Yessotoxins in Mollusks of the Galician Coast from 2014 to 2022: Variability, Biotransformation, and Resistance to Alkaline Hydrolysis



Abstract
:1. Introduction
2. Results
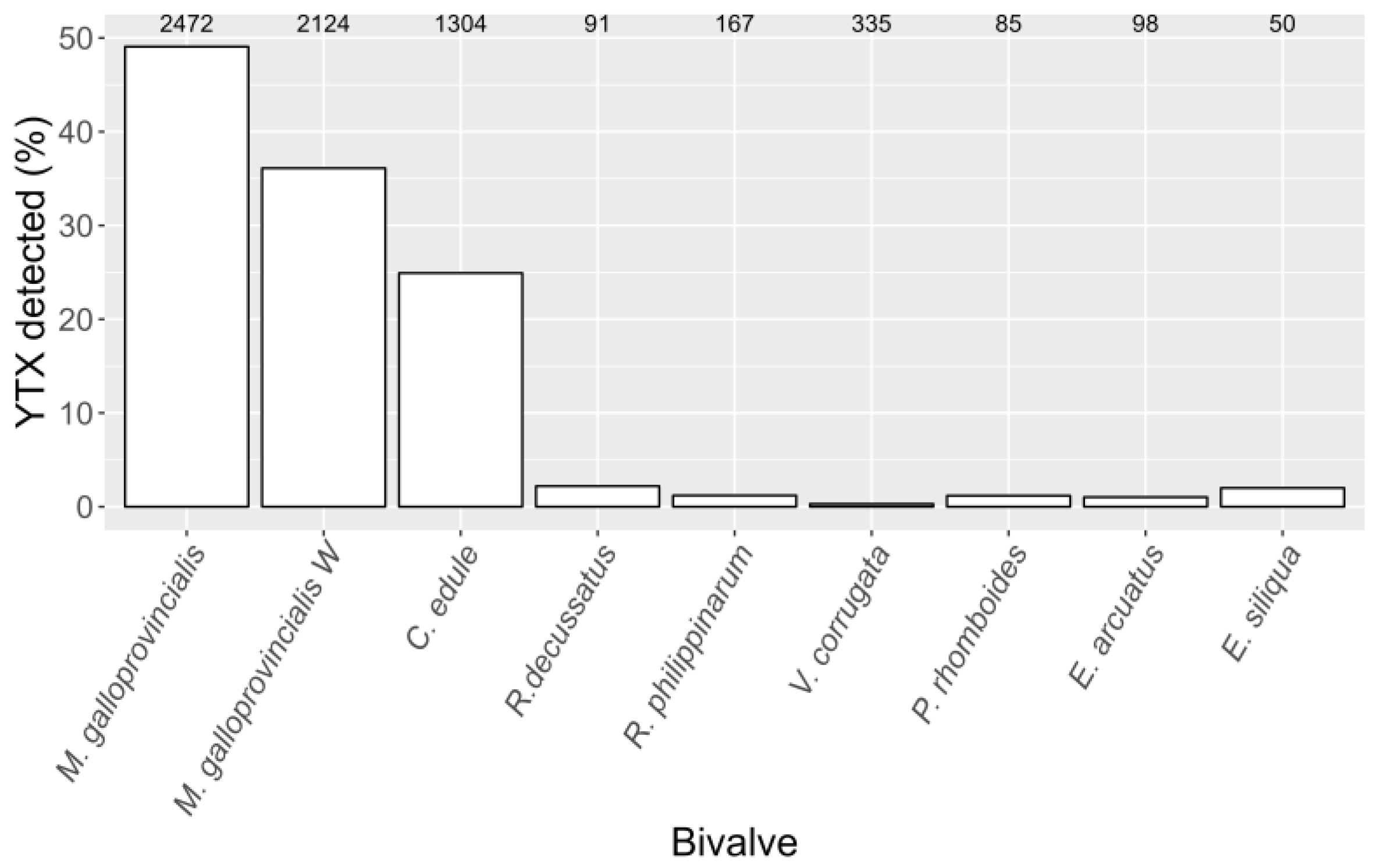
2.1. General
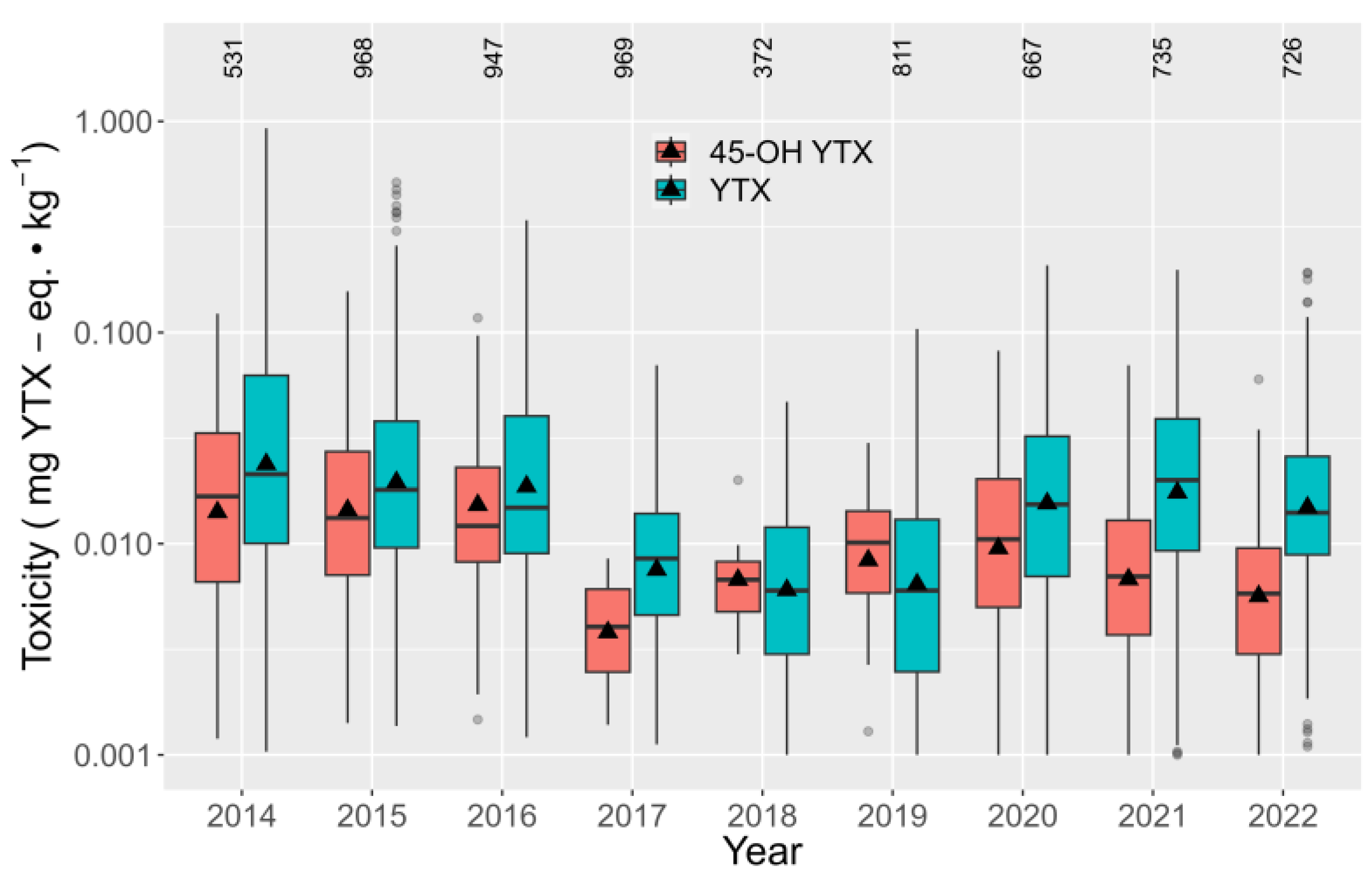
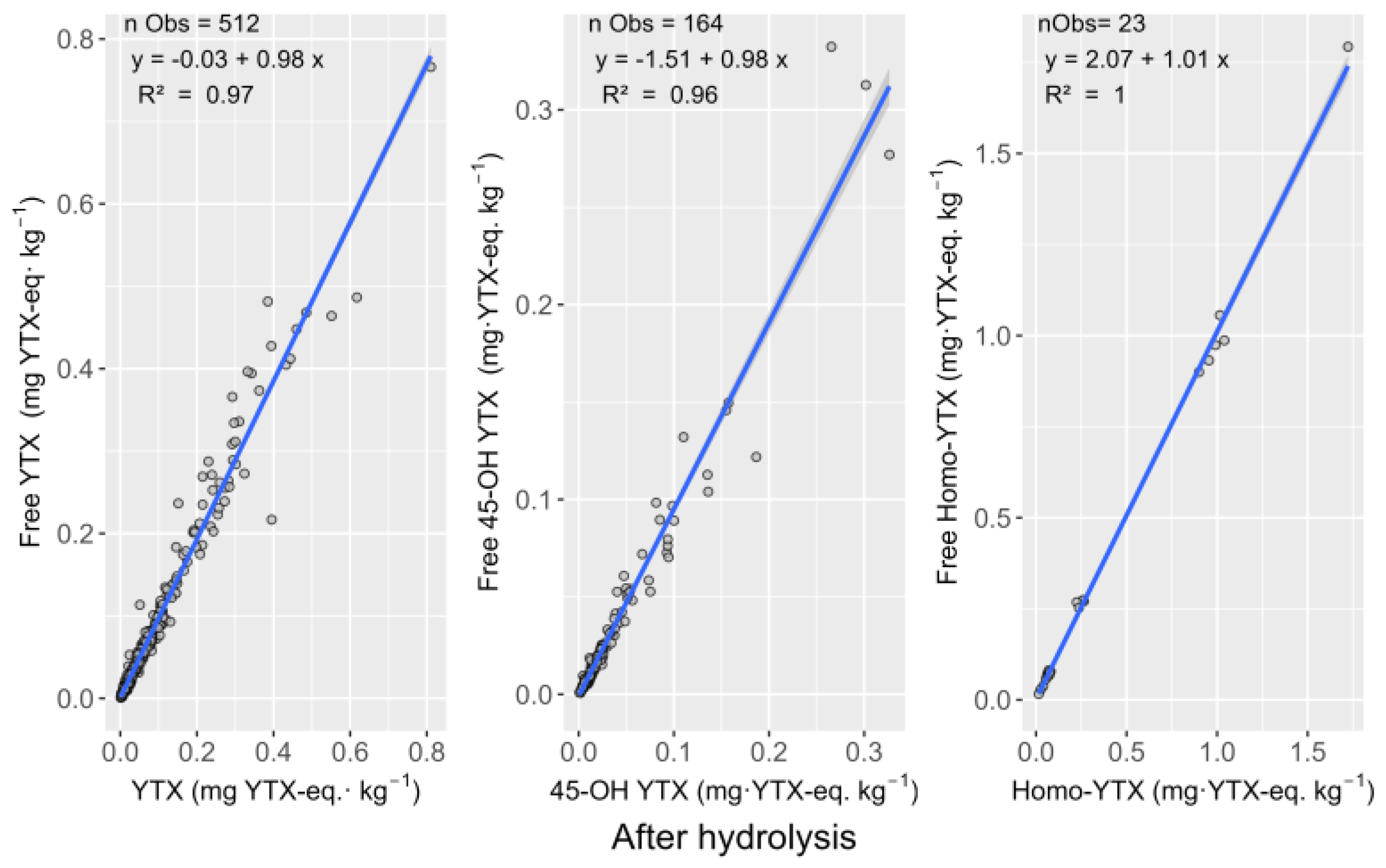
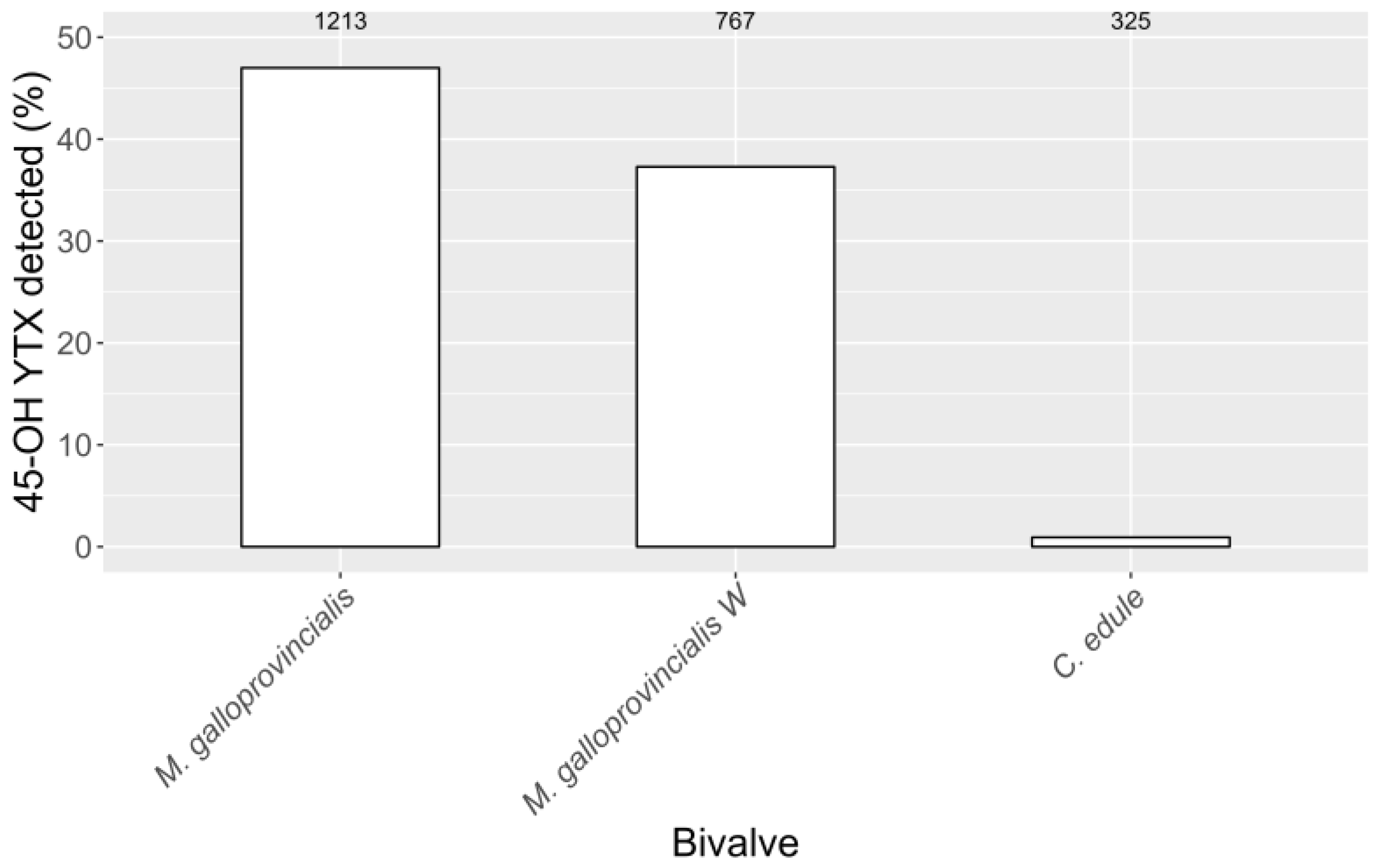
2.2. Hydrolisable Derivatives and Quantification after Alkaline Hydrolisis
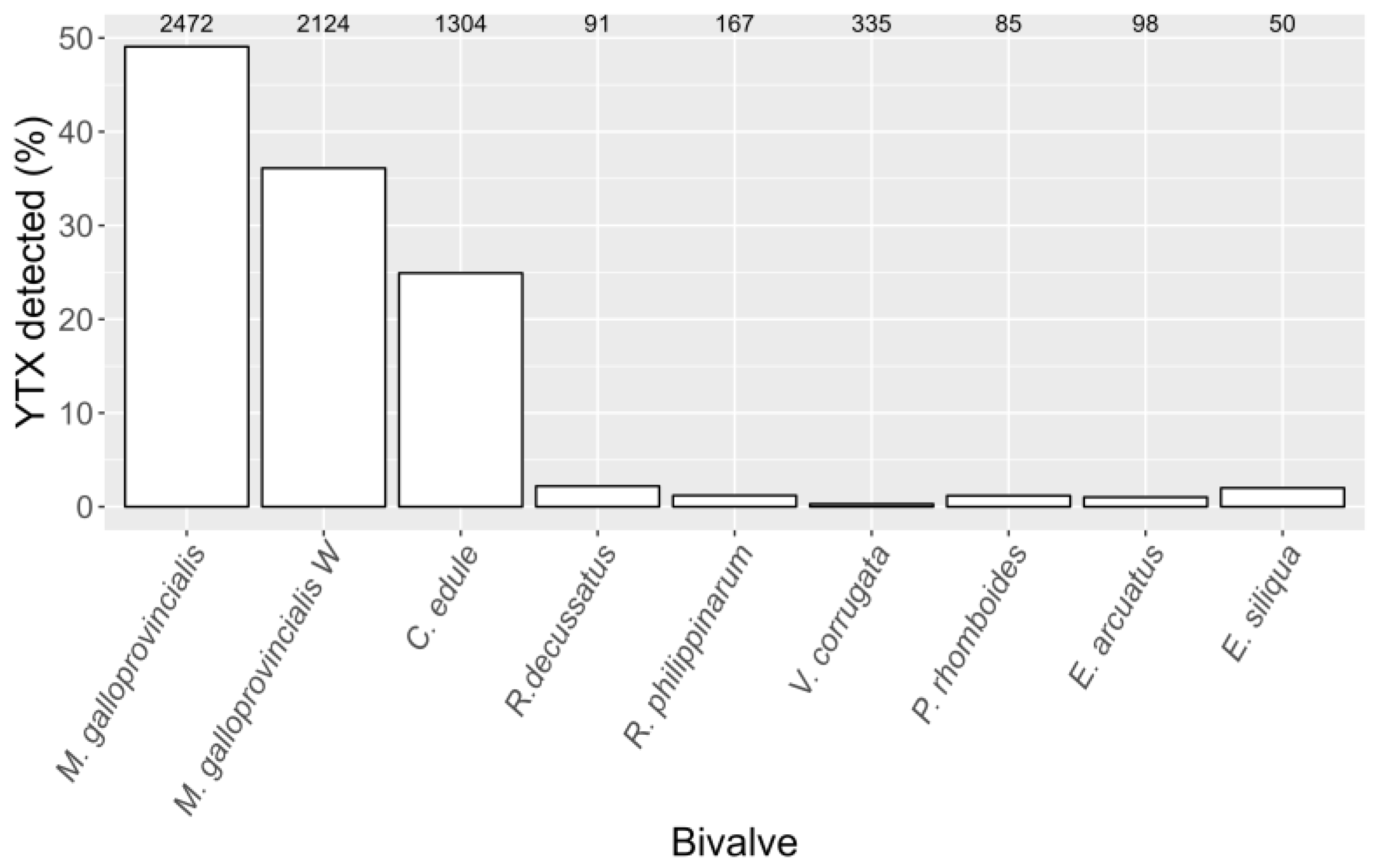
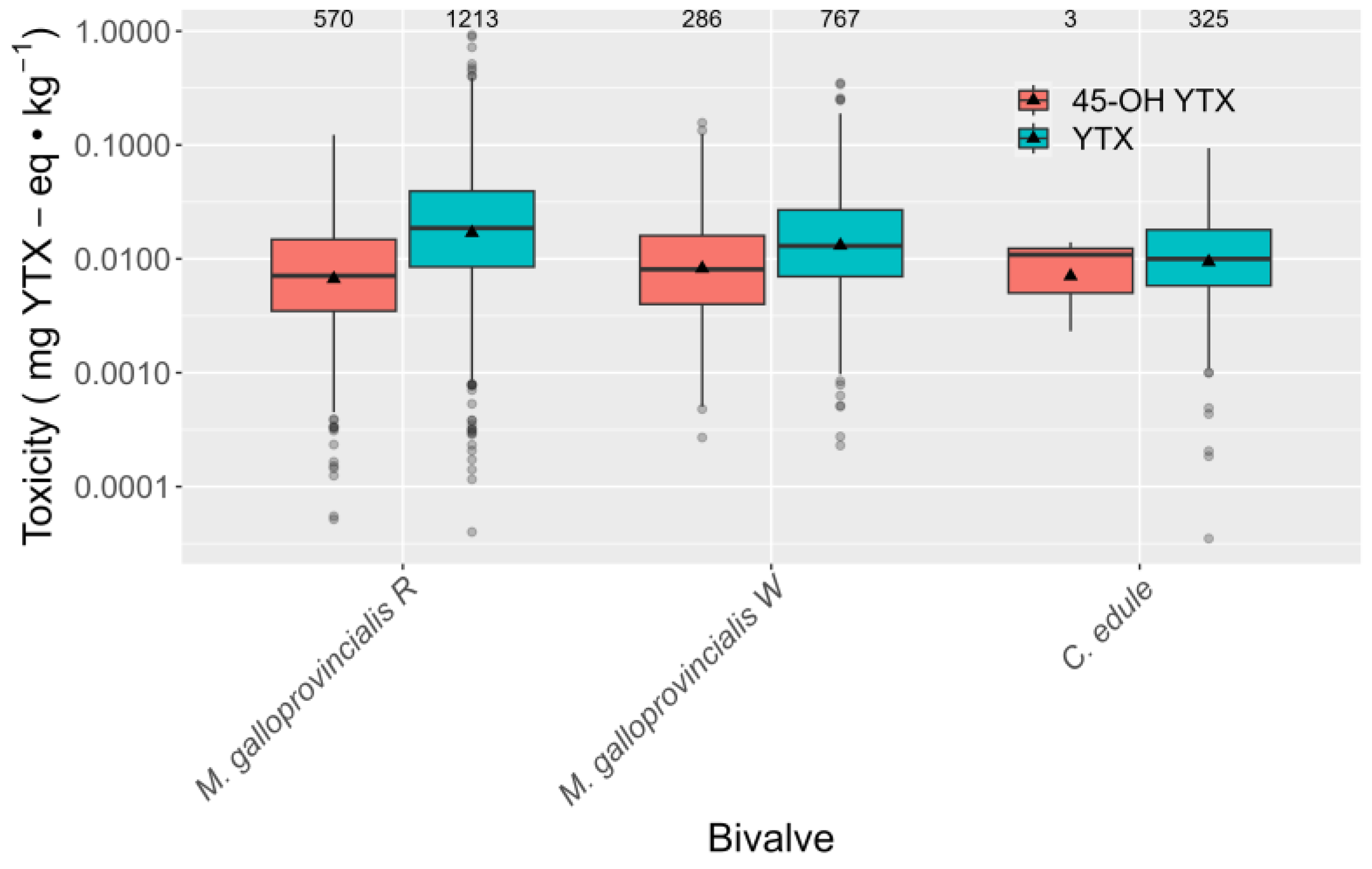
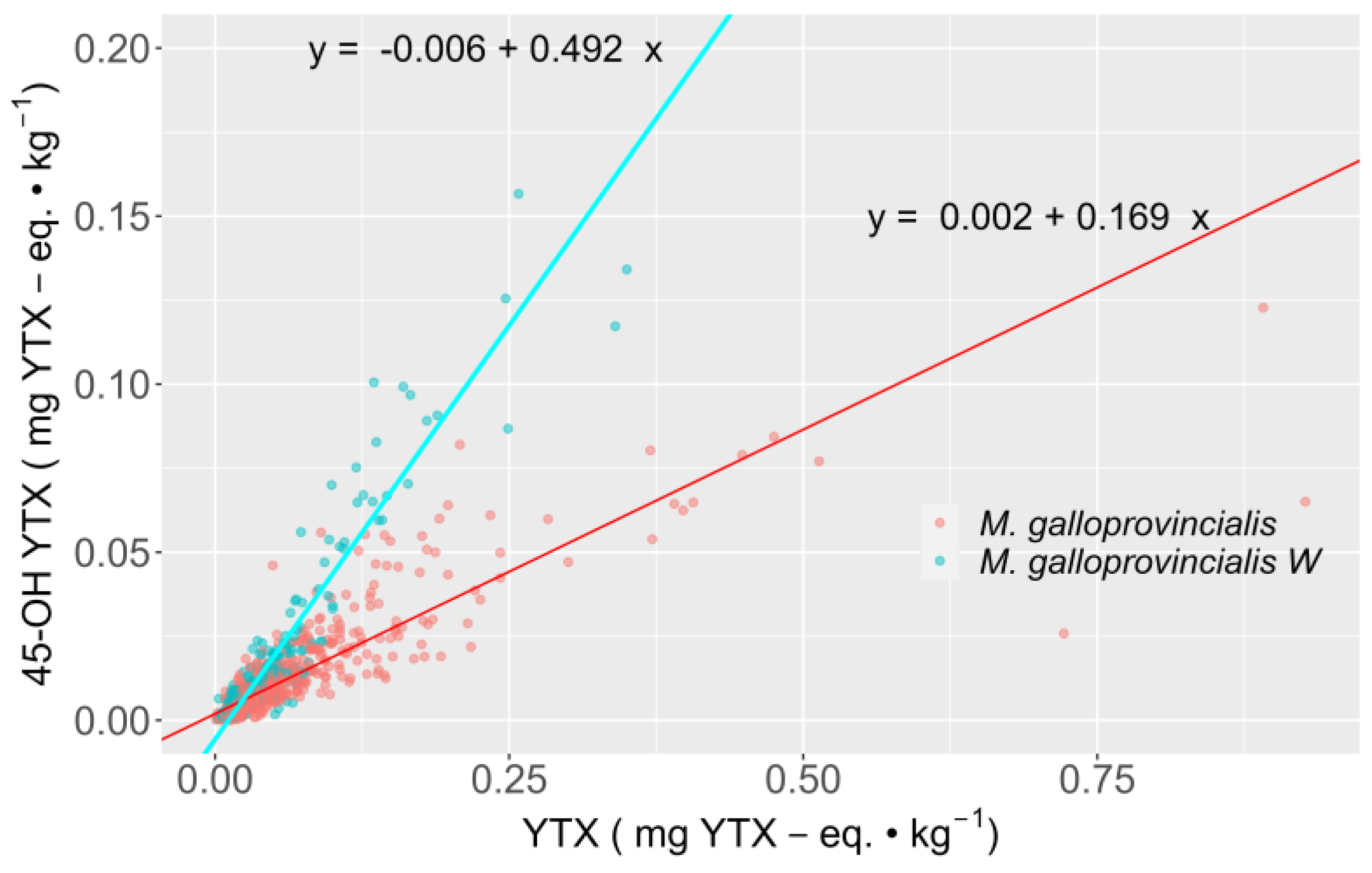
2.3. Interspecific Variation
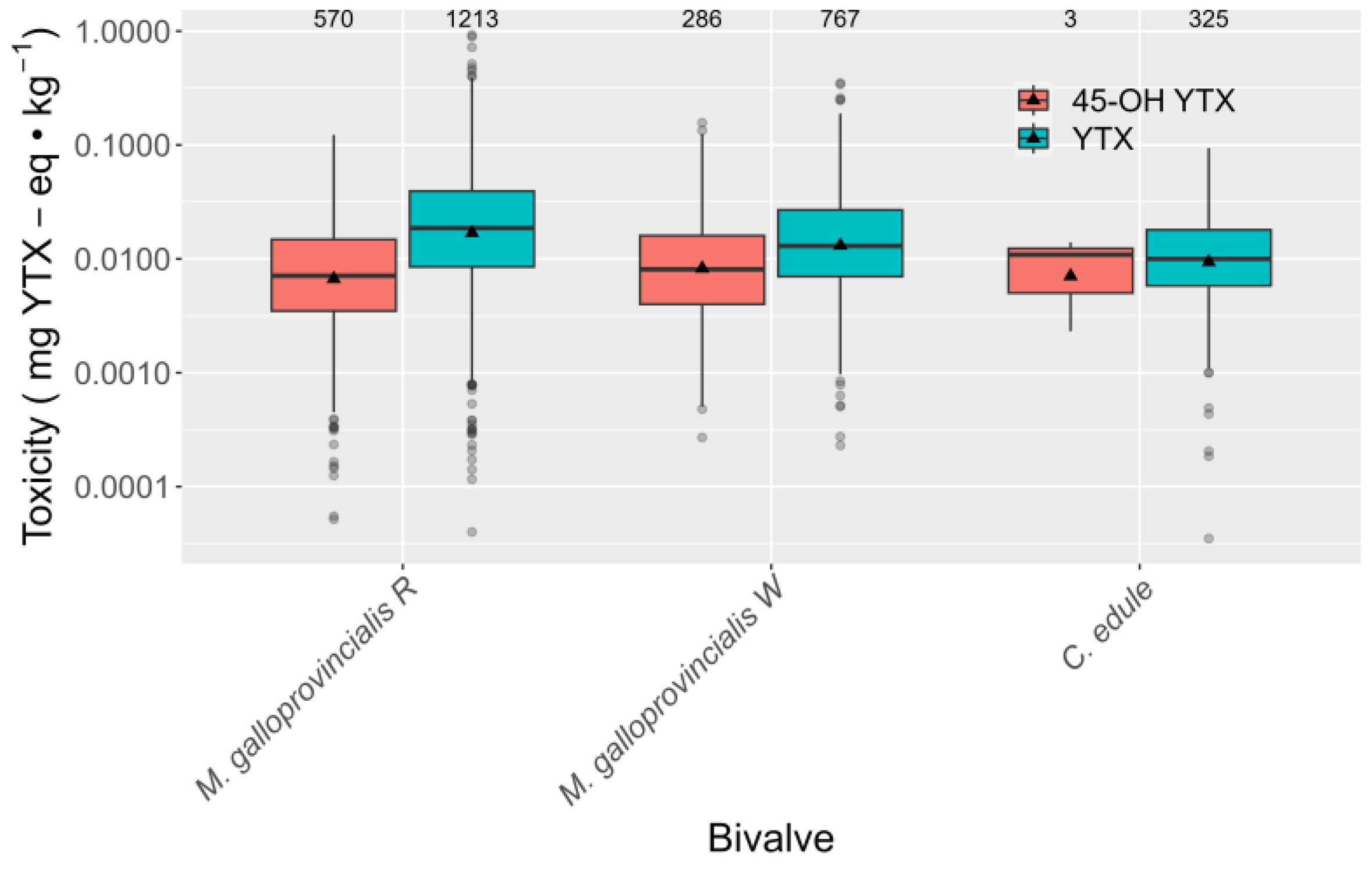
2.4. Relationship between YTX and 45-OH YTX
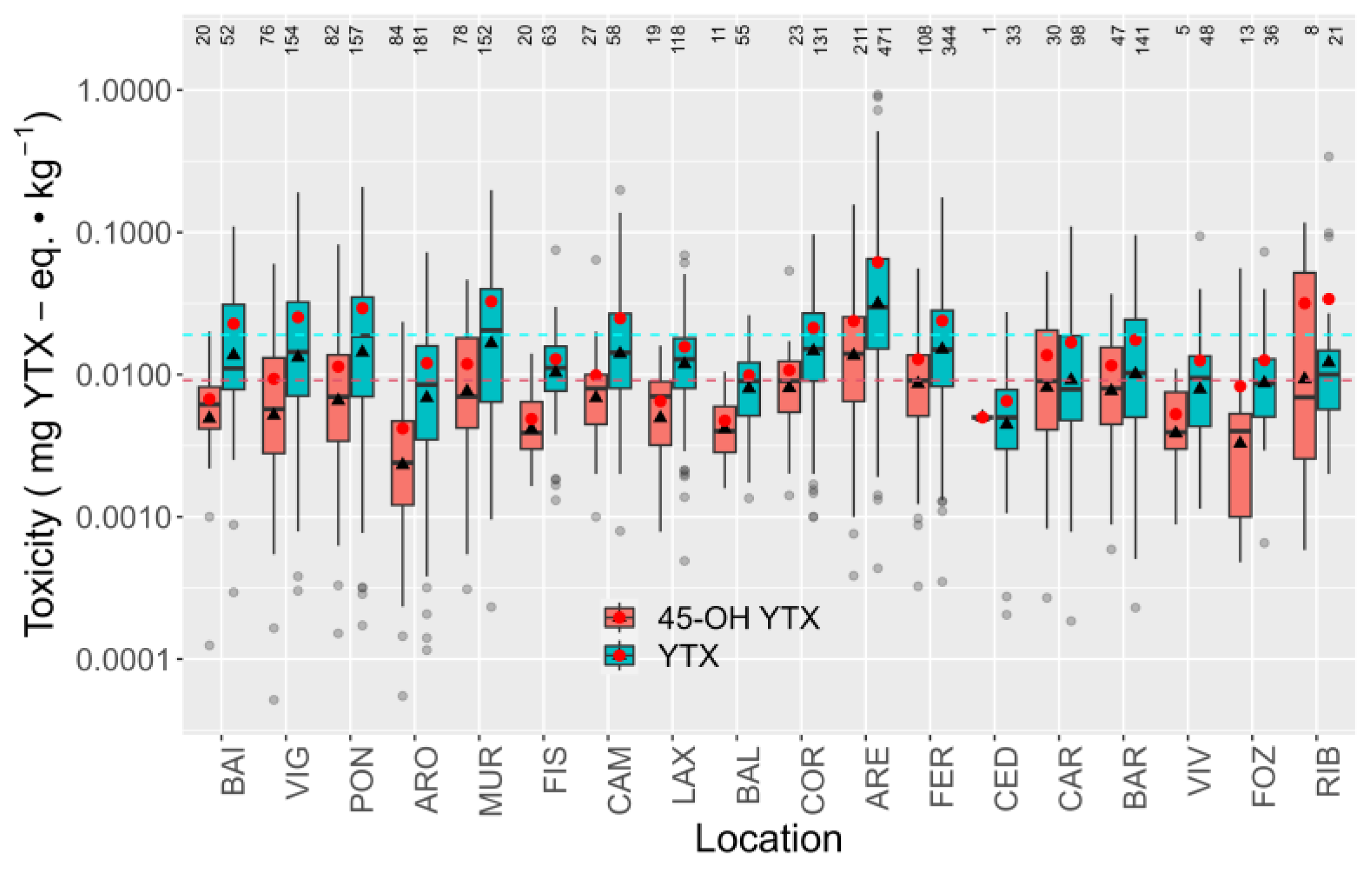
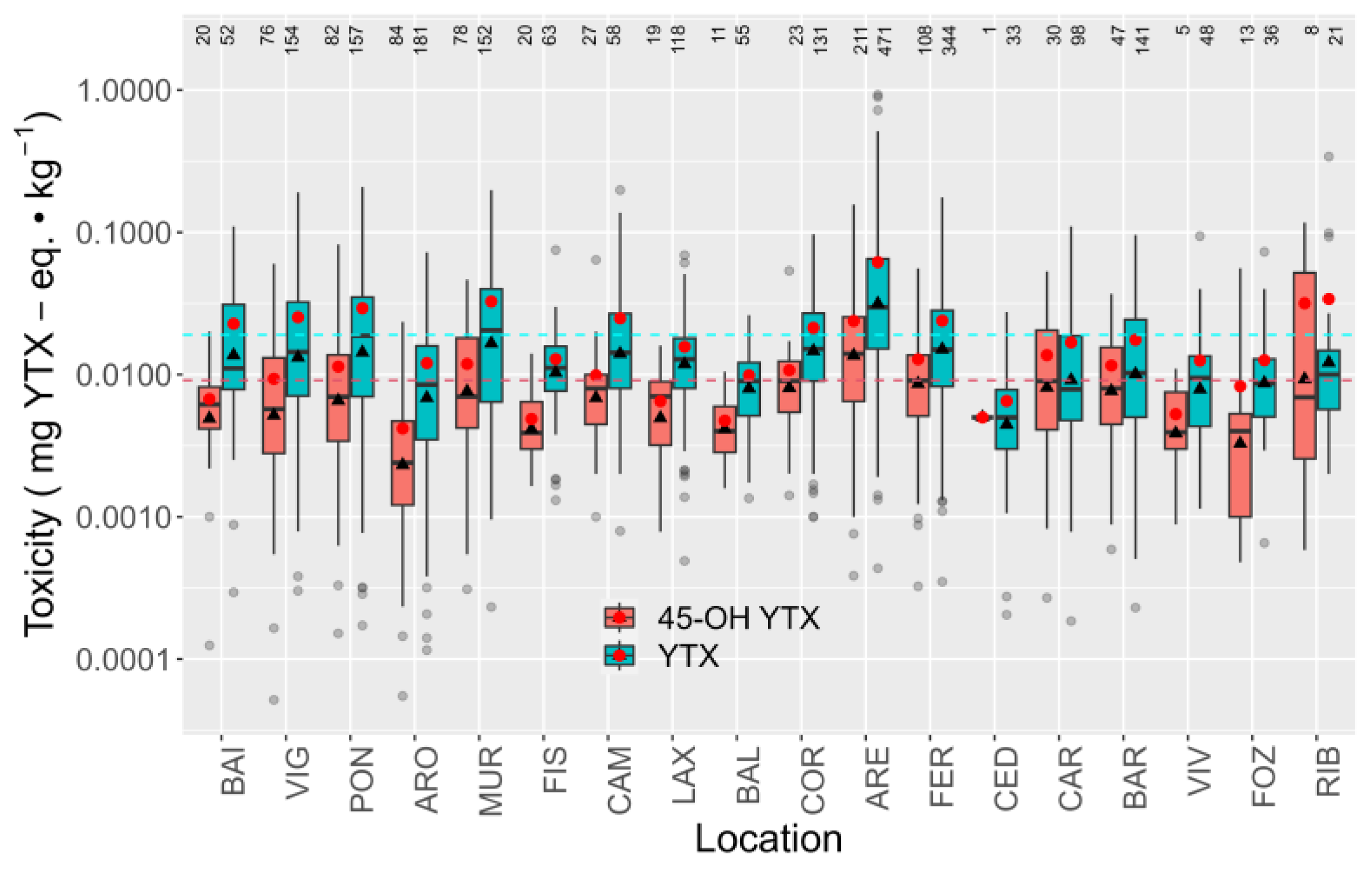
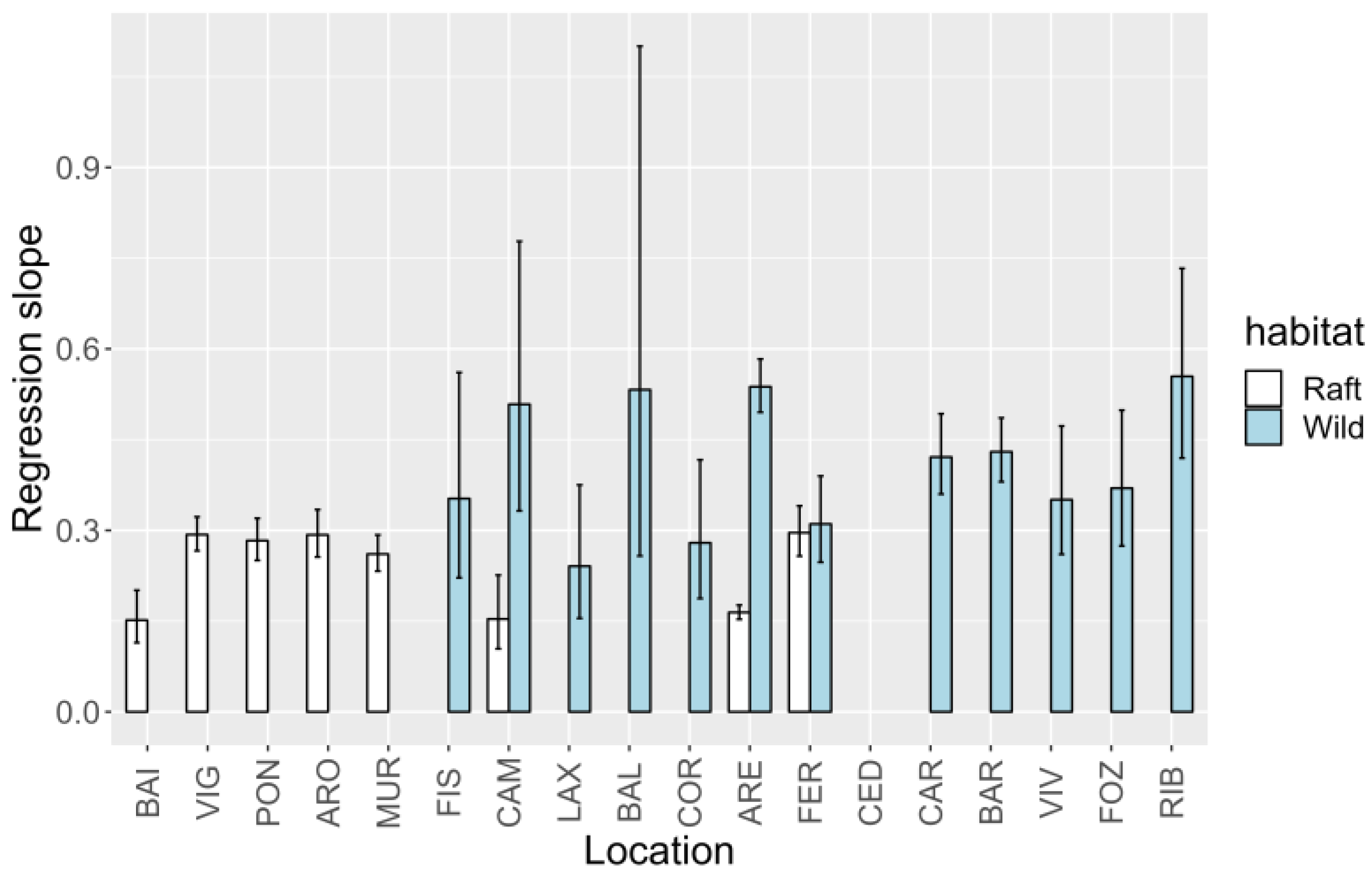
2.5. Spatial Variation
2.5.1. Toxicities
2.5.2. Relationship between YTX and 45-OH YTX
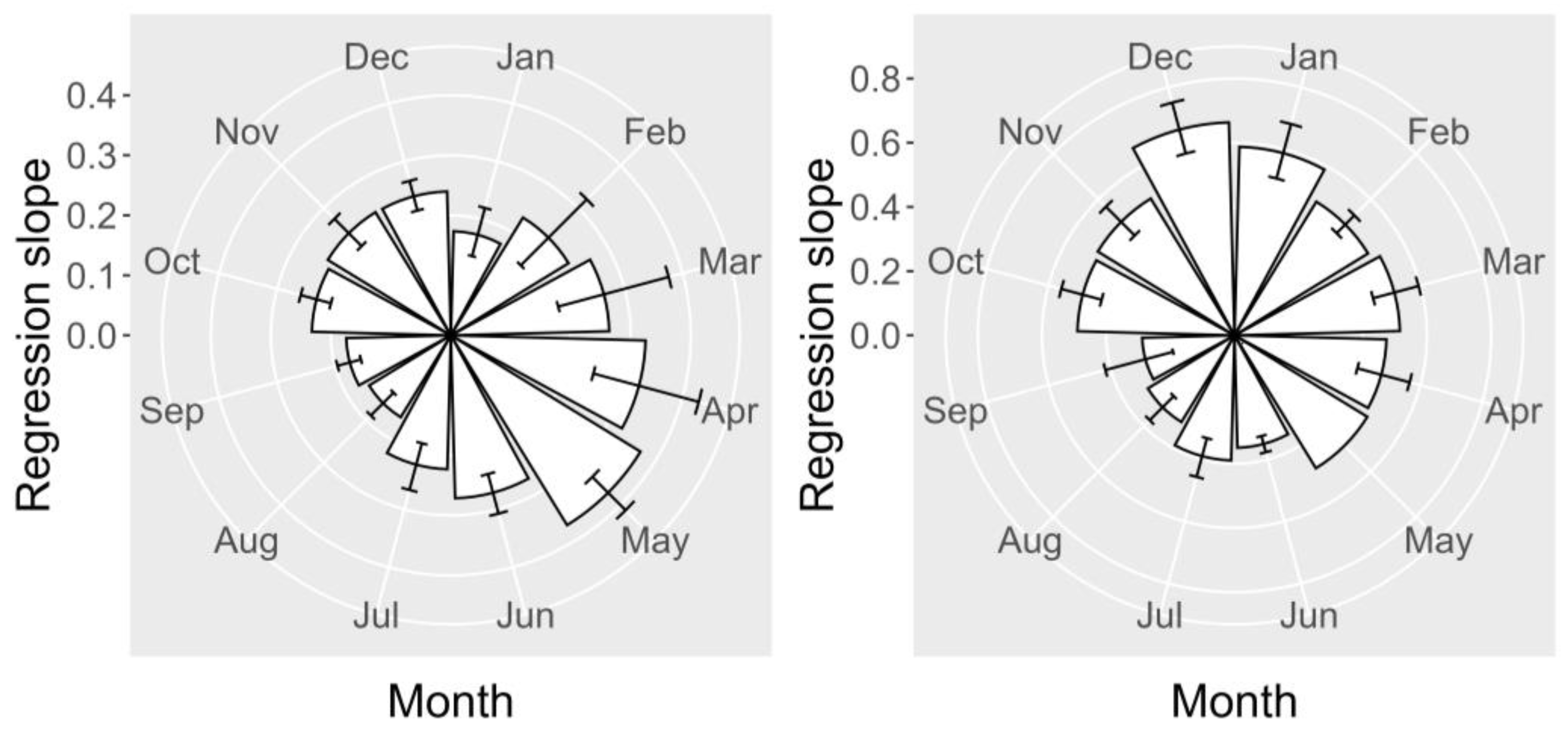
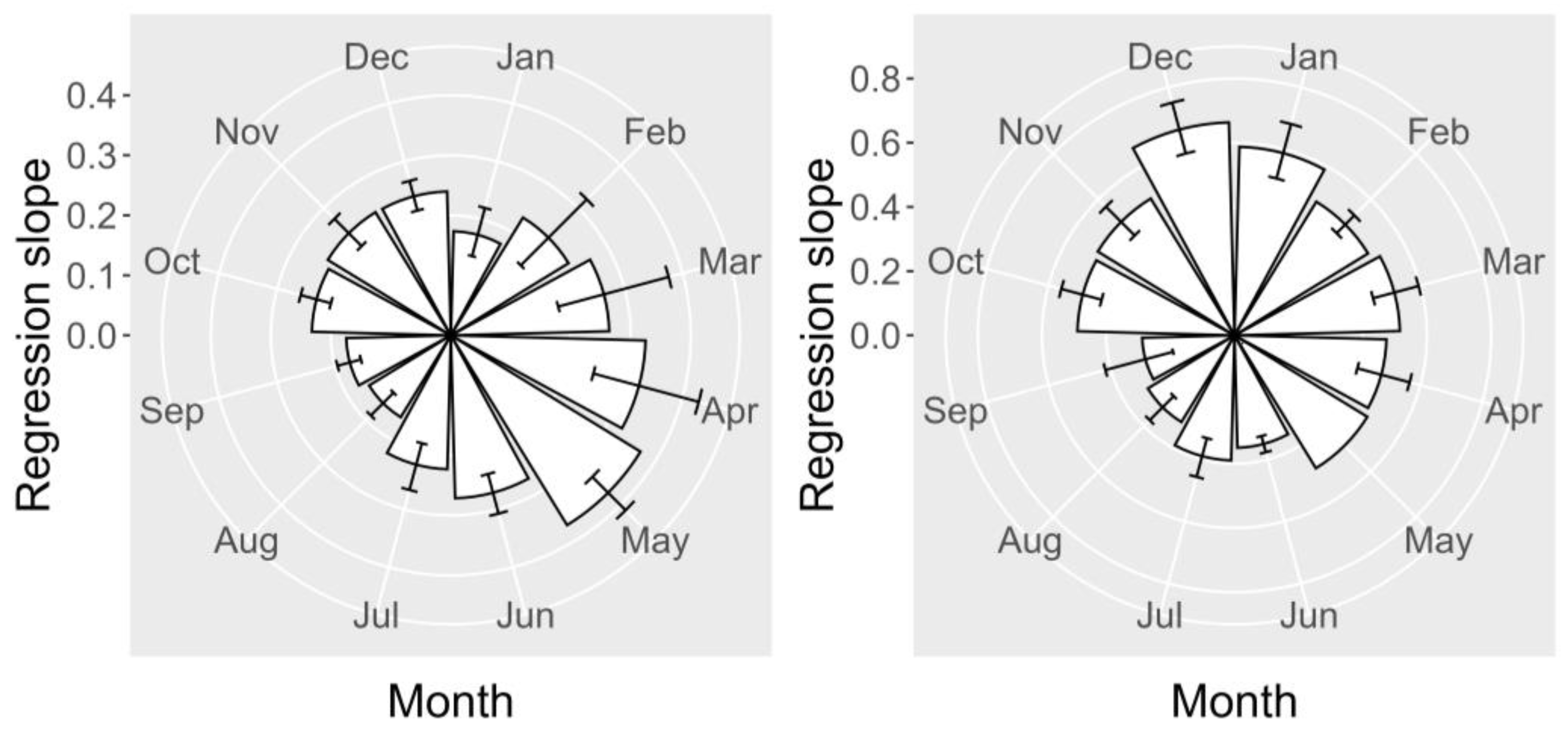
2.6. Seasonal Variation
2.6.1. Toxicities
2.6.2. Relationship between YTX and 45-OH YTX
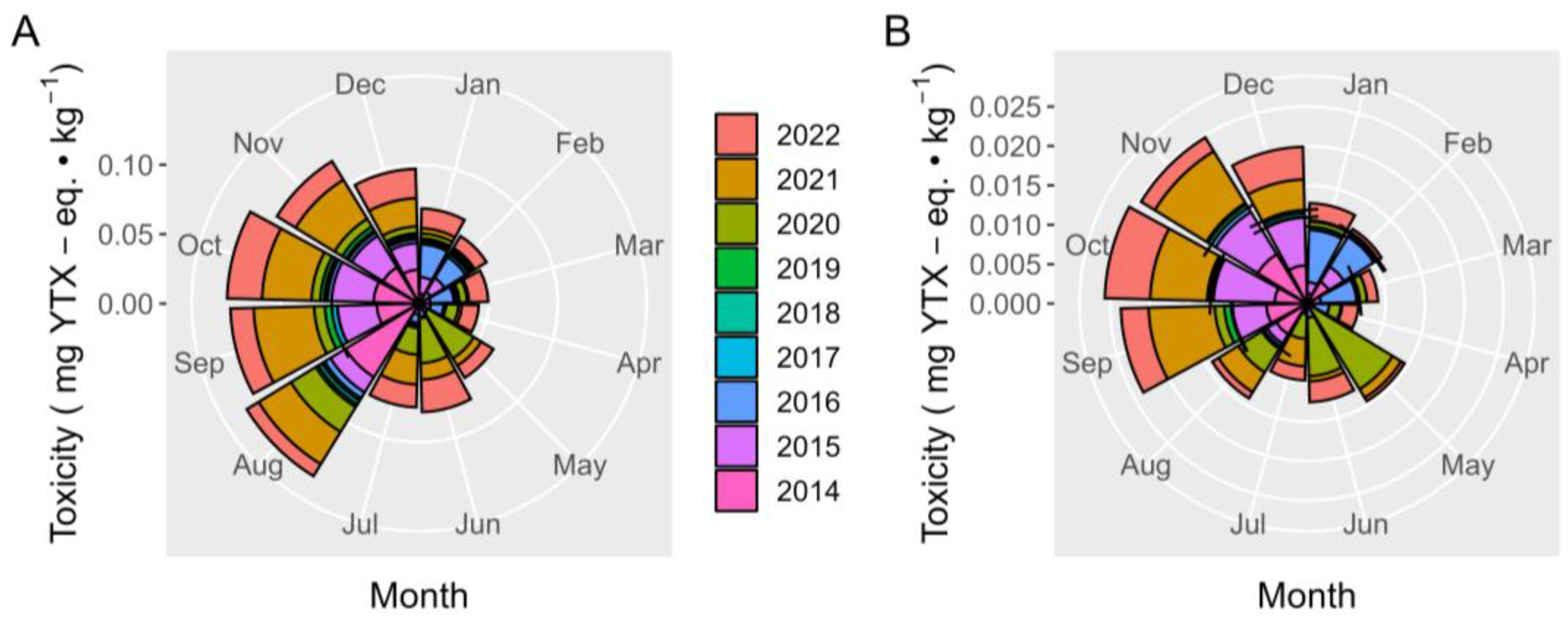
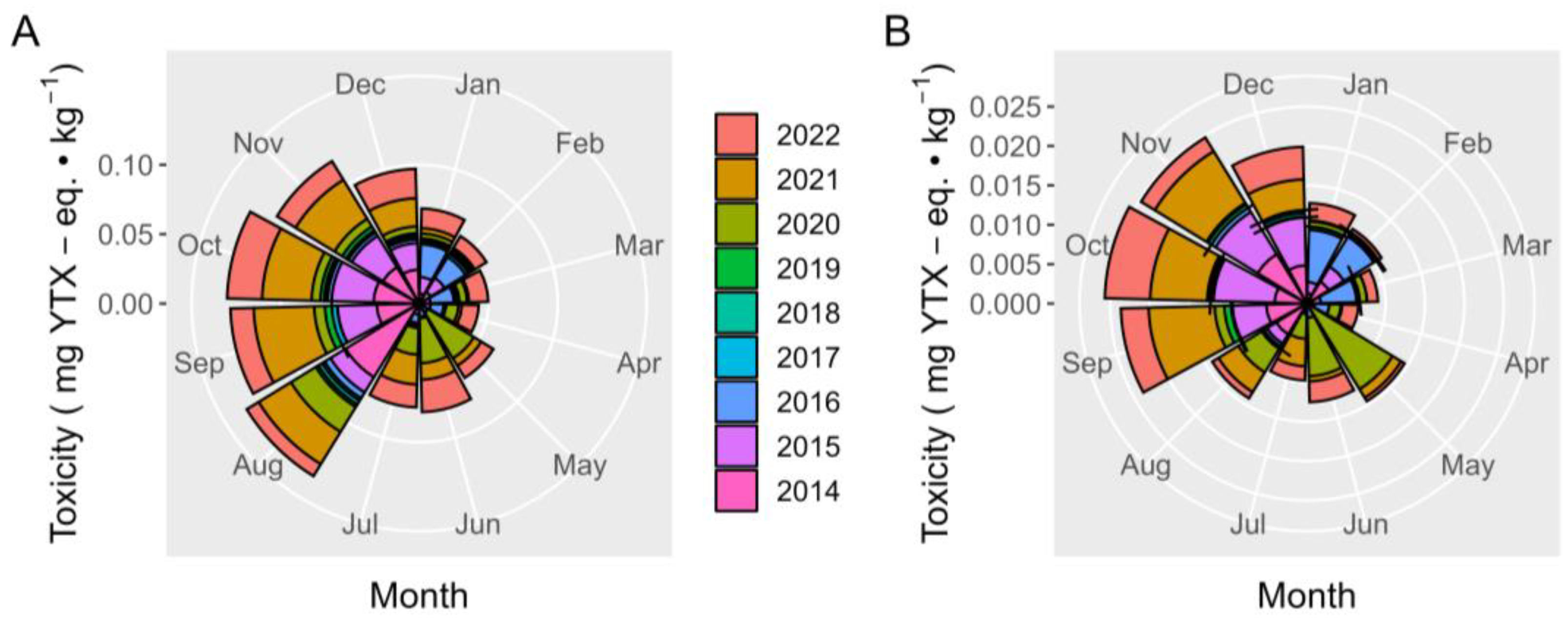
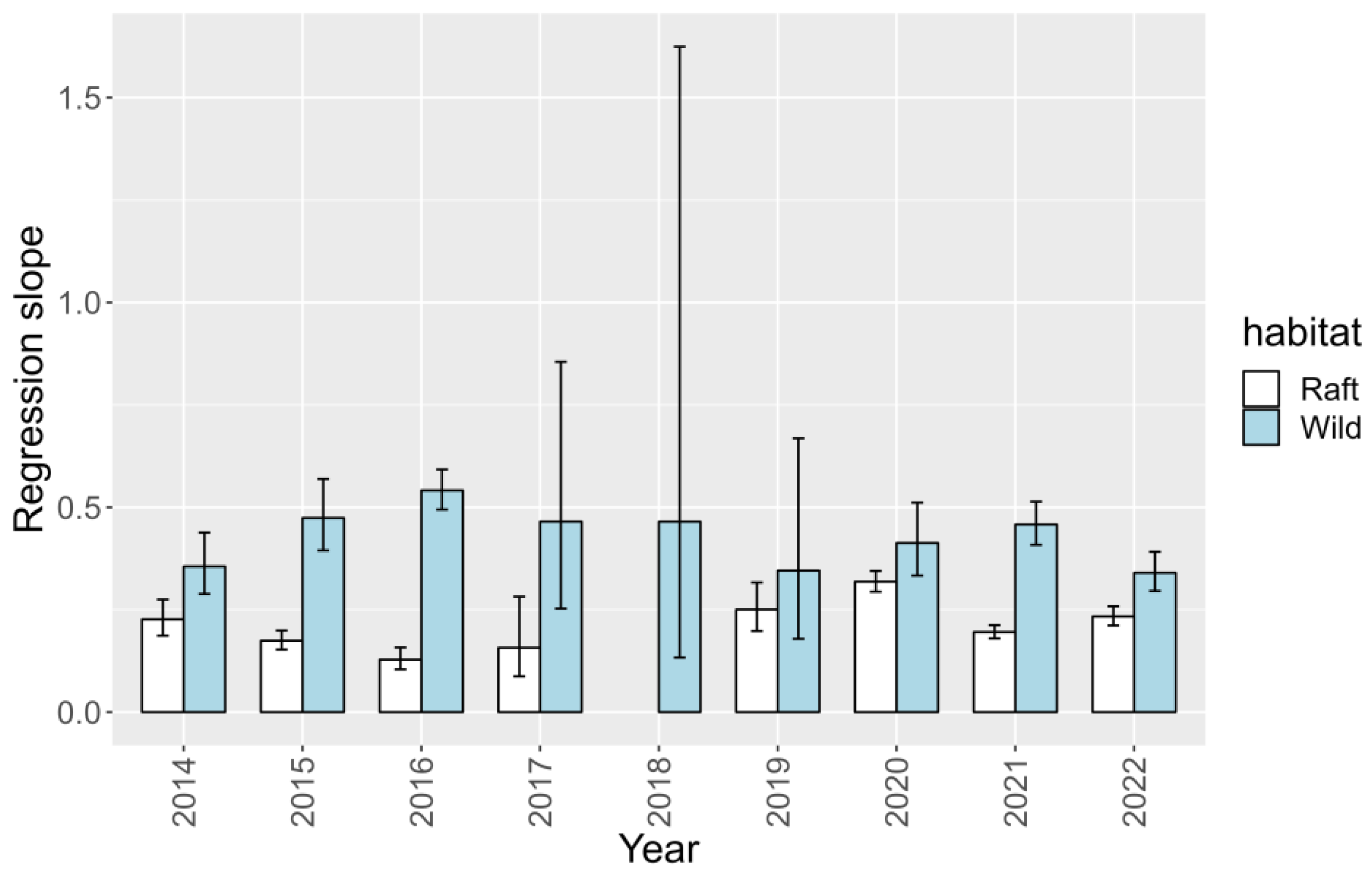
2.7. Interannual Variation
2.7.1. Toxicities
2.7.2. Relationship 45-OH YTX YTX
3. Discussion
3.1. Toxin Profile and Possible Use of Hydrolyzed Extracts to Quantity YTX
3.2. YTX Variability among Bivalve Species
3.3. Spatial Variation of YTXs
3.4. Seasonality
3.5. Interannual Variation
4. Conclusions
5. Material and Methods
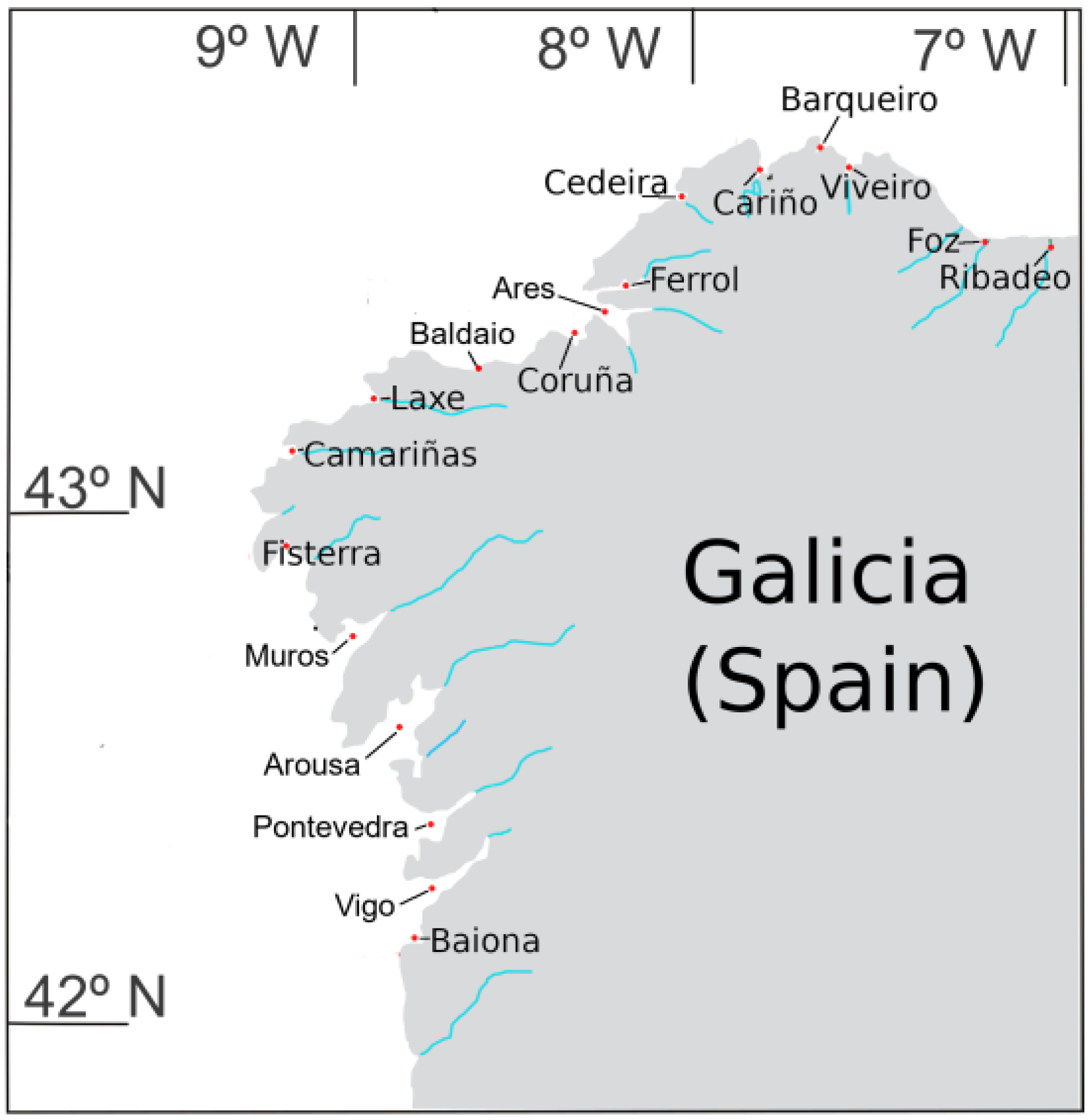
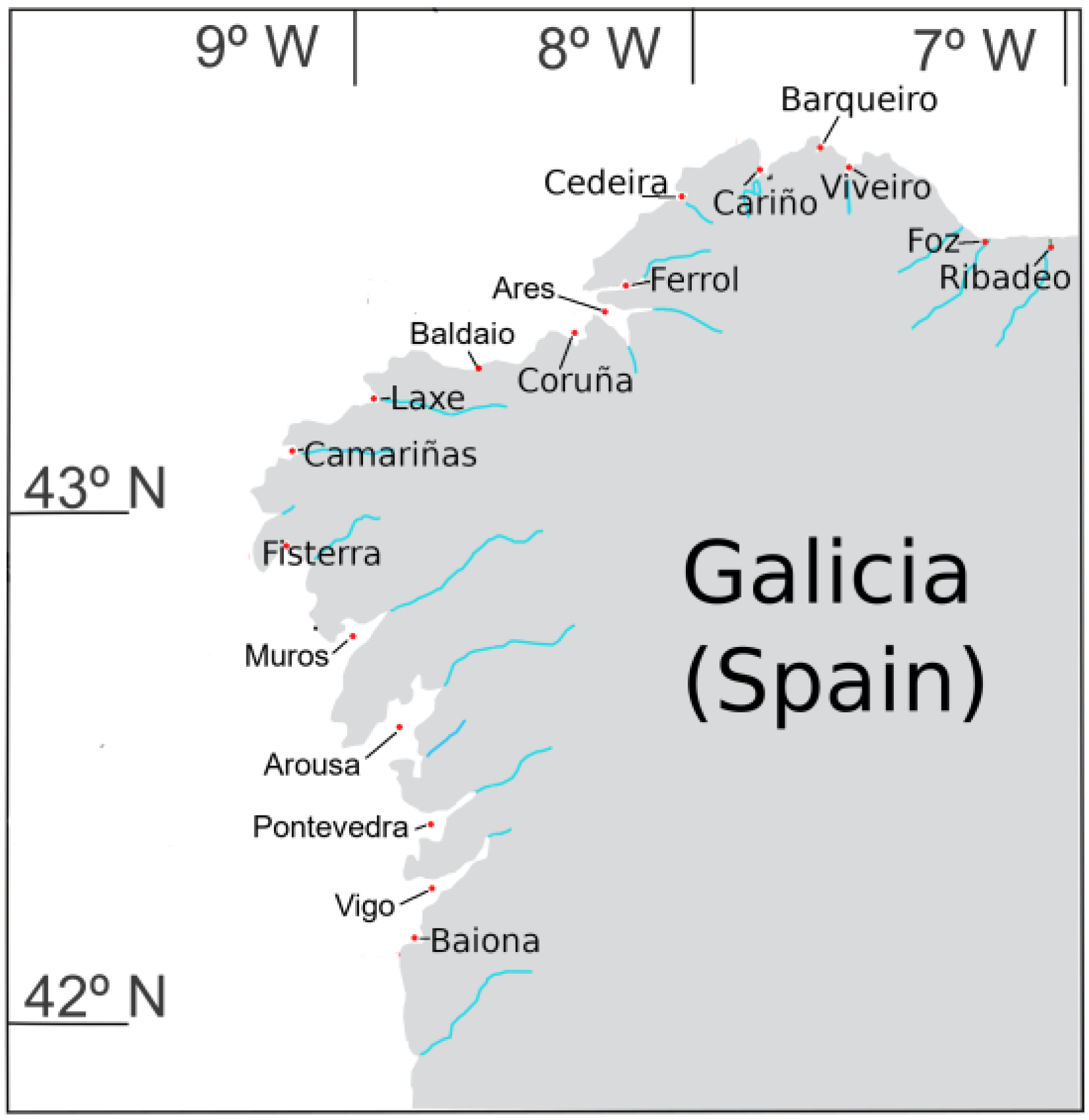
5.1. Sampling
5.2. Analysis
5.2.1. Extraction and Hydrolysis
5.2.2. LC-MS/MS
5.3. Statistical Analysis
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Suzuki, T. Lipophilic Toxins, Pectenotoxins, and Yessotoxins: Chemistry, Metabolism, and Detection. In Seafood and Freshwater Toxins: Pharmacology, Physiology, and Detection; Botana, L.M., Ed.; CRC Press: Boca Ratón, FL, USA, 2014. [Google Scholar]
- Paz, B.; Daranas, A.H.; Norte, M.; Riobo, P.; Franco, J.M.; Fernández, J.J. Yessotoxins, a group of marine polyether toxins: An overview. Mar. Drugs 2008, 6, 73–102. [Google Scholar] [CrossRef] [PubMed]
- Peter, C.; Krock, B.; Cembella, A. Effects of salinity variation on growth and yessotoxin composition in the marine dinoflagellate Lingulodinium polyedra from a Skagerrak fjord system (western Sweden). Harmful Algae 2018, 78, 9–17. [Google Scholar] [CrossRef] [PubMed]
- Suzuki, T.; Horie, Y.; Koike, K.; Satake, M.; Oshima, Y.; Iwataki, M.; Yoshimatsu, S. Yessotoxin Analogues in Several Strains of Protoceratium Reticulatum in Japan Determined by Liquid Chromatography-Hybrid Triple Quadrupole/Linear Ion Trap Mass Spectrometry. J. Chromatogr. A 2007, 1142, 172–177. [Google Scholar] [CrossRef]
- Satake, M.; Eiki, K.; Ichimura, T.; Ota, S.; Sekiguchi, K.; Oshima, Y. Structure of 45,46,47-trinorhomoyessotoxin, a new yessotoxin analog, from Protoceratium reticulatum which represents the first detection of a homoyessotoxin analog in Japan. Harmful Algae 2006, 5, 731–735. [Google Scholar] [CrossRef]
- Miles, C.O.; Samdal, I.A.; Aasen, J.A.G.; Jensen, D.J.; Quilliam, M.A.; Petersen, D.; Briggs, L.M.; Wilkins, A.L.; Rise, F.; Cooney, J.M.; et al. Evidence for Numerous Analogs of Yessotoxin in Protoceratium Reticulatum. Harmful Algae 2005, 4, 1075–1091. [Google Scholar] [CrossRef]
- Ciminiello, P.; Fattorusso, E.; Forino, M.; Poletti, R.; Viviani, R. A new analogue of yessotoxin, carboxyyessotoxin, isolated from Adriatic Sea mussels. Eur. J. Org. Chem. 2000, 2000, 291–295. [Google Scholar] [CrossRef]
- Ciminiello, P.; Fattorusso, E.; Forino, M.; Magno, S.; Poletti, R.; Viviani, R. Isolation of adriatoxin, a new analogue of yessotoxin from mussels of the Adriatic sea. Tetrahedron Lett. 1998, 39, 8897–8900. [Google Scholar] [CrossRef]
- Satake, M.; Tubaro, A.; Lee, J.S.; Yasumoto, Y. Two new analogues of yessotoxin, homoyessotoxin and 45-hydroxyhomoyessotoxin, isolated from mussels of the Adriatic Sea. Nat. Tox. 1997, 5, 107–110. [Google Scholar] [CrossRef]
- Satake, M.; Terasawa, K.; Kadowaki, Y.; Yasumoto, T. Relative configuration of yessotoxin and isolation of two new analogs from toxic scallops. Tetrahedron Lett. 1996, 37, 5955–5958. [Google Scholar] [CrossRef]
- Murata, M.; Kumagai, M.; Lee, J.S.; Yasumoto, T. Isolation and structure of yessotoxin, a novel polyether compound implicated in diarrhetic shellfish poisoning. Tetrahedron Lett. 1987, 28, 5869–5872. [Google Scholar] [CrossRef]
- EFSA Panel on Contaminants in the Food Chain. Marine biotoxins in shellfish—Yessotoxin group-Scientific Opinion of the Panel on Contaminants in the Food chain. EFSA J. 2009, 7, 907. [Google Scholar] [CrossRef]
- Gerssen, A.; Mulder, P.P.J.; McElhinney, M.A.; de Boer, J. Liquid chromatography–tandem mass spectrometry method for the detection of marine lipophilic toxins under alkaline conditions. J. Chromatogr. A 2009, 1216, 1421–1430. [Google Scholar] [CrossRef] [PubMed]
- Ferreiro, S.F.; Vilarino, N.; Carrera, C.; Louzao, M.C.; Cantalapiedra, A.G.; Santamarina, G.; Cifuentes, J.M.; Vieira, A.C.; Botana, L.M. Subacute Cardiotoxicity of Yessotoxin: In Vitro and in Vivo Studies. Chem. Res. Toxicol. 2016, 29, 981–990. [Google Scholar] [PubMed]
- Martin-Lopez, A.; Gallardo-Rodríguez, J.J.; Sanchez-Miron, A.; García-Camacho, F.; Molina-Grima, E. Cytotoxicity of yessotoxin and okadaic acid in mouse T lymphocyte cell line EL-4. Toxicon 2012, 60, 1049–1056. [Google Scholar] [CrossRef] [PubMed]
- Ferreiro, S.F.; Vilarino, N.; Carrera, C.; Louzao, M.C.; Santamarina, G.; Cantalapiedra, A.G.; Cifuentes, J.M.; Vieira, A.C.; Botana, L.M. Subacute immunotoxicity of the marine phycotoxin yessotoxin in rats. Toxicon 2017, 129, 74–80. [Google Scholar] [PubMed]
- Aune, T.; Sorby, R.; Yasumoto, T.; Ramstad, H.; Landsverk, T. Comparison of oral and intraperitoneal toxicity of yessotoxins towards mice. Toxicon 2002, 40, 77–82. [Google Scholar] [CrossRef]
- Tubaro, A.; Sosa, S.; Carbonatto, M.; Altinier, G.; Vita, F.; Melato, M.; Satake, M.; Yasumoto, T. Oral and intraperitoneal acute toxicity studies of yessotoxin and homoyessotoxins in mice. Toxicon 2003, 41, 783–792. [Google Scholar] [CrossRef]
- Tubaro, A.; Sosa, S.; Altinier, G.; Soranzo, M.R.; Satake, M.; Della Loggia, R.; Yasumoto, T. Short-Term Oral Toxicity of Homoyessotoxins, Yessotoxin and Okadaic Acid in Mice. Toxicon 2004, 43, 439–445. [Google Scholar] [CrossRef]
- EU. COMMISSION REGULATION (EU) No 786/2013 of 16 August 2013 amending Annex III to Regulation (EC) No 853/2004 of the European Parliament and of the Council as regards the permitted limits of yessotoxins in live bivalve molluscs. Off. J. Eur. Union 2013, 14, 1. [Google Scholar] [CrossRef]
- Satake, M.; MacKenzie, L.; Yasumoto, T. Identification of Protoceratium reticulatum as the biogenetic origin of yessotoxin. Nat Toxins 1997, 5, 164–167. [Google Scholar] [CrossRef]
- Satake, M.; Ichimura, T.; Sekiguchi, K.; Yoshimatsy, S.; Oshima, Y. Confirmation of yessotoxsin an 45, 46, 47-Trinoryessotoxin production by Protoceratium reticulatum collected in Japan. Nat. Toxins 1999, 7, 147–150. [Google Scholar] [PubMed]
- Armstrong, M.; Kudela, R. Evaluation of California isolates of Lingulodinium polyedrum for the production of yessotoxin. Afr. J. Mar. Sci. 2006, 28, 399–401. [Google Scholar]
- Paz, B.; Riobo, P.; Fernández, M.L.; Fraga, S.; Franco, J.M. Production and release of yessotoxins by the dinoflagellates Protoceratium reticulatum and Lingulodinium polyedrum in culture. Toxicon 2004, 44, 251–258. [Google Scholar] [CrossRef] [PubMed]
- Tubaro, A.; Sirari, L.; Della Loggia, R.; Yasumoto, T. Occurrence of yessotoxin-like toxins in phytoplankton and mussels from the northern Adriatic Sea. In Harmful Algae; Reguera, B., Blanco, J., Fernández, M.L., Wyatt, T., Eds.; Xunta de Galicia and IOC of UNESCO: Santiago de Compostela, Spain, 1998; pp. 470–472. [Google Scholar]
- Rhodes, L.; McNabb, P.; De Salas, M.; Briggs, L.; Beuzenberg, V.; Gladstone, M. Yessotoxin Production by Gonyaulax spinifera. Harmful Algae 2006, 5, 148–155. [Google Scholar] [CrossRef]
- Riccardi, M.; Guerrini, F.; Roncarati, F.; Milandri, A.; Cangini, M.; Pigozzi, S.; Riccardi, E.; Ceredi, A.; Ciminiello, P.; Dell’Aversano, C.; et al. Gonyaulax spinifera from the Adriatic sea: Toxin production and phylogenetic analysis. Harmful Algae 2009, 8, 279–290. [Google Scholar] [CrossRef]
- Álvarez, G.; Uribe, E.; Regueiro, J.; Blanco, J.; Fraga, S. Gonyaulax taylorii, a new yessotoxins-producer dinoflagellate species from Chilean waters. Harmful Algae 2016, 58, 8–15. [Google Scholar]
- Ciminiello, P.; Fatorusso, E.; Forino, M.; Magno, S.; Poletti, R.; Satake, M.; Viviani, R.; Yasumoto, T. Yessotoxin in mussels of the Northern Adriatic Sea. Toxicon 1997, 35, 177–183. [Google Scholar]
- Ciminiello, P.; Fattorusso, E.; Forino, M.; Magno, S.; Poletti, R.; Viviani, R. Isolation of 45-hydroxyyessotoxin from mussels of the Adriatic Sea. Toxicon 1999, 37, 689–693. [Google Scholar]
- Draisci, R.; Ferretti, E.; Palleschi, L.; Marchiafava, C.; Poletti, R.; Milandri, A.; Ceredi, A.; Pompei, M. High levels of yessotoxin in mussels and presence of yessotoxin and homoyessotoxin in dinoflagellates of the Adriatic Sea. Toxicon 1999, 37, 1187–1193. [Google Scholar] [CrossRef]
- Ciminiello, P.; Dell’Aversano, C.; Fattorusso, E.; Forino, M.; Magno, S.; Poletti, R. The detection and identification of 42,43,44,45,46,47,55-heptanor-41-oxoyessotoxin, a new marine toxin from adriatic shellfish, by liquid chromatography-mass spectrometry. Chem. Res. Toxicol. 2002, 15, 979–984. [Google Scholar]
- Ciminiello, P.; Dell’Aversano, C.; Fattorusso, E.; Forino, M.; Magno, S.; Guerrini, F.; Pistocchi, R.; Boni, L. Complex yessotoxins profile in Protoceratium reticulatum from north-western Adriatic sea revealed by LC-MS analysis. Toxicon 2003, 42, 7–14. [Google Scholar] [CrossRef] [PubMed]
- Ciminiello, P.; Dell’Aversano, C.; Fattorusso, E.; Forino, M.; Grauso, L.; Magno, S.G.; Poletti, R.; Tartaglione, L. Desulfoyessotoxins from adriatic mussels: A new problem for seafood safety control. Chem. Res. Toxicol. 2007, 20, 95–98. [Google Scholar] [CrossRef] [PubMed]
- Custovic, S.; Orhanovic, S.; Nincevic-Gladan, Z.; Josipovic, T.; Pavela-Vrancic, M. Occurrence of yessotoxin (YTX) in the coastal waters of the eastern-mid Adriatic Sea (Croatia). Fresenius Environ. Bull. 2009, 18, 1452–1455. [Google Scholar]
- Gladan, Z.N.; Ujevic, I.; Milandri, A.; Marasovic, I.; Ceredi, A.; Pigozzi, S.; Arapov, J.; Skejic, S.; Orhanovic, S.; Isajlovic, I. Is Yessotoxin the Main Phycotoxin in Croatian Waters? Mar. Drugs 2010, 8, 460–470. [Google Scholar] [CrossRef] [PubMed]
- Blanco, J.; Arevalo, F.; Correa, J.; Morono, A. Lipophilic Toxins in Galicia (NW Spain) between 2014 and 2017: Incidence on the Main Molluscan Species and Analysis of the Monitoring Efficiency. Toxins 2019, 11, 612. [Google Scholar] [CrossRef] [PubMed]
- Amorim, A.; Palma, A.S.; Sampayo, M.A.; Moita, M.T. On a Lingulodinium polyedrum bloom in Setúbal Bay, Portugal. In Harmful Algal Blooms 2000; Hallegraeff, G.M., Blackburn, S.I., Bolch, C.J., Lewis, R.J., Eds.; IOC of UNESCO: Paris, France, 2001; pp. 133–136. [Google Scholar]
- Gomes, S.S.; Vale, P.; Botelho, M.J.; Rodrigues, S.M.; Cerejo, M.; Vilarinho, M.G. ELISA Screening of yessotoxins in Portuguese shellfish. In Proceedings of the 12th International Conference on Harmful Algae, Copenhagen, Denmark, 4–8 September 2006; Moestrup, Ø.E.A., Ed.; International Society for the Study of Harmful Algae and IOC of UNESCO: Copenhagen, Denmark, 2008; pp. 290–292. [Google Scholar]
- Arévalo, F.; Pazos, Y.; Correa, J.; Salgado, C.; Moroño, A.; Paz, B.; Franco, J.M. First reported case of yessotoxins in mussels in the Galician Rías during a bloom of Lingulodinium polyedrum STEIN (Dodge). In Proceedings of the 5th International Conference on Molluscan Shellfish Safety, Galway, Ireland, 14–18 June 2004; Henshilwood, K., Deegan, B., McMahon, T., Cusack, C., Keaveney, S., Silke, J., O’Cinneide, M., Lyons, D., Hess, P., Henshilwood, K., et al., Eds.; Marine Institute, Galway, Ireland, 2006; pp. 184–189. [Google Scholar]
- Caballero, I.; Fernandez, R.; Escalante, O.M.; Maman, L.; Navarro, G. New capabilities of Sentinel-2A/B satellites combined with in situ data for monitoring small harmful algal blooms in complex coastal waters. Sci. Rep. 2020, 10, 8743. [Google Scholar] [CrossRef]
- Pitcher, G.C.; Foord, C.J.; Macey, B.M.; Mansfield, L.; Mouton, A.; Smith, M.E.; Osmond, S.J.; van der Molen, L. Devastating farmed abalone mortalities attributed to yessotoxin-producing dinoflagellates. Harmful Algae 2019, 81, 30–41. [Google Scholar] [CrossRef]
- Reinecke, P. Gonyaulax grindleyi sp. nov: A dinoflagellate causing a red tide at Elands Bay, Cape Province, in December 1966. J. S. Afr. Bot. 1967, 33, 157–160. [Google Scholar]
- Grindley, J.R.; Nel, E.A. Red water and mussel poisoning at Edlands Bay, December 1966. Fish. Bull. S. Afr. 1970, 6, 36–55. [Google Scholar]
- Horstman, D.A. Reported red-water outbreaks and their effects on fauna of the west and south coasts of South Africa, 1959–1980. Fish. Bull. S. Afr. 1981, 15, 71–88. [Google Scholar]
- King, T.L.; Nguyen, N.; Doucette, G.J.; Wang, Z.H.; Bill, B.D.; Peacock, M.B.; Madera, S.L.; Elston, R.A.; Trainer, V.L. Hiding in plain sight: Shellfish-killing phytoplankton in Washington State. Harmful Algae 2021, 105, 102032. [Google Scholar] [CrossRef] [PubMed]
- Cassis, D. The Effects of Harmful algae on the Summer Mortalities of Juvenile Pacific oysters (Crassotrea gigas). Master’s Thesis, University of British Columbia, Vancouver, BC, Canada, 2005. [Google Scholar]
- De Wit, P.; Rogers-Bennett, L.; Kudela, R.M.; Palumbi, S.R. Forensic genomics as a novel tool for identifying the causes of mass mortality events. Nat. Commun. 2014, 5, 3652. [Google Scholar] [CrossRef] [PubMed]
- Landsberg, J.H. The effects of harmful algal blooms on aquatic organisms. Rev. Fish. Sci. 2002, 10, 113–390. [Google Scholar] [CrossRef]
- Paz, B.; Blanco, J.; Franco, J.M. Yessotoxins production during the culture of Protoceratium reticulatum strains isolated from Galician Rias Baixas (NW Spain). Harmful Algae 2013, 21–22, 13–19. [Google Scholar] [CrossRef]
- Dhanji-Rapkova, M.; Oneill, A.; Maskrey, B.H.; Coates, L.; Swan, S.C.; Alves, M.T.; Kelly, R.J.; Hatfield, R.G.; Rowland-Pilgrima, S.J.; Lewis, A.M.; et al. Variability and profiles of lipophilic toxins in bivalves from Great Britain during five and a half years of monitoring: Azaspiracids and yessotoxins. Harmful Algae 2019, 87, 101629. [Google Scholar] [CrossRef]
- Paz, B.; Riobo, P.; Ramilo, I.; Franco, J.M. Yessotoxins Profile in Strains of Protoceratium reticulatum From Spain and USA. Toxicon 2007, 50, 1–17. [Google Scholar] [CrossRef]
- Barbosa, M.; Amorim, A.; Costa, P.R. Growth and Toxin Profile of Protoceratium reticulatum (Dinoflagellate) isolated from the Portuguese coast. In Proceedings of the XIV Reunião Ibérica sobre Microalgas Nocivas e Biotoxinas Marinhas, Algés-Lisboa, Portugal, 1–3 June 2022; pp. 27–28. [Google Scholar]
- Krock, B.; Alpermann, T.; Tillmann, U.; Pitcher, G.C.; Cembella, A.D. Yessotoxin profiles of the marine dinoflagellates Protoceratium reticulatum and Gonyaulax spinifera. In Proceedings of the 12th International Conference on Harmful Algae, Copenhagen, Denmark, 4–8 September 2006; Moestrup, Ø.E.A., Ed.; IOC of UNESCO: Copenhagen, Denmark, 2008; pp. 303–305. [Google Scholar]
- Roder, K.; Hantzsche, F.M.; Gebuhr, C.; Miene, C.; Helbig, T.; Krock, B.; Hoppenrath, M.; Luckas, B.; Gerdts, G. Effects of salinity, temperature and nutrients on growth, cellular characteristics and yessotoxin production of Protoceratium reticulatum. Harmful Algae 2012, 15, 59–70. [Google Scholar] [CrossRef]
- Samdal, I.A.; Olseng, C.D.; Sandvik, M.; Miles, C.O.; Briggs, L.; Torgersen, T.; Jensen, D.J.; Cooney, J.M. Profile of yessotoxin analogues in a Norwegian strain of Protoceratium reticulatum. In Proceedings of the 5th International Conference on Molluscan Shellfish Safety, Galway, Ireland, 14–18 June 2004; p. 118. [Google Scholar]
- Samdal, I.A.; Aasen, J.A.B.; Briggs, L.R.; Dahl, E.; Miles, C.O. Comparison of Elisa and LC-MS Analyses for Yessotoxins in Blue Mussels (Mytilus edulis). Toxicon 2005, 46, 7–15. [Google Scholar] [CrossRef]
- Paz, B.; Riobo, P.; Souto, M.L.; Gil, L.V.; Norte, M.; Fernández, J.J.; Franco, J.M. Detection and identification of glycoyessotoxin A in a culture of the dinoflagellate Protoceratium reticulatum. Toxicon 2006, 48, 611–619. [Google Scholar] [CrossRef]
- EC. COMMISSION IMPLEMENTING REGULATION (EU) 2019/627 of 15 March 2019 laying down uniform practical arrangements for the performance of official controls on products of animal origin intended for human consumption in accordance with Regulation (EU) 2017/625 of the European Parliament and of the Council and amending Commission Regulation (EC) No 2074/2005 as regards official controls. Off. J. Eur. Union L 2019, L 131, 51. [Google Scholar]
- Yasumoto, T.; Takizawa, A. Fluorometric measurement of yessotoxins in shellfish by high-pressure liquid chromatography. Biosci. Biotechnol. Biochem. 1997, 61, 1775–1777. [Google Scholar] [CrossRef] [PubMed]
- Diz, A.P.; Presa, P. The genetic diversity pattern of Mytilus galloprovincialis in Galician Rías (NW Iberian estuaries). Aquaculture 2009, 287, 278–285. [Google Scholar] [CrossRef]
- Montagnac, V.; Guyondet, T.; Comeau, L.; Tremblay, R. Physiological differences between wild and cultured bivalves in Prince Edward Island, Canada. Aquat. Living Resour. 2020, 33, 8. [Google Scholar] [CrossRef]
- Cáceres-Martínez, J.; Figueras, A. Long-term survey on wild and cultured mussels (Mytilus galloprovincialis Lmk) reproductive cycles in the Ria de Vigo (NW Spain). Aquaculture 1998, 162, 141–156. [Google Scholar] [CrossRef]
- Rodhouse, P.G.; Roden, C.M.; Hensey, M.P.; Ryan, T.H. Resource-Allocation in Mytilus edulis on the Shore and in Suspended Culture. Mar. Biol. 1984, 84, 27–34. [Google Scholar] [CrossRef]
- Labarta, U.; Fernández-Reiriz, M.J. The Galician mussel industry: Innovation and changes in the last forty years. Ocean Coast. Manag. 2019, 167, 208–218. [Google Scholar] [CrossRef]
- Blanco, J. The distribution of dinoflagellate cysts along the Galician (NW Spain) coast. J. Plankton Res. 1995, 17, 283–302. [Google Scholar] [CrossRef]
- Blanco, J. Quistes de Dinoflagelados de las Costas de Galicia. Doctoral Dissertation, Universidad de Santiago de Compostela, Santiago, Spain, 1988. [Google Scholar]
- Margalef, R. Estructura y dinámica de la “purga de mar” en la ría de Vigo. Inv. Pesq. 1956, 5, 113–134. [Google Scholar]
- Santos, M.; Moita, M.T.; Oliveira, P.B.; Amorim, A. Phytoplankton communities in two wide-open bays in the Iberian upwelling system. J. Sea Res. 2021, 167, 101982. [Google Scholar] [CrossRef]
- EURLMB. EU-Harmonised Standard Operating Procedure for Determination of Lipophilic Marine Biotoxins in Molluscs by LC-MS/MS.Version 5. 2014. Available online: https://www.aesan.gob.es/AECOSAN/docs/documentos/laboratorios/LNRBM/ARCHIVO2EU-Harmonised-SOP-LIPO-LCMSMS_Version5.pdf (accessed on 15 June 2023).
- Regueiro, J.; Rossignoli, A.E.; Álvarez, G.; Blanco, J. Automated on-line solid-phase extraction coupled to liquid chromatography tandem mass spectrometry for determination of lipophilic marine toxins in shellfish. Food Chem. 2011, 129, 533–540. [Google Scholar] [CrossRef]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2019. [Google Scholar]
- Wickham, H. ggplot2: Elegant Graphics for Data Analysis; Springer: Berlin/Heidelberg, Germany, 2016. [Google Scholar]
- Falster, D.S.; Warton, D.I.; Wright, I.J. User’s Guide to SMATR: Standardised Major Axis Tests & Routines Version 2.0, Copyright 2006. 2006. Available online: http://www. bio. mq. edu. au/ecology/SMATR/SMATR_users_guide. pdf (accessed on 5 May 2023).
- Warton, D.I.; Duursma, R.A.; Falster, D.S.; Taskinen, S. smatr 3—An R package for estimation and inference about allometric lines: The smatr 3—An R package. Methods Ecol. Evol. 2012, 3, 257–259. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Toxin | Precursor Ion (m/z) | Product Ion (m/z) | Collision Energy (eV) | Cone Voltage (V) |
|---|---|---|---|---|
| YTX | 570.4 | 467.4 | 30 | 45 |
| 396.3 | 35 | 45 | ||
| 45-OH YTX | 578.4 | 467.4 | 30 | 45 |
| 396.4 | 30 | 45 | ||
| Homo-YTX | 577.5 | 474.4 | 30 | 48 |
| 403.4 | 30 | 48 | ||
| 45-OH-homo YTX | 585.5 | 403.4 | 30 | 45 |
| 474.4 | 30 | 45 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Blanco, J.; Moroño, Á.; Arévalo, F.; Correa, J.; Lamas, J.P. Yessotoxins in Mollusks of the Galician Coast from 2014 to 2022: Variability, Biotransformation, and Resistance to Alkaline Hydrolysis. Toxins 2023, 15, 661. https://doi.org/10.3390/toxins15110661
Blanco J, Moroño Á, Arévalo F, Correa J, Lamas JP. Yessotoxins in Mollusks of the Galician Coast from 2014 to 2022: Variability, Biotransformation, and Resistance to Alkaline Hydrolysis. Toxins. 2023; 15(11):661. https://doi.org/10.3390/toxins15110661
Chicago/Turabian StyleBlanco, Juan, Ángeles Moroño, Fabiola Arévalo, Jorge Correa, and Juan Pablo Lamas. 2023. "Yessotoxins in Mollusks of the Galician Coast from 2014 to 2022: Variability, Biotransformation, and Resistance to Alkaline Hydrolysis" Toxins 15, no. 11: 661. https://doi.org/10.3390/toxins15110661
APA StyleBlanco, J., Moroño, Á., Arévalo, F., Correa, J., & Lamas, J. P. (2023). Yessotoxins in Mollusks of the Galician Coast from 2014 to 2022: Variability, Biotransformation, and Resistance to Alkaline Hydrolysis. Toxins, 15(11), 661. https://doi.org/10.3390/toxins15110661