wheat | 2005–2007 | UPLC-PDA | 55 | DON | 34.5 | 190 | 57.0–423 | Serbia | [105] |
| Wheat | 2016 | LC-MS/MS | 150 | DON | 98.0 | - | 183–2150 | Brazil | [145] |
| Wheat biscuits | 2010 | LC-MS | 201 | NIV | 41.0 | 3.1 | 1.4–35.0 | Japan | [146] |
| DON | 98.0 | 23.0 | 0.90–791 | ||||||
| Wheat | 2007–2010 | LC-MS/MS | 686 | DON | 84.0 | 647 | NA–10,600 | Switzerland | [147] |
| NIV | 19.0 | 15.0 | NA–470 | ||||||
| Wheat | 2013 | HPLC-UV | 50 | DON | 40.0 | 910 | 7.0–4730 | India | [110] |
| Wheat | 2013 | ELISA/HPLC | 51 | DON | 65.0 | 223 | 115–278 | Croatia | [111] |
| Corn (winter) | 2018–2019 | HPLC-UV | 142 | DON | 61.2 | 1122 | <LOQ–2967 | Pakistan | [99] |
| Corn (summer) | 128 | 44.5 | 817 | <LOQ–2490 | |||||
| Maize | 2010 | ELISA/TLC and ELISA/HPLC | 40 | DON | 85.0 | 2150 | 15.0–17,900 | Croatia | [100] |
| Maize | 2008 | LC-MS/MS | 54 | DON | 100.0 | 753 | 238–3680 | Austria, Germany, and Slovakia | [101] |
| DON-3G | 100.0 | 141 | 25.0–763 | ||||||
| Corn | 2011 | GC–ECD | 25 | NIV | 52.0 | 25.4 | NA–129 | Republic of Korea | [84] |
| DON | 96.0 | 110 | NA–492 | ||||||
| FUS-X | 24.0 | 2.1 | NA–19.1 | ||||||
| 15-ADON | 80.0 | 17.3 | NA–98.0 | ||||||
| 3-ADON | 28.0 | 1.1 | NA–6.8 | ||||||
| Corn flour | 2019–2020 | UPLC-MS/MS | 25 | DON | 96.0 | 441 | 0.79–1223 | Republic of Korea | [26] |
| DON-3G | 96.0 | 99.7 | 0.14–419 | ||||||
| NIV | 80.0 | 57.5 | 1.9–234 | ||||||
| 3-ADON | 72.0 | 4.6 | 1.2–11.8 | ||||||
| 15-ADON | 60.0 | 135 | 38.6–298 | ||||||
| FUS-X | 60.0 | 13.0 | 1.2–28.7 | ||||||
| Corn flour | 2020 | UPLC-PDA | 45 | DON | 83.3 | 330 | <LOQ–853 | Egypt | [102] |
| NIV | 74.1 | 147 | <LOQ–462 | ||||||
| DON-3G | 40.7 | 121 | <LOQ–257 | ||||||
| Maize | 2017 | LC-MS/MS | 79 | DON | 8.0 | 311 | NA–807 | Egypt | [148] |
| DON-3G | 5.0 | - | NA–47.5 | ||||||
| NIV | 10.0 | - | NA–38.6 | ||||||
| Maize | 2007 | HPLC-MS | 180 | DON | 22.2 | 226 | 9.6–745 | Southwest Nigeria | [103] |
| 3-ADON | 17.2 | 17.0 | 0.70–72.4 | ||||||
| Maize | 2015 | HPLC-MS | 39 | DON | 70.7 | 0.30 | 0.10–0.70 | Northeast Nigeria | [104] |
| Maize | 2003–2005 | HPLC-MS | 312 | DON | 70.0 | 1104 | NA–10626 | Asia and Oceania | [149] |
| 244 | 80.0 | 1073 | NA–3970 | Europe and the Mediterranean region | |||||
| Maize | 2017 | UPLC-MS/MS | 606 | DON | 99.8 | 175 | <LOQ–4300 | China | [144] |
| 3-ADON | 13.5 | 5.0 | <LOQ–385 | ||||||
| 15-ADON | 76.4 | 115 | <LOQ–4811 | ||||||
| Maize | 2015–2018 | UPLC-MS/MS | 15 | DON | 13.3 | 322 | 313–331 | Turkey | [123] |
| Corn-based food | 2008 | GC-ECD | 175 | DON | 26.8 | 53.9 (median) | 26.1–132 | Spain | [150] |
| NIV | 4.0 | 60.2 (median) | 51.1–107 | ||||||
| Maize | 2013 | LC-MS/MS | 37 | DON | 100.0 | 171 | 43.0–435 | Cameroon | [151] |
| DON-3G | 100.0 | 16.0 | <LOQ–82.0 | ||||||
| NIV | 100.0 | 161 | 3.0–782 | ||||||
| FUS-X | 86.0 | 43.0 | <LOQ–112 | ||||||
| Corn | 2014 | LC-MS/MS | 10 | DON | 90.0 | 627 | 74.0–1382 | China | [19] |
| NIV | 40.0 | 25.0 | 19.0–53.0 | ||||||
| DON-3G | 80.0 | 14.0–121 | |||||||
| FUS-X | ND | - | - | ||||||
| 3-ADON | ND | - | - | ||||||
| 15-ADON | 60.0 | 40.0 | 15.0–79.0 | ||||||
| Corn | 2016 | HPLC-UV | 44 | DON | 66.0 | - | 5.8–9843 | China | [152] |
| Maize | 2005–2007 | UPLC-PDA, ELISA | 216 | DON | 32.4 | 223 | 27.0–2210 | Serbia | [105] |
| Maize | 2009–2011 | LC-MS/MS | 140 | DON | 21.4 | - | 3.0–428 | Italy | [106] |
| Maize | 2015 | LC-MS/MS | 60 | DON | 63.0 | 490 | 68.0–2196 | Tanzania | [107] |
| Maize | 2015 | HPLC-PDA | 30 | DON | 66.6 | 50.8 | <LOQ–90.0 | Poland | [108] |
| Maize | 2016 | LC-MS/MS | 29 | DON | 86.0 | 1872 | 225.0–2963 | Hungary | [109] |
| Maize | 2013 | HPLC-UV | 25 | DON | 24.0 | 450 | 10.0–1070 | India | [110] |
| Maize | 2013 | ELISA/HPLC | 63 | DON | 71.0 | 1565 | 215–2942 | Croatia | [111] |
| Oats | 2015 | LC-MS/MS | 31 | 3-ADON | 77.4 | 341 | NA–2720 | Finland | [14] |
| DON | 100.0 | 2690 | NA–23,800 | ||||||
| DON-3G | 87.1 | 806 | NA–6600 | ||||||
| NIV | 71.1 | 635 | NA–4940 | ||||||
| Oat kernel | 2015–2019 | LC-MS/MS | 100 | DON | 22.0 | 81.4 | 19.1–736 | Spain | [112] |
| 3-ADON | 3.0 | 29.9 | 9.2–42.6 | ||||||
| Oat bran | 2013 | LC-MS/MS | 30 | DON | 17.0 | 230 | NA–276 | Spain | [141] |
| Oat | 2013–2015 | LC-MS/MS | 325 | DON | 49.0 | 56.9 | NA–1328 | Switzerland | [113] |
| NIV | 64.3 | 108 | NA–1653 | ||||||
| Oat flakes | LC-MS/MS | 25 | DON | 60.0 | 44.0 | 16.8–244 | China | [114] | |
| Oat | 2016–2018 | UHPLC-HRMS | 168 | DON | 55.3 | 475 | NA–4143 | Canada | [115] |
| NIV | 92.0 | 210 | NA–795 | ||||||
| Oat | 2013 | LC-MS/MS | 7 | DON | 57.0 | 29.9 | 10.3–83.0 | Italy | [83] |
| 3-ADON | 14.2 | 0.74 | <LOQ–5.23 | ||||||
| 15-ADON | ND | - | - | ||||||
| FUS-X | 42.8 | 23.0 | 26.0–75.0 | ||||||
| NIV | 57.0 | 27.1 | 45.5–50.4 | ||||||
| Oat | 2017–2018 | HPLC-UV | 11 | DON | ND | - | - | Republic of Korea | [116] |
| DON-3G | ND | - | - | ||||||
| NIV | 9.1 | 23.5 | 23.5 | ||||||
| Oat | 2010–2011 | LC-MS/MS | 93 | DON | 95.0 | - | 99.0–5544 | Sweden | [117] |
| NIV | 91.5 | - | 18.0–1743 | ||||||
| Oat | 2006–2008 | LC-MS/MS | 303 | NIV | 73.0 | 57.0 | NA–741 | UK | [118] |
| FUS-X | 1.0 | <10.0 | NA–18.0 | ||||||
| DON | 32.0 | 28.0 | NA–1866 | ||||||
| Oat | 2013 | LC-MS/MS | 10 | DON | 30.0 | - | 22.7–100 | Malaysia | [119] |
| Oat | 2005–2010 | HPLC-UV | 52 | DON | 30.0 | 170 | NA–490 | Slovakia | [120] |
| Oat | 2005–2010 | HPLC-UV | 52 | DON | 4.0 | 90.0 | 90.0–94.0 | Russia | [153] |
| Oat | 2004–2009 | LC-MS/MS | 289 | DON | 90.0 | - | NA–30,000 | Norway | [154] |
| 3-ADON | 71.0 | - | NA–5100 | ||||||
| NIV | 1.0 | - | NA–310 | ||||||
| FUS-X | ND | - | - | ||||||
| Oat | 2013 | ELISA/HPLC | 33 | DON | 21.0 | 145 | 34.0–201 | Croatia | [111] |
| Barley | 2015 | LC-MS/MS | 34 | 3-ADON | 41.2 | - | NA–18.3 | Finland | [14] |
| DON | 82.4 | 234 | NA–802 | ||||||
| DON-3G | 73.5 | 148 | NA–594 | ||||||
| NIV | 73.5 | 96.6 | NA–262 | ||||||
| Barley | 2009 | HPLC-UV | 70 | DON | 54.0 | 9.4 | 3.7–36.8 | Republic of Korea | [121] |
| Barley | 2011 | GC-ECD | 39 | NIV | 59.0 | 16.6 | NA–101 | Republic of Korea | [84] |
| DON | 56.0 | 16.4 | NA–40.1 | ||||||
| FUS-X | 15.0 | 1.0 | NA–9.9 | ||||||
| 15-ADON | 31.0 | 1.1 | NA–7.1 | ||||||
| 3-ADON | 26.0 | 0.70 | NA–3.9 | ||||||
| Barley | 2020 | HPLC-UV | 15 | DON | 33.3 | 75.8 | 11.7–286 | Republic of Korea | [116] |
| DON-3G | 13.3 | 19.3 | 18.0–20.6 | ||||||
| NIV | 40.0 | 90.2 | 17.3–230 | ||||||
| Barley | 2013 | LC-MS/MS | 9 | DON | 11.0 | 3.9 | <LOQ–35.5 | Italy | [83] |
| 3-ADON | ND | - | - | ||||||
| 15-ADON | ND | - | - | ||||||
| FUS-X | 44.4 | 18.4 | 27.5–47.3 | ||||||
| NIV | 33.3 | 25.2 | 21.7–106 | ||||||
| Barley | 2012 | LC-MS/MS | 10 | DON | 50.0 | - | 27.9–72.5 | Malaysia | [119] |
| Barley beer | 2018 | LC-MS/MS | 100 | NIV | 56.0 | 2.4 µg/L | 0.50–7.6 µg/L | Poland | [122] |
| DON | 83.0 | 9.0 µg/L | 1.0–73.6 µg/L | ||||||
| DON-3G | 67.0 | 9.2 µg/L | 2.0–35.8 µg/L | ||||||
| Barley | 2016 | LC-MS/MS | 36 | DON | 48.0 | - | 240–429 | Hungary | [109] |
| Barley | 2013 | HPLC-UV | 25 | DON | 16.0 | 210 | 30.0–530 | India | [110] |
| Barley | 2018 | LC-MS/MS | 76 | DON | 94.0 | 5000 | 310–15500 | Brazil | [85] |
| Barley | 2013 | ELISA/HPLC | 34 | DON | 53.0 | 342 | 74.0–228 | Croatia | [111] |
| Barley | 2009 | HPLC-UV | 72 | DON | 56.9 | 400–2200 | 500–3600 | Tunisia | [155] |
| Barley | 2020 | HPLC-PDA | 15 | DON | 20.0 | 425 | 138–973 | Turkey | [123] |
| Barley | 1997–2000 | HPLC-PDA | 93 | DON | 9.0 | 2553 | 1470–4000 | Saudi Arabia | [156] |
| NIV | 1.0 | 3.1 | 3.1 | ||||||
| Rice | 2020 | HPLC-PDA | 20 | DON | 35.0 | 195 | 136–256 | Turkey | [123] |
| Rice | 2018 | LC-MS/MS | 180 | NIV | 28.0 | 13.8 | <LOQ–116 | Pakistan | [124] |
| DON | 8.0 | 6.9 | <LOQ–115 | ||||||
| White rice | 2018 | HPLC-UV | 241 | NIV | 21.0 | 33.5 | 12.6–2175 | Republic of Korea | [125] |
| DON | 5.0 | 4.0 | 7.1–372 | ||||||
| Brown rice | 2018 | HPLC-UV | 241 | NIV | 34.0 | 52.2 | 17.3–2534 | ||
| DON | 7.0 | 5.4 | 9.1–435 | ||||||
| Rice | 2011 | GC-ECD | 65 | NIV | 35.0 | 9.5 | NA–45.0 | Republic of Korea | [84] |
| DON | 15.0 | 3.7 | NA–31.7 | ||||||
| FUS-X | 15.0 | 1.4 | NA–15.0 | ||||||
| 15-ADON | 46.0 | 2.0 | NA–24.1 | ||||||
| 3-ADON | 12.0 | 0.60 | NA–10.0 | ||||||
| Glutinous rice | 2011 | GC-ECD | 11 | NIV | 64.0 | 11.9 | NA–23.1 | Republic of Korea | [84] |
| DON | 10.0 | 1.7 | NA–18.2 | ||||||
| FUS-X | 18.0 | 1.0 | NA–8.9 | ||||||
| 15-ADON | 45.0 | 1.3 | NA–4.2 | ||||||
| 3-ADON | ND | - | - | ||||||
| Brown rice | 2011 | GC-ECD | 48 | NIV | 60.0 | 14.5 | NA–45.4 | Republic of Korea | [84] |
| DON | 33.0 | 7.1 | NA–24.9 | ||||||
| FUS-X | 2.0 | 0.40 | NA–18.7 | ||||||
| 15-ADON | 25.0 | 0.5 | NA–3.1 | ||||||
| 3-ADON | 29.0 | 1.5 | NA–10.2 | ||||||
| Rice | 2014 | GC-MS/MS | 23 | DON | 13.0 | 5.0 | NA–5.5 | Spain | [126] |
| 3-ADON | ND | - | - | ||||||
| FUS-X | ND | - | - | ||||||
| NIV | ND | - | - | ||||||
| Rice | 2015 | LC-MS | 41 | DON | 70.7 | 0.30 | 0.10–0.7 | Nigeria | [104] |
| Rice | 2011 | HPLC-UV | 21 | DON | 23.8 | 18.9 | 11.2–112 | Nigeria | [157] |
| Rice | 2012 | LC-MS/MS | 50 | DON | 26.0 | - | 12.5–81.2 | Malaysia | [119] |
| Rice flour | 2014–2016 | LC-MS/MS | 93 | DON | 5.3 | 33.0 | NA–125 | Brazil | [127] |
| ADON | 73.3 | 3.8 | NA–17.0 | ||||||
| Rice-based cereals | 2008 | GC-MS | 29 | DON | 3.4 | 1.4 | - | Canada | [158] |
| Sorghum | 2017–2018 | UPLC-UV | DON | 33.3 | 81.7 | 30.0–159 | Republic of Korea | [159] | |
| NIV | 46.7 | 97.6 | 29.0–206 | ||||||
| Sorghum | 2017–2018 | HPLC-UV | 12 | DON | 100.0 | 119 | 18.9–712 | Republic of Korea | [116] |
| DON-3G | 41.7 | 18.8 | 10.4–43.4 | ||||||
| NIV | 91.7 | 45.3 | 4.6–146 | ||||||
| Sorghum | 2018 | LC-MS/MS | 110 | DON | 3.0 | 100 | NA–119 | Nigeria | [128] |
| 15-ADON | 2.0 | 39.0 | NA–44.0 | ||||||
| DON-3G | 23.0 | 24.0 | NA–63.0 | ||||||
| NIV | ND | - | - | ||||||
| FUS-X | ND | - | - | ||||||
| Sorghum | 2021 | LC-MS/MS | 20 | DON | ND | - | - | Nigeria | [129] |
| 3-ADON | ND | - | - | ||||||
| 15-ADON | ND | - | - | ||||||
| Sorghum | 2018 | LC-MS/MS | 1533 | DON | 0.46 | 63.3 | 40.0–112 | Sub-Saharan countries | [130] |
| Sorghum | 1999 | HPLC-UV | 33 | DON | 90.9 | 360 | 50.0–2340 | Ethiopia | [131] |
| NIV | 9.1 | 307 | 50.0–490 | ||||||
| Sorghum | 2019 | LC-MS/MS | 12 | NIV | 17.0 | 49.5 | 47.6–51.4 | Togo | [132] |
| DON | 17.0 | 25.9 | 19.0–32.8 | ||||||
| Sorghum | 2012 | LC-MS/MS | 4 | DON | ND | - | - | Tunisia | [133] |
| NIV | 100.0 | - | 418–667 | ||||||
| Rye | 2013 | LC-MS/MS | 11 | DON | 45.4 | 23.2 | 16.5–79.6 | Italy | [83] |
| 3-ADON | ND | - | - | ||||||
| 15-ADON | ND | - | - | ||||||
| FUS-X | 45.4 | 28.5 | 42.4–70.2 | ||||||
| NIV | 27.3 | 56.9 | 33.9–34.4 | ||||||
| Rye | 2010 | LC-MS/MS | 15 | DON | 100.0 | 270 | 87.0–500 | Canada | [135] |
| ADONs | ND | - | - | ||||||
| Rye | 2009 | LC-MS/MS | 61 | DON | 100.0 | 28 | NA–288 | Germany | [136] |
| 3-ADON | 59.0 | 0.39 | NA–5.0 | ||||||
| 15-ADON | 80.0 | 0.73 | NA–8.6 | ||||||
| NIV | 3.3 | 0.06 | NA–1.8 | ||||||
| FUS-X | 1.6 | 0.01 | NA–1.8 | ||||||
| Rye | 1998–2001 | GC-ECD | 69 | DON | 59.4 | 42.5 | NA–257 | Denmark | [138] |
| NIV | 13.0 | 11.5 | NA–48.0 | ||||||
| Rye | 2014 and 2015 | GC-ECD | 117 | DON | 75.0 | 1060 | <LOQ–10760 | USA | [139] |
| Conventional rye | 2009–2012 | LC-MS/MS | 18 | NIV | ND | - | - | Poland | [160] |
| DON | 83.0 | 29.5 | NA–120 | ||||||
| 3-ADON | ND | - | - | ||||||
| Organic rye | 2009–2012 | LC-MS/MS | 23 | NIV | 4.0 | - | <20 | Poland | [160] |
| DON | 26.0 | <15 | NA–41.6 | ||||||
| 3-ADON | ND | - | - |
| Method | Advantages | Limitations | Reference |
|---|---|---|---|
| Thin layer chromatography | Simple and rapid Low-cost separation technique Reliable quantification method when combined with densitometry | Outdated technique Poor precision and sensitivity Destructive sample preparation Quantitative only when combined with a densitometer Largely substituted by high-performance liquid chromatography for quantitative determination of trichothecenes Inherent need for sample preparation | [2,161,166] |
| High-performance liquid chromatography (HPLC) | Good sensitivity, selectivity, and repeatability Automated Short analysis time Official reference method for the validation and verification of immunochemical tests | Destructive sample preparation Expensive technique Requires dedicated operator Derivatization may be required | [175,243] |
| Liquid chromatography/mass spectrometry | High selectivity and repeatability Very low detection limits (LC-MS/MS) Fast acquisition features Compatibility with a broad range of sample preparation procedures Wide linear dynamic range Simultaneous determination of numerous mycotoxins Ability to generate structural information of analyte (HRMS) No derivatization required Minimum requirement for sample preparation (LC-MS/MS) | Destructive sample preparation Very expensive technique Requires dedicated operator and specialist expertise for data interpretation Sensitivity relies on the ionization method | [8,18,167] |
| Gas chromatography | Good separation ability and repeatability Very low detection limits (GC-MS/MS) Automated Simultaneous analyses of multiple mycotoxins | Expensive technique Requires dedicated operator Matrix interferences Requires derivatization for nonvolatile mycotoxins Carry-over effects from previous samples Narrow scope of analysis | [236,237,241] |
| Enzyme-linked immunosorbent assay (ELISA) | Inexpensive and specific assay Reduced analysis time Visual assessment Easy manipulation Semi-quantitative (screening) or quantitative analysis is possible No dedicated operator required Limited consumption of organic solvents | Easily affected by matrix interferences Affected by potential cross-reactivity with structurally related toxins One-time use only Inefficiency in detection at low concentrations Semiquantitative Confirmatory LC analysis is often required Possible false positives/negatives Narrow detection range | [175,246,274] |
| Lateral flow immunochromatographic assay | Rapid and straightforward (single-step) test No special equipment required Inexpensive onsite screening test No additional chemicals or laborious preparation processes required Portable Reliable quantification method when combined with other modern technology | Semiquantitative (visual assessment) Affected by potential cross-reactivity with structurally related toxins Requiring validation for additional matrices | [255,272,275] |
| Fluorescence polarization immunoassay | Mobility due to portable instrumentation Very sensitive, rapid and user-friendly Homogeneous method performed in the solution phase Faster detection with no additional clean-up and washing steps Convenient for monitoring large-scale samples | Possible cross-reactivity with structurally related toxins Limited validation with HPLC or ELISA Matrix interferences Limited to a single mycotoxin detection at a time | [257,258,272] |
| Biosensors | High transmission and low-cost operation High sensitivity and selectivity User-friendly operation Reduced analysis time Mobility due to portable instrumentation Ability to be recycled Self-contained, simple design | Extensive sample preparation is required to improve sensitivity Limited to a single mycotoxin detection at a time Possible cross-reactivity with structurally related toxins Variable repeatability and reproducibility (enhanced when using novel materials) | [260,261,272] |
| Near-infrared spectroscopy | Reduced analysis time Easy operation Non-destructive testing with minimal or no sample manipulation Quick classification of grains according to mycotoxin contamination | Reliable only when combined with appropriate mathematical tools such as principal component analysis Complicated interpretation of spectral data Knowledge of statistical methods is required Validation of the calibration model is required Expensive equipment Poor sensitivity (high limit of detection) Point-based scanning method which enables only a mean spectrum (average measurement) | [167,264,268] |
| Hyperspectral imaging | Reduced analysis time Easy operation Non-destructive testing with minimal or no sample manipulation Information about the spatial distribution of chemical constituents across the sample is provided (sample heterogeneity can be overcome) High spectral and spatial resolution Quick classification of grains according to mycotoxin contamination | Reliable only when combined with appropriate mathematical tools such as principal component analysis Complicated interpretation of spectral data Knowledge of statistical models is required Validation of the calibration model is required Expensive equipment Poor sensitivity (high limit of detection) | [265,266,267] |
| Electronic nose (EN) | Rapid, inexpensive, and user-friendly screening method to distinguish the microbiological quality of food samples. | Enhancing selectivity and sensitivity is required Reducing interferences (e.g., to humidity) is required Nonvolatile mycotoxins raise difficulties for EN-based detection. Compensation for drift effects is required Narrow scope of analysis and poor validation | [163,269,270] |
| Capillary electrophoresis | Rapid analysis Convenient for separating closely related toxins Limited consumption of organic solvents Good selectivity of analytes from interferences Good sensitivity | Destructive sample preparation Limited to lab use due to cumbersome instrumentation Extensive sample preparation is required to improve sensitivity | [163,272] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gab-Allah, M.A.; Choi, K.; Kim, B. Type B Trichothecenes in Cereal Grains and Their Products: Recent Advances on Occurrence, Toxicology, Analysis and Post-Harvest Decontamination Strategies. Toxins 2023, 15, 85. https://doi.org/10.3390/toxins15020085
Gab-Allah MA, Choi K, Kim B. Type B Trichothecenes in Cereal Grains and Their Products: Recent Advances on Occurrence, Toxicology, Analysis and Post-Harvest Decontamination Strategies. Toxins. 2023; 15(2):85. https://doi.org/10.3390/toxins15020085
Chicago/Turabian StyleGab-Allah, Mohamed A., Kihwan Choi, and Byungjoo Kim. 2023. "Type B Trichothecenes in Cereal Grains and Their Products: Recent Advances on Occurrence, Toxicology, Analysis and Post-Harvest Decontamination Strategies" Toxins 15, no. 2: 85. https://doi.org/10.3390/toxins15020085
APA StyleGab-Allah, M. A., Choi, K., & Kim, B. (2023). Type B Trichothecenes in Cereal Grains and Their Products: Recent Advances on Occurrence, Toxicology, Analysis and Post-Harvest Decontamination Strategies. Toxins, 15(2), 85. https://doi.org/10.3390/toxins15020085