Lipophilic Shellfish Poisoning Toxins in Marine Invertebrates from the Galician Coast

 , , ,
, , , 
Abstract
:1. Introduction
2. Results
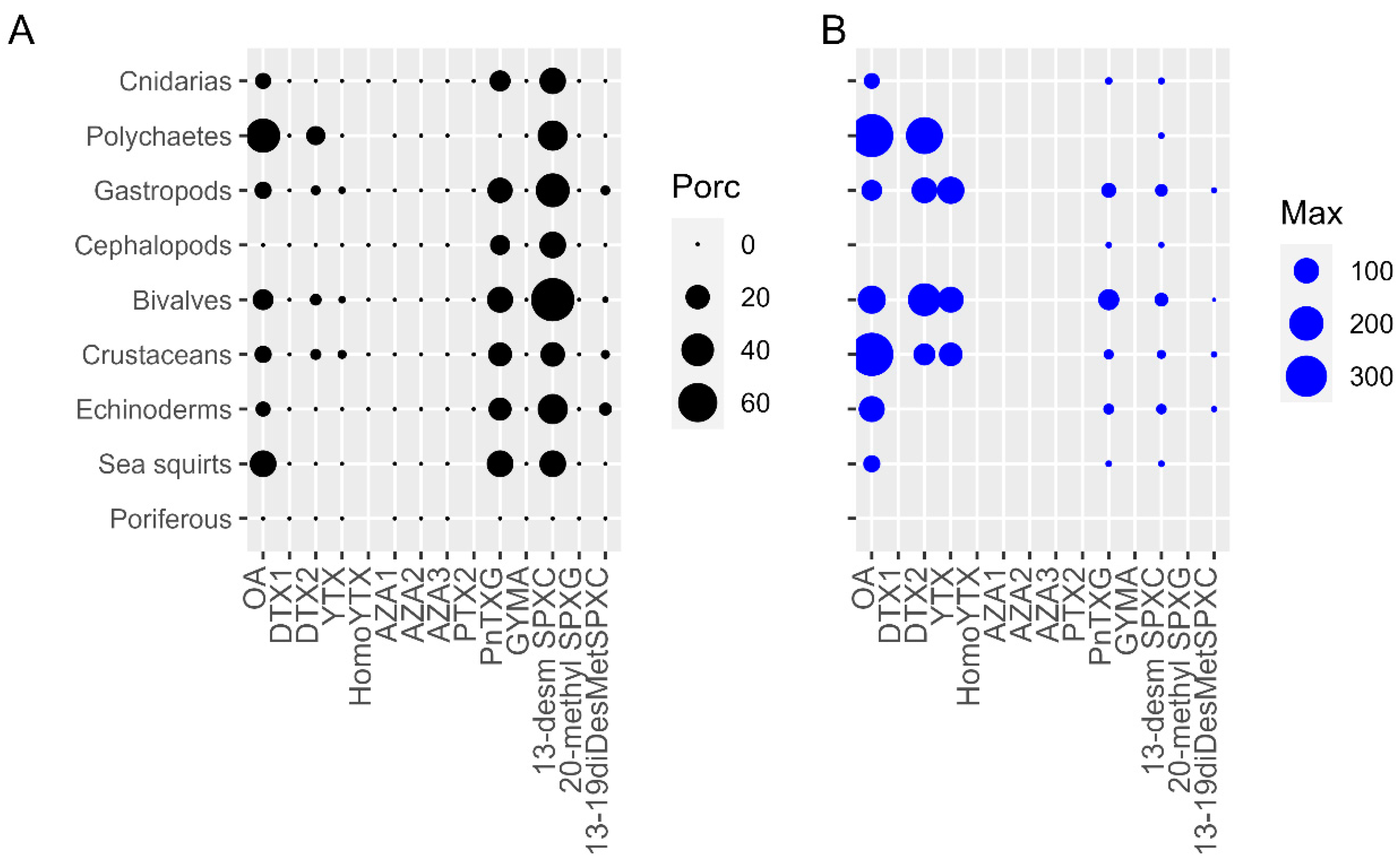
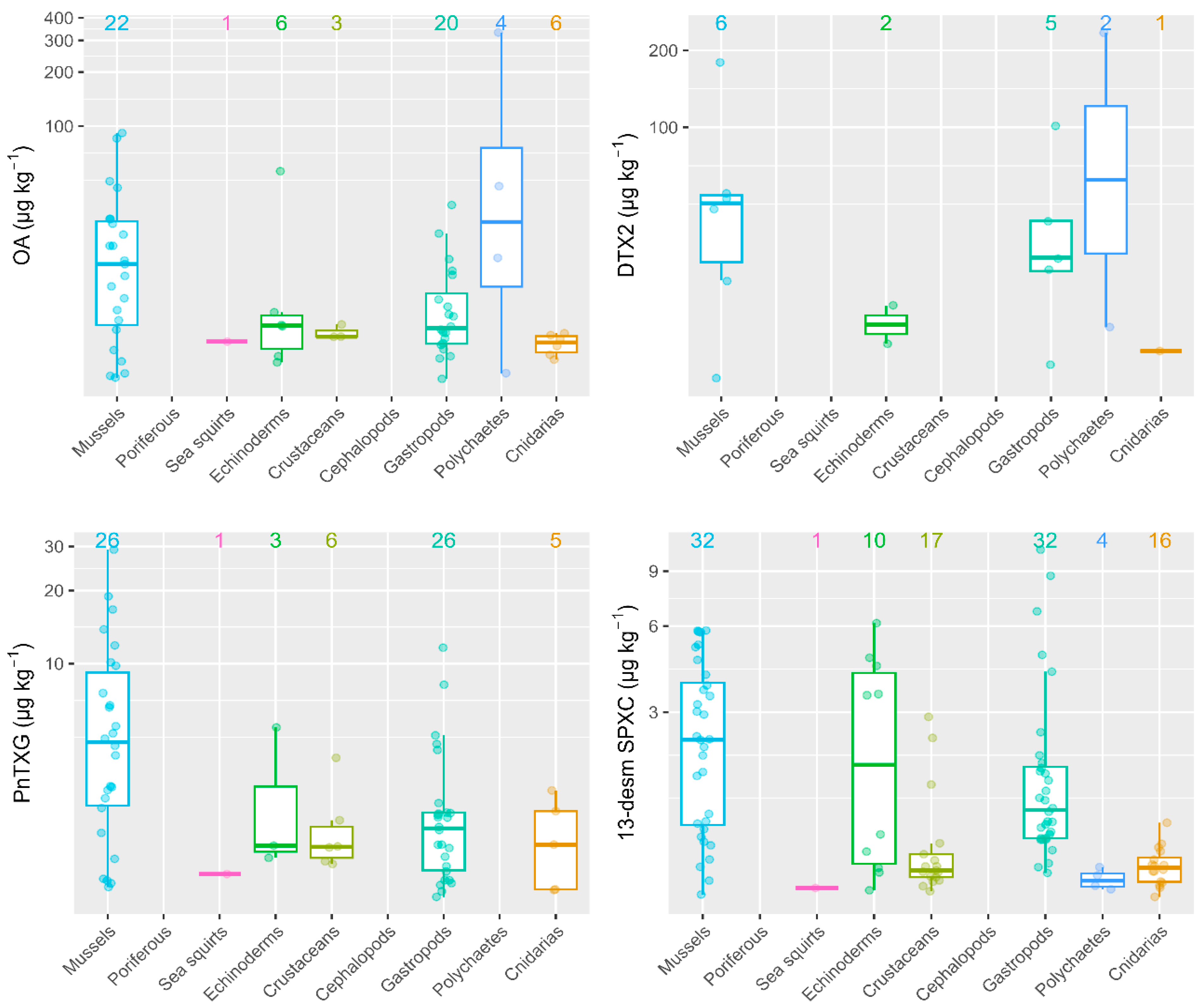
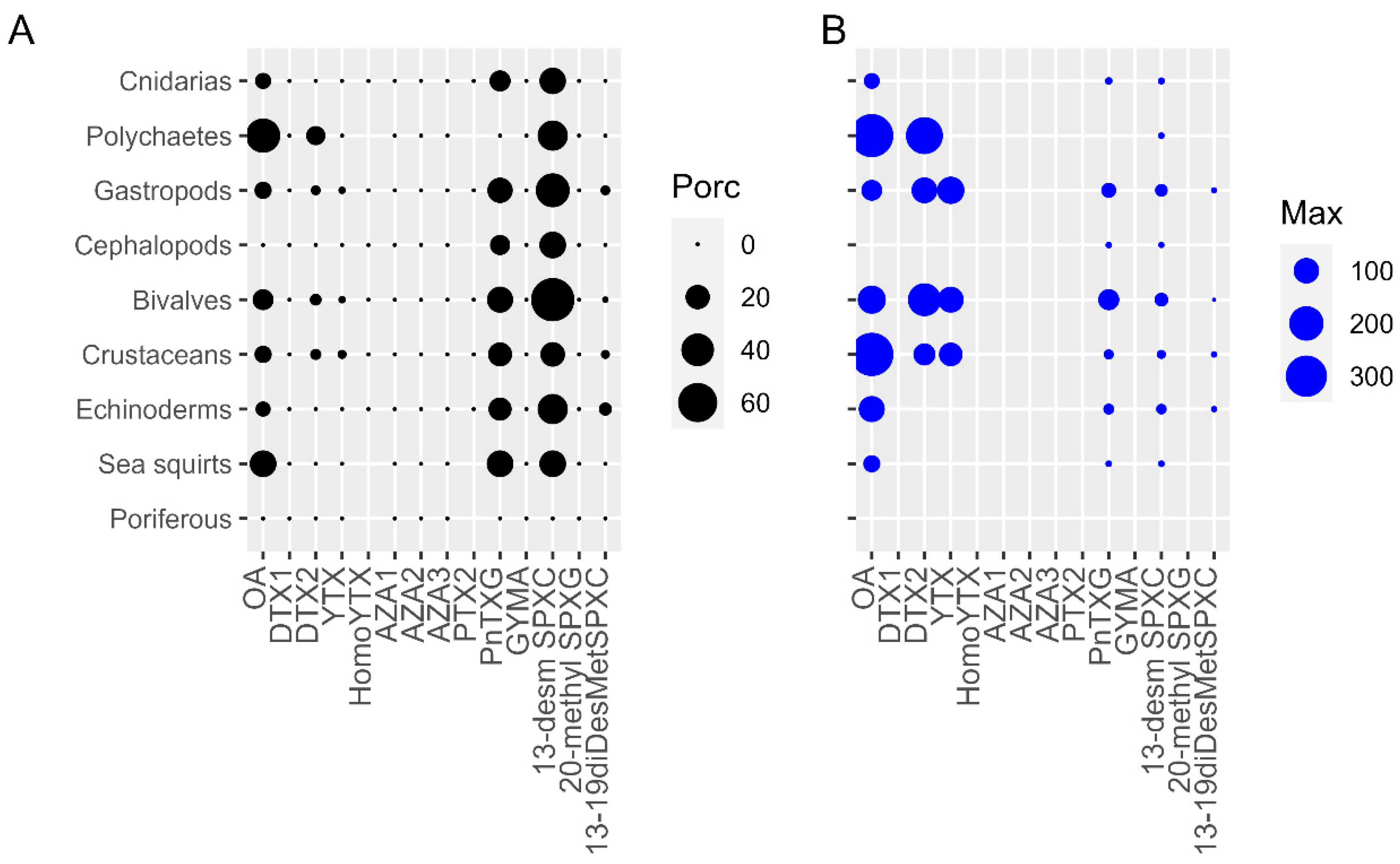
2.1. LTs in Different Marine Invertebrates Groups
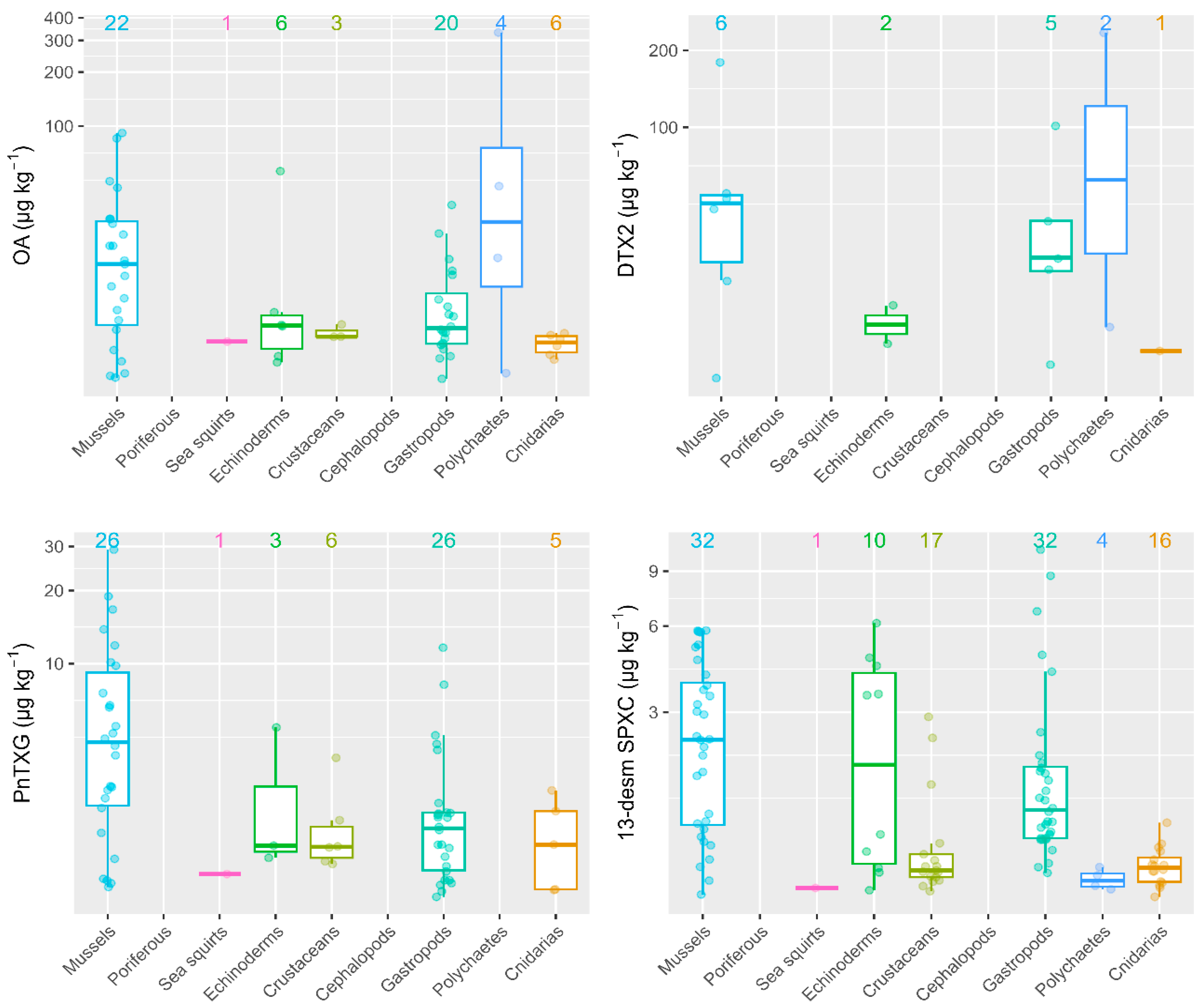
2.2. Mussels versus Different Groups of Marine Invertebrates
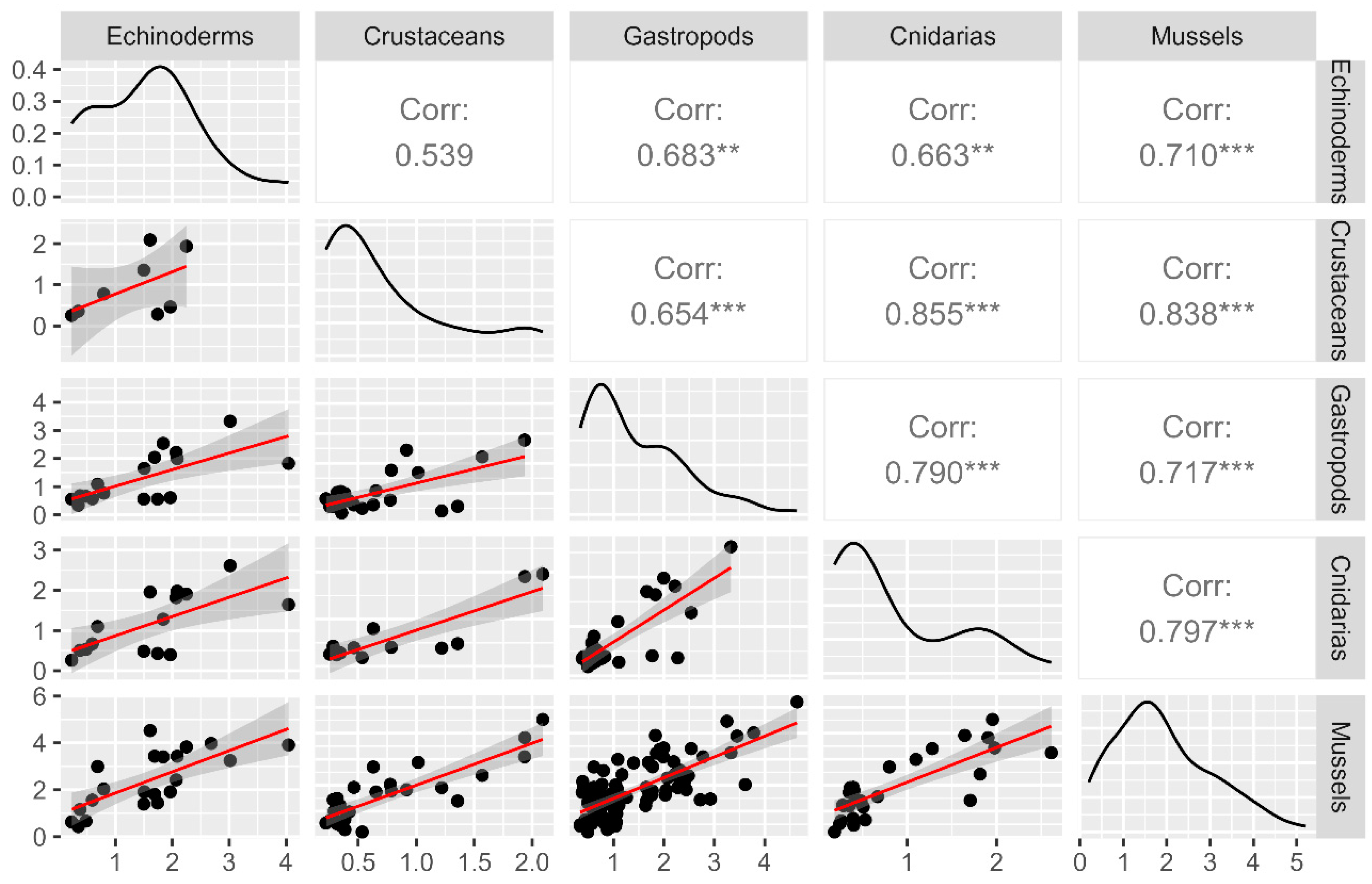
2.3. Correlation between Mussels and Different Groups of Marine Invertebrates
3. Discussion
4. Conclusions
5. Materials and Methods
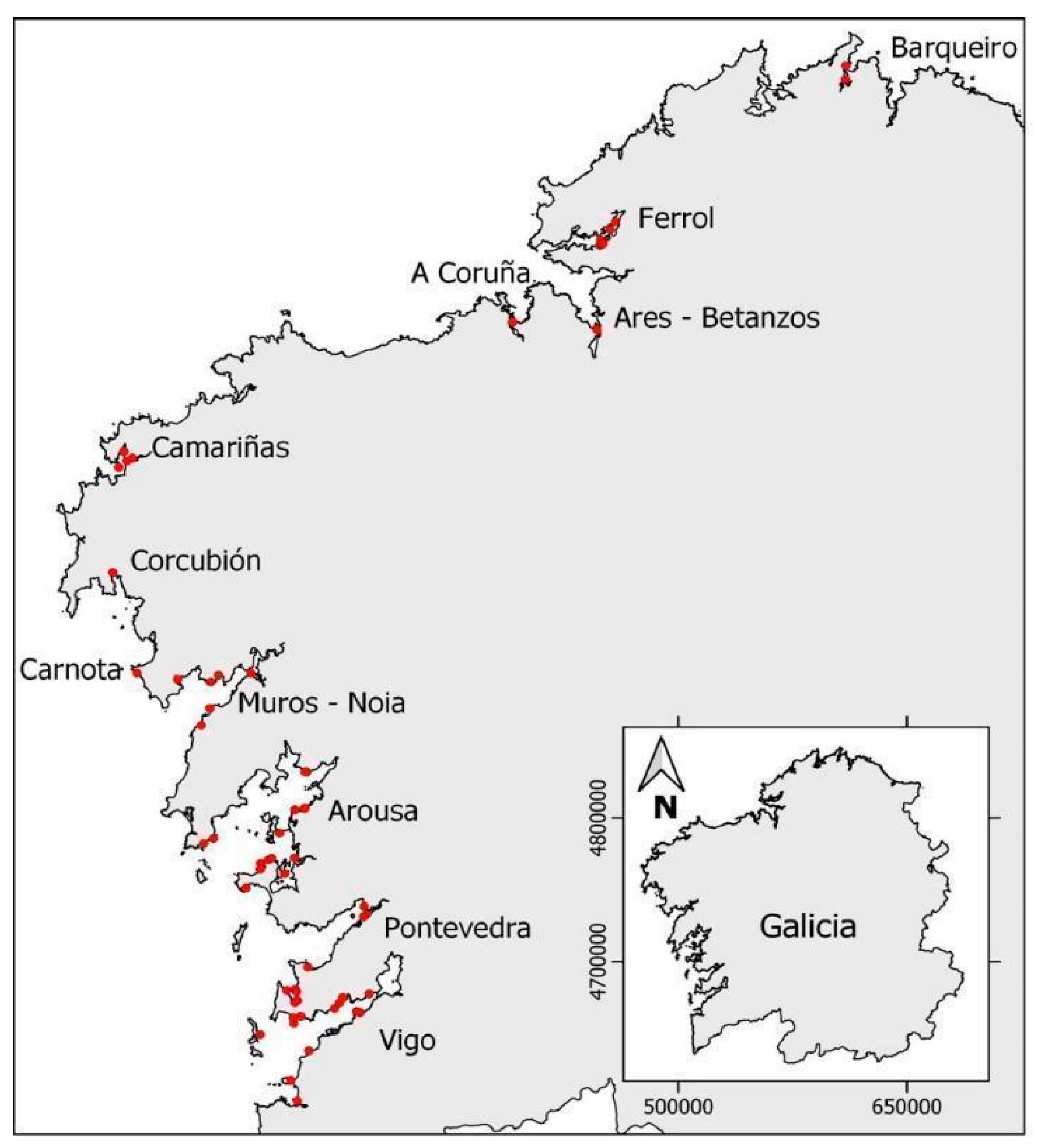
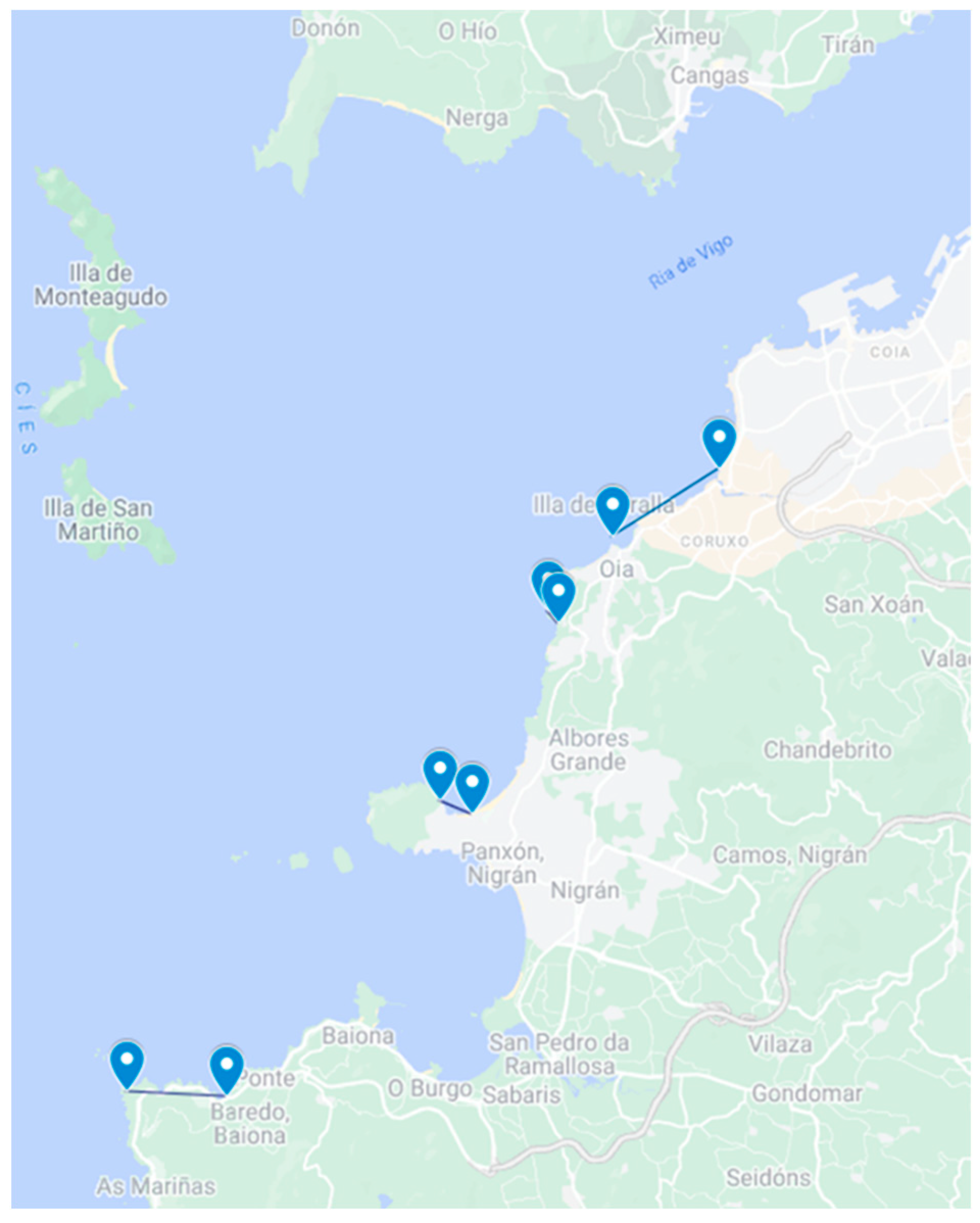
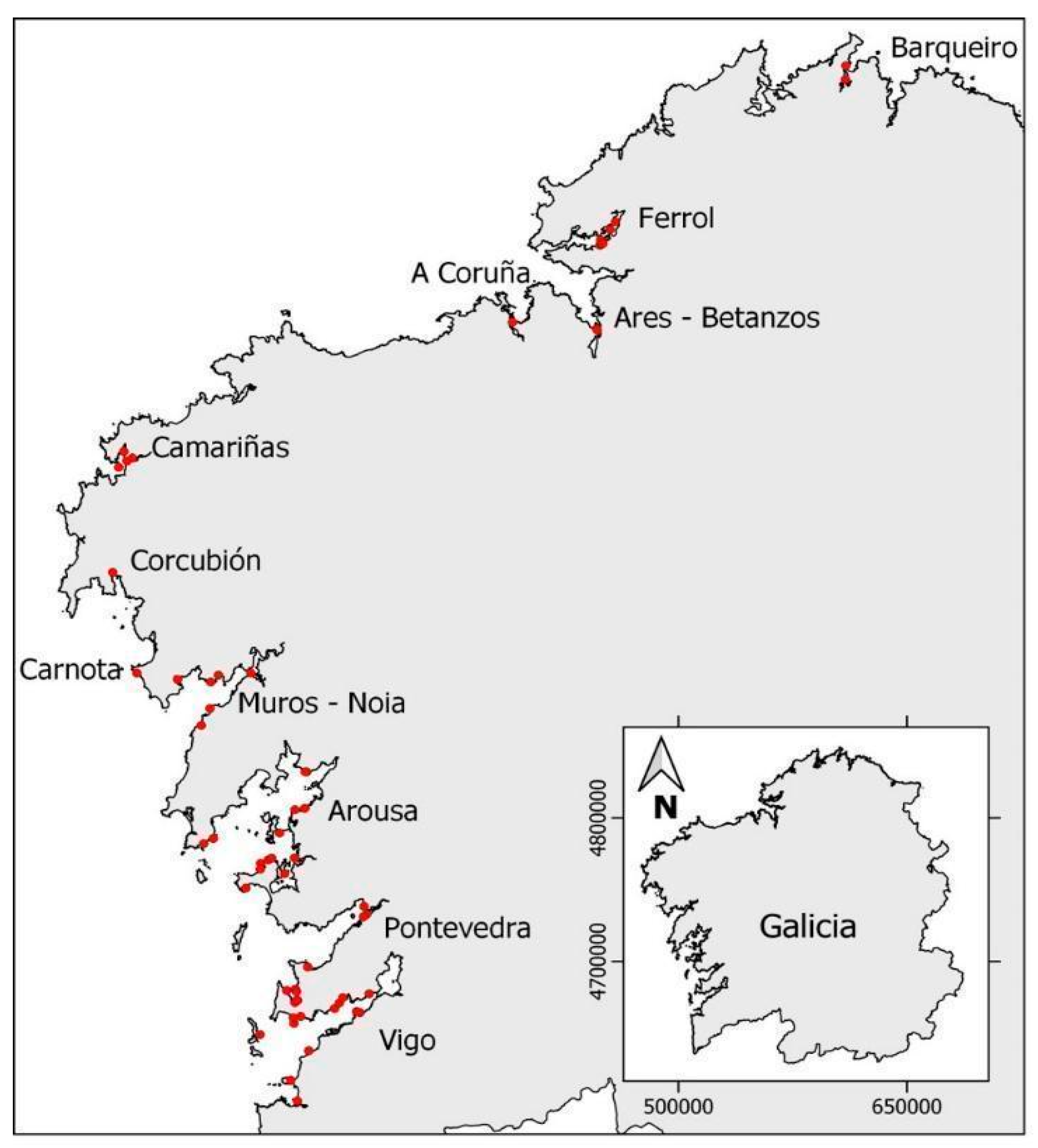
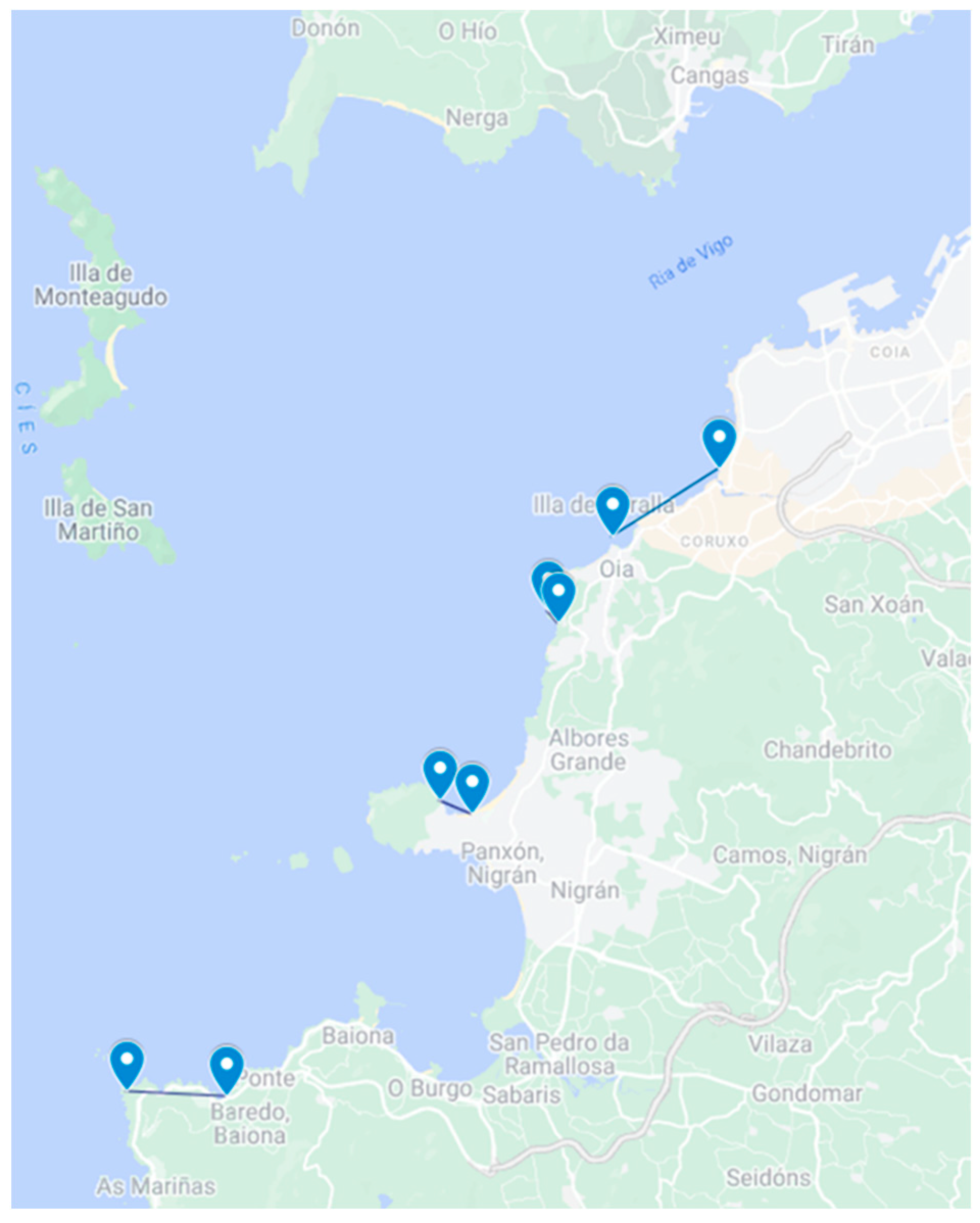
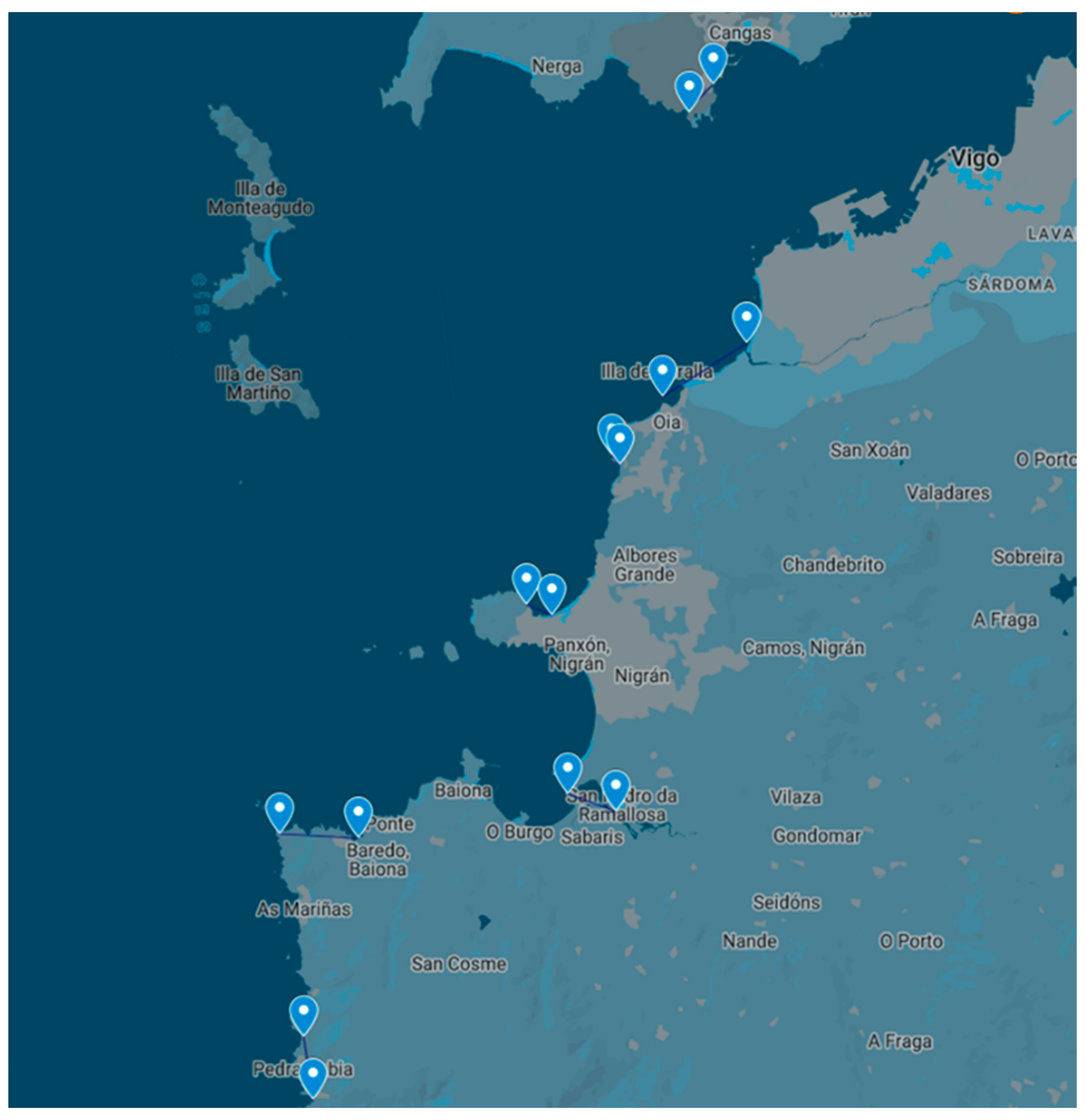
5.1. Sampling of Non-Traditional Invertebrate Vectors
5.1.1. Toxemer
5.1.2. Primrose
5.1.3. Descarsel
5.2. Chemicals and Reagents
5.3. Sample Preparation
5.4. Extraction Procedure
5.5. LC-MS/MS Analysis
5.5.1. Acidic Chromatographic Conditions
5.5.2. Alkaline Chromatographic Conditions
5.6. Statistical Analyses
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Murk, A.J.; Nicolas, J.; Smulders, F.J.M.; Bürk, C.; Gerssen, A. Marine biotoxins: Types of poisoning, underlying mechanisms of action and risk management programmes. In Food Safety Assurance and Veterinary Public Health. Chemical Hazards in Foods of Animal Origin; Smulders, F.J.M., Rietjens, I.M.C.M., Rose, M.D., Eds.; Wageningen Academic Publishers: Wageningen, The Netherlands, 2019; Volume 7, pp. 207–239. [Google Scholar]
- Martínez, A.; Garrido-Maestu, A.; Ben-Gigirey, B.; Chapela, M.J.; González, V.; Vieites, J.M.; Cabado, A.G. Marine Biotoxins. In Springer Handbook of Marine Biotechnology; Kim, S.K., Ed.; Springer Handbooks: Heidelberg, Germany, 2015; pp. 869–904. [Google Scholar]
- Arteaga-Sogamoso, E.; Rodríguez, F.; Amato, A.; Ben-Gigirey, B.; Fraga, F.; Mafra, L.L.; Fernandes, L.F.; Tibiriçá, C.E.J.A.; Chomérat, N.; Nishimura, T.; et al. Morphology and phylogeny of Prorocentrum porosum sp. nov. (Dinophyceae): A new benthic toxic dinoflagellate from the Atlantic and Pacific Oceans. Harmful Algae 2023, 121, 102356. [Google Scholar] [CrossRef] [PubMed]
- Satake, M.; Ofuji, K.; Naoki, H.; James, K.J.; Furey, A.; McMahon, T.; Silke, J.; Yasumoto, T. Azaspiracid, a new marine toxin having unique spiro ring assemblies, isolated from Irish mussels, Mytilus edulis. J. Am. Chem. Soc. 1998, 120, 9967–9968. [Google Scholar] [CrossRef]
- Twiner, M.J.; Rehmann, N.; Hess, P.; Doucette, G.J. Azaspiracid shellfish poisoning: A review on the chemistry, ecology, and toxicology with an emphasis on human health impacts. Mar. Drugs 2008, 6, 39–72. [Google Scholar] [CrossRef] [PubMed]
- Alfonso, A.; Vieytes, M.R.; Botana, L.M. Yessotoxin, a promising therapeutic tool. Mar. Drugs 2016, 14, 30. [Google Scholar] [CrossRef] [PubMed]
- Bacchiocchi, S.; Siracusa, M.; Campacci, D.; Ciriaci, M.; Dubbini, A.; Tavoloni, T.; Stramenga, A.; Gorbi, S.; Piersanti, A. Cyclic imines (CIs) in mussels from North-Central Adriatic Sea: First evidence of Gymnodimine A in Italy. Toxins 2020, 12, 370. [Google Scholar] [CrossRef] [PubMed]
- EFSA Panel on Contaminants in the Food Chain. Scientific opinion on marine biotoxins in shellfish—Cyclic imines (spirolides, gymnodimines, pinnatoxins and pteriatoxins). EFSA J. 2010, 8, 1628. [Google Scholar]
- Davidson, K.; Baker, C.; Higgins, C.; Higman, W.; Swan, S.; Veszelovszki, A.; Turner, A.D. Potential threats posed by new or emerging marine biotoxins in UK waters and examination of detection methodologies used for their control: Cyclic imines. Mar. Drugs 2015, 13, 7087–7112. [Google Scholar] [CrossRef]
- Rambla-Alegre, M.; Miles, C.O.; de la Iglesia, P.; Fernández-Tejedor, M.; Jacobs, S.; Sioen, I.; Verbeke, W.; Samdal, I.A.; Sandvik, M.; Barbosa, V.; et al. Occurrence of cyclic imines in European commercial seafood and consumers risk assessment. Environ. Res. 2018, 161, 392–398. [Google Scholar] [CrossRef]
- Kvrgić, K.; Lešić, T.; Aysal, A.I.; Džafić, N.; Pleadin, J. Cyclic imines in shellfish and ascidians in the northern Adriatic Sea. Food Addit. Contam. Part B Surveill. 2021, 14, 12–22. [Google Scholar] [CrossRef]
- Moreiras, G.; Leao, J.M.; Gago-Martinez, A. Analysis of Cyclic Imines in Mussels (Mytilus galloprovincialis) from Galicia (NW Spain) by LC-MS/MS. Int. J. Environ. Res. Public Health 2019, 17, 281. [Google Scholar] [CrossRef]
- Otero, P.; Silva, M. Emerging marine biotoxins in European waters: Potential risks and analytical challenges. Mar. Drugs 2022, 20, 199. [Google Scholar] [CrossRef] [PubMed]
- Arnich, N.; Abadie, E.; Delcourt, N.; Fessard, V.; Fremy, J.M.; Hort, V.; Lagrange, E.; Maignien, T.; Molgo, J.; Peyrat, M.B.; et al. Health risk assessment related to pinnatoxins in French shellfish. Toxicon 2020, 180, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Nézan, E.; Chomérat, N. Vulcanodinum rugosum gen. et sp. nov. (Dinophyceae), un nouveau dinoflagellé marin de la côte mediterraneenne française. Cryptogam. Algol. 2011, 32, 3–18. [Google Scholar] [CrossRef]
- Rundberget, T.; Aasen, J.A.B.; Selwood, A.I.; Miles, C.O. Pinnatoxins and spirolides in Norwegian blue mussels and seawater. Toxicon 2011, 58, 700–711. [Google Scholar] [CrossRef] [PubMed]
- Lamas, J.P.; Arévalo, F.; Moroño, A.; Correa, J.; Muñíz, S.; Blanco, J. Detection and spatio-temporal distribution of pinnatoxins in shellfish from the Atlantic and Cantabrian coasts of Spain. Toxins 2019, 11, 340. [Google Scholar] [CrossRef] [PubMed]
- Cembella, A.D.; Lewis, N.I.; Quilliam, M.A. The marine dinoflagellate Alexandrium ostenfeldii (Dinophyceae) as the causative organism of spirolide shellfish toxins. Phycologia 2000, 39, 67–74. [Google Scholar] [CrossRef]
- Touzet, N.; Franco, J.M.; Raine, R. Morphogenetic diversity and biotoxin composition of Alexandrium (Dinophyceae) in Irish coastal waters. Harmful Algae 2008, 7, 782–797. [Google Scholar] [CrossRef]
- Blanco, J.; Arévalo, F.; Moroño, A.; Correa, J.; Rossignoli, A.E.; Lamas, J.P. Spirolides in bivalve mollusk of the Galician (NW Spain) coast: Interspecific, spatial, temporal variation and presence of an isomer of 13-desmethyl spirolide C. Toxins 2023, 15, 13. [Google Scholar] [CrossRef]
- Hu, T.; Curtis, J.M.; Oshima, Y.; Quilliam, M.A.; Walter, J.A.; Watson-Wright, W.M.; Wright, J.L.C. Spirolides B and D, two novel macrocycles isolated from the digestive glands of shellfish. J. Chem. Soc. Chem. Commun. 1995, 20, 2159–2161. [Google Scholar] [CrossRef]
- Villar-González, A.; Rodríguez-Velasco, M.L.; Ben-Gigirey, B.; Botana, L.M. First evidence of spirolides in Spanish shellfish. Toxicon 2006, 48, 1068–1074. [Google Scholar] [CrossRef]
- García-Altares, M.; Casanova, A.; Bane, V.; Diogene, J.; Furey, A.; de la Iglesia, P. Confirmation of pinnatoxins and spirolides in shellfish and passive samplers from Catalonia (Spain) by liquid chromatography coupled with triple quadrupole and high-resolution hybrid tandem mass spectrometry. Mar. Drugs 2014, 12, 3706–3732. [Google Scholar] [CrossRef] [PubMed]
- Zurhelle, C.; Nieva, J.; Tillmann, U.; Harder, T.; Krock, B.; Tebben, J. Identification of novel gymnodimines and spirolides from the marine dinoflagellate Alexandrium ostenfeldii. Mar. Drugs 2018, 16, 446. [Google Scholar] [CrossRef] [PubMed]
- Seki, T.; Satake, M.; Mackenzie, L.; Kaspar, H.; Yasumoto, T. Gymnodimine, a new marine toxin of unprecedented structure isolated from New Zealand oysters and the dinoflagellate, Gymnodinium sp. Tetrahedron Lett. 1995, 36, 7093–7096. [Google Scholar] [CrossRef]
- Stewart, M.; Blunt, J.; Munro, M.; Robinson, W. The absolute stereochemistry of the New Zealand shellfish toxin gymnodimine. Tetrahedron Lett. 1997, 38, 4889–4890. [Google Scholar] [CrossRef]
- Bire, R.; Krys, S.; Fremy, J.M.; Dragacci, S.; Stirling, D.; Kharrat, R. First evidence on occurrence of gymnodimine in clams from Tunisia. J. Nat. Toxins 2002, 11, 269–275. [Google Scholar]
- Kharrat, R.; Servent, D.; Girard, E.; Ouanounou, G.; Amar, M.; Marrouchi, R.; Benoit, E.; Molgo, J. The marine phycotoxin gymnodimine targets muscular and neuronal nicotinic acetylcholine receptor subtypes with high affinity. J. Neurochem. 2008, 107, 952–963. [Google Scholar] [CrossRef] [PubMed]
- Krock, B.; Pitcher, G.C.; Ntuli, J.; Cembella, A.D. Confirmed identification of gymnodimine in oysters from the west coast of South Africa by liquid chromatography–tandem mass spectrometry. Afr. J. Mar. Sci. 2009, 31, 113–118. [Google Scholar] [CrossRef]
- Ying, Y.; Che, Y.; Wright, E.J.; McCarron, P.; Hess, P.; Li, A. Fatty acid ester metabolites of gymnodimine in shellfish collected from China and in mussels (Mytilus galloprovincialis) exposed to Karenia selliformis. Harmful Algae 2020, 92, 101774. [Google Scholar] [CrossRef]
- Araoz, R.; Barnes, P.; Sechet, V.; Delepierre, M.; Zinn-Justin, S.; Molgo, J.; Zakarian, A.; Hess, P.; Servent, D. Cyclic imine toxins survey in coastal european shellfish samples: Bioaccumulation and mode of action of 28-O-palmitoyl ester of pinnatoxin-G. First report of portimine-A bioaccumulation. Harmful Algae 2020, 98, 101887. [Google Scholar] [CrossRef]
- Nincevic Gladan, Z.; Ujevic, I.; Milandri, A.; Marasovic, I.; Ceredi, A.; Pigozzi, S.; Arapov, J. Lipophilic toxin profile in Mytilus galloprovincialis during episodes of diarrhetic shellfish poisoning (DSP) in the N.E. Adriatic Sea in 2006. Molecules 2011, 16, 888–899. [Google Scholar] [CrossRef]
- Lamas, J.P.; Arevalo, F.; Morono, A.; Correa, J.; Rossignoli, A.E.; Blanco, J. Gymnodimine A in mollusks from the north Atlantic Coast of Spain: Prevalence, concentration, and relationship with spirolides. Environ. Pollut. 2021, 279, 116919. [Google Scholar] [CrossRef]
- Otero, P.; Miguens, N.; Rodríguez, I.; Botana, L.M. LC-MS/MS analysis of the emerging toxin pinnatoxin-G and high levels of esterified OA group toxins in Galician commercial mussels. Toxins 2019, 11, 394. [Google Scholar] [CrossRef] [PubMed]
- Rossignoli, A.E.; Mariño, C.; Martín, H.; Blanco, J. First report of two gymnodimines and two tetrodotoxin analogues in invertebrates from the North Atlantic coast of Spain. Mar. Drugs 2023, 21, 232. [Google Scholar] [CrossRef] [PubMed]
- Jester, R.; Rhodes, L.; Beuzenberg, V. Uptake of paralytic shellfish poisoning and spirolide toxins by paddle crabs (Ovalipes catharus) via a bivalve vector. Harmful Algae 2009, 8, 369–376. [Google Scholar] [CrossRef]
- Vale, P.; Sampayo, M.A.d.M. First confirmation of human diarrhoeic poisonings by okadaic acid esters after ingestion of razor clams (Solen marginatus) and green crabs (Carcinus maenas) in Aveiro Lagoon, Portugal and detection of okadaic acid esters in phytoplankton. Toxicon 2002, 40, 989–996. [Google Scholar] [CrossRef] [PubMed]
- Torgersen, T.; Aasen, J.; Aune, T. Diarrhetic shellfish poisoning by okadaic acid esters from Brown crabs (Cancer pagurus) in Norway. Toxicon 2005, 46, 572–578. [Google Scholar] [CrossRef] [PubMed]
- Torgersen, T.; Bremnes, N.B.; Rundberget, T.; Aune, T. Structural confirmation and occurrence of azaspiracids in Scandinavian brown crabs (Cancer pagurus). Toxicon 2008, 51, 93–101. [Google Scholar] [CrossRef]
- Reizopoulou, S.; Strogyloudi, E.; Giannakourou, A.; Pagou, K.; Hatzianestis, I.; Pyrgaki, C.; Granéli, E. Okadaic acid accumulation in macrofilter feeders subjected to natural blooms of Dinophysis acuminata. Harmful Algae 2008, 7, 228–234. [Google Scholar] [CrossRef]
- Silva, M.; Rodríguez, I.; Barreiro, A.; Kaufmann, M.; Neto, A.I.; Hassouani, M.; Sabour, B.; Alfonso, A.; Botana, L.M.; Vasconcelos, V. New invertebrate vectors of okadaic acid from the North Atlantic waters-Portugal (Azores and Madeira) and Morocco. Toxins 2015, 7, 5337–5347. [Google Scholar] [CrossRef]
- García, C.; Pérez, F.; Contreras, C.; Figueroa, D.; Barriga, A.; López-Rivera, A.; Araneda, O.F.; Contreras, H.R. Saxitoxins and okadaic acid group: Accumulation and distribution in invertebrate marine vectors from Southern Chile. Food Addit. Contam. Part A Chem. Anal. Control. Expo. Risk Assess. 2015, 32, 984–1002. [Google Scholar] [CrossRef]
- Alves, T.P.; Mafra, L.L. Diel variations in cell abundance and trophic transfer of diarrheic toxins during a massive Dinophysis bloom in Southern Brazil. Toxins 2018, 10, 232. [Google Scholar] [CrossRef] [PubMed]
- Pires, E.; de Melo Tarouco, F.; Alves, T.P.; da Rosa, C.E.; da Cunha Lana, P.; Mafra, L.L. Antioxidant responses and okadaic acid accumulation in Laeonereis acuta (Annelida) exposed to the harmful dinoflagellate Prorocentrum cf. lima. Toxicon 2021, 203, 104–114. [Google Scholar] [CrossRef] [PubMed]
- Stirling, D. Survey of historical New Zealand shellfish samples for accumulation of gymnodimine. N. Z. J. Mar. Freshw. Res. 2001, 35, 851–857. [Google Scholar] [CrossRef]
- El Rahman Hassoun, A.; Ujević, I.; Mahfouz, C.; Fakhri, M.; Roje-Busatto, R.; Jemaa, S.; Nazlić, N. Occurrence of domoic acid and cyclic imines in marine biota from Lebanon-Eastern Mediterranean Sea. Sci. Total Environ. 2021, 755, 142542. [Google Scholar] [CrossRef] [PubMed]
- Silva, M.; Barreiro, A.; Rodriguez, P.; Otero, P.; Azevedo, J.; Alfonso, A.; Botana, L.M.; Vasconcelos, V. New invertebrate vectors for PST, spirolides and okadaic acid in the North Atlantic. Mar. Drugs 2013, 11, 1936–1960. [Google Scholar] [CrossRef] [PubMed]
- Farabegoli, F.; Blanco, L.; Rodríguez, L.P.; Vieites, J.M.; Cabado, A.G. Phycotoxins in marine shellfish: Origin, occurrence and effects on humans. Mar. Drugs 2018, 16, 188. [Google Scholar] [CrossRef]
- Ruiz-Villarreal, M.; Sourisseau, M.; Anderson, P.; Cusack, C.; Neira, P.; Silke, J.; Rodriguez, F.; Ben-Gigirey, B.; Whyte, C.; Giraudeau-Potel, S.; et al. Novel methodologies for providing in situ data to HAB early warning systems in the European Atlantic Area: The PRIMROSE experience. Front. Mar. Sci. 2022, 9, 791329. [Google Scholar] [CrossRef]
- Amzil, Z.; Derrien, A.; Terre Terrillon, A.; Savar, V.; Bertin, T.; Peyrat, M.; Duval, A.; Lhaute, K.; Arnich, N.; Hort, V.; et al. Five years monitoring the emergence of unregulated toxins in shellfish in France (EMERGTOX 2018–2022). Mar. Drugs 2023, 21, 435. [Google Scholar] [CrossRef]
- Pires, E.; Cunha Lana, P.; Mafra, L.L. Phycotoxins and marine annelids—A global review. Harmful Algae 2023, 122, 102373. [Google Scholar] [CrossRef]
- Costa, P.R.; Costa, S.T.; Braga, A.C.; Rodrigues, S.M.; Vale, P. Relevance and challenges in monitoring marine biotoxins in non-bivalve vectors. Food Control 2017, 76, 24–33. [Google Scholar] [CrossRef]
- Amzil, Z.; Sibat, M.; Royer, F.; Masson, N.; Abadie, E. Report on the First Detection of Pectenotoxin-2, Spirolide-a and Their Derivatives in French Shellfish. Mar. Drugs 2007, 5, 168–179. [Google Scholar] [CrossRef] [PubMed]
- Aasen, J.; Mackinnon, S.L.; Leblanc, P.; Walter, J.A.; Hovgaard, P.; Aune, T.; Quilliam, M.A. Detection and identification of spirolides in Norwegian shellfish and plankton. Chem. Res. Toxicol. 2005, 18, 509–515. [Google Scholar] [CrossRef] [PubMed]
- Hess, P.; Abadie, E.; Herve, F.; Berteaux, T.; Sechet, V.; Araoz, R.; Molgo, J.; Zakarian, A.; Sibat, M.; Rundberget, T.; et al. Pinnatoxin G is responsible for atypical toxicity in mussels (Mytilus galloprovincialis) and clams (Venerupis decussata) from Ingril, a French Mediterranean lagoon. Toxicon 2013, 75, 16–26. [Google Scholar] [CrossRef] [PubMed]
- Louzao, M.C.; Vilariño, N.; Vale, C.; Costas, C.; Cao, A.; Raposo-García, S.; Vieytes, M.R.; Botana, L.M. Current trends and new challenges in marine phycotoxins. Mar. Drugs 2022, 20, 198. [Google Scholar] [CrossRef] [PubMed]
- EURLMB. EU-Harmonised Standard Operating Procedure for Determination of Lipophilic Marine Biotoxins in Molluscs by LC-MS/MS. Version 5. Available online: https://www.aesan.gob.es/CRLMB/docs/docs/metodos_analiticos_de_desarrollo/EU-Harmonised-SOP-LIPO-LCMSMS_Version5.pdf (accessed on 28 July 2023).
- Rossignoli, A.E.; Mariño, C.; Martín, H.; Blanco, J. Development of a fast liquid chromatography coupled to mass spectrometry method (LC-MS/MS) to determine fourteen lipophilic shellfish toxins based on fused–core technology: In-house validation. Mar. Drugs 2021, 19, 603. [Google Scholar] [CrossRef]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2019. [Google Scholar]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| (a) | |||
| Flow: [0.25–0.40] mL/min | |||
| Injection Volume: 5 µL, Injector Temp: 4 °C Column Temp: 30 °C | |||
| Time (min) | Flow | %A | %B |
| 0.10 | 0.25 | 90 | 10 |
| 6.50 | 0.25 | 20 | 80 |
| 8.50 | 0.25 | 0 | 100 |
| 9.61 | 0.25 | 0 | 100 |
| 10.00 | 0.40 | 0 | 100 |
| 11.50 | 0.40 | 0 | 100 |
| 11.60 | 0.25 | 90 | 10 |
| 15.00 | 0.25 | 90 | 10 |
| (b) | |||
| Parameters | Positive Mode | Negative Mode | |
| Curtain Gas (CUR) | 25 | 25 | |
| Collision Gas (CAD) | High | High | |
| Voltage (IS) V | 4500 | −4500 | |
| Temperature (TEMP) °C | 500 | 500 | |
| Gas 1 (GS1) psi | 50 | 50 | |
| Gas 2 (GS2) psi | 50 | 50 | |
| (a) | |||
| Flow: 0.40 mL/min | |||
| Injection Volume: 1 µL, Injector Temp: 4 °C Column Temp: 40 °C | |||
| Time (min) | Flow | %A | %B |
| 0.50 | 0.40 | 78 | 22 |
| 3.85 | 0.40 | 5 | 95 |
| 6.25 | 0.40 | 5 | 95 |
| 6.75 | 0.40 | 78 | 22 |
| 8.75 | 0.40 | 78 | 22 |
| (b) | |||
| Parameters | Positive Mode | Negative Mode | |
| Curtain Gas (CUR) | 30 | 30 | |
| Collision Gas (CAD) | Medium | Medium | |
| Voltage (IS) V | 5000 | −4500 | |
| Temperature (TEMP) °C | 600 | 600 | |
| Gas 1 (GS1) psi | 75 | 75 | |
| Gas 2 (GS2) psi | 75 | 75 | |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Rossignoli, A.E.; Ben-Gigirey, B.; Cid, M.; Mariño, C.; Martín, H.; Garrido, S.; Rodríguez, F.; Blanco, J. Lipophilic Shellfish Poisoning Toxins in Marine Invertebrates from the Galician Coast. Toxins 2023, 15, 631. https://doi.org/10.3390/toxins15110631
Rossignoli AE, Ben-Gigirey B, Cid M, Mariño C, Martín H, Garrido S, Rodríguez F, Blanco J. Lipophilic Shellfish Poisoning Toxins in Marine Invertebrates from the Galician Coast. Toxins. 2023; 15(11):631. https://doi.org/10.3390/toxins15110631
Chicago/Turabian StyleRossignoli, Araceli E., Begoña Ben-Gigirey, Mónica Cid, Carmen Mariño, Helena Martín, Soledad Garrido, Francisco Rodríguez, and Juan Blanco. 2023. "Lipophilic Shellfish Poisoning Toxins in Marine Invertebrates from the Galician Coast" Toxins 15, no. 11: 631. https://doi.org/10.3390/toxins15110631
APA StyleRossignoli, A. E., Ben-Gigirey, B., Cid, M., Mariño, C., Martín, H., Garrido, S., Rodríguez, F., & Blanco, J. (2023). Lipophilic Shellfish Poisoning Toxins in Marine Invertebrates from the Galician Coast. Toxins, 15(11), 631. https://doi.org/10.3390/toxins15110631