1H NMR Profiling of the Venom from Hylesia continua: Implications of Small Molecules for Lepidopterism

 ,
, 

Abstract

1. Introduction
2. Results
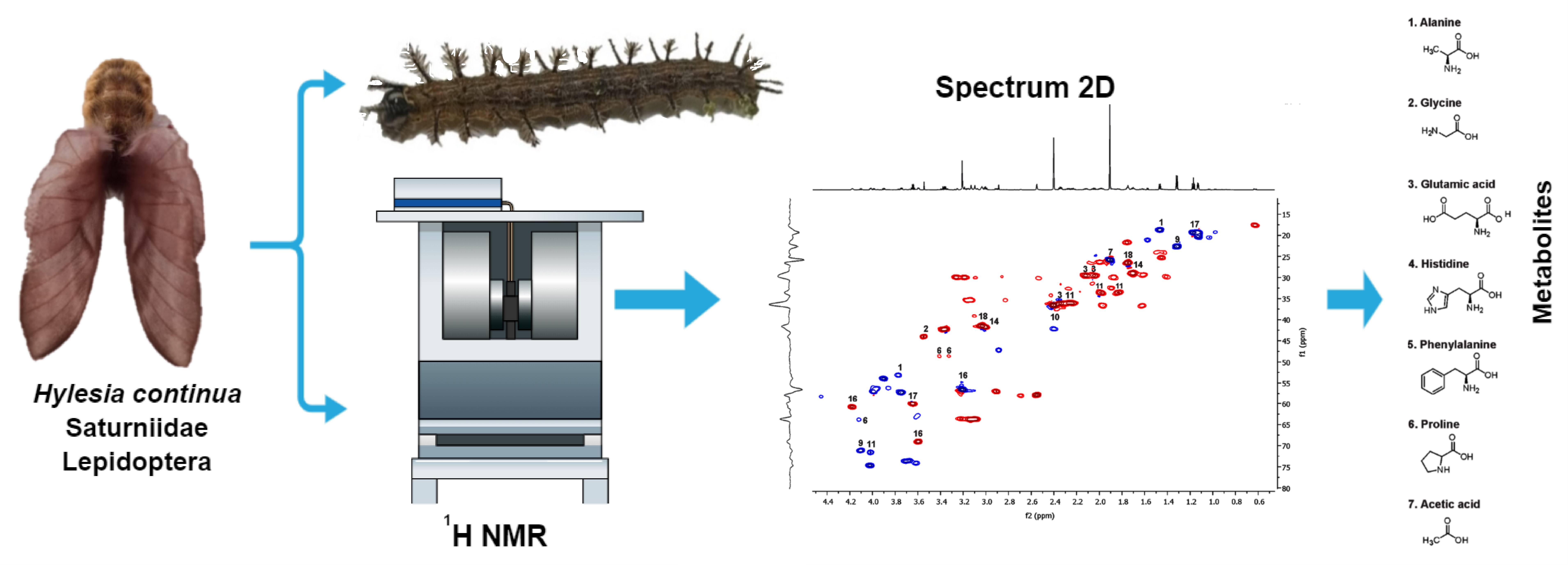
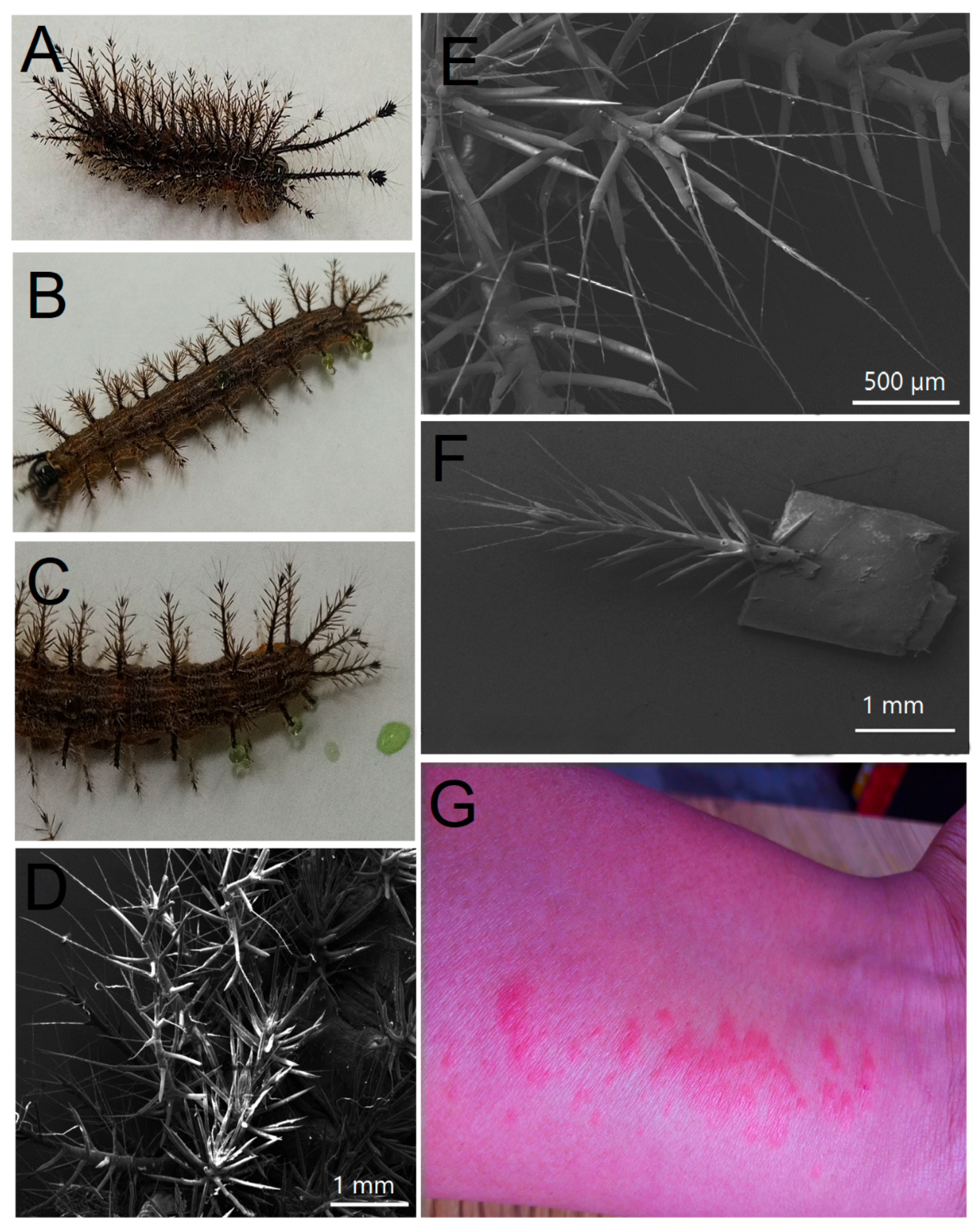
2.1. Morphological and Molecular Features of Hylesia continua
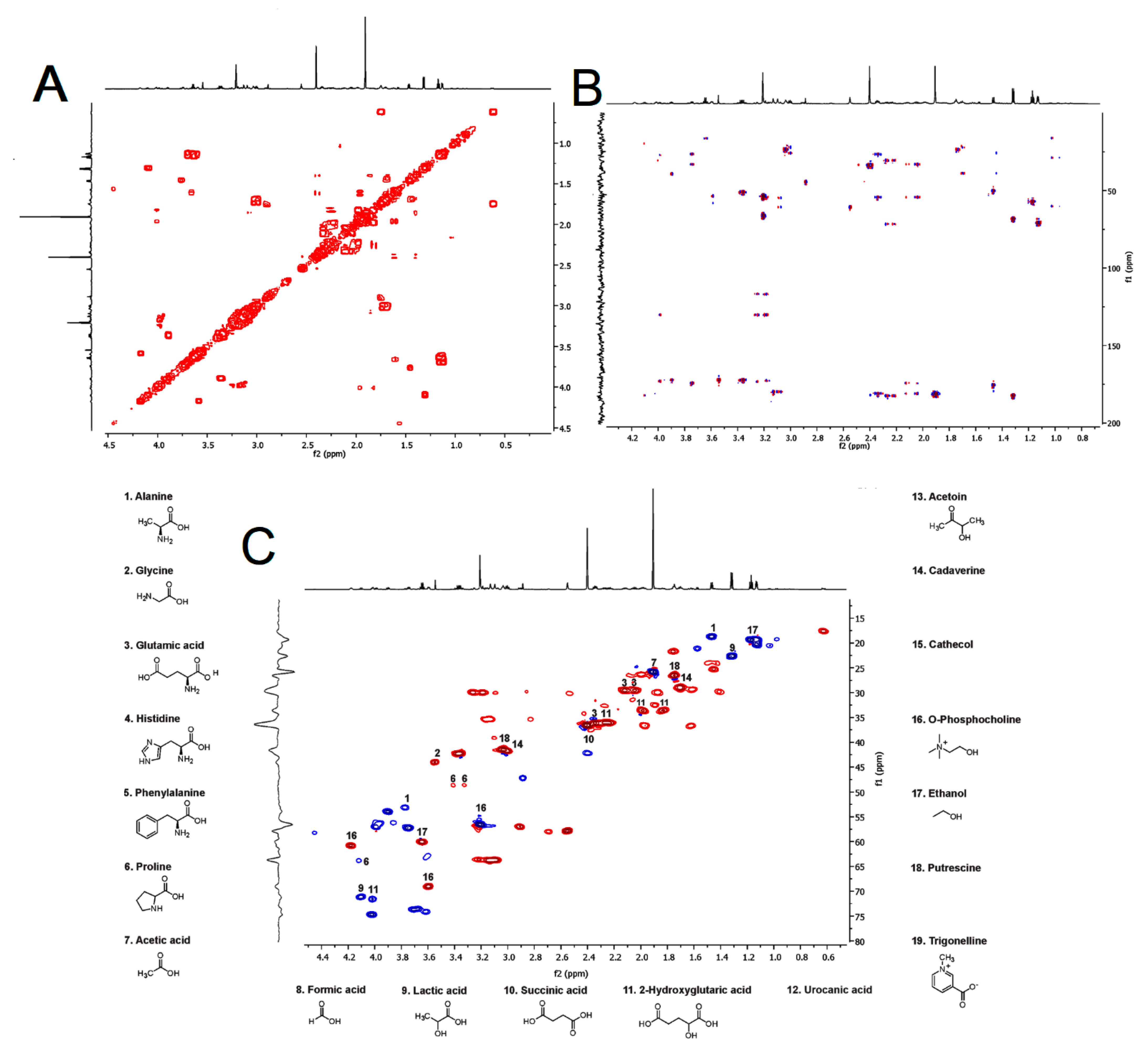
2.2. 1H NMR Profiling of the Venom from Hylesia continua
3. Discussion
4. Conclusions
5. Materials and Methods
5.1. Collection and Identification of Hylesia continua
5.2. Venom Extraction and Sample Preparation
5.3. NMR Analysis
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Hossler, E.W. Caterpillars and moths. Dermatol. Ther. 2009, 22, 353–356. [Google Scholar] [CrossRef] [PubMed]
- Lee, D.; Pitetti, R.D.; Casselbrant, M.L. Oropharyngeal manifestations of lepidopterism. Arch. Otolaryngol. Head Neck Surg. 1999, 125, 50–52. [Google Scholar] [CrossRef] [PubMed]
- Konstat-Korzenny, E.; Yudovich, A.; Morgenstern-Kaplan, D. Lepidopterism: Case report and review of the literature. Cureus 2020, 12, e6567. [Google Scholar] [CrossRef] [PubMed]
- Gómez, J.P.C. Lepidopterismo y Euricismo en Colombia. Rev. Biosalud 2014, 13, 59–83. [Google Scholar]
- Seldeslachts, A.; Peigneur, S.; Tytgat, J. Caterpillar venom: A health hazard of the 21st century. Biomedicines 2020, 8, 143. [Google Scholar] [CrossRef]
- Alvarez-Flores, M.P.; Gomes, R.N.; Trevisan-Silva, D.; Oliveira, D.S.; Batista, I.D.F.C.; Buri, M.V.; Alvarez, A.M.; DeOcesano-Pereira, C.; de Souza, M.M.; Chudzinski-Tavassi, A.M. Lonomia obliqua envenoming and innovative research. Toxins 2021, 13, 832. [Google Scholar] [CrossRef]
- Eyer-Silva, W.A.; Silva, G.A.R.D.; Neves-Motta, R. Two cases of lepidopterism caused by indoor exposure to moths. Rev. Soc. Bras. Med. Trop. 2016, 49, 393. [Google Scholar] [CrossRef]
- Rodríguez-Morales, A.J.; Arria, M.; Rojas-Mirabal, J.; Borges, E.; Benitez, J.A.; Herrera, M.; Villalobos, C.; Maldonado, A.; Rubio, N.; Franco-Paredes, C. Lepidopterism due to exposure to the moth Hylesia metabus in northeastern Venezuela. Am. J. Trop. Med. Hyg. 2005, 73, 991–993. [Google Scholar] [CrossRef]
- Paniz-Mondolfi, A.E.; Pérez-Alvarez, A.M.; Lundberg, U.; Fornés, L.; Reyes-Jaimes, O.; Hernández-Pérez, M.; Hossler, E. Cutaneous lepidopterism: Dermatitis from contact with moths of Hylesia metabus (Cramer 1775) (Lepidoptera: Saturniidae), the causative agent of caripito itch. Int. J. Dermatol. 2011, 50, 535–541. [Google Scholar] [CrossRef]
- Jourdain, F.; Girod, R.; Vassal, J.M.; Chandre, F.; Lagneau, C.; Fouque, F.; Guiral, D.; Raude, J.; Robert, V. The moth Hylesia metabus and French Guiana lepidopterism: Centenary of a public health concern. Parasite 2012, 19, 117–128. [Google Scholar] [CrossRef]
- Herrera-Chaumont, C.; Sojo-Milano, M.; Pérez-Ybarra, L. Conocimientos y prácticas sobre lepidopterismo por Hylesia metabus (Cramer, 1775) (Lepidoptera: Saturniidae) en la parroquia Yaguaraparo, Estado Sucre, noreste de Venezuela. Rev. Biomed. 2016, 27, 11–23. [Google Scholar] [CrossRef]
- Iserhard, C.A.; Kaminski, L.A.; Marchiori, M.A.; Teixeira, E.C.; Romanowski, H.P. Occurrence of Lepidopterism caused by the moth Hylesia nigricans (Berg) (Lepidoptera: Saturniidae) in Rio Grande do Sul State, Brazil. Neotrop. Entomol. 2007, 36, 612–614. [Google Scholar] [CrossRef]
- Cabrerizo, S.; Spera, M.; de Roodt, A. Accidents due to Lepidoptera: Hylesia nigricans (Berg, 1875) or “mariposa negra”. Arch. Argent. Pediatr. 2014, 112, 179–182. [Google Scholar]
- Alvarez Garcia, D.M.; Díaz Pérez, J.A.; Amarillo-Suárez, A. Hylesia continua (Walker, 1865) (Lepidoptera: Saturniidae) in a locality of northern Colombia: Pupae dimorphism and lepidopterism. Acta Zool. Mex. 2015, 31, 327–330. [Google Scholar] [CrossRef]
- Pacheco-Hernández, Y.; Villa-Ruano, N.; Rubio-Rosas, E.; Vásquez-Lara, I. Chemical profiling of the urticating trichomes from Cnidoscolus multilobus (“mala mujer”) and their antimicrobial activity. Polibotanica 2020, 50, 137–146. [Google Scholar] [CrossRef]
- Lundberg, U.; Salazar, V.; León, E.; González, L.J.; Hernández, Y.T. Toxinología de la polilla Hylesia metabus. In Estudio Mutidiscuplinario de la Palometa Peluda Hylesia Metabus, 1st ed.; Hernández, E.J.V., Osborn, F., Conde, J.L., Eds.; Ediciones IVIC: Caracas, Venezuela, 2012; Volume 1, pp. 57–68. [Google Scholar]
- Salazar, V.; Lundberg, U.; Gamez, R.; Méndez, R. Efectos de la toxina de las setas urticantes de la polilla Hylesia metabus (Cramer [1775]) (Lepidoptera, Saturniidae), en modelos experimentales. Estudio histopatológico. In Estudio Mutidiscuplinario de la Palometa Peluda Hylesia Metabus, 1st ed.; Hernández, E.J.V., Osborn, F., Conde, J.L., Eds.; Ediciones IVIC: Caracas, Venezuela, 2012; Volume 1, pp. 69–106. [Google Scholar]
- Casafús, M.G.; Gritti, M.A.; González, K.Y.; Sánchez, M.N.; Sciani, J.M.; Martínez, M.M.; Teibler, G.P.; Peichoto, M.E. Unraveling the distinctive venomous features of the saturniid Hylesia sp.: An integrative approach of a public health concern in Argentina. Acta Trop. 2022, 231, 106428. [Google Scholar] [CrossRef]
- Villa-Ruano, N.; Rosas-Bautista, A.; Rico-Arzate, E.; Cruz-Narvaez, Y.; Zepeda-Vallejo, L.G.; Lalaleo, L.; Hidalgo-Martínez, D.; Becerra-Martínez, E. Study of nutritional quality of pomegranate (Punica granatum L.) juice using 1H NMR-based metabolomic approach: A comparison between conventionally and organically grown fruits. LWT-Food Sci. Technol. 2020, 134, 110222. [Google Scholar] [CrossRef]
- Hylesia continua (Saturniidae). Available online: https://www.acguanacaste.ac.cr/paginas-de-especies/insectos/102-saturniidae/368-i-hylesia-continua-i-saturniidae (accessed on 25 November 2022).
- Villa, R.T. Personal Communication; Mexican Institute of Social Security-Rural Clinic of Yaonáhuac: Puebla, México, 2022. [Google Scholar]
- Rabl, D.; Gottsberger, B.; Brehm, G.; Hofhansl, F.; Fiedler, K. Moth assemblages in Costa Rica rain forest mirror small-scale topographic heterogeneity. Biotropica 2019, 52, 288–301. [Google Scholar] [CrossRef]
- Schroeder, F.C.; Taggi, A.E.; Gronquist, M.; Malik, R.U.; Grant, J.B.; Eisner, T.; Meinwald, J. NMR-spectroscopic screening of spider venom reveals sulfated nucleosides as major components for the brown recluse and related species. Proc. Natl. Acad. Sci. USA 2008, 105, 14283–14287. [Google Scholar] [CrossRef]
- Lee, S.; Kalcic, F.; Duarte, I.F.; Titera, D.; Kamler, M.; Mrna, P.; Hyrsl, P.; Danihlik, J.; Dobes, P.; Kunc, M.; et al. 1H NMR profiling of honey bee bodies revealed metabolic differences between summer and winter bees. Insects 2022, 13, 193. [Google Scholar] [CrossRef]
- Brütsch, T.; Jaffuel, G.; Vallat, A.; Turlings, T.C.J.; Chapuisat, M. Wood ants produce a potent antimicrobial agent by applying formic acid on tree-collected resin. Ecol. Evol. 2017, 7, 2249–2254. [Google Scholar] [CrossRef] [PubMed]
- Nishio, S.; Yamaguchi, M.; Yamada, N.; Uenishi, N.; Kageyama, Y.; Umetsu, T. Effect of beraprost sodium (TRK-100) on acetic acid-induced skin ulcer. Res. Commun. Chem. Pathol. Pharmacol. 1989, 64, 381–393. [Google Scholar] [PubMed]
- Gawade, S.P. Acetic acid induced painful endogenous infliction in writhing test on mice. J. Pharmacol. Pharmacother. 2012, 3, 348. [Google Scholar] [CrossRef] [PubMed]
- Torres, A.F.C.; Quinet, Y.P.; Havt, A.; Rádis-Baptista, G.; Martins, A.M.C. Molecular pharmacology and toxinology of venom from ants. In An Integrated View of the Molecular Recognition and Toxinology- From Analytical Procedures to Biomedical Applications, 1st ed.; Baptista, G.R., Ed.; IntechOpen: London, UK, 2013; Volume 1, pp. 1–16. [Google Scholar] [CrossRef]
- Manosalva, C.; Quiroga, J.; Hidalgo, A.I.; Alarcon, P.; Anseoleaga, N.; Hidalgo, M.A.; Burgos, R.A. Role of lactate in inflammatory processes: Friend or foe. Front. Inmunol. 2022, 12, 808799. [Google Scholar] [CrossRef] [PubMed]
- Williams, N.C.; Ryan, D.G.; Costa, A.S.H.; Mills, E.L.; Jedrychowski, M.P.; Cloonan, S.M.; Frezza, C.; O’Neill, L.A. Signaling metabolite L-2-hydroxyglutarate activates the transcription factor HIF-1α in lipopolysaccharide-activated macrophages. J. Biol. Chem. 2022, 298, 101501. [Google Scholar] [CrossRef]
- Lachenmeier, D.W. Safety evaluation of topical applications of ethanol on the skin and inside the oral cavity. J. Occup. Med. Toxicol. 2008, 3, 26. [Google Scholar] [CrossRef]
- In, H.S.; Kim, D.W.; Park, Y.M.; Kim, B. Experimental intraperitoneal injection of alcohol in rats: Peritoneal findings and histopathology. Toxicol. Rep. 2014, 1, 31–35. [Google Scholar] [CrossRef]
- Wozniak, K.M.; Rojas, C.; Wu, Y.; Slusher, B.S. The role of glutamate signaling in pain processes and its regulation by GCP II inhibition. Curr. Med. Chem. 2012, 19, 1323–1334. [Google Scholar] [CrossRef]
- Lewerenz, J.; Maher, P. Chronic glutamate toxicity in neurodegenerative diseases—What is the evidence? Front. Neurosci. 2015, 9, 469. [Google Scholar] [CrossRef]
- Lovell, T.M.; Woods, R.J.; Butlin, D.J.; Brayley, K.J.; Manyonda, I.T.; Jarvis, J.; Howell, S.; Lowry, P.J. Identification of a novel mammalian post-translational modification, phosphocholine, on placental secretory polypeptides. J. Mol. Endocr. 2007, 39, 189–198. [Google Scholar] [CrossRef]
- Wu, X.; Zheng, Y.; Ma, J.; Yin, J.; Chen, S. The effects of dietary glycine on the acetic acid-induced mouse model of colitis. Mediat. Inflamm. 2020, 2020, 5867627. [Google Scholar] [CrossRef]
- Jin, S.J.; Moon, H.W.; Kim, J.S.; Kim, H.Y. Effects of 4 weeks of beta-alanine intake on inflammatory cytokines after 10 km long distance running exercise. Exerc. Sci. 2022, 31, 188–196. [Google Scholar] [CrossRef]
- Niu, Y.C.; Feng, R.N.; Hou, Y.; Li, K.; Kang, Z.; Wang, J.; Sun, C.H.; Li, Y. Histidine and arginine are associated with inflammation and oxidative stress in obese women. Br. J. Nut. 2012, 108, 57–61. [Google Scholar] [CrossRef]
- Delwing, D.; Bavaresco, C.S.; Wannmacher, C.M.; Wajner, M.; Dutra-Filho, C.S.; Wyse, A.T. Proline induces oxidative stress in cerebral cortex of rats. Int. J. Dev. Neurosci. 2003, 21, 105–110. [Google Scholar] [CrossRef]
- del Rio, B.; Redruello, B.; Linares, D.M.; Ladero, V.; Ruas-Madiedo, P.; Fernandez, M.; Martin, M.C.; Alvarez, M. The biogenic amines putrescine and cadaverine show in vitro cytotoxicity at concentrations that can be found in foods. Sci. Rep. 2019, 9, 120. [Google Scholar] [CrossRef]
- Fan, G.C.; Rong, X.Z.; Wang, X.M.; Liu, S.R.; Zhou, Y.P.; Li, Q.H. Influence of exogenous putrescine and cadaverine on pro-inflammatory factors in the peripheral blood of rabbits. Zhonghua Shao Shang Za Zhi 2012, 28, 451–454. [Google Scholar]
- Cesselin, B.; Henry, C.; Gruss, A.; Gloux, K.; Gaudu, P. Mechanisms of acetoin toxicity and adaptive responses in an acetoin-producing species. Lact. Lactis. Appl Environ. Microbiol. 2021, 87, e01079-21. [Google Scholar] [CrossRef]
- Gibbs, N.K.; Norval, M. Urocanic acid in the skin: A mixed blessing? J. Investig. Dermatol. 2011, 131, 14–17. [Google Scholar] [CrossRef]
- De La Cruz, J.P.; Ruiz-Moreno, M.I.; Guerrero, A.; López-Villodres, J.A.; Reyes, J.J.; Espartero, J.L.; Labajos, M.T.; González-Correa, J.A. Role of the catechol group in the antioxidant and neuroprotective effects of virgin olive oil components in rat brain. J. Nutr. Biochem. 2015, 26, 549–555. [Google Scholar] [CrossRef]
- Chowdhury, A.A.; Gawali, N.B.; Munshi, R.; Juvekar, A.R. Trigonelline insulates against oxidative stress, proinflammatory cytokines and restores BDNF levels in lipopolysaccharide induced cognitive impairment in adult mice. Metab. Brain Dis. 2018, 33, 681–691. [Google Scholar] [CrossRef]
- Sotler, R.; Poljšak, B.; Dahmane, R.; Jukić, T.; Jukić, D.P.; Rotim, C.; Trebše, P.; Starc, A. Prooxidant activities of antioxidants and their impact on health. Acta Clin. Croat. 2019, 58, 726–736. [Google Scholar] [CrossRef]
- Scoble, M. The Lepidoptera: Form, Function and Diversity Natural History Museum Publications, 1st ed.; Oxford University Press: New York City, NY, USA, 1992; pp. 203–404. [Google Scholar]
- Folmer, O.; Black, M.; Hoeh, W.; Lutz, R.; Vrijenhoek, R. DNA primers for amplification of mitochondrial cytochrome C oxidase subunit I from diverse metazoan invertebrates. Mol. Mar. Biol. Biotechnol. 1994, 3, 294–299. [Google Scholar]
- Villa-Ruano, N.; Pérez-Hernández, N.; Zepeda-Vallejo, L.G.; Quiroz-Acosta, T.; Mendieta-Moctezuma, A.; García-Nava, M.L.; Becerra-Martínez, E. 1H NMR-based metabolomics profiling of citrus juices produced in Veracruz, México. Chem. Biodivers. 2019, 16, e1800479. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
{kind=link}
| Metabolite | Experimental Data | Confirmation | HMDB Data | Assignment | |||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| δ 1H (ppm) | Multiplicity | J (Hz) | δ 13C (ppm) | δ 1H (ppm) | Multiplicity | J (Hz) | δ 13C (ppm) | ||||
| Amino acids | |||||||||||
| 1 | Alanine | 1.47 3.76 - | d q - | 7.2 7.2 - | 18.67 53.06 176.38 | 1H, COSY, HSQC, HMBC | 1.46 3.77 - | d q - | 7.1 7.20 - | 19.03 53.55 177.35 | CH3-6 CH2-4 C=O |
| 2 | Glycine | 3.55 - | s - | - - | 44.03 172 64 | 1H, HSQC, HMBC | 3.54 - | s - | - - | 44.30 173.59 | CH2-4 C=O |
| 3 | Glutamic acid | 2.04 2.12 2.35 - - | m m m - - | - - - - - | 29.69 29.69 36.07 174.87 181.05 | 1H, COSY HSQC | - 2.08 2.34 - - | m m m - - | - - - - - | 29.82 29.82 36.35 175.82 182.43 | CH2-3 CH2-3 CH2-2 C=O C=O |
| 4 | Histidine | 7.17 8.11 | d d | - - | 119.73 138.19 | 1H, HSQC | 7.09 7.90 | d d | 0.6 1.1 | 119.99 138.36 | CH-5 CH-2 |
| 5 | Phenylalanine | 7.32 7.36 7.41 | d m m | 7.5 - - | 131.32 129.36 130.92 | 1H, HSQC | 7.32 7.36 7.42 | d m m | 6.9 - - | 132.11 130.42 131.81 | CH-6, 2 CH-4 CH-3, 5 |
| 6 | Proline | 3,32 3.41 4.12 | m m m | - - - | 48.67 48.67 63.57 | 1H, HSQC | 3.33 3.41 4.12 | dt dt dd | 14.0, 7.1 11.6, 7.0 8.8, 6.4 | 48.95 48.95 64.03 | CH2-2 CH2-2 CH-5 |
| Organic acids | |||||||||||
| 7 | Acetic acid | 1.91 - | s - | - - | 25.86 180.81 | 1H, HSQC, HMBC | 1.91 - | s - | - - | 26.08 176.55 | CH3-4 C=O |
| 8 | Formic acid | 8.44 | s | - | - | 1H | 8.44 | s | - | 172.41 | CH-2 |
| 9 | Lactic acid | 1.32 4.10 - | d q - | 6.9 7.0 - | 22.71 71.15 183.46 | 1H, COSY, HSQC, HMBC | 1.32 4.10 | d q - | 7.0 7.0 - | 22.90 71.37 185.08 | CH3-3 CH2-2 C=O |
| 10 | Succinic acid | 2.40 | s | - | 36.61 | 1H, HSQC | 2.39 | s | - | 36.82 | CH2-4, 5 |
| 11 | 2-Hydroxyglutaric acid | 1.84 1.99 2.29 4.02 - | m m m m - | - - - - - | 33.84 33,84 36.24 71.39 182.57 | 1H, HSQC, HMBC | 1.83 1.98 2.25 4.01 - | m m m m - | - - - - - | 33.66 33.74 36.02 74.80 183.83 | CH2-2 CH2-2 CH2-3 CH-1 C=O |
| 12 | Urocanic acid | 6.41 7.28 7.43 | d d s | 16.0 16.0 - | - - - | 1H, COSY, HSQC | 6.38 7.29 7.38 | d d m | 16.0 16.1 - | 124.55 133.42 123.94 | CH-4 CH-3 CH-2 |
| Other compounds | |||||||||||
| 13 | Acetoin | 1.36 2.22 | d s | 7.2 - | - - | 1H | 1.37 2.21 | d s | 7.2 - | 20.91 27.55 | CH3-3 CH3-6 |
| 14 | Cadaverine | 1.41 1.70 3.00 | m m m | - - - | - 29.05 41.36 | 1H, COSY, HSQC | 1.34 1.49 2.68 | m m m | - - - | 25.90 32.86 42.82 | CH2-3 CH2-2 CH2-1 |
| 15 | Catechol | 6.85 6.93 | m m | - - | 123.88 119.15 | 1H, COSY, HSQC | 6.88 6.96 | m m | - - | 123.81 119.12 | CH-2 CH-1 |
| 16 | O-Phosphocholine | 3.21 3.60 4.18 | s m m | - - - | 56.61 68.85 60.90 | 1H, COSY, HSQC | 3.19 3.57 4.15 | s m m | - - - | 56.52 68.90 60.60 | CH3-5, 6, 7 CH2-2 CH2-3 |
| 17 | Ethanol | 1.17 3.64 | t q | 7.1 7.1 | 19.46 59.93 | 1H, COSY, HSQC | 1.17 3.65 | t q | 7.1 7.1 | 19.59 60.30 | CH3-3 CH2-2 |
| 18 | Putrescine | 1.75 3,03 | m m | - - | 26.53 40.59 | 1H, COSY, HSQC | 1.75 | m m | - - | 26.72 41.73 | CH2-2 CH2-1 |
| 19 | Trigonelline | 4.43 8.06 8.82 8.83 9.11 | s m m m s | - - - - - | - - - - - | 1H | 4.43 8.07 8.78 8.87 9.16 | s m m m s | - - - - - | 51.06 130.09 148.71 147.51 148.38 | CH3-9 CH-4 CH-5 CH-3 CH-1 |
| Metabolite | Concentration (mM) | |
|---|---|---|
| 1 | Alanine | 2.7708 |
| 2 | Glycine | 3.2829 |
| 3 | Glutamic acid | 5.4098 |
| 4 | Histidine | 1.3714 |
| 5 | Phenylalanine | 0.0755 |
| 6 | Proline | 1.2543 |
| 7 | Acetic acid | 28.1704 |
| 8 | Formic acid | 0.4487 |
| 9 | Lactic acid | 6.4347 |
| 10 | Succinic acid | 9.7295 |
| 11 | 2-Hydroxyglutaric acid | 6.4660 |
| 12 | Urocanic acid | 0.1029 |
| 13 | Acetoin | 0.2760 |
| 14 | Cadaverine | 1.3077 |
| 15 | Catechol | 0.1029 |
| 16 | O-Phosphocholine | 3.0128 |
| 17 | Ethanol | 9.1453 |
| 18 | Putrescine | 1.5733 |
| 19 | Trigonelline | 0.0259 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Villa-Ruano, N.; Becerra-Martínez, E.; Cunill-Flores, J.M.; Torres-Castillo, J.A.; Horta-Valerdi, G.M.; Pacheco-Hernández, Y. 1H NMR Profiling of the Venom from Hylesia continua: Implications of Small Molecules for Lepidopterism. Toxins 2023, 15, 101. https://doi.org/10.3390/toxins15020101
Villa-Ruano N, Becerra-Martínez E, Cunill-Flores JM, Torres-Castillo JA, Horta-Valerdi GM, Pacheco-Hernández Y. 1H NMR Profiling of the Venom from Hylesia continua: Implications of Small Molecules for Lepidopterism. Toxins. 2023; 15(2):101. https://doi.org/10.3390/toxins15020101
Chicago/Turabian StyleVilla-Ruano, Nemesio, Elvia Becerra-Martínez, José María Cunill-Flores, Jorge Ariel Torres-Castillo, Guillermo M. Horta-Valerdi, and Yesenia Pacheco-Hernández. 2023. "1H NMR Profiling of the Venom from Hylesia continua: Implications of Small Molecules for Lepidopterism" Toxins 15, no. 2: 101. https://doi.org/10.3390/toxins15020101
APA StyleVilla-Ruano, N., Becerra-Martínez, E., Cunill-Flores, J. M., Torres-Castillo, J. A., Horta-Valerdi, G. M., & Pacheco-Hernández, Y. (2023). 1H NMR Profiling of the Venom from Hylesia continua: Implications of Small Molecules for Lepidopterism. Toxins, 15(2), 101. https://doi.org/10.3390/toxins15020101